
Abstract
Capsule Feeding habits of the Black-billed Magpie are of interest for researchers, conservationists and hunters since magpies are considered as predators of eggs and chicks of both songbirds and gamebirds.
Aims To characterize the feeding habits of magpies during their breeding season in agricultural environments of central Spain, and to assess the occurrence and incidence of birds and eggs in the magpie's diet.
Methods Diet was determined by the analysis of gizzard contents from 118 culled magpies. The diet was described as the frequency of occurrence (FO) and the percentage of volume (VOL) of a certain food item and for each gizzard.
Results Magpies presented a generalist diet, which included a wide range of foods. Arthropods and cereal seeds were the most frequently consumed food groups (FO > 60%). Eggs and birds were consumed only occasionally (FO < 6% and 17%, respectively; percentage of volume, VOL < 4%).
Conclusion Our findings suggest that other birds and their eggs do not represent an important food for magpies in Mediterranean agricultural environments under the conditions found during this study. Nevertheless, more complex studies in different scenarios (i.e. different population sizes of magpies and prey), and over longer temporal scales, are still necessary to clarify this controversial issue.
Feeding habits are an important and widely studied aspect of animal ecology and a fundamental component for understanding the biology and ecology of species. Some species are frequently perceived as harmful for human interests because of their feeding habits. These include, for instance, some predators that consume game species or livestock (Woodroffe et al. Citation2005). From this point of view, the information provided by studies on predator feeding habits may be relevant to guide appropriate policy and management decisions that facilitate human–wildlife coexistence (López-Bao et al. Citation2013).
The feeding habits of the Black-billed Magpie Pica pica (hereafter the Magpie) gives rise to controversial interpretations between researchers, conservationists and hunters. In Europe, Magpies are considered as a harmful bird species by some conservationists and hunters because of their predation on eggs and chicks of songbirds and gamebirds (Birkhead Citation1991, Herranz Citation2000). As a consequence, control of Magpie populations is widespread in Europe (Hadjisterkotis Citation2003, Chiron & Julliard Citation2013, Díaz-Ruiz & Ferreras Citation2013). In Spain, Magpie control is mostly performed by hunters and game managers, who consider these birds as highly efficient predators of nests of Red-legged Partridges Alectoris rufa (Delibes-Mateos et al. Citation2013, Díaz-Ruiz & Ferreras Citation2013), a small game species of socio-economic relevance (Díaz-Fernández et al. Citation2012).
The Magpie diet has been the object of several studies focusing on different issues; e.g. seasonal differences, food selection, diet of nestlings or differences between rural and urban magpies (Birkhead Citation1991, Soler & Soler Citation1991, Martínez et al. Citation1992, Ponz et al. Citation1999, Kryštofková et al. Citation2011). According to these studies, Magpies are generalist predators that feed on a broad spectrum of food types, included both vegetal and animal resources, which ranged from seeds and small invertebrates to larger vertebrate carcasses and human discarded waste. In general, most studies agree that eggs form only a small proportion of the Magpie diet (Birkhead Citation1991, Martínez et al. Citation1992), although on some occasions Magpies are one of the main predators of artificial and natural nests (Groom Citation1993, Herranz Citation2000, Miller & Hobbs Citation2000, Roos & Pärt Citation2004). Nevertheless, the impact of Magpies on bird populations remains unclear, due to contrasting results (Gooch et al. Citation1991, Thomson et al. Citation1998, Stoate & Szczur Citation2001, Chiron & Julliard Citation2007, Newson et al. Citation2010), particularly in the Iberian Peninsula, where the number of studies on this issue is low.
In the present study, our main goal was to characterize the feeding habits of Magpies during their breeding season in agricultural areas of central Iberia, paying particular attention to the occurrence and importance of birds and eggs in the diet.
MATERIAL AND METHODS
Study Area
Magpie feeding habits were studied in two hunting estates located in central Spain (Area 1: 960 ha, 39°4.5ʹN, 3°54ʹW; Area 2: 547 ha, 39°33ʹN, 3°12ʹW), during spring 2006. Both study areas were within the Mediterranean bioclimatic region (Rivas-Martínez et al. Citation2004), and were similar in habitat composition: an agricultural dominated landscape with some interspersed patches of natural vegetation, mainly Mediterranean bushes, some trees in riparian areas and hedgerows. The main crops were cereals (∼50% and 70% of total surface, respectively in Area 1 and Area 2) and, to a lesser extent, vineyards and olive groves. Hunting was an important activity in both estates, and the main game species were Iberian Hare Lepus granatensis, European Rabbit Oryctolagus cuniculus and Red-legged Partridge. Partridge density was low in both estates (less than 0.36 partridges/ha, authors unpubl. data) and within the range of other agricultural regions of the Iberian Peninsula (Borralho et al. Citation1996, Duarte & Vargas Citation2001). Both hunting estates harbour an important community of small breeding birds, including species of families such as larks Alaudidae and finches Fringillidae (Martí & Del Moral Citation2003). Magpie density in both study areas (Area 1: 0.23 Magpies/ha, Area 2: 0.39 Magpies/ha, before the breeding season; see Díaz-Ruiz et al. Citation2010) was above the average values reported in other European regions (Birkhead Citation1991).
Sample collection
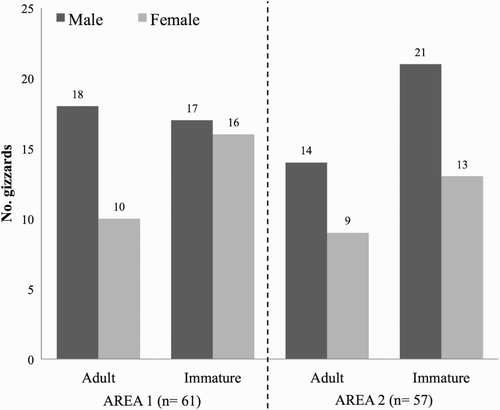
Magpies were captured during an experimental evaluation of cage-traps as live capture methods for Magpie population management (for more details see Díaz-Ruiz et al. Citation2010). Captures took place during the Magpie breeding season of 2006. Magpies were captured earlier in Area 1 (May) than in Area 2 (late May–early June). Birds were humanely euthanized using standard procedures and following current guidelines on animal welfare (Close et al. Citation1997). Age was determined from the shape and appearance of the first outermost primaries; this method allows differentiation between first-year (hereafter immature) and older magpies (hereafter adult) (Erpino Citation1968, Birkhead Citation1991). Sex was determined for each individual by the assessment of gonadal development during laboratory autopsies. Gizzard contents were extracted and stored in 70% alcohol in labelled plastic tubes for subsequent analyses. A total of 118 gizzards were collected, achieving a similar sample size for each study area (61 from Area 1 and 57 from Area 2), age (51 adult and 67 immature) and sex (48 females and 70 males) ().
Figure 1. Summary of the sample sizes for both study sites, according to the sex and age of sampled Magpies.
Diet analysis
Magpie diet was determined through the analysis of gizzard contents, a frequent method used in diet studies of several bird species (Jiguet Citation2002, Kopij Citation2005, Bur et al. Citation2008). Gizzard contents were analysed following the methods described in other corvid diet studies (Soler et al. Citation1990, Soler & Soler Citation1991, Herranz Citation2000). Food items were identified to the lowest possible taxonomic level using published literature (Day Citation1966, Barrientos Citation1988, Devesa Citation1991, Teerink Citation1991, Chinery Citation1997), as well as a dedicated reference collection of seeds, invertebrates, bird eggs and mammal hairs. The thickness of eggshells was measured with a digital calliper (precision 0.01 mm) to assign the eggs at least to the family level (Herranz Citation2000). All identified items were pooled in nine food classes: arthropod, gastropod, cereal seed, fruit, other vegetal, bird, bird egg, reptile and mammal, and two non-food items: gastrolith and plastic ().
Table 1. Detailed description of Magpie diet composition. The number of analysed gizzards containing each food group is shown. For each food group, we present the frequency of occurrence (FO) and the average % volume (VOL). Data are independently presented in terms of overall Magpie diet (Total) and for each study area (A1 and A2). Bold values are the values estimated for each main food class, which summarises other food classes. There are 9 main food classes (i.e. arthropod, gastropod, cereal seed, fruit, other vegetal, bird, bird egg, reptile and mammal), which should be highlighted in bold to differentiate from subclasses.
For the diet description we calculated two dietary indices frequently used in diet studies (Soler et al. Citation1993, Herranz Citation2000, Hadjisterkotis Citation2003, Kryštofková et al. Citation2011): the frequency of occurrence (FO), expressed as the percentage of gizzards in which a food item was found, and the percentage of volume (VOL), estimated as the percentage of total volume corresponding to a certain food item upon the total content of each gizzard. For VOL estimation, the remains from each gizzard were spread by group in a Petri dish; this enabled quantification of percentage volume by comparing the different sizes of remains for the food groups.
To detect possible biases in the interpretation of diet description, we explored potential differences in overall Magpies’ diet between study areas, sex and age classes. We pooled all food classes in four main categories to avoid groups with very low FO (<5%; e.g. fruits, reptiles and mammals). The four categories were invertebrates (arthropods and gastropods), cereal seeds, vegetal (encompassing fruits and other vegetal material, see below) and vertebrates (eggs, birds, reptiles and mammals). We used multivariate analysis of variance (manova) with the VOL of each main food category as response variables and the study area, age and sex and all interactions between them as fixed factors. We used VOL because this index considers the amount of each food class in each Magpie gizzard (Reynolds & Aebischer Citation1991). The individual gizzard was considered as the sampling unit in the statistical analyses. Prior to statistical analyses, the VOL for each food class (dependent variables) were log (x + 1) transformed to achieve normality (Zar Citation1984). Statistical analyses were performed using Statistica 10.0 software (Statsoft INC Citation2011) and the significance level was set at α = 0.05.
RESULTS
Overall, we identified 1016 food items in the gizzard contents belonging to 26 taxonomic groups (). Magpies consumed a wide range of food items among which arthropods and cereal seeds were the most frequent classes, followed by other vegetal material and birds (). Other food classes (gastropods, mainly small snails, bird eggs, fruits, mammals and reptiles) were present in much lower FO (<10%, ). Coleoptera and formicidae species represented 90% of the items consumed among the arthropoda (). We were able to identify 84% of the seeds found in the gizzards, and most of them corresponded to Hordeum sp. (64%), Avena sp. (27%) and Triticum sp. (9%) (). The ‘other vegetal' class was composed mainly by grass stalks and leaves of unidentified herbaceous plants, likely from cereal crops. We only could differentiate bird remains to the taxonomic order level by the microscopic structure of feathers (Day Citation1966). Most bird remains belonged to passeriformes, and only one of them corresponded to galliformes (). Bird egg remains always appeared highly fragmented, making the identification of the species very difficult. Nevertheless, according to the thickness of eggshells, four (<0.09 mm) were compatible with eggs of small birds (likely passeriformes), one (0.14 mm) with those of doves and one with those of partridges (0.23 mm, Herranz Citation2000). The rest of the vertebrate prey items were remains of two Wood Mice Apodemus sylvaticus, hairs from cats Felis sp., one undetermined mammal and one undetermined reptile species ().
manova results showed that diet composition only varied significantly between study areas (F4, 107 = 9.15, P < 0.001) and that there was a statistically significant effect of the sex–area interaction (F4, 107 = 3.48, P = 0.01).
DISCUSSION
Our findings show that, during the breeding season, Magpies fed on a range of different food types, with arthropods and cereal seeds being the most frequently consumed food classes. Invertebrates are the principal contribution of protein for a large number of bird species (Capinera Citation2010), including the Magpies in this study. Our results are in agreement with previous studies conducted in Spain, which indicate that, although invertebrates and seeds are consumed throughout the year, the consumption of the former increases during the breeding season, when their availability is higher and nestling demand is higher (Buitron Citation1988, Soler & Soler Citation1991, Martínez et al. Citation1992, Ponz et al. Citation1999, Herranz Citation2000).
Eggs were detected in a low proportion and volume in Magpie gizzards (<6%), which is in agreement with most previous studies (Birkhead Citation1991). A higher occurrence of eggs in magpie diet has been recorded in a previous study conducted in central Spain (FO = 13–20%, Herranz Citation2000); a large proportion of which were attributed to Red-legged Partridges (77–80%). In contrast, only one of the egg remains found in our study (17%) coincided with partridge egg thickness. This suggests that partridge eggs do not represent an important food for Magpies during the breeding season in our study areas. However, several studies conducted in the Iberian Peninsula have shown that Magpies are one of the main predators of dummy partridge nests (Herranz Citation2000, Blanco-Aguiar et al. Citation2001, Ferreras et al. Citation2010). From this perspective, we cannot reject the hypothesis that nest predation by Magpies could represent some risk for partridge breeding success. In addition, partridge nest predation by Magpies may be underestimated in diet studies, which hardly detect remains of eggshells (Chiron & Julliard Citation2007). This is probably because magpie behaviour during egg predation and ingestion varies with egg size. While smaller eggs are entirely swallowed, including the eggshell, larger ones are broken and only the egg content and small eggshell pieces are swallowed (Suvorov et al. Citation2012), decreasing the likelihood of eggshell ingestion. Also, we do not know if nest predation intensity by Magpies varies with their breeding stage in our study area. In this sense, Suvorov et al. (Citation2012) showed that Magpies predated dummy nests more frequently during incubation than during the nestling phase, probably because magpies preferentially feed invertebrates to nestlings (Martínez et al. Citation1992). From this perspective, diet studies from different breeding stages may be biased and non-comparable, since detection probability of eggshells could be higher in the early breeding stage (incubation) than during later stages (nestling provisioning).
We found a relatively high consumption of passerines (12.7% FO) in comparison to other studies performed during the breeding season (FO < 8%; Birkhead Citation1991, Herranz Citation2000, Kryštofková et al. Citation2011). It has been suggested that Magpie predation on breeding birds may be related to high bird densities (Birkhead Citation1991). However, Fernández-Juricic et al. (Citation2004) found that Magpie predation on other bird species was opportunistic and was mainly observed during the breeding season. Magpies might increase their predatory pressure on birds when invertebrates, the main animal component of their diet, are less available. Nevertheless, we lack data to test this hypothesis, which may be worth exploring in future studies. In any case, our findings agree with previous studies, which suggest that birds and eggs may be a secondary source of protein for Magpies during the breeding season (Birkhead Citation1991).
Magpie diet varied between localities but not in relation to Magpie age or sex. This result was potentially related to food availability, as suggested by the similar consumption of cereal seeds between areas, which had similar cereal crop land cover. Nevertheless, we must be cautious with this interpretation for two reasons. First, we lacked data about the availability of the other food groups and, second, Magpies can select food items independently of their availability, as reported for some invertebrate groups (Martínez et al. Citation1992, Kryštofková et al. Citation2011).
Overall, we found no evidence that Magpies pose a big threat for other birds. However, the possible sources of bias associated with our methodology, such as how we quantified the bird remains and eggs, as well as the fact that even a low rate of predation may affect a prey species when the predator is abundant, should make us cautious about this conclusion. Therefore, more complex and experimental studies over greater time and spatial scales are necessary, including localities with different densities of Magpies and potential prey species. Diet data should be complemented with the monitoring of the abundances of potential prey species and Magpies, prey breeding success and predation rates of Magpies on nests, chicks and adult birds.
Acknowledgements
We are very grateful to land owners and game managers who allowed us to work in their hunting estates, and to Ian Hartley and an anonymous referee for their helpful comments on a previous version of the manuscript. We thank people who assisted us during the fieldwork, especially S. Luna and L.E. Minguez. We acknowledge Dr. J.T. García and Dr. E. Pérez-Ramírez for autopsies and sexing of Magpies.
Ethical standards: This work was performed in compliance with current Spanish legislation, and follows the European Union's recommendations regarding animal welfare. All procedures were carried out with all legal permits required by the concerned administrations.
Funding
The study was funded by Consejería de Medio Ambiente of Junta de Comunidades de Castilla-La Mancha (Project PREG-05-23). F. Díaz-Ruiz holds a postdoctoral contract financed by the European Social Fund (ESF) and the Junta de Comunidades de Castilla-La Mancha (Operational Programme FSE 2007/2013). M. Delibes-Mateos is supported by the Talentia Postdoc Program launched by the Andalusian Knowledge Agency, co-funded by the European Union's Seventh Framework Program, Marie Skłodowska-Curie actions (COFUND – Grant Agreement n° 267226) and the Ministry of Economy, Innovation, Science and Employment of the Junta de Andalucía.
References
- Barrientos, J.A. 1988. Bases para un curso práctico de entomología. Asociación española de Entomología. Department of Animal Biology, University of Salamanca, Salamanca, Spain.
- Birkhead, T.R. 1991. The Magpies: The Ecology and Behaviour of Black-billed and Yellow-billed Magpies. T and A.D. Poyser, London.
- Blanco-Aguiar, J.A., García, J.F., Ferreras, P., Viñuela, J. & Villafuerte, R. 2001. Effect of game management on artificial nest predation in central Spain. In 25th International Union of Game Biologists (IUGB) and the 9th International Symposium Perdix, Limasol, Chipre.
- Borralho, R., Rego, F. & Vaz-Pinto, P. 1996. Is driven transect sampling suitable for estimating red-legged partridge Alectoris rufa densities? Wildlife Biol. 2: 259–268.
- Buitron, D. 1988. Female and male specialization in parental care and its consequences in black-billed magpies. Condor 90: 29–39. doi: 10.2307/1368429
- Bur, M.T., Stapanian, M.A., Bernhardt, G. & Turner, M.W. 2008. Fall diets of Red-breasted Merganser (Mergus serrator) and Walleye (Sander vitreus) in Sandusky Bay and adjacent waters of western Lake Erie. Am. Midl. Nat. 159: 147–161. doi: 10.1674/0003-0031(2008)159[147:FDORMM]2.0.CO;2
- Capinera, J.L. 2010. Insects and Wildlife: Arthropods and Their Relationships with Wild Vertebrate Animals. Wiley-Blackwell, Oxford.
- Chinery, M. 1997. Guía de campo de los insectos de España y de Europa. Ediciones Omega, S.A., Barcelona.
- Chiron, F. & Julliard, R. 2007. Responses of songbirds to magpie reduction in an urban habitat. J. Wildlife. Manage. 71: 2624–2631. doi: 10.2193/2006-105
- Chiron, F. & Julliard, R. 2013. Assessing the effects of trapping on pest bird species at the country level. Biol. Conserv. 158: 98–106. doi: 10.1016/j.biocon.2012.08.001
- Close, B., Banister, K., Baumans, U., Bernoth, E.M., Bromage, N., Bunyan, J., Erhardt, W. & Flecknell, P. 1997. Recommendations for euthanasia of experimental animals: Part 2. Laboratory Animals 31: 1–32. doi: 10.1258/002367797780600297
- Day, M.G. 1966. Identification of hair and feather remains in the gut a faeces of stoats and weasels. J. Zool. 148: 201–217. doi: 10.1111/j.1469-7998.1966.tb02948.x
- Delibes-Mateos, M., Díaz-Fernández, S., Ferreras, P., Viñuela, J. & Arroyo, B. 2013. The role of economic and social factors driving predator control in small game estates in central Spain. Ecol. Soc. 18: article no. 28. http://www.ecologyandsociety.org/vol18/iss2/art28/.
- Devesa, J.A. 1991. Las gramineas de Extremadura. University of Extremadura, Badajoz, Spain.
- Díaz-Fernández, S., Viñuela, J. & Arroyo, B. 2012. Harvest of red-legged partridge in central Spain. J. Wildlife Manage. 76: 1354–1363. doi: 10.1002/jwmg.391
- Díaz-Ruiz, F. & Ferreras, P. 2013. Conocimiento científico sobre la gestión de depredadores generalistas en España: el caso del zorro (Vulpes vulpes) y la urraca (Pica pica). Ecosistemas 22: 40–47. doi: 10.7818/ECOS.2013.22-2.07
- Díaz-Ruiz, F., García, J.T., Pérez-Rodríguez, L. & Ferreras, P. 2010. Experimental evaluation of live cage-traps for black-billed magpies Pica pica management in Spain. Eur. J. Wildl. Res. 56: 239–248. doi: 10.1007/s10344-009-0310-3
- Duarte, J. & Vargas, J.M. 2001. Survey methods for red-legged partridge (Alectoris rufa) in olive groves in Southern Spain. Game Wildlife Sci. 18: 141–156.
- Erpino, M.J. 1968. Age determination in the Black-billed Magpie. Condor 70: 91–92. doi: 10.2307/1366522
- Fernández-Juricic, E., Jokimäki, J., McDonald, J.C., Melado, F., Toledano, A., Mayo, C., Martín, B., Fresneda, I. & Martín, V. 2004. Effect of opportunistic predation on anti-predator behavioural responses in a guild ground foragers. Oecologia 140: 183–190. doi: 10.1007/s00442-004-1560-0
- Ferreras, P., Mateo-Moriones, A. & Villafuerte, R. 2010. Influencia de la depredación sobre la perdiz roja en Navarra. Final Report. Gobierno de Navarra, IREC-CSIC.
- Gooch, S., Baillie, S.R. & Birkhead, T.R. 1991. Magpie Pica pica and songbirds populations: restrospective investigation of trends in population density and breeding success. J. Appl. Ecol. 28: 1068–1086. doi: 10.2307/2404226
- Groom, D.W. 1993. Magpie Pica pica predation on Blackbird Turdus merula nests in urban areas. Bird Study 40: 55–62. doi: 10.1080/00063659309477129
- Hadjisterkotis, N. 2003. The effect of corvid shooting on the populations of owls, kestrels and cuckoos in Cyprus, with notes on corvid diet. Z. Jagdwiss. 49: 50–60.
- Herranz, J. 2000. Efectos de la depredación y del control de depredadores sobre la caza menor en Castilla-La Mancha. PhD Thesis, Autonoma University, Madrid, Spain.
- Jiguet, F. 2002. Arthropods in diet of Little Bustards Tetrax tetrax during the breeding season in western France. Bird Study 49: 105–109. doi: 10.1080/00063650209461253
- Kopij, G. 2005. Diet of some insectivorous passerines in semi-arid regions of South Africa. Ostrich 76: 85–90. doi: 10.2989/00306520509485478
- Kryštofková, M., Fousová, P. & Exnerová, A. 2011. Nestling diet of the common magpie (Pica pica) in urban and agricultural habitats. Ornis Fenn. 88: 138–146.
- López-Bao, J.V., Sazatornil, V., Llaneza, L. & Rodríguez, A. 2013. Indirect effects on heathland conservation and wolf persistence of contradictory policies that threaten traditional free-ranging horse husbandry. Conserv. Lett. 6: 448–455. doi: 10.1111/conl.12014
- Martí, R. & Del Moral, J.C. (eds) 2003. Atlas de las Aves Reproductoras de España. SEO/BirdLife y Direccion General de Conservacion de la Naturaleza, Madrid.
- Martínez, J.G., Soler, M., Soler, J.J., Paracuellos, M. & Sánchez, J. 1992. Alimentación de los pollos de urraca (Pica pica) en relación con la edad y disponibilidad de presas. Ardeola 39: 35–48.
- Miller, J.R. & Hobbs, N.T. 2000. Recreational trails, human activity, and nest predation in lowland riparian areas. Landscape Urban Plan 50: 227–236. doi: 10.1016/S0169-2046(00)00091-8
- Newson, S.E., Rexstad, E.A., Baillie, S.R., Buckland, S.T. & Aebischer, N.J. 2010. Population change of avian predators and grey squirrels in England: is there evidence for an impact on avian prey populations? J. Appl. Ecol. 47: 244–252. doi: 10.1111/j.1365-2664.2010.01771.x
- Ponz, A., Gil-Delgado, J.A. & Barba, E. 1999. Factors affecting prey preparation by adult Magpies feeding nestlings. Condor 101: 818–823. doi: 10.2307/1370069
- Reynolds, J.C. & Eabischer, N.J. 1991. Comparison and quantification of carnivore diet by faecal analysis: a critique, with recommendations, based on a study of the Fox Vulpes vulpes. Mammal Rev. 21: 97–122. doi: 10.1111/j.1365-2907.1991.tb00113.x
- Rivas-Martínez, S., Penas, A. & Díaz, T.E. 2004. Mapa Bioclimático de Europa, Bioclimas, http://www.globalbioclimatics.org/form/maps.
- Roos, S. & Pärt, T. 2004. Nest predators affect spatial dynamics of breeding red-backed shrikes (Lanius collurio). J. Anim. Ecol. 73: 117–127. doi: 10.1111/j.1365-2656.2004.00786.x
- Soler, J.J. & Soler, M. 1991. Análisis comparado del régimen alimenticio durante el periodo otoño-invierno de tres especies de córvidos en un área de simpatría. Ardeola 38: 69–89.
- Soler, M., Alcalá, N. & Soler, J.J. 1990. Alimentación de la grajilla Corvus monedula en tres zonas del sur de España. Doñana Acta Vertebrata 17: 17–48.
- Soler, J.J., Soler, M. & Martínez, J.G. 1993. Grit ingestion and cereal consumption in five corvid species. Ardea 81: 143–149.
- StatSoft. 2011. Statistica© (Data Analysis Software System), Version 10, Tulsa.
- Stoate, C. & Szczur, J. 2001. Could game management have a role in the conservation of farmland passerines? A case study from a Leicestershire farm. Bird Study 48: 279–292. doi: 10.1080/00063650109461228
- Suvorov, P., Svobodová, J., Koubová, M. & Dohnalová, L. 2012. Ground nest depredation by European Black-billed Magpies Pica pica: an experimental study with artificial nests. Acta Ornithol. 47: 55–61. doi: 10.3161/000164512X653926
- Teerink, B.J. 1991. Hair of West-European Mammals: Atlas and Identification Key. Cambridge University Press, Cambridge.
- Thomson, D.L., Green, R.E., Gregory, R.D., & Baillie, S.R. 1998. The widespread declines of songbirds in rural Britain do not correlate with the spread of their avian predators. Proc. R. Soc. Lond. B. 265: 2057–2062. doi: 10.1098/rspb.1998.0540
- Woodroffe, R., Thirgood, S. & Rabinowitz, A. 2005. People and Wildlife, Conflict or Co-existence? Cambridge University Press, Cambridge.
- Zar, J.H. 1984. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, NJ.