
Abstract
Capsule We analysed ring recovery data of Siskins from the EURING database (N = 42,707 individuals) and found 18 groups of birds that moved together a median distance of 200 km (range 51–1296 km). The median time elapsed between capture and recovery was 21 days (range 4–1390 days). These data suggest that Siskins have stable social bonds and move in stable social units.
Female choice of sexual partners in birds may be based on social familiarity as well as plumage ornaments or displays. Recently, Senar et al. (Citation2013) suggested that female Siskins Carduelis spinus consider social familiarity when choosing a mate. Hence, experiments with captive birds clearly showed that belonging to the same social flock enhanced the possibilities of an individual male being chosen by a female. The topic of familiarity is significant because by preferring familiar individuals as mates, the potential for local adaptation may be favoured (Joly Citation2011).
This work is based on the assumption that Siskin flocks are socially stable. Stability of membership and composition of bird groups have been studied in larger species, such as geese Anserini or crows Corvidae (Brown Citation1963, Jónsson & Afton Citation2008, Kurvers et al. Citation2013, Racine & Thompson Citation1983), and in sedentary species like tits, Paridae and Aegithalidae (Ficken et al. Citation1981, Hatchwell et al. Citation2001, Matthysen Citation1990, Nowicki Citation1983). Knowledge of social membership stability in small-sized migrant birds is more sparse (Shizuka et al. Citation2014), probably because of the difficulties in following the movements of their groups. Recently, however, some progress has been made in this area by analysing large data sets of ring recoveries. By analysing re-encounter data of birds ringed at the same location and period, and subsequently recaptured elsewhere together, Wilson (Citation2006) showed that Pine Siskins Carduelis pinus and Common Redpolls Acanthis flammea maintained some flock stability. Wilson restricted his analyses to birds ringed and re-encountered during the same winter. Here, we replicate his analyses with the Siskin but focus on longer distance movements over 50 km.
For this analysis, we used the ring recovery data for Siskin from the European Union for Bird Ringing (EURING) database. This data set contained 91,513 records of 42,707 individual Siskins, trapped between 5 January 1907 and 29 July 2011. We restricted analyses to those individuals that had moved a distance between ringing and recovery of at least 50 km, a movement which is further than the maximum movements that Siskin transients can achieve in a single day (10–40 km; Senar et al. Citation1992).
Individuals were defined as being in a group if they were trapped in the same locality on the same day. We used the program SOCPROG to detect associations (Whitehead Citation2009). The analysed data included ring number, date and locality; locality was transformed to a number. First, we defined the association based on both time and place, by including the variable [Day*100 + Locality]. We then computed the Joint Occurrences Association Index, saved it as a SOCPROG association matrix and opened it in the ‘Analysis of multiple association measures' module. Then we calculated dyadic values for the association index and sorted it, in MS Access, on association index, to assign individual Siskins to groups.
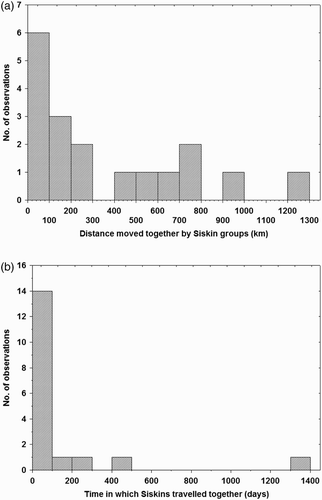
We found 18 groups of Siskins that travelled together. Most of the groups were formed by 2 individuals, 1 by 3, and 1 by 58 individuals that travelled a distance of 165 km in 19 days. The different groups were recovered at a median distance of 200 km, with one pair travelling together as far as 1296 km (a). The median time elapsed between ringing and recovery was 21 days (range 4–1390 days; b). Groups were either single-sex or mixed-sex, but the time for which the birds stayed together did not differ between single- and mixed-sex groups (F1,10 = 0.74, P = 0.41). Despite the fact that the sample size was small, single-sex groups travelled further (mean ± se = 572 ± 103 km) than mixed-sex groups (mean ± se = 180 ± 122 km; F1,10 = 6.0, P = 0.03).
Figure 1. Frequency distribution of (a) the distances moved by Siskin groups and (b) the time for which birds within groups remained together. All the groups, except two, were formed by two individuals ringed together (same locality and day) and again recovered together more than 50 km away from the ringing site. N = 18 groups.
Previous work analysing ringing data in wintering Pine Siskins, Common Redpolls and Golden-crowned Sparrows Zonotrichia atricapilla supported the idea of stability of group membership in small, non-sedentary passerines within the wintering period (Shizuka et al. Citation2014, Wilson Citation2006). We now extend the evidence with this analysis, which showed that individual Siskins can remain in the same group for as long as 1390 days (nearly 4 years) and can travel together for distances up to 1296 km. Although some groups were of mixed-sex and so could perhaps be breeding pairs, other groups were single-sex. These exclude the confounding issue of pair bonds and confirm that social units of Siskins can remain together, and form stable groups for long periods and across long distances. These results support the view that Siskins really form stable social units for long periods, a point that had been previously raised either by anecdotal observations (Cawthorne Citation1985) or work with captive birds (Senar et al. Citation1990). This stability of social groups may increase mate choice opportunities within the flock. As mate choice in Siskins takes place within flocks at the end of the winter (Senar & Borras Citation2004, Senar & Copete Citation1990), these data support the assumption that Siskin groups are stable enough to allow social familiarity to appear as a mating criterion during pair formation (Senar et al. Citation2013).
ACKNOWLEDGEMENTS
We thank EURING for allowing access to their data, and specially to Chris du Feu for his help and kindness. We thank Hal Whitehead for advice on the use of SOCPROG; Lluïsa Arroyo for help with the analysis and W.H.J. Wilson for earlier discussions on the paper.
ORCID
Juan Carlos Senar http://orcid.org/0000-0001-9955-3892
Additional information
Funding
REFERENCES
- Brown, J.L. 1963. Social organization and behavior of the Mexican jay. Condor 65: 126–153. doi: 10.2307/1365491
- Cawthorne, A. 1985. Ringing recoveries. BTO News 136: 11.
- Ficken, M.S., Witkin, S.R. & Weise, C.M. 1981. Associations among members of a black-capped chickadee flock. Behav. Ecol. Sociobiol. 8: 245–249. doi: 10.1007/BF00299522
- Hatchwell, B.J., Anderson, C., Ross, D.J., Fowlie, M.K. & Blackwell, P.G. 2001. Social organization of cooperatively breeding long-tailed tits: kinship and spatial dynamics. J. Anim. Ecol. 70: 820–830. doi: 10.1046/j.0021-8790.2001.00541.x
- Joly, E. 2011. The existence of species rests on a metastable equilibrium between inbreeding and outbreeding. An essay on the close relationship between speciation, inbreeding and recessive mutations. Biol. Direct 6: 62. doi: 10.1186/1745-6150-6-62
- Jónsson, J.E. & Afton, A.D. 2008. Lesser Snow Geese and Ross's Geese form mixed flocks during winter but differ in family maintenance and social status. Wilson J. Ornithol. 120: 725–731. doi: 10.1676/07-124.1
- Kurvers, R.H.J.M., Adamczyk, V.M.A.P., Kraus, R.H.S., Hoffman, J.I., van Wieren, S.E., van der Jeugd, H.P., Amos, W., Prins, H.H.T. & Jonker, R.M. 2013. Contrasting context dependence of familiarity and kinship in animal social networks. Anim. Behav. 86: 993–1001. doi: 10.1016/j.anbehav.2013.09.001
- Matthysen, E. 1990. Nonbreeding social organization in Parus. Curr. Ornithol. 7: 209–249.
- Nowicki, S. 1983. Flock-specific recognition of chickadee calls. Behav. Ecol. Sociobiol. 12: 317–320. doi: 10.1007/BF00302899
- Racine, R.N. & Thompson, N.S. 1983. Social organization of wintering blue jays. Behaviour 87: 237–255. doi: 10.1163/156853983X00444
- Senar, J.C. & Borras, A. 2004. Sobrevivir al invierno: estratégias de las aves invernantes en la Península Ibérica. Ardeola 51: 133–168.
- Senar, J.C. & Copete, J.L. 1990. Observación de alimentación de cortejo en Lúganos Carduelis spinus invernantes. Butll. GCA 7: 11–12.
- Senar, J.C., Camerino, M. & Metcalfe, N.B. 1990. Familiarity breeds tolerance: the development of social stability in flocking Siskins (Carduelis spinus). Ethology 85: 13–24. doi: 10.1111/j.1439-0310.1990.tb00381.x
- Senar, J.C., Burton, P.J.K. & Metcalfe, N.B. 1992. Variation in the nomadic tendency of a wintering finch Carduelis spinus and its relationship with body condition. Ornis Scand. 23: 63–72. doi: 10.2307/3676428
- Senar, J.C., Mateos-González, F., Uribe, F. & Arroyo, L. 2013. Familiarity adds to attractiveness in matters of siskin mate choice. Proc. R. Soc. Lond. B 280: 20132361. doi: 10.1098/rspb.2013.2361
- Shizuka, D., Chaine, A.S., Anderson, J., Johnson, O., Laursen, I.M. & Lyon, B.E. 2014. Across-year social stability shapes network structure in wintering migrant sparrows. Ecol. Lett. 17: 998–1007. doi: 10.1111/ele.12304
- Whitehead, H. 2009. SOCPROG programs: analysing animal social structures. Behav. Ecol. Sociobiol. 63: 765–778. doi: 10.1007/s00265-008-0697-y
- Wilson, W.H.J. 2006. Evidence of flock fidelity in common redpolls and pine siskins. N. Am. Bird Bander 31: 105–109.