
Abstract
Capsule House Sparrows preferred millet over seeds of prairie forbs and grasses native to the USA. By reducing the abundance and availability of some grains, changing agricultural practices may contribute to the population declines of House Sparrows seen in some parts of their range.
As human commensals, House Sparrows Passer domesticus occur on all continents except Antarctica, on many oceanic islands, and from sea level up to 4500 m (Anderson Citation2006). This distribution challenges the House Sparrow with a wide variety of environmental conditions. Studies document local climatic adaptations, often over remarkably short time periods, in a variety of morphological characters, including plumage colour and wing length (Johnston & Selander Citation1964), skeletal size (Johnston & Selander Citation1971), and morphology (Förschler & Geiter Citation2010).
House Sparrows favour agricultural grains over wild seeds; those of grasses and forbs native to the USA. In an early study, Judd (Citation1901) analysed gut contents in the USA and reported that over 70% of the House Sparrow's diet comprised cereal grain seeds, with small amounts of seeds of native grasses and forbs, and a miniscule quantity of insects. Generally, similar dietary preferences have been reported from around the globe (UK: Perkins et al. Citation2007; New Zealand: MacMillan Citation1981; Germany: Keil Citation1972; India: Beri et al. Citation1972).
Human selection on these cereal grains produce grains rich in carbohydrates, generally poorer in fats and proteins, and free of most toxins and secondary chemicals (Harlan et al. Citation1973, Wink Citation1988). The nutritional composition of these grains differs from their wild progenitors and from seeds of most native forb and grass species. While House Sparrows may be habitat generalists, as adults they may be dependent upon agricultural grains. Physiological studies indicate that digestive flexibility declines during development in House Sparrows (Caviedes-Vidal & Karasov Citation2001, Brzek et al. Citation2010), likely owing to nearly total granivory in the adult. The abundance and success of House Sparrows may be dictated by the availability of cereal grains either in agricultural fields, as spillage, as processed refuse (discarded bread or crumbs), or as birdseed, particularly millet Panicum miliaceum (Gavett & Wakeley Citation1986, Lowther & Cink Citation2006).
The evolution, success, and spread of the House Sparrow may truly be the product of the dawn of human agriculture and movement out of the Fertile Crescent of the Middle East. The success of the House Sparrow owes directly to the niche construction that humans inadvertently perform, unwittingly, on its behalf. Humans convert native habitat to an agriculture-dominated landscape. In these newly created landscapes, humans often cultivate the very foods that favour the adaptations and success of the House Sparrow.
Here, we report results of an experiment in which the seeds of plants native to the prairies of the upper Midwest, USA, were offered to House Sparrows in an urban residential habitat. We ask how consumption of the seeds of these native plant species compares to a classic ‘white’ grain: Proso Millet (a common constituent of commercial bird seed). We use the results of this experiment to propose a tentative hypothesis to explain both the spread, with humans, of House Sparrows around the globe, and its perplexing recent demise in some parts of it established range.
We measured the giving-up density (GUD – the amount of food left in a depletable patch when a forager quits the patch) of House Sparrows on seeds in experimental food patches to determine House Sparrow dietary preferences. A higher GUD indicates a less valuable or harder to find food, a higher perceived risk of predation, higher physiological stresses, or an environment with a higher background level of food (Brown Citation1988, Olsson & Molokwu Citation2007). Here, we controlled for the predation, missed opportunity, and physiological costs of foraging. Hence, differences in GUDs between food patches with different seed species reflect dietary preferences and/or conspicuousness.
Ten foraging stations were established in a residential backyard in the village of Oak Park, a suburb of Chicago, Illinois, USA. Each station consisted of 3 food patches placed side by side, and stations were placed approximately 5–7 m apart (ensuring that all foragers could easily see and forage from all stations). Patches consisted of plastic greenhouse saucers (32 cm diameter) containing 1.5 L of commercial bank sand as foraging substrate, into which 2 g of seed was well-mixed. At each of the 10 stations, 1 patch contained 2 g of millet, 1 contained 2 g of a native seed, and 1 contained 2 g of millet mixed with 2 g of that native seed. In a given foraging trial, each of the 10 stations offered 1 of the 10 native seeds. Native seed species were rotated amongst the ten stations following a Latin square, with the position of each patch type within the station randomized for each foraging trial. At the conclusion of the ten-day experiment, each station had offered seeds of each of the ten plant species and millet. The experiment was conducted under animal welfare protocol 11–082 and complies with all current laws of the USA.
Foraging data were collected on ten days from September through November 2007. During this time, 8–16 individuals were seen feeding from the patches, and 2 digital camera traps rotated among the patches confirmed that all foraging was by House Sparrows. Patches were available to sparrows from approximately 08:00 until 16:00, when any remaining food was sifted from the trays and placed into plastic specimen bags. Experimental foraging patches were then restocked and covered to prevent nocturnal foragers. The specimen bags with remaining food were transported to a laboratory at the University of Illinois at Chicago. Seeds from each bag were cleaned of debris, dried, and weighed. The weight of each sample represents the GUD for that seed species on that day, stratified by occurrence (alone or mixed).
GUDs from millet alone versus GUDs from millet mixed with a particular seed species allows one to compare millet with that seed species in terms of ease of finding, and true preference (reward per unit handling time; see Garb et al. Citation2000). For instance, the GUD on millet in its own patch may be lower than that of another seed in its own patch either because it is easier to find or because millet is preferred. When seeds are mixed together in a patch, the GUDs on each seed species should reflect how easily each is found. It is possible to tease apart properties of conspicuousness and preference by comparing selectivities (see below for details) for millet when seed species are alone (Sa) versus mixed (Sm) together in patches with another seed species. Sa > Sm indicates preference for millet. Sm > 0.5 indicates that millet is either easier to find within the food patch, or that the forager actually rejects some or all of the other seeds (e.g. expanding specialist diet strategy, Heller Citation1980, Brown & Mitchell Citation1989, Brown & Morgan Citation1995). Using selectivity for millet, we compared millet to ten different native seeds of Illinois. Sa = S m = 0.5 provides the null expectation for two seeds that do not differ in encounter probability or reward to handling time ratio.
To calculate the selectivities of the House Sparrow for millet with respect to each of the ten prairie seeds, we used Chesson's (Citation1983) modification of Manly's index:
where S equals the selectivity for prey type 1, N1 and N2 are GUDs for prey types 1 and 2, respectively, and R1 and R2 are the initial abundances of prey in the resource patch. Manly's index gives values between 0 and 1; when S = 1, that food type is always accepted and the alternative food type is always rejected. When S = 0, that food type is always rejected, and the alternative is always accepted. When S = 0.5, the 2 foods are harvested in proportion to their abundances. For our experiments this index provides 20 separate preference measures for millet (10 native seed species contingent upon mixed versus alone in patches).
We chose millet to serve as a standard for comparison and as a representative agricultural cereal grain, and ten native prairie species () to examine dietary preferences in the House Sparrow. House Sparrows readily consume and favour millet (Gavett & Wakeley Citation1986, Rosenberg & Bonney Citation1994). The ten native seed species are common in North American prairies, and all are frequently included in prairie restorations. The plant species were chosen because their seeds represent a range of seed masses and represent both grasses and forbs, factors known to influence diet selection in granivorous birds (Pulliam Citation1985, Diaz Citation1990). Native seeds were obtained from Prairie Moon Seed Company in Winona, Minnesota. We used commercial millet obtained from a local feed store.
Table 1. Native plant species used in seed selectivity foraging experiment with House Sparrows.
In 10 days of foraging, House Sparrows consumed 214 g (53.5% consumption rate) of millet, 71 g (35.5% consumption rate) of the 5 native grass species, and only 23 g (11.5% consumption rate) of the 5 native forb species. In all, the total amount of native seed consumed by House Sparrows was less than half of the total amount of millet consumed. These different levels of consumption are reflected in overall mean GUDs of 0.947 g for millet, 1.28 g for the native grasses, and 1.77 g for the native forbs. Sparrows harvested almost equal amounts of their millet from alone and mixed patches (53% from alone patches) while harvesting less of their native seeds from alone patches (31% from alone patches). This suggests that millet experiences little associational effects when co-occurring with the native seeds, whereas the native seeds experience greater consumption when mixed with millet (shared doom: Emerson et al. Citation2012).
House sparrow GUDs for the 10 native seed species had no relation to seed mass (P > 0.25) but they differed significantly from each other (F9,162 = 32.8, P < 0.001; A). In general, House Sparrows had higher GUDs on the forbs than on the grass species. House Sparrows had significantly lower GUDs on these native seeds when the seeds were mixed with millet (GUD = 1.46 ± 0.021 g) than when offered alone (GUD = 1.59 ± 0.021 g; F1,162 = 19.5, P < 0.001). This pattern of short-term apparent competition did not vary with the native seed species, and the interaction effect of seed species with mixed versus alone was not significant (F9,162 = 0.79, P > 0.6; A). Rudbeckia hirta was the only forb species for which the GUD from the mixed patch was greater than the GUD from the patch in which the seeds occurred alone (A).
Figure 1. GUDs and selectivities of House Sparrows in seed selection experiment. (A) GUDs of native seeds when offered alone (GUD Alone) and when offered together with 2 g millet (GUD mixed). The dotted line marks 1.8 g, or 90% of the offered 2 g of native seed. Species with GUDs above this line were considered non-foraged. The solid line marks 0.94 g, the mean GUD of millet. Different lower case letters above bars indicate significant differences in overall mean GUDs among the ten native plant species (Tukey HSD post hoc test). (B) Selectivities for millet when offered alone (Selectivity Alone) and when mixed with 2 g of each of 10 native seeds (Selectivity mixed). The dotted line marks 0.5, the selectivity value at which both seed types are consumed randomly (as encountered). Different lower case letters above bars indicate significant differences in overall mean selectivities for millet when paired with each of the ten native plant species (Tukey HSD post hoc test).
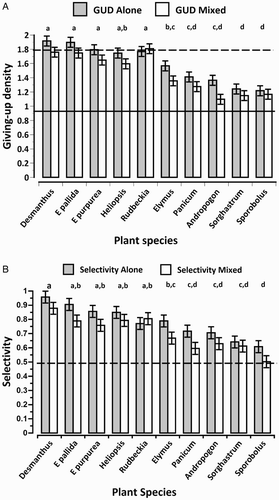
Post hoc Tukey honest significant difference (HSD) paired comparisons found no significant differences among the GUDs of the five forb species. The GUD of Heliopsis helianthoides, a forb, did not differ significantly from that of Elymus canadensis, a grass. The GUD of Heliopsis was significantly greater than those of each of the four remaining grass species. The GUD of Elymus did not differ from that of Panicum virgatum, but it was significantly greater than the other three grass species, Andropogon gerardii, Sporobolus heterolepis, and Sorghastrum nutans. The GUD of Panicum did not differ significantly from Andropogon, Sporobolus, and Sorghastrum.
House Sparrows' selectivity for millet depended strongly on the associated native species (F9,158 = 14.2, P < 0.001; B). House Sparrow selectivity for millet was greater when alone (Sa = 0.78 ± 0.14) than when mixed with each native seed species (Sm = 0.703 ± 0.13; F1,158 = 15.2, P < 0.001). Millet selectivity when alone or mixed was not influenced by the native seed species (non-significant interaction effect: F9,158 = 0.67, P > 0.7; B). For all native species, whether mixed or alone, selectivities were greater than 0.5: Sa > Sm > 0.5 indicating a preference for millet. Furthermore, either millet was always easier to find than the native seeds or when mixed together the sparrows actively sought out millet seeds while rejecting some of the native seeds within the same patch. The only exception to these patterns was for S. heterolepis where Sa > Sm = 0.5 suggesting a preference for millet but equal encounter probabilities and acceptances when the two seeds were mixed together in a patch.
The sparrows' GUDs on millet varied with the native species (F9,158 = 2.2, P = 0.024). Overall, millet GUDs were higher when mixed with a native seed (0.989 ± 0.033) than when alone (0.906 ± 0.034). This pattern held for seven of the native species, suggesting that the presence of the native seed species sometimes reduced the sparrows' perception of patch quality, perhaps because of the recognition time (Kotler & Mitchell Citation1995) required to distinguish the millet. This further suggests that Sm > 0.5 results from the active selection for millet – not because millet is easier to find.
Three of the forb seeds, Desmanthus illinoensis, Echinacea pallida, and Echinacea purpurea may not constitute actual foods for the House Sparrow. For each, the mean GUD is within 90% of the initial seed density, an amount that could reflect slight variation in initial seed density (determined by volume and not by weight). Rejection of these seeds may be due to nutritional content, secondary metabolites (toxins), or other seed properties that may affect energy:handling time ratios. Regardless, seeds of some of these native species are highly favoured by birds such as the native American goldfinch Spinus tristis in Illinois prairies. If House Sparrows are dietary specialists on domesticated grains and human grain products, then their distribution and abundance relative to native seed-eating birds may be dictated by the relative availabilities of domesticated grains versus native seeds. In this way plantings of native grasses and, especially, forbs may feed and encourage native birds and actually discourage House Sparrows.
The finding that House Sparrows prefer introduced cereals over native species could explain their historic success and also may contribute to their recent declines as availability of agricultural seeds decrease. Having evolved as humans domesticated early cereal grains in the Middle East, House Sparrows spread along with humans, taking advantage of the human-modified environment and benefiting from our agricultural crops. Now, changing agricultural practices (Hart Citation1986, Wilson et al. Citation2007, Schnitkey Citation2013), coupled with other cultural shifts in modern society, result in a general reduction in the abundance and availability of the major human cereal grains on which the species may be uniquely dependent. We see this as a tentative but important hypothesis. Our research provides an initial piece of support, and we hope that our work inspires additional tests. Specifically, similar diet selection experiments using different agricultural cereals as the standard may help elucidate the importance of these foods in House Sparrow population dynamics.
ACKNOWLEDGEMENTS
We thank two anonymous reviewers for helpful comments and suggestions on a previous draft of the manuscript.
REFERENCES
- Anderson, T.R. 2006. Biology of the Ubiquitous House Sparrow. From Genes to Populations. Oxford Univ. Press, Oxford.
- Beri, Y., Raizada, P.M. & Bhatnagar, R.K. 1972. Food in adults of common sparrow Passer domesticus Linnaeus. Entomol. Newslett. 2: 65–66.
- Brown, J.S. 1988. Patch use as an indicator of habitat preference, predation risk, and competition. Behav. Ecol. Sociobiol. 22: 37–47. doi: 10.1007/BF00395696
- Brown, J.S. & Mitchell, W.A. 1989. Diet selection on depletable resources. Oikos 54: 33–43. doi: 10.2307/3565894
- Brown, J.S. & Morgan, R.A. 1995. Effects of foraging behavior and spatial scale on diet selectivity: a test with fox squirrels. Oikos 74: 122–136. doi: 10.2307/3545681
- Brzek, P., Caviedes-Vidal, E. & Karasov, W.H. 2010. House Sparrow fledglings leave the nest digestively immature but more flexible than adults. Int. Comp. Biol. 50: E19–E19.
- Caviedes-Vidal, E. & Karasov, W.H. 2001. Developmental changes in digestive physiology of nestling House Sparrows, Passer domesticus. Physiol. Biochem. Zool. 74: 769–782. doi: 10.1086/322966
- Chesson, J. 1983. The estimation and analysis of preference and its relationship to foraging models. Ecology 64: 1297–1304. doi: 10.2307/1937838
- Diaz, M. 1990. Interspecific patterns of seed selection among granivorous passerines: effects of seed size, seed nutritive value and bird morphology. Ibis 132: 467–476. doi: 10.1111/j.1474-919X.1990.tb01064.x
- Emerson, S.E., Brown, J.S., Whelan, C.J. & Schmidt, K.A. 2012. Scale-dependent neighborhood effects: shared doom and associational refuge. Oecologia 168: 659–670. doi: 10.1007/s00442-011-2144-4
- Förschler, M.I. & Geiter, O. 2010. Morphological shifts in the House Sparrows Passer domesticus of Helgoland: insular syndrome or founder effects? Ornis Fenn. 87: 93–98.
- Garb, J., Kotler, B.P. & Brown, J.S. 2000. Foraging and community consequences of seed size for coexisting Negev Desert granivores. Oikos 88: 291–300. doi: 10.1034/j.1600-0706.2000.880207.x
- Gavett, A.P. & Wakeley, J.S. 1986. Diets of House Sparrows in urban and rural habitats. Wilson Bull. 98: 137–144.
- Harlan, J.R., deWet, J.M.J. & Glen Price, E. 1973. Comparative evolution of cereals. Evolution 27: 311–325. doi: 10.2307/2406971
- Hart, J.F. 1986. Change in the corn belt. Geog. Rev. 76: 51–72. doi: 10.2307/214784
- Heller, R. 1980. On optimal diet in a patchy environment. Theor. Pop. Biol. 17: 201–214. doi: 10.1016/0040-5809(80)90006-4
- Johnston, R.F. & Selander, R.K. 1964. House Sparrows: Rapid evolution of races in North America. Science 144: 548–550. doi: 10.1126/science.144.3618.548
- Johnston, R.F. & Selander, R.K. 1971. Evolution in the House Sparrow. II. Adaptive differentiation in North American populations. Evolution 25: 1–28.
- Judd, S.D. 1901. The Relation of Sparrows to Agriculture. USDA Division of Economic Ornithology and Mammalogy Bulletin 15. Government Printing Office, Washington, DC.
- Keil, W. 1972. Investigations of food of house- and tree sparrows in a cereal-growing area during winter. In Kendeigh, S. C., Pinkowski, J. (eds.) Productivity, Population Dynamics and Systematics of Granivorous Birds, 253–262. PWN-Polish Scientific Publishers, Warszawa.
- Kotler, B.P. & Mitchell, W.A. 1995. The effect of costly information in diet choice. Evol. Ecol. 9: 18–29. doi: 10.1007/BF01237693
- Lowther, P.E. & Cink, C.L. 2006. House Sparrow (Passer domesticus). In Poole, A. (ed.) The Birds of North America Online, Cornell Lab of Ornithology, Ithaca. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu.proxy2.library.illinois.edu/bna/species/012
- MacMillan, B.W.H. 1981. Food of House Sparrows and greenfinches in a mixed farming district, Hawke's Bay, New Zealand. New Zeal. J. Zool. 8: 93–104. doi: 10.1080/03014223.1981.10427946
- Olsson, O. & Molokwu, M.N. 2007. On the missed opportunity cost, GUD, and estimating environmental quality. Isr. J. Ecol. Evol. 53: 263–278. doi: 10.1560/IJEE.53.3.263
- Perkins, A.J., Anderson, G. & Wilson, J.D. 2007. Seed food preferences of granivorous farmland passerines: lark buntings prefer cereal grains whilst sparrows also take oily seeds. Bird Study 54: 46–53. doi:10.1080/00063650709461455.
- Pulliam, H.R. 1985. Foraging efficiency, resource partitioning, and the coexistence of sparrow species. Ecology 66: 1829–1836. doi: 10.2307/2937378
- Rosenberg, K.V. & Bonney, R. 1994. The national science experiments: birds like millet and milo too! Birdscope 8: 1.
- Schnitkey, G. 2013. Geographical acreage changes between 2006 and 2012 in corn, soybeans, wheat, and cotton. http://farmdocdaily.illinois.edu/2013/03/geographical-acreage-changes.html
- Wilson, J.D., Boyle, J., Jackson, D.B., Lowe, B. & Wilkinson, N.I. 2007. Effect of cereal harvesting method on a recent population decline of Corn Buntings Emberiza calandra on the Western Isles of Scotland. Bird Study 54: 362–370. doi: 10.1080/00063650709461496
- Wink, M. 1988. Plant-breeding importance of plant secondary metabolites for protection against pathogens and herbivores. Theor. Appl. Gen. 75: 225–233. doi: 10.1007/BF00303957