
ABSTRACT
Capsule: Northern Gannet Morus bassanus counts obtained from a coastal monitoring network in northern Spain were highly determined by wind conditions.
Aims: To determine how external factors including weather conditions affect seabird counts in a land-based seabird monitoring project (Red de observación de Aves y Mamíferos marinos, RAM project; Seabird & Marine Mammal Observation Network).
Methods: Seabird counts were obtained from four study capes situated in the southeastern part of the Bay of Biscay in northern Spain. Counts were conducted on a monthly-basis for a period of 3 hours starting at dawn on each sampling day. Data used here were compiled over a 10-year period (2006–15).
Results: Northern Gannet numbers were chiefly influenced by meteorological conditions at a local spatial scale. Northwesterly winds were found to positively influence gannet abundance, whereas high relative humidity caused a decrease in abundance. Moreover, gannets were more abundant during the non-breeding period as compared to breeding period.
Conclusion: We recommend considering the effect of the wind and relative humidity on land-based seabird counts in the Bay of Biscay when handling data from the RAM protocol in order to avoid biases and misinterpretation of data. Since seabirds approach the coast during harsh weather conditions, the RAM protocol should be improved in order to better account for the distance at which the birds are seen.
When monitoring seabirds, it is not only important to survey populations within their breeding range, but also to consider their migration routes and wintering areas. Such estimates of abundance have biological and ecological interest, but are also essential from a conservation and management standpoint (Mateos-Rodríguez et al. Citation2012). Land-based counts are an efficient technique for studying migrating pelagic seabird populations (Camphuysen et al. Citation2004), but these surveys have to consider the possible effects of weather conditions on data collected (Jones et al. Citation2014).
In Spain, such coastal counts have been performed since 2006 within the Iberian Seabird and Marine Mammal Monitoring Network (Red de observación de Aves y Mamíferos marinos (RAM) Valeiras et al. Citation2011). This project involved monthly land-based surveys during the whole annual cycle, in order to obtain a better understanding on the spatio-temporal distribution of seabirds along the coast of Spain. The Bay of Biscay in particular is an important migratory corridor for seabirds crossing southern European waters, either breeding in northern Europe to overwinter in the South Atlantic (Garthe et al. Citation2012), or breeding in the southern hemisphere to overwinter in the northern Atlantic (Brereton et al. Citation2003, Arcos Citation2009, Stenhouse et al. Citation2012, Jones et al. Citation2014, Louzao et al. Citation2015). In addition, recent papers have determined that the Bay of Biscay is an important wintering area for some seabird species (Fort et al. Citation2012). Since the Bay of Biscay is an obligate migratory corridor for a number of seabird populations, seabird counts can be used to monitor the long-term trends in seabird populations. Furthermore, the population status of marine predators, including seabirds, can be used as an indicator of the biodiversity and conservation status of marine ecosystems (Furness & Greenwood Citation1993, Mateos-Rodríguez et al. Citation2012).
Meteorological conditions could influence counts of seabirds along the coast of the Bay of Biscay. Specifically, under winds with a strong northwest component (Puillat et al. Citation2006), birds might drift towards inshore waters within the Bay of Biscay (i.e. southeastern parts of the Bay), so it could be predicted that seabird numbers would increase with strong northwest winds, but decrease in southeast winds. In the Bay of Biscay, northwest winds are often accompanied by unstable meteorological conditions, so it may also be predicted that seabirds will be more abundant on days of rain, with decreasing temperatures, higher relative humidity and low air pressures.
For seabirds moving from northwest France to northwest Iberia, passing through the southeastern part of the Bay of Biscay may entail a remarkable detour in their route of migration, which might be costly from an energetic viewpoint. Seabird abundance tends to decrease from the west to the east of the bay. Accordingly, seabird numbers are predicted to be higher at those capes located further to the west and, at the same time, should be expected to increase with strong northwesterly winds, with this effect being more marked at capes located further east.
The Northern Gannet Morus bassanus (hereafter Gannet) is a large marine avian predator that is widely distributed in the North Atlantic. It breeds in densely populated colonies in latitudes ranging between 48° and 66°N (Nelson Citation2002) and spends the winter within a broad geographic range from the North Sea area to the northwestern African coast. A fraction of the population has an intermediate distance of migration and overwinters within the Bay of Biscay (Fort et al. Citation2012). The Gannet is one of the main seabird species detected along the coast of Spain during the RAM counts (for details see the TREKTELLEN database, www.trektellen.nl). The RAM survey has been running over a period of 10 years, but the effect of meteorological conditions on seabird counts has never been assessed.
We used the Gannet as an avian model in order to test for the effect of meteorological conditions on the spatio-temporal abundance patterns of seabirds obtained through coastal counting methods. Specifically, we were interested in (1) describing abundance patterns of Gannets within the southeast part of the Bay of Biscay (from west to east: capes of Ajo, Matxitxako, St. Anton and Higuer) covering the 10 years from 2006 to 2015, and (2) investigating the role of weather on counts of Gannets explored at different spatial scales.
Methods
Data collection
The study was carried out using data obtained at four capes along the coast of the southeastern Bay of Biscay (), within the RAM survey (Higuer, 43°23′76″N 01°47′60″W; St. Anton 43°18′68″N 02°12′19″W; Matxitxako 43°27′27″N 02°45′4″W; Ajo 43°30′83″N 03°35′30″W). The census protocol consisted of counting seabirds from each cape, during a period of 3 hours starting at 07:00 UTC, every first Saturday of each month (http://redavesmarinas.blogspot.com.es). Accordingly, all counts were performed on the same day and time at all the capes regardless of weather conditions. Counts were performed using telescopes and binoculars. Between 30% and 50% of the sea under the horizon line was observed with the telescope and the remaining sea surface was observed with binoculars. The following variables were recorded on each sampling event: sighting conditions (semi-quantitatively: cloudiness, visibility, wind velocity and direction and sea conditions), the number of individuals seen for each seabird species, flight direction and behaviour (landed, moving randomly, migration, foraging movement, etc.) and the age of each individual (if possible). RAM volunteers used standard protocols and sheets to collect data. In each cape and sampling day, a single person was responsible for collecting the data, and there were always at least two people counting seabirds. The protocol did not consider any method to avoid double-counting. However, in most cases birds were observed to show single-direction movements (e.g. west- or east-wards migration), so double-counting was assumed to be minimal. Here, we use data collected on the Gannet over a period of 10 years (2006–15).
Figure 1. Map of the Bay of Biscay showing the area where the coastal counts were performed in the southeastern the Bay of Biscay.
Meteorological data
Meteorological data were obtained for the 2006–15 period from NCAR’s Research Data Archive (NCAR/UCAR, http://rda.ucar.edu, 2006–07) and the Galician Meteorological Agency (www.meteogalicia.es, 2008–15). A range of representative variables was selected including precipitation, temperature, relative humidity, pressure and the northwest-wind component. For each cape and survey day, we obtained the meteorological variables matching the closest count starting time. The precipitation data were represented as the rainfall (kg m−2) at sea level at the time of the census. We extracted the temperature (°C) and the relative humidity at 2 m above sea level. We used the mean sea level atmospheric pressure (Pa) to represent the atmospheric pressure at the time of the census. Wind field data were provided at a height of 10 m above sea level. The zonal u and meridional v winds describe the west–east and north–south direction and are represented by positive values toward the north and east, respectively. The two perpendicular components can be derived into the modulus of the wind speed in m s−1 (w) and direction (α in degrees, ranging from 0° to 360°). Wind speed and direction were estimated as follows: w = √(u2 + v2) and α = 180 + atan2(u,v) × (180/π). Then, we calculated the northwest-wind component as follows:(1) The northwest-wind component (T) takes into account the wind speed (w, m s−1), preferential wind direction (αt) and wind direction at the moment of the counts (αd). In this study, αt was considered the northwest wind since this wind direction causes the movement of seabirds into the Bay of Biscay. Therefore, the northwest-wind component would reach the highest values when the wind was blowing from the northwest.
Regarding the spatial scale of analysis, meteorological variables were extracted at different radii from each cape: 0 (local scale), 50, 100, 250 and 500 km. Meteorological data were provided on a grid basis, hence for a given spatial scale (radius) we calculated the median values of those grid cells within the radius, after removing grid points over the continent.
Meteorological data were divided into four time periods based on the Gannet’s life cycle: wintering (November–January), pre-breeding (February–March), breeding (April–August) and post-breeding (September–October). Such periods were determined according to already existing knowledge on the timing of the birds’ life cycle (Fort et al. Citation2012).
In order to determine the temporal scale that better explained Gannet counts, we investigated the correlation (Pearson’s r) between Gannet counts at one specific day and the meteorological variables conditions from that day to the previous 15 days of the observation in a 3-hourly basis to match the temporal conditions of counts. Therefore, we obtained 120 values of correlation for each count. In the correlation analysis, all variables showed maximum correlation to the moment of the census, and the values of variables in the days before had less influence than the meteorological conditions at the time of the survey (see online Figure S1). Therefore, we considered only the variables at the moment of the census. Moreover, we analysed the correlation between all the meteorological variables. The analysis showed a low correlation (Pearson’s r) in all analysed pairs (online Figure S2).
Statistical analysis
To test for the effect of meteorological variables on Gannet counts (the response variable), we used generalized linear models (GLMs) with a log-linear link function and a negative binomial error distribution. Predictor variables considered were: cape, period, northwest-wind component, precipitation (as binary variable, rain/no rain), temperature, relative humidity and air pressure, as well as all the two-way interactions of cape with the other variables.
Before starting to select models to determine which variables had an effect on Gannet counts, we determined which spatial scale was better at explaining Gannet counts. With that goal, we ran all the possible models for each radius. To determine how alternative models fitted to the data, we used Akaike’s Information Criterion corrected for small sample sizes (AICc) (Akaike Citation2011). According to this approach, models with a lower AICc value show a better fit to the data, because they explain a greater amount of variance with a (reduced) number of parameters. Thus, we calculated the mean and the 95% confidence interval of all models run for each spatial scale and found that, overall, there was a high overlap between spatial scales, with the mean AICc for the models considering a radius of 0 km being slightly lower than the mean of models considering larger spatial scales. Therefore, meteorological data at a local spatial scale (radius: 0 km, data for the grid cell that contained each cape) were best at explaining Gannet counts.
Model selection was carried out using a selection procedure based on AICc values. Models were considered to fit the data equally well when they differed by less than 2 AICc units (Williams et al. Citation2002). All statistical models were run with R software (R Core Team, Citation2014).
Results
Regarding abundance patterns at a local scale, the model with the lowest AICc value included an effect of period, cape, northwest-wind component, precipitation, temperature, relative humidity and pressure and all the two-way interactions of cape with the rest of the variables on Gannet counts (). This top model showed an AICc difference higher than 20 when compared with the second one, so hereafter we will only consider the top model.
Table 1. Mean (±se), range and total number of Gannets counted by hour and sampling day during the 3-hour survey period at each sampling cape and period (wintering, pre-breeding, breeding and post-breeding) within the Bay of Biscay, for the period 2006–15. The number of sampling hours is shown in parenthesis.
A total of 88 781 Gannets were observed over 1101 hours in 367 days of observations across the 10 years period (). Overall, we observed a mean passage rate of 81 Gannets/h. The mean passage rate tended to decrease from west (Ajo: 117 Gannets/h) to east (Higuer: 58 Gannets/h), but the difference was non-significant ().
Table 2. Beta-parameter estimates (±se, P values) obtained from the best model used to predict coastal Gannet counts in the southeastern Bay of Biscay. Only significant parameter estimates are shown.
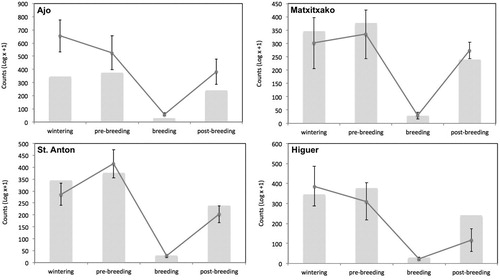
Globally, Gannets were more abundant during the non-breeding period (from September to March; , ). The GLM showed that numbers of Gannets did not vary significantly among capes, but there was a significant interaction of period × cape (), indicating that there were local differences in Gannet abundance during specific periods (). In general, Gannets were significantly more abundant during the post-breeding, wintering and pre-breeding periods than in the breeding period at the four capes ().
Figure 2. Mean (±se) number of Gannets counted by cape and four phenological periods. Grey bars represent the global mean number of counted Gannets for all four capes.
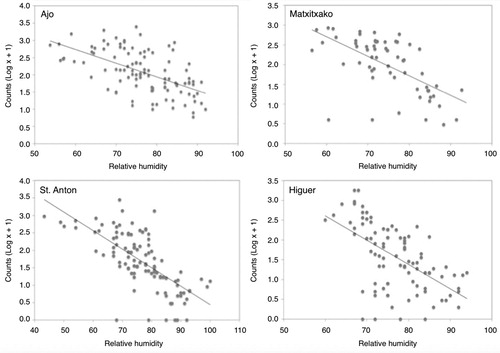
The northwest-wind component showed a positive effect on Gannet counts. Hence Gannets were more abundant with strong northwest winds, while they were scarcer under southeast winds (). The lack of a significant interaction of this variable with cape allows us to conclude that the wind effect was generalized and did not vary at a local level (). The relative humidity showed an overall significant negative correlation with Gannet counts (). In this case we also detected a significant interaction with cape (); at St. Anton, the effect of the variable was stronger than in the other three capes (). The other three meteorological variables did not have any significant effect on bird counts ().
Figure 3. Gannet counts in the capes in relation to northwest-wind component values. Data from the four study capes have been pooled due to the lack of a significant cape × wind interaction ().
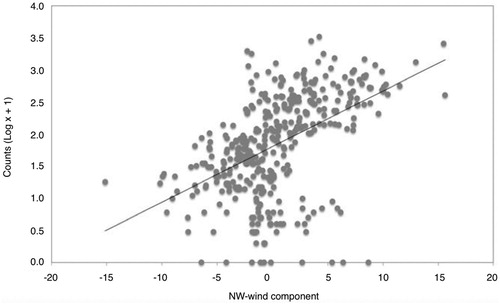
Figure 4. Relationship between relative humidity and Gannet counts at the capes. Relative humidity values were calculated for each count value (log x + 1) for each cape separately. The four capes are represented individually since there were differences in the relative humidity effect between the capes (P < 0.05).
Discussion
This is the first study to determine the effect of meteorological conditions on counts of a seabird species from the coast in the southeastern part of the Bay of Biscay. Overall, the number of Gannets counted at four selected capes situated along a coast transect of 151 km did not differ between capes, while a month × cape interaction showed slight temporal variations at a local scale level.
In general, Gannets showed maximum numbers during the non-breeding period (from September to March), coinciding with the migration and wintering of the species. Moreover, many Gannets choose this area to spend the wintering period (Fort et al. Citation2012). Therefore, the area is characterized as an important migration corridor but also as a major wintering area for the species. These wintering birds have been found to come from different colonies of northern Europe: Starstappen (Norway), Store UlvØyholmen (Norway), Bass Rock (Scotland, UK) and Grassholm (Wales, UK) (Fort et al. Citation2012). Wintering areas for colonies are mainly linked to the respective latitude of each colony and strongly suggest a chain migration pattern in which populations move uniformly southward, with more northern breeders wintering in northernmost grounds (Newton Citation2008, Fort et al. Citation2012). The seasonal patterns detected in our study fit well with the phenological cycle of the species and support previous studies indicating that Gannets are present in the Bay of Biscay during the whole non-breeding period, either during the migration period or as wintering birds (Abad et al. Citation2006, Fort et al. Citation2012, Garthe et al. Citation2012).
The number of Gannets recorded tended to increase under northwest winds. The causes underlying this pattern remain unknown to us: a possibility is that birds are blown by these northwest wind towards the shore and, therefore, are more easily detected from capes, which result in higher numbers; an alternative explanation is that northwest winds would push the rich-of-fish upper water layers closer to shore (as in the Baltic Sea), and Gannets follow them. Probably both are true, but this has consequences in terms of data interpretation. Given this result, it would be worth obtaining some kind of standardized population index, hence allowing us to adjust the counts to meteorological conditions, (as it has been done for other areas; Walker Citation1996, Abad et al. Citation2006, Mateos & Arroyo Citation2011). For that goal, we recommend to consider all capes situated along the coast of northern Spain in order to obtain an index that would be applicable to the entire region, and not only to the southeastern part of the Bay of Biscay (this article).
Moreover, we also observed that Gannet numbers tended to decrease with increasing relative humidity. This result is consistent with previous studies that have demonstrated that humidity, along with other environmental factors, can shape the daily movement patterns and behaviour of seabirds (Moore et al. Citation2008). In addition, high relative humidity can affect counts as visibility is reduced when this parameter increases. Interestingly, at one of the capes (St. Anton) the effect was even stronger when compared to the other three study capes, but the reason behind this finding remains unknown to us. Meteorological factors (fog, sun glare, humidity) can influence counts directly and limit the visibility for seabird detection (Tasker et al. Citation1984).
In conclusion, Gannet counts from a number of capes of a coastal monitoring network in the southeastern Bay of Biscay were mostly determined by period and two meteorological factors: northwest-wind component and relative humidity. Our results suggest that the programme was adequate to monitor the species’ abundance pattern throughout the annual cycle and might be used to estimate population trends within the region. However, count analysis must control for the effect of weather, and in this context future studies should try to obtain standardized indices of seabird counts. Accounting for the distance at which the birds are observed from each cape is also recommended.
Supplemental Material
Download MS Word (1 MB)Acknowledgements
We are grateful to the volunteers and entities (Sociedad Ornitológica Lanius, Arkamurka, Itsas Enara Ornitologi Elkartea, SEO/BirdLife, GIAM) supporting bird counts at the study capes. We also thank the two RAM Spanish General Coordinators (Xulio Valeiras, Salvador García). Almudena Fontán provided useful comments on meteorological conditions in the Bay of Biscay. This study is a contribution to the CHALLENGES (CTM2013-47032-R) project. Two anonymous referees provided very valuable comments that helped us to improve an earlier version of this work.
Additional information
Funding
Related Research Data
References
- Abad, E., Valeiras, X., Serrano, A., Sánchez, F., Preciado, I. & Olaso, I. 2006. Influence of fisheries discards and environmental variables on seabirds in northern Spanish waters (Cantabrian Sea) ICES CM 50:19.
- Akaike, H. 2011. Akaike’s Information Criterion. In: Lovric, M. (ed) International Encyclopedia of Statistical Science. Springer, Berlin.
- Arcos, J. M. 2009. Áreas importantes para la conservación de las aves marinas en España: Important areas for the conservation of seabirds in Spain. Sociedad Española de Ornitologia (SEO/BirdLife).
- Brereton, T. M., Weir, C., Hobbs, M. & Williams, A. D. 2003. A low-cost, year-round seabird monitoring programme in the English channel and Bay of Biscay: preliminary results 1995-2001. Ornis Hungarica 12: 105–113.
- Camphuysen, C. J., Fox, A. D., Leopold, M. F. & Petersen, I. K. 2004. Towards Standardised Seabirds at Sea Census Techniques in Connection with Environmental Impact Assessments for Offshore Wind Farms in the UK: a comparison of ship and aerial sampling methods for marine birds and their applicability to offshore wind farm assessments. Cowrie.
- Fort, J., Pettex, E., Tremblay, Y., Lorentsen, S. H., Garthe, S., Votier, S., Pons, J.B., Siorat, F., Furness, R. W., Grecian, W. J., Bearhop, S., Montevecchi, W. A. & Grémillet D. 2012. Meta-population evidence of oriented chain migration in northern gannets (Morus bassanus). Front. Ecol. Environ. 10: 237–242. doi: 10.1890/110194
- Furness, R. W. & Greenwood, J. J. D. (eds) 1993. Birds as Monitors of Environmental Change. Springer, Dordrecht.
- Garthe, S., Ludynia, K., Hüppop, O., Kubetzki, U., Meraz, J. F., Furness & R. W. 2012. Energy budgets reveal equal benefits of varied migration strategies in northern gannets. Mar. Biol. 159: 1907–1915. doi: 10.1007/s00227-012-1978-6
- Jones, A. R., Wynn, R. B., Yésou, P., Thébault, L., Collins, P., Suberg, L., Lewis, K. M. & Brereton, T. M. 2014. Using integrated land-and boat-based surveys to inform conservation of the Critically Endangered Balearic shearwater Puffinus mauretanicus in northeast Atlantic waters. Endangered Species Res. 25: 1–18. doi: 10.3354/esr00611
- Louzao, M., Afán, I., Santos, M. & Brereton, T. 2015. The role of climate and food availability on driving decadal abundance patterns of highly migratory pelagic predators in the Bay of Biscay. Front. Ecol. Evol. 3: 90. doi: 10.3389/fevo.2015.00090
- Mateos, M. & Arroyo, G. M. 2011. Ocean surface winds drive local-scale movements within long-distance migrations of seabirds. Mar. Biol. 158: 329–339. doi: 10.1007/s00227-010-1561-y
- Mateos-Rodríguez, M., Thomas, L. & Arroyo, G. M. 2012. The development and use of a method to fill time gaps in migration counts. Condor 114: 513–522. doi: 10.1525/cond.2012.100053
- Moore, A. L., Damania, S. P., Henson, S. M. & Hayward, J. L. 2008. Modeling the daily activities of breeding colonial seabirds: dynamic occupancy patterns in multiple habitat patches. Math. Biosci. Eng. 5: 831–842. doi: 10.3934/mbe.2008.5.831
- Nelson, B. 2002. The Atlantic Gannet, 2nd ed. Fenix Books, Norfolk.
- Newton, I. 2008. The Migration Ecology of Birds. Academic Press.
- Puillat, I., Lazure, P., Jegou, A. M., Lampert, L. & Miller, P. 2006. Mesoscale hydrological variability induced by northwesterly wind on the French continental shelf of the Bay of Biscay. Sci. Mar. 70: 15–26. doi: 10.3989/scimar.2006.70s115
- R. C. Team 2014. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
- Stenhouse, I. J., Egevang, C. & Phillips, R. A. 2012. Trans-equatorial migration, staging sites and wintering area of Sabine’s Gulls Larus sabini in the Atlantic Ocean. Ibis 154: 42–51. doi: 10.1111/j.1474-919X.2011.01180.x
- Tasker, M. L., Jones, P. H., Dixon, T. & Blake, B. F. 1984. Counting seabirds at sea from ships: a review of methods employed and a suggestion for a standardized approach. Auk 101: 567–577.
- Valeiras, X., García, S. & Abad, E. 2011. Observación de aves marinas desde cabos: la Red de observación de Aves y Mamíferos marinos (RAM). In Valeiras, X., Muñoz, G., Bermejo, A., Arcos, J.M. & Paterson, A.M. (eds) Actas del VI Congreso del GIAM y el Taller internacional sobre la Ecología de Paiños y Pardelas en el sur de Europa. Boletín del Grupo Ibérico de Aves Marinas, 37–42.
- Walker, F. 1996. Observations of movements of North Atlantic Gannets Morus bassanus from Cape St Vincent, Portugal. Seabirds 18: 44–48.
- Williams, B. K., Nichols, J. D. & Conroy, M. J. 2002. Analysis and Management of Animal Populations: modeling, estimation, and decision making. Academic Press, San Diego, CA.