
ABSTRACT
Capsule: Grazing by livestock can have complex effects on drivers of population change in the Clamorous Reed Warbler Acrocephalus stentoreus and Dead Sea Sparrow Passer moabiticus.
Aims: To investigate the effect on two specialist bird species on wetland degradation in the Jordan Valley.
Methods: The direct and indirect effects of grazing on the probability of occurrences of two specialist bird species, Clamorous Reed Warbler A. stentoreus and Dead Sea Sparrow P. moabiticus, were analysed during the breeding season at the patch scale, using path analysis.
Results: Tamarix shrub density was a strong predictor for the presence of both species. Grazing had a negative total effect on both; a significant indirect effect on Dead Sea Sparrow via its impact on the mean height of shrubs, and a significant, negative indirect effect on Clamorous Reed Warbler by reducing reed cover. Intensive grazing and browsing by livestock including goats, sheep and camels, apparently had a negative effect on the overall density of native Tamarix shrubs, while promoting encroachment by invasive Prosopis juliflora.
Conclusion: This may be part of a long-term cascade leading to an ecological transition and loss of important wetland habitats in the arid Jordan Valley.
Degradation of dry rangelands can lead to a reduction of the vegetation cover, and subsequent increase in soil erosion and stream flow (Wilcox Citation2007, D’Odorico et al. Citation2013). This degradation is particularly manifested where pastoral rangeland exists in combination with cultivated land use (Millennium Ecosystem Assessment Citation2005). Wetland habitat patches typical of the arid Jordan Valley have been impacted by an increasing need for limited water resources, agricultural expansion and overgrazing (Albert et al. Citation2004, Waitzbauer & Petutsching Citation2004a), which apparently facilitated the recent spread of invasive plant species (Dufour-Dror & Shmida Citation2017).
Previous studies in arid environments found that a decrease in natural vegetation cover lowers bird diversity (Tomoff Citation1974, Newton & Newton Citation1997, Khoury et al. Citation2007). Overgrazing in drylands apparently reduces bird diversity in a bottom-up cascade by reducing standing biomass, food abundance and habitat complexity (Joubert & Ryan Citation1999, Ogada et al. Citation2008). Grazing may influence various species differently, that is positively or negatively, directly or indirectly. Thinning of the shrub layer is one of the indirect effects that may increase access to competitors and predators (James Citation2003, Krueper et al. Citation2003). In various drylands across the world, including the Jordan Valley, grazing is contributing to the spread of alien, invasive shrub species like Prosopis juliflora (Dufour-Dror & Shmida Citation2017). P. juliflora has several features that make it a powerful and noxious invader (Pasiecznik et al. Citation2001, Shiferaw et al. Citation2004). Invasive Prosopis can have multiple effects on ecosystems and bird habitats, for example, by lowering groundwater tables and outcompeting native shrubs and trees (Dzikiti et al. Citation2017, Kumar & Mathur Citation2014), some of which appear to be important for specialist birds in the Jordan Valley (Khoury & Korner Citation2018). The spread of invasive alien plants can significantly reduce food abundance and availability of suitable nesting sites for birds (Skórka et al. Citation2010, Schirmel et al. Citation2016).
It has previously been demonstrated that the breeding occurrence of various birds that build nests in reeds and shrubs was lower in habitat patches that were intensively grazed by livestock (Khoury & Korner Citation2018). However, the exact mechanism of grazing impact was not clarified. Birds can be affected by grazing during the breeding season either directly, by disturbance and nest destruction, or indirectly, by reducing vegetation cover important for successful breeding, and by facilitating bio-invasions in the breeding habitats. Thus we used a path analysis to investigate how grazing affects the occurrences of two specialist bird species at the local patch scale (Clough Citation2012, Korner et al. Citation2018). Quantifying the relative magnitudes of direct and indirect effects of land use will help in guiding conservation and restoration actions (Feeley & Terborgh Citation2008). For example, if intensive grazing has a negative indirect effect via the change in vegetation features, management could be adapted to regulate grazing spatially and temporally, to allow the regeneration of native vegetation. But managing grazing alone may not be sufficient in all cases, and further interventions may be necessary to minimize indirect effects previously facilitated by grazing, such as the advanced encroachment by alien species.
In this study, we used two resident species, the Clamorous Reed Warbler Acrocephalus stentoreus, an insectivore and the Dead Sea Sparrow Passer moabiticus, a granivore. Both have a restricted breeding range and are specialized on marsh-like/riparian-like habitats. These two species often shared the same habitat but occupied different niches as their diets and behaviour differ considerably. We recorded two further specialist species of wetland habitats with a restricted breeding distribution in Jordan, the Little Bittern Ixobrychus minutus and Common Nightingale Luscinia megarhynchos, but the frequency of their occurrences was too low to carry out further analysis. Specialist bird species are considered important in conservation, and are useful indicators of ecosystem integrity (Julliard et al. Citation2006), and indicators of processes related to invasion of alien plants in salt marsh and riparian habitats (Ma et al. Citation2011, Hajzlerová & Reif Citation2014) and desertification in drylands (Whitford Citation1997).
Methods
Study area
Fieldwork was conducted along the narrow and shallow tributaries of the River Jordan (in this text also called wadis), in the flat parts of the Jordan Valley below sea level. The mean annual precipitation is around 100 mm and mean annual temperature of 23°C. Most rain falls in winter when temperatures are mild, whereas the summer is very hot and dry (45°C absolute maximum temperature; Waitzbauer & Petutsching Citation2004b). The wadis were surrounded by an arid landscape mostly used as rangeland, or intensive, irrigated agriculture. Agricultural expansion in the Jordan Valley caused widespread degradation and fragmentation of wetland habitats (Andrews Citation1995, Albert et al. Citation2004). Bedouin camps were frequent in accessible parts and this caused localized overgrazing by livestock (sheep, goats and camels) and woodcutting. The wadis usually contained continuous or intermittent perennial streams, locally flowing slowly to create marsh-like conditions with reeds Phragmites australis, which were always associated with water availability, Tamarisk shrubs Tamarix nilotica and Tamarix tetragyna, cat-tail, sedges and grasses. The current sources of the brackish water were springs and/or treated water from wastewater plants, drainage from farms and small desalination plants. The width of the stream with its dry banks usually ranged from 10 to 100 m; water depth varied from a few centimetres to around 50 cm. Disturbed sections (e.g. intensively grazed) had either impoverished vegetation or were encroached by alien P. juliflora shrubs.
Bird species
The Dead Sea Sparrow breeds in shrubs (mainly Tamarix) in lowland areas with water. It has a disjunct distribution from Turkey, Cyprus and parts of the Middle East, east to Afghanistan (Snow & Perrins Citation1997). In Jordan, it is mainly a resident with some dispersion after the breeding season. Its breeding range is restricted to a narrow area in the Jordan Valley, namely the banks of the River Jordan, along some of its tributaries, and along drainage canals southeast of the Dead Sea. Breeding colonies near the northern and southeastern shores of the Dead Sea have mostly disappeared during the last two decades (Andrews Citation1995, F. Khoury pers. obs.). Tamarix thickets growing on or near inundated or moist ground and slow-flowing streams or drainage canals are the typical breeding habitat of Dead Sea Sparrow in Jordan, but they usually feed on seeds in adjacent fields and scrub vegetation (Khoury et al. Citation2006, Khoury & Korner Citation2018). This species builds huge nests in high Tamarix shrubs, occasionally in cultivated citrus trees, date palms, ornamental shrubs, and rarely also P. juliflora, if located within or very close to its native habitat. Dead Sea sparrows tend to form very loose colonies of various size and density.
The Clamorous Reed Warbler occurs as several subspecies that breed from Egypt eastwards through Asia to China (Snow & Perrins Citation1997). In Jordan, it is currently restricted to the Jordan Valley and its margins (Andrews Citation1995) where it breeds in mature reed beds and thickets (preferably Tamarix) mixed with reed and sedge growing on inundated ground and along streams; it occasionally occurs in the same habitat with the Dead Sea Sparrow, preferring the densest vegetation, where territorial males can easily be heard singing during the breeding season.
Line transects
Fifty-eight line transects, each with a length of 200 m, were carried out during the breeding season of 2016 to count birds along the wadis (). The line transect method was adopted here instead of point counts due to the linear, narrow shaped and relatively open habitat (Bibby et al. Citation2000). Locations were selected randomly, but several had to be relocated due to accessibility restrictions. This caused some spatial clustering, but transects were generally well spaced, with a minimum distance of 200 m between neighbouring transects (). Each transect was assessed twice between late March and early June, and the larger count was considered for analysis. The observer recorded all birds seen or heard within the shallow wadis to a distance of 50 m while walking slowly along the wadi. In the case of both species, these were singing males, and for further analysis, we used presence/absence data only. Dense reedbeds did not hinder walking along transects as they occurred locally only as patches (up to 30 m broad) or narrow strips along streams. The surveys were carried out by the same observer (F. Khoury) and performed in the morning from sunrise to around 10:00 am. Habitat variables were quantified at three points (near the two ends and at the middle) along each line transect to a range of 50 m, then summed up for the entire transect. The variables included mainly structural attributes related to the vegetation in addition to signs of grazing intensity by goats, sheep and/or camels ().
Figure 1. Distribution of transects in the study area; lines indicate clustering of more than one transect along the wadis, see text for details.
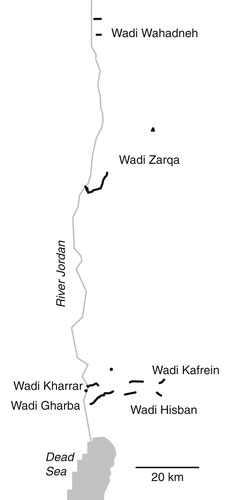
Table 1. Variables used in the models with range, mean and standard deviation of the observed values in n = 58 transects.
Data analysis
We explored the interrelationships between grazing, Tamarix sp., P. juliflora, reed and the occurrence probability of each of the two studied bird species using linear models in a path analysis (Clough Citation2012). Path analysis was used to disentangle the effects of grazing on the bird presence that operate through indirect effects on Tamarix, Prosopis and/or reed from remaining, direct effects of grazing on the bird presence. In this way, path analysis attempts to model cascading effects.
Our path analyses consist of three (Dead Sea Sparrow) or four (Clamorous Reed Warbler) sub-models ( & , ): Simple linear regressions predicting invasive Prosopis, mean height of shrubs and reed (for Clamorous Reed Warbler only) from grazing, and a multiple logistic regression (with a logit-link function) predicting the presence probability of the target species dependent on grazing, Prosopis, mean height of shrubs, reed (Clamorous Reed Warbler) and Tamarix. Tamarix was added here as a separate predictor rather than building it into a path grazing-Tamarix-species occurrence because the relationship between grazing and Tamarix does not appear to be causal in one direction. With less grazing, Tamarix is most common and, as expected, decreases with increasing grazing. However, at high Tamarix density values, grazing is again intense due to the attractiveness of accessible habitat with dense Tamarix for shepherds (according to our observations and short interviews with the shepherds).
Figure 2. Path analysis used for the Dead Sea Sparrow occurrence probability, with estimated slopes for the simple linear regressions of P. juliflora and mean shrub height (MHS) on grazing, and with estimated partial effects of a logistic regression (with logit-link) predicting Dead Sea Sparrow occurrence from the other four variables. Numeric variables centred and scaled. Values are point estimates and 95% intervals in brackets.
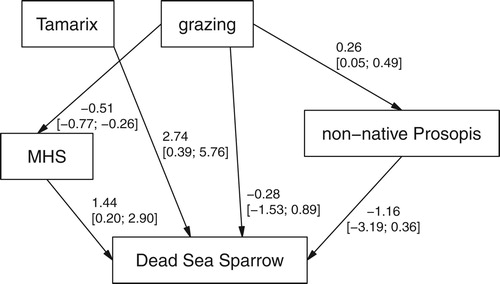
The path model included three indirect effects of grazing on the target bird species: (a) the spread of P. juliflora which indicates disturbances and alters the breeding habitat by affecting the native vegetation cover (e.g. by competition), (b) the reduction of mean shrub height (mainly consisting of Tamarix) by browsers (e.g. camels) and shepherds who cut large branches of shrubs for firewood, and (c) the reduction of reed cover by grazing livestock (only for the Clamorous Reed Warbler model). Grazing, in addition, may also have direct effects on the target species, reflecting possible true direct effects (e.g. disturbance at nesting sites, destruction of nests by livestock) as well as various other indirect effects via covariates not included in the model (e.g. effects of grazing activities on water and nutrient availability). All numeric variables were centred and scaled. In all sub-models, we included a random factor ‘area’, which identified transects that were from the same wadi or section of a wadi (there were 12 areas with 1–13 transects each), hence the sub-models were hierarchical models. The sub-models were fitted within a Bayesian framework using the open-source statistical software R (linear regressions; R 3.4.3, R Core Team 2017) and Stan (logistic regression; used via rstanarm 2.17.3 in R, Stan Development Team 2017). Weakly informative priors were used (we used the Stan default setting, i.e. priors that aim not to influence the posterior distribution) and 4000 samples from the posterior distribution were generated. For error propagation along the paths, we followed the analyses outlined by Clough (Citation2012). Model assumptions were tested graphically (e.g. observed vs. fitted values, residuals vs. predictors).
Results
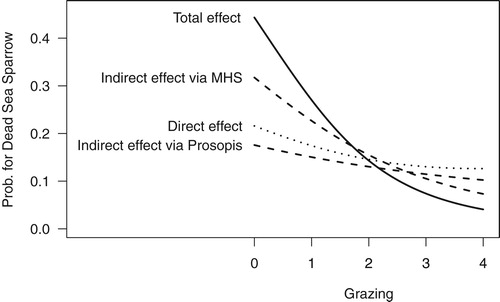
Dead Sea Sparrows were recorded on 28%, Clamorous Reed Warbler on 19% of the transects. The expected positive effect of Tamarix density was obvious on both species ( & ). The two species existed as breeding species only at the highest Tamarix scores. The correlation between Tamarix and grazing was not, as might be expected, negative and linear (; see also methods and discussion). The direct effect of grazing on Dead Sea Sparrow presence was low and non-significant (; see below), whereas the total effect (sum of direct and indirect effects) of grazing on Dead Sea Sparrow occurrence probability was relatively strong and significant (estimate = −1.23, 95% intervals = −2.55 and −0.13, ). Much of the total grazing effect on the Dead Sea Sparrow appears to be mediated through the effect of grazing on mean shrub height (estimate = −0.62, 95% intervals = −1.37 and −0.07, ).
Figure 3. Path analysis used for the Clamorous Reed Warbler occurrence probability, with estimated slopes for the simple linear regressions of P. juliflora, mean shrub height (MHS) and reed on grazing, and with estimated partial effects of a logistic regression (with logit-link) predicting Clamorous Reed Warbler occurrence from the other five variables. Numeric variables centred and scaled. Values are point estimates and 95% intervals in brackets.
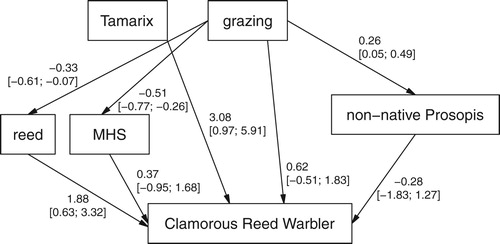
Figure 4. Observed correlation between grazing and Tamarix density (small grey dots, jittered to prevent overlap). Mean Tamarix values per grazing score are given as larger black dots connected with a line.
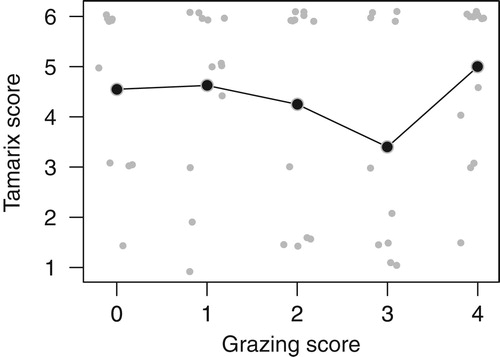
Figure 5. Total, direct and indirect effects of grazing on Dead Sea Sparrow occurrence probability from the path analysis (). For the graph, the Tamarix density value was set to 5.5 since Dead Sea Sparrows appear to be very scarce at Tamarix values below 5. See text for effect sizes and uncertainties.
The direct effect of grazing on Clamorous Reed Warbler occurrence was not significant and with a large uncertainty (estimate = 0.62, 95% intervals = −0.51 and 1.83, ) making it difficult to interpret.
Figure 6. Total, direct and indirect effects of grazing on Clamorous Reed Warbler occurrence probability from the path analysis (). For the graph, the Tamarix value was set to 5.5 since Clamorous Reed Warbler appear to be scarce at Tamarix values below 5. See text for effect sizes and uncertainties.
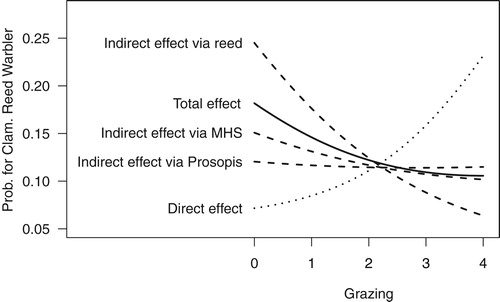
In the Clamorous Reed Warbler the expected negative total effect of grazing was not supported well by the data (estimate = −0.28, 95% intervals = −1.44 and 0.81, ). However, the data supported a negative indirect effect via reed cover (estimate = −0.64, 95% intervals = −1.50 and −0.08). The indirect effect of grazing on Clamorous Reed Warbler occurrence via mean shrub height was non-significant (estimate = −0.19, 95% intervals = −0.95 and 0.51), unlike in Dead Sea Sparrow. A similarly weak indirect effect via P. juliflora (estimate = −0.07, 95% intervals = −0.59 and 0.37) was observed for this species, too, while the indirect effect of grazing via Prosopis tended to be negative but (marginally) non-significant in the Dead Sea Sparrow (estimate = −0.33, 95% intervals = −1.04 and 0.09).
Discussion
This study provides strong evidence of indirect effects of grazing on two passerine species specialized on remnant marsh-like habitats in the Rift Valley. The indirect effects are reasonably interpretable, unlike the direct effects of grazing on the presence of the two specialist species. Significant indirect effects of heavy grazing include the reduction of the shrub sizes (mean heights) that lowers the probability of occurrence of Dead Sea Sparrow and the reduction of reed cover that is important for breeding Clamorous Reed Warbler. Tamarix density is a strong predictor of the breeding occurrence of both species in the Jordan Valley, but it could not be modelled in a path from grazing to Tamarix to bird occurrence due to the bidirectional effects as indicated by the U-shaped curve describing the relationship between grazing intensity and Tamarix density. This relationship can be explained according to our results and observations in the field. Intensive grazing apparently reduces shrub density with time, and high shrub density at accessible sites currently attracts herders, which is then expected to increase grazing intensity in the less degraded habitats. The practices of nomadic or semi-nomadic herders are thus relevant for conservation as some are leading to a circle of further habitat degradation due to the long-term effects of heavy grazing and other disturbances. Intensive grazing also affects the herbaceous vegetation, reed cover and small shrubs, very likely also Tamarix seedlings, thus suppressing the regeneration of native Tamarix thickets. Similar to our results, a high stocking rate of large herbivores has been found to negatively impact birds nesting in the shrub layer of riparian and semi-arid savanna habitats, whereas birds of open ground and low scrub are less affected (Krueper et al. Citation2003, James Citation2003, Ogada et al. Citation2008). Camels were the largest herbivores in our study area and were locally present in herds of up to 30 in the wadis during the rainy season when Tamarix is more palatable (local shepherds, pers. comm.). In addition to reducing the height of Tamarix canopies, they spread, along with other livestock, the seeds of invasive P. juliflora (Dufour-Dror & Shmida Citation2017). At low density, P. juliflora can be considered an indicator of ecological disturbance but with low impact. Depending on local conditions, however, it may dominate and contribute to considerable alterations in hydrology and vegetation (Dzikiti et al. Citation2017). Dead Sea Sparrows avoided areas with high P. juliflora densities because such sites were highly disturbed and rendered unsuitable for breeding, as indicated by a marginally significant negative direct effect of Prosopis. This confirms the findings of other studies (Schirmel et al. Citation2016) that demonstrated negative effects of invasive, alien plants on the diversity of birds, especially in riparian habitats, perhaps because food resources and suitable nesting sites are reduced. The encroachment of alien, invasive Prosopis along with the disappearance of breeding occurrences of specialist birds can thus be used as indicators of wetland habitat and ecosystem degradation in the Jordan Valley. This degradation is manifested by the decline of water flow and reduction of large Tamarix shrubs, reeds and other native vegetation, important for both bird species. Features of the vegetation (density, height, vertical layers) are important for birds in many ways: the dense vegetation contains food, including arthropods for the insectivorous Clamorous Reed Warbler. Reeds and high shrubs offer suitable structures for placing nests by both bird species, well concealed and/or well above the ground, thus protecting the nests from predation and flooding (Martin Citation1993, Lambert and Kleindorfer Citation2006, Khoury et al. Citation2009).
The large uncertainty in the direct and total effects of grazing on the occurrence of Clamorous Reed Warbler may be due to methodological restrictions in data analysis; the direct effect estimates bird occurrence for various grazing intensities but with fixed average reed and shrub height values. Situations with intensive grazing and an average reed or average shrub height were, however, rare in our study area because heavy grazing appears to have a strong negative impact on shrub size and reed cover. Secondly, we observed no Clamorous Reed Warblers in ten transects with low grazing scores although some contained reeds. Possible reasons for this are local temporal reductions in water flow and recent encroachment by drought-tolerant shrubs due to protection measures, such as at a managed ‘baptism site’ where some reed still occurs, but where Clamorous Reed Warbler stopped breeding several years ago (Khoury et al. Citation2006). This shows the need for further research on the specific habitat requirements of this species during the breeding season. Mérő et al. (Citation2016), for example, showed that Great Reed Warblers Acrocephalus arundinaceus respond to water availability and depth, with dominant males preferring more stable conditions and deeper water.
Our path analysis has its practical uses, as management could also target indirect effects. P. juliflora may invade the small wadis even if grazing is locally reduced because the droppings containing the seeds can be transported into the wadis by surface water from surrounding areas, and because livestock is frequently moved from one site to another. Controlling invasive Prosopis may thus be necessary to reduce or reverse negative changes within remnant, but important bird habitat patches. Overgrazing may slowly change an ecosystem so that new undesirable conditions are established (Reynolds et al. Citation2007). The drying out of some stream sections may be the result of cascading effects with positive feedback driven by local intensive grazing and consequent changes in soil properties and spread of invasive Prosopis (D’Odorico et al. Citation2013, Dzikiti et al. Citation2017). It is not our goal to disseminate our results as a condemnation of livestock grazing, given that marginal or moderate grazing intensity does not appear to have negative impacts on specialist birds. We rather point at the importance of controlled, sustainable grazing and stocking rates based on the true carrying capacity for livestock which is expected to be relatively low in an arid environment. Our results also suggest protecting specific nesting sites in remnant wetland patches from camel grazing to sustain local populations of Dead Sea Sparrow, Clamorous Reed Warbler and probably many other bird species. Land use in the surrounding landscape and further upstream also influences water flow in the tributaries of the River Jordan, and detailed knowledge of its cascading effects (including social and economic) at the landscape and regional scales (Kinzig et al. Citation2006), could be important for a more integrated management of key bird habitats. The two specialist bird species can, therefore, be used as indicators of the integrity or degradation of important wetland habitat patches.
Additional information
Funding
References
- Albert, R., Petutschnig, B. & Watzka, M. 2004. Zur vegetation und Flora Jordaniens. Denisia 14: 133–220.
- Andrews, I.J. 1995. The Birds of the Hashemite Kingdom of Jordan. I.J. Andrews, Musselburgh.
- Bibby, C.J., Burgess, N.D., Hill, D.A. & Mustoe, S. 2000. Bird Census Techniques, 2nd edn. Academic Press, New York.
- Clough, Y. 2012. A generalized approach to modeling and estimating indirect effects in ecology. Ecology 93: 1809–1815. doi: 10.1890/11-1899.1
- D’Odorico, P., Bhattachan, A., Davis, K.F., Ravi, S. & Runyan, C.W. 2013. Global desertification: drivers and feedbacks. Adv. Water Resour. 51: 326–344. doi: 10.1016/j.advwatres.2012.01.013
- Dufour-Dror, J.M. & Shmida, A. 2017. Invasion of alien Prosopis species in Israel, the West Bank and western Jordan: characteristics, distribution and control perspectives. BioInv. Rec. 6: 1–7. doi: 10.3391/bir.2017.6.1.01
- Dzikiti, S., Ntshidi, Z., Le Maitre, D.C., Bugan, R.D., Mazvimavi, D., Schachtschneider, K., Jovanovic, N. & Pienaar, H.H. 2017. Assessing water use by Prosopis invasions and Vachellia karroo trees: implications for groundwater recovery following alien plant removal in an arid catchment in South Africa. For. Ecol. Manage. 398: 153–163. doi: 10.1016/j.foreco.2017.05.009
- Feeley, K.J. & Terborgh, J.W. 2008. Direct versus indirect effects of habitat reduction on the loss of avian species from tropical forest fragments. Anim. Conserv. 11: 353–360. doi: 10.1111/j.1469-1795.2008.00182.x
- Hajzlerová, L. & Reif, J. 2014. Bird species richness and abundance in riparian vegetation invaded by exotic Reynoutria spp. Biologia 69: 247–253. doi: 10.2478/s11756-013-0296-x
- James, C.D. 2003. Response of vertebrates to fenceline contrasts in grazing intensity in semi-arid woodlands of eastern Australia. Austral. Ecol. 28: 137–151. doi: 10.1046/j.1442-9993.2003.01259.x
- Joubert, D.F. & Ryan, P.G. 1999. Differences in mammal and bird assemblages between commercial and communal rangelands in the Succulent Karoo, South Africa. J. Arid Environ. 43: 287–299. doi: 10.1006/jare.1999.0553
- Julliard, R., Clavel, J., Devictor, V., Jiguet, F. & Couvet, D. 2006. Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 9: 1237–1244. doi: 10.1111/j.1461-0248.2006.00977.x
- Khoury, F. & Korner, P. 2018. The effects of habitat variables and land use on breeding birds in remnant wetland strips in an arid, rural landscape. J. Arid Environ. 153: 24–31. doi: 10.1016/j.jaridenv.2018.01.001
- Khoury, F., Al-Omari, K., Azar, J. & Al-Hasani, I., 2006. Observations on the avifauna of the eastern Jordan Valley, during July-August 2005. Sandgrouse 28: 119–126.
- Khoury, F., Al-Shamlih, M., Sultan, H. & Abu-Ghalyun, Y. 2007. The effects of vegetation cover on the structure of bird communities in a hyperarid desert. Zool. Middle East 40: 11–20. doi: 10.1080/09397140.2007.10638199
- Khoury, F., Janaydeh, M. & Al-Hmoud, A.R. 2009. Nest placement and nesting success in two finch species colonizing a recently established plantation in an arid region. J. Ornithol. 150: 29–37. doi: 10.1007/s10336-008-0314-x
- Kinzig, A.P., Ryan, P., Etienne, M., Elmqvist, T., Allison, H.E. & Walker, B.H. 2006. Resilience and regime shifts: assessing cascading effects. Ecol. Soc. 11: 20. [online] URL: http://www.ecologyand-society.org/vol11/iss1/art20. doi: 10.5751/ES-01678-110120
- Korner, P., Graf, R. & Jenni, L. 2018. Large changes in the avifauna in an extant hotspot of farmland biodiversity in the Alps. Br. Conserv. Int. 28: 263–277. doi: 10.1017/S0959270916000502
- Krueper, D., Bart, J. & Rich, T. 2003. Response of vegetation and breeding birds to the removal of cattle on the San Pedro River, Arizona (USA). Conserv. Biol. 17: 607–615. doi: 10.1046/j.1523-1739.2003.01546.x
- Kumar, S. & Mathur, M. 2014. Impact of invasion by Prosopis juliflora on plant communities in arid grazing lands. Trop. Ecol. 55: 33–46.
- Lambert, S. & Kleindorfer, S. 2006. Nest concealment but not human visitation predicts predation of New Holland honeyeater nests. Emu 106: 63–68. doi: 10.1071/MU05006
- Ma, Z., Gan, X., Cai, Y., Chen, J. & Li, B. 2011. Effects of exotic Spartina alterniflora on the habitat patch associations of breeding saltmarsh birds at Chongming Dongtan in the Yangtze River estuary, China. Biol. Inv. 13: 1673–1686. doi: 10.1007/s10530-010-9924-3
- Martin, T.E. 1993. Nest predation and nest sites. BioScience 43: 523–532. doi: 10.2307/1311947
- Mérő, T.O., Žuljević, A., Varga, K. & Lengyel, S. 2016. Wing size-related reed habitat selection by Great Reed Warbler (Acrocephalus arundinaceus) males. Auk 133: 205–212. doi: 10.1642/AUK-15-175.1
- Millennium Ecosystem Assessment. 2005. Ecosystems and Human Well-Being: Desertification Synthesis. World Resources Institute, Washington, DC.
- Newton, S.F. & Newton, A.V. 1997. The effect of rainfall and habitat on abundance and diversity of birds in a fenced protected area in the central Saudi Arabian desert. J. Arid Environ. 35: 715–735. doi: 10.1006/jare.1996.0182
- Ogada, D.L., Gadd, M.E., Ostfeld, R.S., Young, T.P. & Keesing, F. 2008. Impacts of large herbivorous mammals on bird diversity and abundance in an African savanna. Oecologia 156: 387–397. doi: 10.1007/s00442-008-0994-1
- Pasiecznik, N.M., Felker, P., Harris, P.J.C., Harsh, L.N., Cruz, G., Tewari, J.C., Cadoret, K. & Maldonado, L.J. 2001. The Prosopis juliflora – Prosopis pallida Complex: A Monograph. HDRA, Coventry, UK, p. 172.
- Reynolds, J. F., et al. 2007. Global desertification: building a science for dryland development. Science 316: 847–851. doi: 10.1126/science.1131634
- Schirmel, J., Bundschuh, M., Entling, M.H., Kowarik, I. & Buchholz, S. 2016. Impacts of invasive plants on resident animals across ecosystems, taxa, and feeding types: a global assessment. Glob. Change Biol. 22: 594–603. doi: 10.1111/gcb.13093
- Shiferaw, H., Teketay, D., Nemomissa, S. & Assefa, F. 2004. Some biological characteristics that foster the invasion of Prosopis juliflora (Sw.) DC. at Middle Awash Rift Valley Area, north-eastern Ethiopia. J. Arid Environ. 58: 135–154. doi: 10.1016/j.jaridenv.2003.08.011
- Skórka, P., Lenda, M. & Tryjanowski, P. 2010. Invasive alien goldenrods negatively affect grassland bird communities in Eastern Europe. Biol. Conserv. 143: 856–861. doi: 10.1016/j.biocon.2009.12.030
- Snow, D. & Perrins, C. 1997. The Birds of the Western Palearctic, Concise Edition. Vol. 2: Passerines. Oxford University Press, Oxford.
- Tomoff, C.S. 1974. Avian species diversity in desert scrub. Ecology 55: 396–403. doi: 10.2307/1935227
- Waitzbauer, W. & Petutsching, B. 2004a. Zur Hydrogeologie Jordaniens. Denisia 14: 55–76.
- Waitzbauer, W. & Petutsching, B. 2004b. Zum Klima Jordaniens. Denisia 14: 77–87.
- Wilcox, B.P. 2007. Does rangeland degradation have implications for global streamflow? Hydrol. Process. 21: 2961–2964. doi: 10.1002/hyp.6856
- Whitford, W.G. 1997. Desertification and animal biodiversity in the desert grasslands of North America. J. Arid Environ. 37: 709–720. doi: 10.1006/jare.1997.0313