
ABSTRACT
The local and migratory movements of Greylag Geese Anser anser wintering in southwest and northwest Europe were revealed with global positioning system tracking. Geese in the southwest had smaller home ranges than those from the northwest, and birds also moved large distances over wintering grounds and showed flexibility in the annual choice of wintering sites.
Within Europe, the Greylag Goose Anser anser has four biogeographic populations: Icelandic, British and Irish, which is resident, central European and northwest/southwest European, where the latter population accounts for approximately three quarters of the total number of birds (Fox et al. Citation2010). The strong increase of the northwest/southwest European population over recent decades has raised new issues for the conservation and management of this population due to the recent conflicts with human activities (Powolny et al. Citation2018). In this context, understanding the migration strategy of Greylag Geese and how they use their stopover and wintering areas in space and time is a major issue for population and habitat management (Powolny et al. Citation2018). Greylag Geese breeding in Norway are of special interest because these individuals become more abundant among Greylags migrating over France and reaching Spain relative to the birds originating from other Scandinavian countries or from the Netherlands (Pistorius et al. Citation2006a, Citation2007, Voslamber et al. Citation2010, Nilsson et al. Citation2013). The aim of the present study was to investigate the local and migratory movements of Greylag Geese depending on their breeding sites in north and central-western Norway (East-Finnmark, Helgeland, Møre and Romsdal) and wintering regions (southwest vs. northwest Europe) and to test the hypothesis that birds wintering farther from their breeding grounds tend to arrive later on their nesting sites than those wintering closer (Alves et al. Citation2013).
We analysed data from 33 individuals (birds were not sexed), older than 1 year, equipped with global positioning system (GPS) and global systems for mobile communications (GSM) solar powered transmitters (less than 3% of the bird’s body mass, Ecotone Telemetry, Poland) set to 12 fixes per day and for which we recorded regular geolocations for more than two months. Attachment methods followed the previous recommendations (Demers et al. Citation2003, Barron et al. Citation2010). Fourteen birds were trapped with canon-nets in southern Spain (December 2011 and 2013) at the Doñana National Park (n = 5) and at the Los Canchales reservoir (n = 9); 19 were trapped with sweep-nets at sea during the moulting period (July 2012–14) at the central-western (n = 12) and northeast (n = 7) coasts of Norway.
Birds’ home ranges were calculated by means of 50% and 95% kernel utilization distribution (KUD) (Worton Citation1989) using ArcMap 10.2 with Minimum Boundary Tools (ArcGIS 10.2, Esri 2013). Fifty per cent KUD provides an estimate of an animal’s core activity area, while 95% KUD also includes exploratory behaviours (Chevallier et al. Citation2010). To differentiate migratory geese and intermediate flights behaviour from their sedentary activities we used the distance of 30 km between two consecutive GPS fixes as it is known to be the maximum daily distance a goose covers between roosting and foraging sites (Van Wijk et al. Citation2012). KUDs were calculated for five different periods of the annual cycle: (i) stopovers during autumn migration, (ii) wintering – period from the end of autumn migration up to 10 days before the beginning of spring migration, (iii) spring premigration – a 10-day period before spring migratory flights, (iv) spring migration stopovers and (v) nesting.
To differentiate between intermediate and migratory flights we used previously established criteria (Ebbinge & Buurma Citation2000, Van Wijk et al. Citation2012) and information on the long distance round trips from this study. Successive flights over at least 250 km in one direction with no backward displacements exceeding 30 km were considered as migratory, and the flights within the 30 to 250 km range were considered as intermediate. To examine the effect of wintering regions on the number of intermediate or migratory flights we used generalized linear mixed models with Poisson distribution. To test how home ranges differed according to the wintering regions and annual cycle periods we used linear mixed models on the log-transformed data. In both types of models the diagnostic plots of the residuals were checked to make sure that normality assumptions were satisfied. Likelihood-ratio test (LRT) was used to test for the significance of the interaction term in both types of models. We used Tukey correction to account for the multiple comparisons when necessary. The analyses were performed using R software (R Core Team Citation2016).
Among Greylag Geese tagged on the breeding sites in Norway, 68% of the individuals wintered in northwest Europe (Belgium, Netherlands or northwest Germany), and 32% migrated to southwest Europe (France or Spain). This is in accordance with the earlier observed northerly shift of the wintering core of the northwest/southwest European population with more than 50% of the population now wintering in northern Europe (Ramo et al. Citation2015, Powolny et al. Citation2018). Interestingly, one individual tracked over four consecutive years wintered alternatively in the Netherlands and in the southern part of the Iberian Peninsula. These observations raise important questions regarding the flexibility of the annual choices of the wintering sites and should be explored further.
On average, duration of migration journeys over France was longer during spring migration (median = 5 days) than during autumn migration (median = 1 day; Mann Whitney U = 8, P = 0.004). This shows that for southwest geese, migratory stopovers in France were relatively short. Overall, mean (±se) migration speed was 69 ± 2 km h−1 (range = 45–130 km h−1). Regardless of their wintering location and migratory routes, Greylag Geese returned the following spring to their initial breeding grounds within less than a 500 m radius of their capture location. This provides additional evidence for the high breeding site fidelity or philopatry that has been previously documented (Andersson et al. Citation2001, Kruckenberg & Borbach-Jaene Citation2004). Whether the site fidelity is characteristic for all sub-populations of the flyway, whether it persists across the years, and how it depends on bird’s social status need to be further investigated over the long-term period. Noteworthy, the high breeding site fidelity does not seem to preclude genetic heterogeneity and admixture among the northwest/southwest European population (Pellegrino et al. Citation2015).
Interestingly, Greylag Geese from the Eastern Finnmark region migrated via the Swedish coast (Bothnia Gulf). Greylag Geese stayed for one to four weeks at the same stopover site along the Swedish coast (Umeå region) or in the southern region of Sweden (Västmanland county) before moving to Denmark and the Netherlands after mid-October. The spring migratory routes back to their breeding areas and stopover locations were similar.
The number of intermediate flights taken by birds depended on their wintering zone and the month of the year (LRT: χ2 = 27.3, df = 7, P < 0.001, N = 235 flights; (A)). Specifically, in September, birds that two months later would migrate and winter in southwest Europe took more flights than birds that would winter in northwest Europe (LSMeans post-hoc comparison test, means ± se: southwest = 2.15 ± 0.76, northwest = 0.45 ± 0.15, z = −3.282, P = 0.001). The number of intermediate flights taken by southwest and northwest populations during the other months was not significantly different (P > 0.052).
Figure 1. (A) Number of intermediate flights per individual (mean ± se) performed per month by Greylag Geese wintering in southwest and northwest Europe. (B) Chronology (mean date ± se) of spring migration stages for geese wintering in southwest Europe and breeding in Norway (precise mean dates are indicated besides the bars).
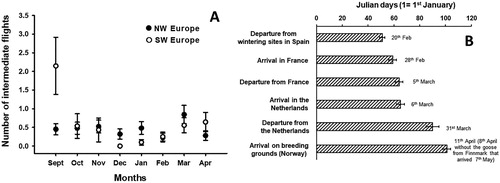
During wintering, in late December and January, especially in northwest Europe, 25% of the geese performed relatively large displacements northeast (45–250 km between two consecutive geolocations). They staged for several days outside their previous wintering core area and then came back southwest one to three weeks later. These intermediate movements (mean ± se = 87.3 ± 15.6 km), different from local and migratory flights, showed that some individuals or flocks redistributed over a large range within their wintering area, perhaps, to find alternative food sources.
The mean (±se) number of migratory flights per individual changed over months ranging from 0.11 ± 0.05 in December to 1.05 ± 0.19 in March (LRT: χ2 = 47.6, df = 6, P < 0.001). Greylag Geese wintering in southwest Europe performed more migratory flights overall than birds in northwest Europe (means ± se; southwest = 0.82 ± 0.12, northwest = 0.55 ± 0.08, LRT: χ2 = 5.0, df = 1, P < 0.05). Irrespective of the wintering grounds, no migratory flights were observed in January.
Over all years of tracking, Greylag Geese wintering in southwest Europe started their spring migration between the 11th February and 10th March ((B)). For geese that wintered in Spain and France, the Netherlands was the major spring migration stopover (median = 31 days). Possibly, geese breeding in Norway start their spring migration from southwest Europe later than those breeding in southern Sweden (Nilsson et al. Citation2013). This hypothesis should be tested with a larger dataset using GPS tagged Greylag Geese in Norway and Sweden, and migrating over long distances.
Based on Greylag Geese (n = 13) that joined the same breeding areas in central-western Norway (excluding the birds breeding in Finnmark), spring migration departure dates from northwest Europe (used as stopover areas) and arrival dates on the breeding grounds were respectively 7 days later and 8 days earlier for geese that wintered in southwest Europe compared to those that wintered in northwest Europe. However, these differences between southwest and northwest birds were not significant (Mann Withney U = 11, P = 0.22). Thus, our results do not support the idea that arrival dates on similar breeding grounds depend on migration distances. Potentially, as for other migratory species, the migration costs could be compensated during the four-week stopovers in the Netherlands (see also Alves et al. Citation2013). The median arrival date on breeding grounds (8 April) regardless of the wintering regions is consistent with previous reports (Pistorius et al. Citation2006b).
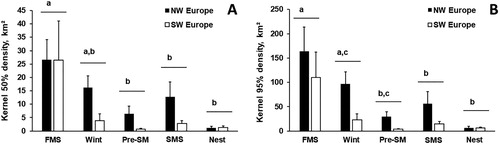
Mean (±se) local movements recorded between two GPS fixes averaged 1.18 ± 0.01 km. They happened mostly in the second part of the night (29%: 00–06 h GMT) and the day (29%: 12–18 h GMT). The birds that overwintered in southwest Europe had smaller exploratory and core activities home ranges than birds in northwest Europe, independently of the periods within their annual cycle (50% KUD: LRT, χ2 = 10.8, df = 1, P = 0.001, n = 254; 95% KUD: LRT, χ2 = 10.7, df = 1, P = 0.001, n = 254; ). A possible explanation is that the increasing number of birds staging in northwest Europe force geese to explore larger areas to forage in order to avoid food competition (Legagneux et al. Citation2009). Both exploratory and core activities home ranges changed by periods (50% KUD: LRT, χ2 = 26.6, df = 4, P < 0.001, n = 254; 95% KUD: LRT, χ2 = 31.2, df = 4, P < 0.001, n = 254; ). The smallest home ranges were observed during nesting while the biggest were observed during autumn migration stopovers, which consequently decreased during wintering. This decrease in home ranges during wintering was also recorded in Greylag Geese tracked in Germany (Kleinhenz & Koenig Citation2018).
Figure 2. (A) Home range areas (95% KUD density, km2, mean ± se) and (B) core areas (Kernel 50% density, km2, mean ± se) according of the wintering region of Greylag Geese and at different stages of the annual cycle periods (FMS – autumn migration stopovers, Wint – wintering, Pre-SM – spring premigration, SMS – spring migration stopover, Nest – nesting). The period × wintering region interaction was not significant (LRT: χ2 < 5,53, df = 4, P > 0.24, n = 254). Mean values (for merged sites) sharing the same letters were not significantly different (P > 0.05).
To conclude, our study using continuous GPS tracking of Greylag Geese provides new and valuable insights into understanding individual migration strategies and spatial ecology of the northwest/southwest population of Greylag Geese, in addition to the visual surveys of neck-banded geese. The geolocational data can help better to define management units and their functionalities in order to achieve goals set in the international management plan adopted for this species at the 7th Session of the Meeting of the Parties to the African-Eurasian Migratory Waterbird Agreement (Powolny et al. Citation2018). In addition, tracking will facilitate the investigation of wintering strategies of Greylag Geese in the context of climate change, population size changes and carrying capacities of natural and agricultural habitats.
Acknowledgements
We thank for the purchase of some GPS devices the French National Hunting Federation (FNC), the North-East Scientific Atlantic Institute (ISNEA) and local associations: Fédérations Départementales des Chasseurs de l'Oise, du Nord et de Charente-Maritime, ANCGE, GASSAUGI (33), ASSELMM (33), ALCGE (40), ASSDH (59), ACGECE (59), ASVS (59), GDCGEN (59), ACMCOC (50), UPACGEM (80,60), ACTCM (17), ASCGE (17). We are also grateful to the Office National de la Chasse et de la Faune Sauvage, to José Masero and José Mª Abad Gómez-Pantoja (University of Badajoz) and to Andy Green and Juan Amat (CSIC Sevilla) for their help with the field work in Spain. Procedures for capture, handling and marking were reviewed and approved by the Spanish and Norwegian Animal Care and Use Committees.
Additional information
Funding
References
- Alves, J.A., Gunnarsson, T.G., Hayhow, D.B., Appleton, G.F., Potts, P.M., Sutherland, W.J. & Gill, J.A. 2013. Costs, benefits, and fitness consequences of different migratory strategies. Ecology 94: 11–17. doi: 10.1890/12-0737.1
- Andersson, Å., Follestad, A., Nilsson, L. & Persson, H. 2001. Migration pattern of Nordic Greylag Geese Anser anser. Ornis Svec. 11: 19–58.
- Barron, D.G., Brawn, J.D. & Weatherhead, P.J. 2010. Meta-analysis of transmitter effects on avian behaviour and ecology. Methods Ecol. Evol. 1: 180–187. doi: 10.1111/j.2041-210X.2010.00013.x
- Chevallier, D., Le Maho, Y., Baillon, B., Duponnois, R., Dieulin, C., Brossault, P., De Franclieu, P., Lorge, P., Aurouet, A. & Massemin, S. 2010. Human activity and the drying up of rivers determine abundance and spatial distribution of Black Storks Ciconia nigra on their wintering grounds. Bird Study 57: 369–380. doi: 10.1080/00063651003678467
- Demers, F., Giroux, J. F., Gauthier, G. & Bêty, J. 2003. Effects of collar-attached transmitters on behaviour, pair bond and breeding success of snow geese Anser caerulescens atlanticus. Wildlife Biol. 9: 161–170. doi: 10.2981/wlb.2003.047
- Ebbinge, B. S. & Buurma, L.S. 2000. Mid winter movements of geese in the Netherlands as a risk to aviation safety. International Bird Strike Committee, Amsterdam, 17–21 April 2000, 9 p.
- Fox, A.D., Ebbinge, B.S., Mitchell, C., Heinicke, T., Aarvak, T., Colhoun, K., Clausen, P., Dereliev, S., Faragó, S., Koffijberg, K., Kruckenberg, H., Loonen, M.J.J.E., Madsen, J., Mooij, J., Musil, P., Nilsson, L., Pihl, S. & Van Der Jeugd, H. 2010. Current estimates of goose population sizes in Western Europe, a gap analysis and assessment of trends. Ornis Svec. 20: 115–127. doi: 10.34080/os.v20.19922
- Kleinhenz, A. & Koenig, A. 2018. Home ranges and movements of resident graylag geese (Anser anser) in breeding and winter habitats in Bavaria, South Germany. Plos One 13: e0202443. doi: 10.1371/journal.pone.0202443
- Kruckenberg, H. & Borbach-Jaene, J. 2004. Do Greylag geese Anser anser use traditional roosts? Site fidelity of colour-marked Nordic Greylag geese during spring migration. J. Ornithol. 145: 117–122. doi: 10.1007/s10336-004-0021-1
- Legagneux, P., Blaize, C., Latraube, F., Gautier, J. & Bretagnolle, V. 2009. Variation in home-range size and movements of wintering dabbling ducks. J. Ornithol. 150: 183–193. doi: 10.1007/s10336-008-0333-7
- Nilsson, L., Follestad, A., Guillemain, M., Schricke, V. & Voslamber, B. 2013. France as a staging and wintering area for Greylag geese Anser anser. Wildfowl 63: 24–39.
- Pellegrino, I., Cucco, M., Follestad, A. & Boos, M. 2015. Lack of genetic structure in Greylag goose Anser anser populations along the European Atlantic flyway. PeerJ. 3: e1161. doi: 10.7717/peerj.1161
- Pistorius, P.A., Follestad, A. & Taylor, F.E. 2006a. Declining winter survival and fitness implications associated with latitudinal distribution in Norvegian Greylag Geese Anser anser. Ibis 148: 114–125. doi: 10.1111/j.1474-919X.2006.00498.x
- Pistorius, P.A., Follestad, A. & Taylor, F.E. 2006b. Temporal changes in spring migration phenology in the Norwegian Greylag goose Anser anser, 1971–2004. Wildfowl 56: 23–36.
- Pistorius, P.A., Follestad, A., Nilsson, L. & Taylor, F.E. 2007. A demographic comparison of two Nordic populations of Greylag Geese Anser anser. Ibis 149: 553–563. doi: 10.1111/j.1474-919X.2007.00682.x
- Powolny, T., Jensen, G.H., Nagy, S., Czajkowski, A., Fox, A.D., Lewis, M. & Madsen, J. 2018. AEWA International Single Species Management Plan for the Greylag Goose (Anser anser) - Northwest/Southwest European population. AEWA Technical Series No. xx, Bonn, Germany.
- R Core Team. 2016. R: A Language and Environment for Statistical Computing. Vienna: R.Foundation for Statistical Computing. URLhttps://www.R-project.org/.
- Ramo, C., Amat, J.A., Nilsson, L., Schricke, V., Rodríguez-Alonso, M., Gómez-Crespo, E. & Green, A.J. 2015. Latitudinal-related variation in wintering population trends of Greylag Geese (Anser Anser) along the Atlantic flyway: a response to climate change? Plos One, 10: e0140181. doi: 10.1371/journal.pone.0140181
- Van Wijk, R.E., Kölzsch, A., Kruckenberg, H., Ebbinge, B.S., Müskens, G.J. & Nolet, B.A. 2012. Individually tracked geese follow peaks of temperature acceleration during spring migration. Oikos 121: 655–664. doi: 10.1111/j.1600-0706.2011.20083.x
- Voslamber, B., Knecht, E. & Kleijn, D. 2010. Dutch Greylag geese Anser anser: migrants or residents? Ornis Svec. 20: 207–214.
- Worton, B.J. 1989. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70: 164–168. doi: 10.2307/1938423