
ABSTRACT
Capsule: The nesting habitat of the Common Buzzard Buteo buteo and Lesser Spotted Eagle Clanga pomarina extensively overlap, indicating that they exploit similar resources.
Aim: We aimed to determine the overlap in the nest platforms, nest trees and nest stands used by these raptors, find any evidence for the avoidance of the larger Lesser Spotted Eagle by the smaller Common Buzzard, and provide conservation implications for habitat protection of the former species in habitats that overlap extensively.
Methods: Nest sites were mapped during 2012–2014 in the Biržai Forest Spatial Protection Area, northern Lithuania. Fifty-three nest sites occupied by Common Buzzards and 26 by Lesser Spotted Eagles were compared.
Results: The nest platforms of both raptors were similarly placed in the tree canopies. Most Lesser Spotted Eagle nests were built in spruce, while the Common Buzzard usually nested in birch. The nest stands of the eagles were on wetter soil and located closer to the forest edge than those of the buzzards, otherwise, the nest stands did not differ significantly. There was no evidence for spatial avoidance of the larger raptor by the Common Buzzard.
Conclusions: The different components of the nesting habitats extensively overlapped, and the distribution of the interspecific pairs lacked spatial avoidance. We suggest that the nest sites of both raptors were a largely shared resource, especially if located close to the forest edges. We propose, as a rule of thumb, applying protection by way of buffer zones around buzzard nest sites if they are located close to eagle nest sites and the forest edge.
The availability of prime nesting sites suitable for forest-dwelling raptors is usually restricted by proximity to optimal foraging habitats (Lõhmus Citation2001), avoidance of anthropogenic disturbance (Ortego Citation2007), preference for mature stands (Treinys & Mozgeris Citation2010 and references therein), and the presence of some key physical structures (e.g. large, malformed trees within otherwise suitable stands) (Lõhmus & Sellis Citation2003). Timber harvesting is a crucial driver in the forest ecosystem, changing the availability of mature stands that would otherwise be suitable for a nesting raptor (Barrientos & Arroyo Citation2014). As most long-lived raptors occupy the same territories for many years (Poole Citation1989, Meyburg et al. Citation2004), harvesting restrictions in buffer zones of a certain size around nest sites occupied by focal species are a useful tool for protecting nesting habitats, as well as the birds themselves during periods of extreme sensitivity (Richardson & Miller Citation1997). The protection of nest sites (whether occupied or not) in the buffer zones is also vital for the persistence of local raptor populations under timber harvesting pressures (Jiménez-Franco et al. Citation2018).
Limited resources, together with overlap in resource use, are the basis for interspecific competition between sympatric species (Dhondt Citation2012). Previous studies have demonstrated extensive overlap in nest sites among sympatric and similar-sized diurnal raptors occupying stick nests (Krüger Citation2002, Katzner et al. Citation2003, Hakkarainen et al. Citation2004). In habitat overlap and resource shortage situations, one species may occupy the nests of other species (Skuja & Budrys Citation1999, Hakkarainen et al. Citation2004, Jiménez-Franco et al. Citation2014). Understanding nesting habitat differences, and the potential for biotic interactions between sympatric species when at least one of them is protected, is also important from a conservation perspective to avoid mismanagement (Sarà et al. Citation2016). If a focal raptor species shares a similar habitat with an abundant, but unprotected, species, management decisions to protect the nesting habitat with buffer zones are not straightforward because decision-makers face a trade-off situation between increasing the area of protected nesting/potential habitat of the focal species and increasing income losses for forestry entities due to the restriction of timber harvesting.
While there are existing studies on nesting habitat differences among various forest-dwelling raptor species (Selås Citation1997, Krüger Citation2002, Katzner et al. Citation2003, Hakkarainen et al. Citation2004, Poirazidis et al. Citation2007, Treinys et al. Citation2011, Kajtoch et al. Citation2015), studies focusing on habitat segregation between two ecologically similar diurnal birds of prey – the Lesser Spotted Eagle Clanga pomarina (1.1–2.2 kg) and the Common Buzzard Buteo buteo (0.6–1.2 kg) – are scarce (but see Lõhmus Citation2006). These two generalist predators are abundantly distributed in the forests of Central and Eastern Europe (report under Article 12 of the Birds Directive Period 2008–2012, https://bd.eionet.europa.eu/article12/ summary). The mean densities in suitable habitats vary from 14 to 21 pairs per 100 km2 for the Common Buzzard (Dombrovski & Ivanovski Citation2005) and from 3 to 26 pairs per 100 km2 for the Lesser Spotted Eagle (Bergmanis et al. Citation2015). Both species prefer mature forest stands and trees for nesting (Skuja & Budrys Citation1999, Drobelis Citation2004, Bielański Citation2006, Treinys & Mozgeris Citation2006) and, despite body size differences, their stick nest structures are very similar in their dimensions (Drobelis Citation2004). In fact, there have been observed cases where Common Buzzards have successfully reproduced in the nests of the larger, site-tenacious Lesser Spotted Eagle (Skuja & Budrys Citation1999, SS, RT unpublished data). These observations have led to the expectation of extensive nesting habitat overlap between these two raptors.
The Lesser Spotted Eagle, but not the Common Buzzard, is under the protection of an European Union Bird Directive, and consequently, member states are obliged to designate Special Protection Areas (SPAs) for their conservation. In Lithuania, following its accession to the European Union in 2004, new areas were designated as SPAs, including those where the focal species is the Lesser Spotted Eagle. This eagle species is threatened by forestry operations in their breeding grounds (Meyburg et al. Citation2001); hence, Lesser Spotted Eagle nest sites are legally protected with buffer zones within SPAs (as well as outside the Natura 2000 network). However, the nest sites of the Common Buzzard are not protected by the same approach (at least in Lithuania, according to the Forest Felling Rules approved by the Ministry of the Environment). The key question we aimed to answer here is, should the nest sites of the unprotected Common Buzzard also be considered when protecting the Lesser Spotted Eagle in a Natura 2000 network, if the nesting habitats of these raptors significantly overlaps?
We used data on Common Buzzard and Lesser Spotted Eagle pairs that bred in a large commercial forest that was designated as an SPA for the conservation of the eagles, in order to understand the extent of habitat overlap between the two species. More specifically, we compared: (1) nest platforms, (2) nest trees and (3) nest stands used by these two raptors and analysed (4) whether the smaller Common Buzzard spatially avoided the larger Lesser Spotted Eagle when occupying nest sites. Based on the findings, we also discuss the implications for their conservation.
Methods
Study area and data collection
Data on nest sites were collected in the Biržai Forest, northern Lithuania. The study area (approximately 170 km2; 56°17′N, 24°56′E) was characterized by flat, fertile, wet or water-logged terrain covered by forest (). The stands were dominated by Silver Birch Betula pendula (31%), then Norway Spruce Picea abies (29%) and Black Alder Alnus glutinosa (13%). The study area was covered by a dense network of gravel roads and drainage channels (total lengths 450 and 700 km, respectively). The stands in the study area were used for commercial forestry, but with some restrictions related to the protection of rare species.
Figure 1. Location of Biržai Forest in the region, and distribution of Common Buzzard and Lesser Spotted Eagle nests.
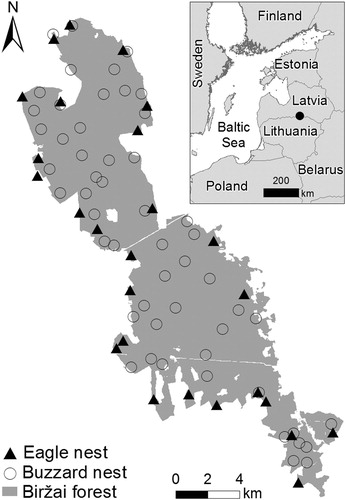
The nest sites occupied by raptors were mapped during fieldwork undertaken between 2012 and 2014. The Lesser Spotted Eagle was searched for between April and August, by observing their behaviour in open areas, mapping potential nest sites (according to the method described by Dombrovski & Ivanovski Citation2005) and, later, checking these forest patches for nests. The Common Buzzard was searched for during April to June, by checking nests found in October–March searches, and by observing bird behaviour. A nest platform was considered to be occupied by one of the two species if it had been attended to by the birds in the spring or early summer, or if it contained eggs, nestlings or their remains, following the common techniques used in raptor research (Steenhof & Newton Citation2007). Only the most recently occupied nest in the distinct territory of a pair was included in the analysis where the pair used two nests between 2012 and 2014. Altogether, 53 nest sites occupied by the Common Buzzard and 26 by the Lesser Spotted Eagle were included in the study.
Data analyses
For each nest site, characteristics were measured representing different components of the habitat (Gamauf et al. Citation2013): (1) nest platform, (2) nest tree and (3) nest stand (the variables used are presented in ). All the spatial data were processed using ArcGIS 10.0 software.
Table 1. Explanatory variables used to characterize Common Buzzard and Lesser Spotted Eagle nest sites.
Nest platform analysis
The proportions of the nest platforms placed by the raptors in trunk forks, side branches and top branches were compared using a chi-squared test. The difference in height of the nest platform from the ground between the raptors was analysed through a generalized linear model (GLM; binomial error structure, logit link function). The binary response variable was species (0 – Common Buzzard, and 1 – Lesser Spotted Eagle), and the height of the nest platform from the ground was included as the explanatory variable.
Nest tree analysis
The proportions of tree species used for nesting by the Common Buzzard and Lesser Spotted Eagle were compared using the chi-squared test. The differences in the trunk diameters of the nest trees were analysed through a generalized linear mixed model (GLMM; binomial error structure, logit link function). In this analysis, we included Norway Spruce, Black Alder, Aspen Populus tremula and Silver Birch nest trees because both raptors exploited these for nesting. The binary response variable was raptor species, the nest tree trunk diameter was included as the explanatory variable and tree species was included as the random variable.
Nest stand analysis
We built GLMs that included the same species binary response and explanatory variables for all possible combinations (i.e. stand age, fertility, humidity, proportion of tree species, distance to forest edge) using the dredge function (MuMIn package; Bartoń Citation2019). To better understand the underlying mechanism for the differences between raptor nest stands, we tested each species’ preferences by comparing the nest stands with randomly selected stands available in the studied forest (hereafter termed ‘control stands’). This approach is widely used to test habitat preferences (Ortego Citation2007, Treinys & Mozgeris Citation2010). The minimal 0.5 km distance between stands was the only restriction applied when selecting control stands. The nest stand variables () were extracted for 53 control stands. The nest stand preferences of the Common Buzzard and Lesser Spotted Eagle were analysed by comparing each to the control stands using GLMs. We built GLMs that included a binary response (0 – control forest stand, 1– Common Buzzard or Lesser Spotted Eagle nest site) and explanatory variables for all possible combinations, as listed above.
Competition analysis
The conspecific distribution patterns (i.e. random, regular or aggregated) for the Common Buzzard and Lesser Spotted Eagle in the study area were estimated using the average nearest neighbour tool in the ArcGIS 10.0 software. To test whether the Common Buzzard avoided interspecific competitor, we built a GLM that included the binary response variable (0 – control stand, 1 – Common Buzzard) and distance to the nearest Lesser Spotted Eagle nest as an explanatory variable.
An information-theoretical approach was used for model selection and a multi-model inference procedure (Burnham & Anderson Citation2002). The Akaike information criterion with a correction for small sample size (AICc), was used to select the best models from the corresponding model sets. The relative importance of each model was estimated through ranking the models by ΔAICc = AICci − AICcmin, where AICcmin was the best model in the corresponding model set. To separate good models from less supported models, the threshold ΔAICc ≤ 2 was used. When several models in the best model subsets were similar in model weight, we used an averaging procedure to estimate the relative importance values for the explanatory variables. The packages lme4 (Bates et al. Citation2015) and MuMIn (Bartoń Citation2019) in the statistical environment R were used for these calculations (R v.3.5.1 R Core Team Citation2018).
Results
The descriptive characteristics of the nest sites of the two species are presented in .
Table 2. Descriptive statistics for the nest sites of the Lesser Spotted Eagle and the Common Buzzard in the Biržai Forest Special Protection Area.
Nest platform differences between raptor species
The largest proportion (over 3/4) of the nest platforms occupied by the two bird species was placed in trunk forks, whilst the other nests were placed on the side and the top branches in similar proportions (χ2 = 1.1, df = 2, P < 0.6; ). The model showing the differences between species in height of the nest platform from the ground (ΔAICc = 1.33; weight = 0.34) was 1.9 times less likely compared to the null model (ΔAICc = 0.00; weight = 0.66), indicating that the nest platforms of both raptors were located at similar heights from the ground.
Nest tree differences between raptor species
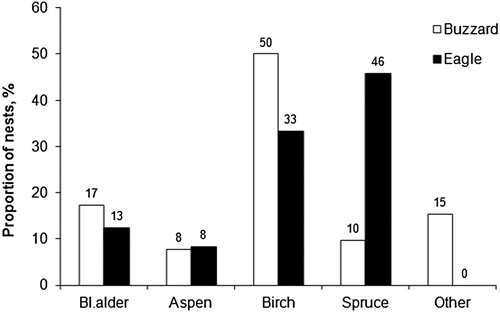
The Common Buzzard differed in tree species usage for nesting compared to the Lesser Spotted Eagle (χ2 = 38.2, df = 4, P < 0.0001; ). The largest proportion of nests occupied by the Common Buzzard was in Silver Birch, whereas Norway Spruce was most frequently used by the Lesser Spotted Eagle. The model showing the nest tree diameter differences between the species (ΔAICc = 1.53, weight = 0.32) was 2.1 times less likely compared to the null model (ΔAICc = 0.00, weight = 0.68), indicating that the raptors nested in trees of similar size.
Figure 2. Proportion of nest tree species used by the Common Buzzard (white bars) and the Lesser Spotted Eagle (black bars).
Nest stand differences between raptor species
The nest stands occupied by Lesser Spotted Eagles differed from those occupied by Common Buzzards, as indicated by the nine models supported by the data (). After model-averaging, the relative importance values (RIV) for the explanatory variables, included in the supported model subsets, indicated that most of the raptor species differences between the nest stands were in distance to forest edge and humidity (RIVs of 1.00 and 0.93, respectively). The nest stands of the Lesser Spotted Eagle were located closer to the forest edge () and on wetter soil than those of the Common Buzzard. Occupancy probability was only moderately explained by these two variables included in the best model (34% coefficient of determination). The nest stands were similar in tree species composition, with only the proportion of Silver Birch tending to differ. In summary, the results supported extensive overlap in the nest stands used by the two raptor species.
Figure 3. Distance to forest edge from the nest stands of the Common Buzzard (n = 53) and the Lesser Spotted Eagle (n = 26). The hollow square shows the median, the grey box gives the 25–75% range and the lines indicate the non-outlier range.
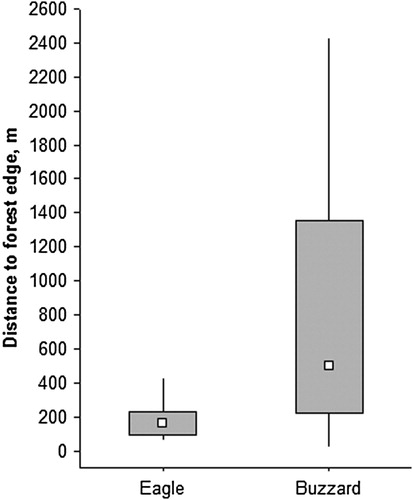
Table 3. Summary of supported models (GLMs) used to compare nest stands between species and with control stands. W – AICc model weights calculated for models ΔAICc ≤ 2. Negative estimates - decrease in values of explanatory variables in Lesser Spotted Eagle nest stands compared with the Common Buzzard nest stand, in the nest stand of the Common Buzzard compared to the control stands, and in the nest stand of the Lesser Spotted Eagle compared to the control stands. RIV – relative importance values estimated for variables after supported models averaging. Explanatory variables used: Fr.edge – distance to forest edge, St.age – stand age, Humid. – humidity of stand soil, Fertil. – fertility of stand soil, Bl.alder – proportion of Black Alder in stand composition, Aspen, Silver Birch, Norway Spruce – proportion of these tree species in stand composition.
Nest stand differences between the Common Buzzard and control stands
The nest stands of the Common Buzzard differed markedly compared to the control stands, as revealed by the subset of best models supported by the data (). Model-averaging indicated that the Common Buzzard nested in older stands with a higher proportion of Black Alder and less wet soils, compared to the control stands. The best-ranked model including these three variables explained a moderate proportion of response variable variation (31% coefficient of determination).
Nest stand differences between the Lesser Spotted Eagle and control stands
The Lesser Spotted Eagle nest sites differed from the control stands, as indicated by the best model’s subset (). Model-averaging indicated that the most important two variables were distance to forest edge and stand age, followed by the proportion of Aspen and Silver Birch in the stand composition. The nest stands occupied by Lesser Spotted Eagles were closer to the forest edge, older, with more Aspen and less Silver Birch in the stand composition, compared to the control stands. The best-ranked model including three variables explained a moderate proportion of the response variable variation (42% coefficient of determination).
Distribution of nest sites
The mean (± sd) distance between the nearest nest sites of the Common Buzzard was 1.28 ± 0.52 km (n = 53), while the minimum distance was only 0.5 km. Pairs of Common Buzzards in the study area were distributed randomly (average nearest neighbour analysis: z-score = 0.97, P = 0.33). The mean (± sd) conspecific inter-nest distance for the Lesser Spotted Eagle was 2.01 ± 0.97 km (n = 26), but the two closest pairs nested only 0.65 km apart. Analysis of the Lesser Spotted Eagle distribution indicated that conspecific pairs tended to be regularly dispersed (z-score = 1.77, P = 0.08). The mean (± sd) distance from Common Buzzard nests to the nearest Lesser Spotted Eagle nest was 1.51 ± 0.97 km (n = 53), however, 15% of Common Buzzard pairs nested within 0.51 km of a Lesser Spotted Eagle pair. The modelling results indicated that breeding Common Buzzards did not avoid proximity to Lesser Spotted Eagle nests because the model with that explanatory variable (ΔAICc = 1.72, weight = 0.3) was 2.4 times weaker than the null model (ΔAICc = 0.00, weight = 0.7), based on the evidence ratio.
Discussion
A comparison of the nest sites occupied by the sympatric Common Buzzard and Lesser Spotted Eagle in the large Biržai Forest SPA has revealed a strong overlap in the nesting habitat at three levels of spatial scale. The nest platforms of these raptors were similarly placed in the tree canopies. Most Lesser Spotted Eagle nests were built in Norway Spruce, while the Common Buzzard usually nested in Silver Birch, with other tree species being used at a similar frequency and being of similar size. The nest stands of the eagles were on wetter soil and located closer to the forest edge than those of the buzzards, but otherwise their nest stands did not differ significantly. Both raptors preferred slightly different stands for nesting compared to available stands in the study forest, suggesting that these small differences between the species were because of species-specific preferences, not the outcome of direct competition. Lesser Spotted Eagle pairs tended to space themselves regularly apart from each other, whilst the Common Buzzard pairs were distributed randomly. There was no evidence for spatial avoidance of the larger raptor by the Common Buzzard. In summary, because different components of the nesting habitats were extensively overlapping, and the distribution of interspecific pairs lacked spatial avoidance, we suggest that the nest sites of both raptors were largely a shared resource, especially when located close to the forest edges.
The nest platforms of the two raptors were placed similarly in the canopy, however, trees with trunk forks were especially important. The stick nest platforms built by these two raptors were also similar in their dimensions (Drobelis Citation2004). Bielański (Citation2006) found that nest platform placement in trunk forks was the most frequent for the Common Buzzard and Northern Goshawk Accipiter gentilis. Other studies of forest-dwelling sympatric raptors have not reported differences in nest platform placement height from the ground (Krüger Citation2002, Bielański Citation2006, Lõhmus Citation2006, Poirazidis et al. Citation2007), suggesting an extensive overlap of the nest platform placements, at least among raptors of similar body size.
The nest trees were similar in diameter, but the use of tree species was different. In other areas, sympatric raptors have also usually nested in trees of similar size (Krüger Citation2002, Lõhmus Citation2006, Poirazidis et al. Citation2007, but see Bielański Citation2006). Nearly half of the eagle pairs nested in Norway Spruce, while half of the Common Buzzards preferred Silver Birch; however, the frequency of nesting in Aspen and Black Alder trees was similar. For the Lesser Spotted Eagle, the use of Norway Spruce as the main nest tree species is evident over a large geographical area: 52% in Lithuania (Treinys Citation2015), 67% in NE Poland (Mirski Citation2009), 47% in Belarus (Ivanovski & Tishechkin Citation1993), 42% in Latvia (Bergmanis Citation2004) and 71% in Estonia (Väli Citation2003). The importance of the spruce for the eagle cannot be explained by the availability of that tree species in local breeding areas. Lõhmus (Citation2006) found that Lesser Spotted Eagles over-selected spruce as nest trees compared with the availability of the tree species in the study area, as well as in nest stands; a finding supported by Mirski (Citation2009) in Poland. A similar pattern was observed in our study area (the mean proportion of spruce in the stand composition was 25%, but 46% of nest platforms were in spruce). The proportions of Common Buzzard nest trees in Lithuania were similarly distributed across birch (25%), pine (20%), oak (20%) and spruce (19%) (Drobelis Citation2004). Common Buzzard nest tree species proportions and dominant nest tree species vary spatially (Bielański Citation2006, Lõhmus Citation2006, Väli Citation2015), with no consistent preferences for tree species compared to the Lesser Spotted Eagle. All of this suggests that: (1) the differences in the main nest trees used is not a result of interspecific competition; (2) the association of the Lesser Spotted Eagle with spruce indicates a limited plasticity of nest tree selection compared to the Common Buzzard and (3) the use of other tree species for nesting by these raptors overlaps significantly.
Direct comparison of the nest stands occupied by the two species indicates that the Common Buzzard nested further away from the forest edge, in forest stands growing on drier soil. Other characteristics were similar, although it is worth mentioning that there was some indication of more abundant birch in the nest stands of the Common Buzzard. Previous studies have repeatedly demonstrated that subdominant raptor species avoid habitats associated with dominant heterospecific raptors (Björklund et al. Citation2016, Michel et al. Citation2016), or selected nest sites in habitats avoided by the dominant species (Gamauf et al. Citation2013). Could these results indicate that the Common Buzzard avoids the larger raptor through space-mediated and habitat-mediated mechanisms? We provide three arguments that negate the interference competition mechanism as an explanation of the observed nest stand differences:
The Lesser Spotted Eagle occupied nests located close to the forest edge in our study area, similarly to elsewhere in Lithuania (Treinys et al. Citation2011) and other parts of its distribution range (Estonia, Lõhmus Citation2006; Poland, Mirski Citation2009). The nest stands of the Common Buzzard, however, were dispersed over the study area, with no indication of forest edge avoidance or a preference for the forest interior. Similar spatial patterns of the Common Buzzard (i.e. distribution in a forest according to landscape availability) have been found in two different areas studied in Poland (Bielański Citation2006). Moreover, in Estonia, the Common Buzzard nested close to the forest edge (Lõhmus Citation2006, Väli Citation2015), at a similar distance to the Lesser Spotted Eagle (Lõhmus Citation2006). These findings suggest that the nest stand location difference is caused by the Lesser Spotted Eagle’s strong association with the forest edge, not by the Common Buzzard’s avoidance of forest edges.
The Lesser Spotted Eagle nest stands only weakly differed from the available stands in the studied forest, based on soil wetness, contrary to the Common Buzzard, which strongly preferred drier stands compared to the available ones. The reverse pattern was observed with birch, which was strongly avoided by the Lesser Spotted Eagle, and only marginally preferred by the Common Buzzard. Furthermore, this marginal preference for birch may be related to its importance as the principal nest tree of the smaller raptor in the study area. Hence, the nest stand differences that may have been explained as species-specific preferences are not related to interspecific interactions.
We did not find any indication that the Common Buzzard avoided the Lesser Spotted Eagle, as has also been demonstrated in other studies, where subdominant raptors avoid dominant heterospecific raptors by increasing the distance to that species (Gamauf et al. Citation2013, Rebollo et al. Citation2017). Furthermore, the minimal distance between neighbouring pairs of Common Buzzard was 0.5 km in our study area, similarly to two study areas in Poland (0.46 and 0.55 km) (Bielański Citation2006) and in North Wales (Dare & Barry Citation1990). In this study, 15% of Common Buzzards nested within 0.51 km of Lesser Spotted Eagle pairs, and the minimum interspecific distance was less than 0.1 km. A smaller minimal distance to an interspecific compared to an intraspecific pair was also found in a community of similar-sized eagles, whose distribution was not explained by interspecific competition interactions (Katzner et al. Citation2003).
Considering these results, it is likely that nest platform and nest tree are the most similar component in the nesting habitat between the widespread Common Buzzard and the protected Lesser Spotted Eagle. The nest stands also overlapped extensively, but less so in patterns observed at lower spatial scales. Similarly, the strongest differentiation in the habitat components at a higher spatial scale was found previously and this is woodland size and hunting habitat (Kostrzewa Citation1996, Krüger Citation2002). It can be concluded that one of the most important habitat partitions in our study area were realized because eagles nested close to the forest edge and buzzards were spaced over the entire forested area. This also leads us to suggest that the highest degree of nesting habitat overdispersion occurs near the forest edges. We did not find any support for the Common Buzzard spatially avoiding the larger raptor. The observed slight habitat differentiation may be unlikely to be a consequence of interference competition. These results, however, could be interpreted as a high potential for exploitative competition for a shared resource if its supply decreases.
Can a similar habitat-overlap pattern be temporally stable? The availability of suitable forests plots for nesting may decrease because of timber harvesting, thus stimulating competitive interactions, leading to dominant raptor establishment in nest sites previously used by other raptors (Hakkarainen et al. Citation2004). Contrarily, the occupation of each other’s nest sites by three raptor species has been observed (albeit rarely) in an area with a large supply of vacant sites (Jiménez-Franco et al. Citation2011). The local density, especially in vole-eating raptors, may vary considerably over several years (Newton Citation2003). The density of the Lesser Spotted Eagle, however, has been quite stable over the years across a large region, despite fluctuations in the main prey (Treinys et al. Citation2017), however, the number of breeding pairs of Common Buzzards can change considerably, even between two consecutive years, in response to increased prey abundance (Tóth Citation2014). Hence, different numbers of heterospecific pairs might share a similar supply of suitable forest patches from year to year. This has led us to suppose that the degree of habitat overlap and interspecific interaction might be time-dependent.
Can a similar habitat-overlap pattern be expected elsewhere? Raptors vary spatially in their local densities in response to resource supply (Newton Citation2003), including the Common Buzzard and the Lesser Spotted Eagle (Dombrovski & Ivanovski Citation2005, Treinys et al. Citation2017), resulting in different numbers and ratios of heterospecific pairs within communities. Furthermore, raptor habitat preferences vary spatially due to environmental heterogeneity, as demonstrated by Väli et al. (Citation2004) for the Lesser Spotted Eagle and by Bielański (Citation2006) for the Common Buzzard. As a consequence of spatial variability in species habitat along environmental gradients, it might be expected that various degrees of habitat overlap will occur between the same sympatric forest-dwelling species (Skuja et al. Citation2019).
Conservation implications
The protection of Common Buzzard nest sites applying buffer zones for the entire local population in an SPA cannot be justified as a measure to considerably improve the nesting habitat protection of the Lesser Spotted Eagle because the latter species is strongly associated with forest edges. We propose applying the protection of Common Buzzard nest sites if they are located: (1) up to 0.5 km from Lesser Spotted Eagle nest sites (see below); and (2) less than 0.43 km from the forest edge (90% of eagle nests are within this distance from the forest edge; herein). The reasons behind this proposal are: (1) the Lesser Spotted Eagle in a defined territory alternates among a few nests located several hundreds of metres apart in different years; approximately 80% of movements of the same pair between alternative nests has been observed within this distance in Lithuania (RT unpublished data); (2) some Common Buzzard pairs in the local population nest close to Lesser Spotted Eagle pairs; (3) the dimensions of the nest platform (Drobelis Citation2004), nest placement in the tree canopy and nest trees (in terms of trunk diameter and spectrum of tree species) are similar (this study); (4) the same nest platform can be occupied by buzzards or eagles in any particular year (Skuja & Budrys Citation1999), probably in cases when heterospecific pairs nest in close proximity and (5) this approach would eliminate mistakes in identifying bird species associated with newly found nests in the vicinity of old Lesser Spotted Eagle nests in years of unsuccessful breeding or during non-breeding periods. Finally, the protection of nest sites of the Common Buzzard, when they are located close to nest sites of the Lesser Spotted Eagle and in apparently similar habitats, may prevent suitable nesting patches from decreasing and may act as a precautionary measure to eliminate possible competitive interactions.
Additional information
Funding
References
- Barrientos, R. & Arroyo, B. 2014. Nesting habitat selection of Mediterranean raptors in management pinewoods: Searching for common patterns to derive conservation recommendations. Bird Conserv. Internat. 24: 138–151. doi: 10.1017/S0959270913000270
- Bartoń, K. 2019. MuMIn: Multi-Model Inference. R Package Version 1.43.6.
- Bates, D., Maechler, M., Bolker, B. & Walker, S. 2015. Fitting linear mixed-effects models using lme4. J. Stat. Soft. 67: 1–48. doi: 10.18637/jss.v067.i01
- Bergmanis, U. 2004. Analysis of breeding habitats of the Lesser Spotted Eagle Aquila pomarina in Latvia. In Chancellor, R.D. & Meyburg, B.U. (eds) Raptors Worldwide, 537–550. WWGBP/MME, Budapest.
- Bergmanis, U., Auninš, A., Petrinš, A., Cīrulis, V., Granāts, J., Opermanis, O. & Soms, A. 2015. Population size, dynamics and reproduction success of the Lesser Spotted Eagle (Aquila pomarina) in Latvia. Slovak. Raptor J. 9: 45–54. doi: 10.1515/srj-2015-0003
- Bielański, W. 2006. Nesting preferences of Common Buzzard Buteo buteo and Goshawk Accipiter gentilis in forest stands of different structure (Niepolomice Forest, Southern Poland). Biologia 61: 597–603. doi: 10.2478/s11756-006-0096-7
- Björklund, H., Santangeli, A., Blanchet, F.G., Huitu, O., Lehtoranta, H., Lindén, H., Valkama, J. & Laaksonen, T. 2016. Intraguild predation and competition impacts on a subordinate predator. Oecologia 181: 257–269. doi: 10.1007/s00442-015-3523-z
- Burnham, K.P. & Anderson, D.R. 2002. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, second ed. Springer, New York.
- Dare, P.J. & Barry, J.T. 1990. Population size, density and regularity in nest spacing of Buzzards Buteo buteo in two upland regions of North Wales. Bird Study 37: 23–29. doi: 10.1080/00063659009477033
- Dhondt, A.A. 2012. Interspecific Competition in Birds. Oxford University Press, Oxford.
- Dombrovski, V.C. & Ivanovski, V.V. 2005. New data on numbers and distribution of birds of prey breeding in Belarus. Acta Zool. Lituan. 15: 218–227. doi: 10.1080/13921657.2005.10512615
- Drobelis, E. 2004. Lietuvos miškų plėšrieji paukščiai [ Birds of Prey of Lithuanian Forests]. Ministry of the Environment of the Republic of Lithuania, Vilnius.
- Gamauf, A., Tebb, G. & Nemeth, E. 2013. Honey Buzzard Pernis apivorus nest-site selection in relation to habitat and the distribution of the Goshawk Accipiter gentilis. Ibis 155: 258–270. doi: 10.1111/ibi.12023
- Hakkarainen, H., Mykrä, S., Kurki, S., Tornberg, R. & Jungell, S. 2004. Competitive interactions among raptors in boreal forests. Oecologia 141: 420–424. doi: 10.1007/s00442-004-1656-6
- Ivanovski, V.V. & Tishechkin, A.K. 1993. Monitoring of Lesser Spotted Eagle (Aquila pomarina) in Byelorussia. Ring 15: 267–273.
- Jiménez-Franco, M.V., Martínez, J.E. & Calvo, J.F. 2011. Territorial occupancy dynamics in a forest raptor community. Oecologia 166: 507–516. doi: 10.1007/s00442-010-1857-0
- Jiménez-Franco, M.V., Martínez, J.E. & Calvo, J.F. 2014. Lifespan analyses of forest raptor nests: patterns of creation, persistence and reuse. PLoS ONE 9 (4): e93628. doi: 10.1371/journal.pone.0093628
- Jiménez-Franco, M.V., Martínez-Fernández, J., Martínez, J.E., Pagán, I., Calvo, J.F. & Esteve, M.A. 2018. Nest sites as a key resource for population persistence: A case study modelling nest occupancy under forestry practices. PLoS ONE 13 (10): e0205404. doi: 10.1371/journal.pone.0205404
- Kajtoch, Ł, Żmihorski, M. & Wieczorek, P. 2015. Habitat displacement effect between two competing owl species in fragmented forests. Popul. Ecol. 57: 517–527. doi: 10.1007/s10144-015-0497-y
- Katzner, T.E., Bragin, E.A., Knick, S.T. & Smith, A.T. 2003. Coexistence in a multispecies assemblage of eagles in central Asia. Condor 105: 538–551. doi: 10.1093/condor/105.3.538
- Kostrzewa, A. 1996. A comparative study of nest-site occupancy and breeding performance as indicators for nesting habitat quality in three European raptor species. Ethol. Ecol. Evol. 8: 1–18. doi: 10.1080/08927014.1996.9522931
- Krüger, O. 2002. Analysis of nest occupancy and nest reproduction in two sympatric raptors: Common Buzzard Buteo buteo and Goshawk Accipiter gentilis. Ecography 25: 523–532. doi: 10.1034/j.1600-0587.2002.250502.x
- Lõhmus, A. 2001. Habitat selection in a recovering osprey Pandion haliaetus population. Ibis 143: 651–657. doi: 10.1111/j.1474-919X.2001.tb04893.x
- Lõhmus, A. 2006. Nest-tree and nest-stand characteristics of forest-dwelling raptors in east-central Estonia: Implications for forest management and conservation. Proc. Estonian Acad. Sci. Biol. Ecol. 55: 31–50.
- Lõhmus, A. & Sellis, U. 2003. Nest trees – A limiting factor for the black stork population in Estonia. Aves 40: 84–91.
- Meyburg, B.-U., Haraszthy, L., Strazds, M. & Schäffer, N. 2001. European species action plan for Lesser Spotted Eagle. In Schäffer, N. & Gallo-Ursi, U. (eds) European Union Actions Plans for Eight Priority Species, 1–24. Office for Official Publications of the European Communities, Luxemburg.
- Meyburg, B.-U., Meyburg, C., Bělka, T., Šreibr, O. & Vrana, J. 2004. Migration, wintering and breeding of a Lesser Spotted Eagle (Aquila pomarina) from Slovakia tracked by satellite. J. Ornithol. 145: 1–7. doi: 10.1007/s10336-003-0014-5
- Michel, V.T., Jiménez-Franco, M.V., Naef-Daenzer, B. & Grüebler, M.U. 2016. Intraguild predator drives forest edge avoidance of a mesopredator. Ecosphere 7 (3): e01229. doi:10.1002/ecs2.1229.
- Mirski, P. 2009. Selection of nesting and foraging habitat by the Lesser Spotted Eagle Aquila pomarina (Brehm) in the Knyszyńska forest (NE Poland). Pol. J. Ecol. 57: 581–587.
- Newton, I. 2003. The role of natural factors in the limitation of bird of prey numbers: A brief review of the evidence. In Thompson, D.B.A., Redpath, S.M., Fielding, A.H., Marquiss, M. & Galbraith, C.A. (eds) Birds of Prey in a Changing Environment, 5–23. Scottish Natural Heritage/The Stationery Office, Edinburgh.
- Ortego, J. 2007. Consequences of Eagle Owl nest-site habitat preference for breeding performance and territory stability. Ornis Fenn. 84: 78–90.
- Poirazidis, K., Goutner, V., Tsachalidis, E. & Kati, V. 2007. Comparison of nest-site selection patterns of different sympatric raptor species as a tool for their conservation. Anim. Biodiv. Conserv. 30 (2): 131–145.
- Poole, A.F. 1989. Regulation of Osprey Pandion haliaetus populations: The role of nest side availability. In Meyburg, B.-U. & Chancellor, R.D. (eds) Raptors in the Modern World, 227–234. WWGBP, Berlin.
- R Core Team. 2018. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/.
- Rebollo, S., Martínez-Hesterkamp, S., García-Salgado, G., Pérez-Camacho, L., Fernández-Pereira, J.M. & Jenness, J. 2017. Spatial relationships and mechanisms of coexistence between dominant and subordinate top predators. J. Avian Biol. 48: 1226–1237. doi: 10.1111/jav.01337
- Richardson, C.T. & Miller, C.K. 1997. Recommendations for protecting raptors from human disturbance: A review. Wildlife Soc. B. 25: 634–638.
- Sarà, M., Mascara, R. & López-López, P. 2016. Understanding the coexistence of competing raptors by Markov chain analysis enhances conservation of vulnerable species. J. Zool. 299: 163–171. doi: 10.1111/jzo.12340
- Selås, V. 1997. Nest-site selection by four sympatric forest raptors in southern Norway. J. Raptor Res. 31: 16–25.
- Skuja, S. & Budrys, R.R. 1999. Nesting sites of Black Stork, Lesser Spotted Eagle and Common Buzzard and their nest exchange in the forests of north, north-east and central Lithuania. Balt. For. 5: 67–74.
- Skuja, S., Mozgeris, G. & Treinys, R. 2019. May sympatric Lesser Spotted Eagles and Black Storks compete for nesting sites in spatially varying environments? Balt. For. 25: 63–68.
- Steenhof, K. & Newton, I. 2007. Assessing nesting success and productivity. In Bird, D.M. & Bildstein, K.L. (eds) Raptor Research and Management Techniques Manual, 181–192. Hancock House, Surrey, British Columbia, Canada.
- Tóth, L. 2014. Numerical response of the Common Buzzard Buteo buteo to the changes in abundance of small mammals. Ornis Hungarica 22: 48–56. doi: 10.2478/orhu-2014-0010
- Treinys, R. 2015. Species Action Plan for the Lesser Spotted Eagle. Accepted by the Ministry of Environment of Republic Lithuania on 27 February 2015 by Order No. D1-185.
- Treinys, R. & Mozgeris, G. 2006. Past and present nest-site requirements of the Lesser Spotted Eagle (Aquila pomarina C.L. Brehm) and their possible conflicts with timber harvesting. Balt. For. 12: 252–258.
- Treinys, R. & Mozgeris, G. 2010. Relationship between the productivity of the Lesser Spotted Eagle Aquila pomarina and forest characteristics at the macrohabitat level. Balt. For. 16: 87–92.
- Treinys, R., Dementavičius, D., Mozgeris, G., Skuja, S., Rumbutis, S. & Stončius, D. 2011. Coexistence of protected avian predators: Does a recovering population of white-tailed eagle threaten to exclude other avian predators? Eur. J. Wildlife Res. 57: 1165–1174. doi: 10.1007/s10344-011-0529-7
- Treinys, R., Bergmanis, U. & Väli, Ü. 2017. Strong territoriality and weak density-dependent reproduction in Lesser Spotted Eagles Clanga pomarina. Ibis 159: 343–351. doi: 10.1111/ibi.12454
- Väli, Ü. 2003. The Lesser Spotted Eagle and its conservation in Estonia. Hirundo Supplementum 6: 1–64.
- Väli, Ü. 2015. Habitat selection of the Common Buzzard (Buteo buteo) on the island of Hiiumaa, Western Estonia. Hirundo 2015: 19–28.
- Väli, Ü, Treinys, R. & Lõhmus, A. 2004. Geographical variation in macrohabitat use and preferences of the Lesser Spotted Eagle Aquila pomarina. Ibis 146: 661–671. doi: 10.1111/j.1474-919x.2004.00300.x