
ABSTRACT
Capsule
Population trends are estimated for the first time for 12 species based on Shetland Breeding Bird Survey data collected between 2002 and 2019.
Aim
To provide population trends for a selection of commoner breeding waders and passerines on the Shetland Isles, the most northerly region of the UK, which has historically been under-represented in national bird monitoring schemes.
Methods
One hundred and forty-five self-selected 1-km2 squares were surveyed between 2002 and 2019 using similar methods to the BTO/JNCC/RSPB Breeding Bird Survey (BBS), the main breeding bird monitoring scheme in the UK.
Results
Over 17 years, Eurasian Oystercatcher Haematopus ostralegus, Northern Lapwing Vanellus vanellus, and Common Redshank Tringa totanus populations declined whilst Eurasian Curlew Numenius arquata and Common Snipe Gallinago gallinago showed no significant changes in their population trends. For passerines, Common Skylark Alauda arvensis and Northern Wheatear Oenanthe oenanthe populations declined, whilst Common Starling Sturnus vulgaris, Common Blackbird Turdus merula, and House Sparrow Passer domesticus increased. Winter Wren Troglodytes troglodytes and Meadow Pipit Anthus pratensis showed no significant population changes. Comparisons with BBS trends for the UK and for Scotland are provided. For Eurasian Curlew, Common Starling, and House Sparrow, trends were more favourable in Shetland than elsewhere in UK.
Conclusion
The Shetland Breeding Bird Survey provides useful trends for several bird species, which include important breeding numbers of Winter Wren and Common Starling (both endemic subspecies), as well as nationally important wader populations. We recommend increasing the number of survey squares sampled each year, which would increase the precision of population estimates. The inclusion of a random or stratified element to square selection would help to ensure that these trends are representative of the wider region.
Birds are not only of wide interest in their own right but are also used to indicate the state of the environment (Furness & Greenwood Citation1993). Breeding bird surveys create robust indices of abundance that can be used to assess temporal change. In the UK, the British Trust for Ornithology’s (BTO) Common Birds Census (CBC), which ran from 1962 to 2000, was based on observers selecting their own survey plots and formed a highly useful resource (Freeman et al. Citation2007). Since 1994, the CBC has been replaced by the Breeding Bird Survey (BBS; organized by BTO/Joint Nature Conservation Committee (JNCC)/Royal Society for the Protection of Birds (RSPB)), using random sampling methods to provide trends for a range of common and widespread species (Harris et al. Citation2020). Data from these surveys have been used for a wide range of purposes, including the identification of range expansions due to global climate change (Massimino et al. Citation2015), assessing the effectiveness of environmental stewardship for farmland birds (Baker et al. Citation2013), and detecting declines in Afro-Palearctic migrant birds linked to changes in habitat at breeding and/or wintering grounds (Ockendon et al. Citation2012). Across the UK, regional variation occurs in the trends of some species, driven by regional- and local-scale processes such as climate change and breeding habitat modification (Morrison et al. Citation2010, Citation2015). This highlights the importance of ensuring a representative geographical spread of breeding bird survey sites to detect regional differences in trends over time, to help identify local drivers of population change, and to identify which regions nationally are most important for species of conservation concern. Regional trend variation is particularly important where populations are isolated; for example, resident island populations of otherwise widespread species that might harbour important intra-specific variation, such as unique genetic forms or subspecies.
Shetland’s avifauna is largely known for its globally important seabird populations (Mitchell et al. Citation2004) and rarer UK breeding species, such as nationally important breeding populations of Red-necked Phalarope Phalaropus lobatus and Eurasian Whimbrel Numenius phaeopus (Massey et al. Citation2016, Holling & the Rare Breeding Birds Panel Citation2019). Whilst these rare species have been relatively well studied, commoner wader and passerine species have received less attention. These include several endemic subspecies found in Shetland for which there is a lack of available data. Among these are two subspecies of Winter Wren (the Fair Isle Winter Wren Troglodytes troglodytes fridariensis, which is present only on Fair Isle, Shetland’s most southerly island; and the Shetland Wren T. t. zetlandicus, which is present across the rest of Shetland); and a sub-species of the Common Starling, Sturnus vulgaris zetlandicus.
Uptake of the national BBS has been low in Shetland due to the logistics of conducting counts at the randomly allocated set of 1-km survey squares, many of which are located in remote areas and could include areas of dangerous terrain. Surveys at these squares are made still more difficult by the high number of summer days in Shetland with wind speeds exceeding the recommended survey limit of Beaufort scale four; and by the very changeable weather, which can make it difficult to take advantage of short windows of suitable conditions to carry out surveys at remote sites. To address the lack of data from BBS, the Shetland Breeding Bird Survey (hereafter referred to as SBBS) was established in 2002 as a partnership between Shetland Amenity Trust (a local community organization) and Shetland Bird Club. Here, we use data collected for the SBBS from 2002 to 2019 to provide estimated population trends for several common wader and passerine species in Shetland, acknowledging the limitations associated with the non-random allocation of survey sites. This is the first long-term analysis carried out on the SBBS data.
Methods
Field methods
Across Shetland, 145 1-km2 squares were surveyed (2002–2019) by volunteers (). Survey squares were self-selected by the surveyors and, as a result, were in easily accessible locations. Following comparable recording protocols to the BBS (www.bto.org/bbs), surveys were carried out during the breeding season with a first visit between 20th April and 10th May and a second visit between 20th May and 10th June. The higher latitude of Shetland means that breeding phenology is expected to be later; therefore the first visit was later than the BBS start date of early April. Surveys started before 09:00, in dry conditions, with Beaufort wind scale force 4 or less. Repeated survey routes comprised of approximately 2 km routes through a 1-km square to ensure as much of each square was covered as possible, whilst avoiding residential areas and freshwater lochs. Birds were recorded as being located either within 100 m of the transect route or over 100 m from the route. Several behaviours, including singing, displaying, alarming, defending territories, carrying food, or carrying a faecal sac, were recorded and used to define possible/probable breeding pairs. Seabirds, including divers (Gaviidae), were covered by separate monitoring schemes and so were not included in the SBBS.
Figure 1. Locations of the 145 Shetland Breeding Bird Survey 1-km2 squares monitored between 2002 and 2019. 1-km2 are coloured along a gradient based on the number of years they have been surveyed, from yellow (one year) to dark red (all 18 years).
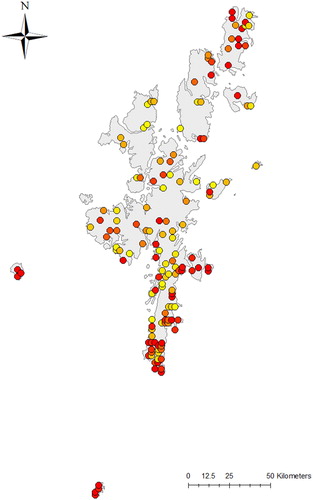
SBBS indices for each year are based on interpretation of data to produce a minimum number of pairs per 1-km2 square. Country and regional trends from the BBS are routinely reported for species recorded in a minimum annual mean of 30 survey squares during the reporting period (Harris et al. Citation2020). For results of the SBBS presented here, we have adopted the same reporting threshold. There was variation in annual survey square coverage, ranging from 1 to 18 years (), with an overall increase in the number of squares covered over the 18-year period.
Data analysis
We fitted log-linear trend models to evaluate Shetland trends during the period 2002–2019 using TRIM (Trends and Indices for Monitoring data) models (Pannekoek & van Strien Citation2006a, Citation2006b), creating mean indices, and associated standard errors in the R package rtrim (Bogaart et al. Citation2018). The overall slopes of these models are the results of linear regressions over time, plus site, and year effects that measure deviation from the overall slope. An advantage of TRIM is that it allows for the absence of data in some years at some squares. TRIM uses a stepwise selection of change-points in trends using Wald-tests to test the significance of change-points. The best-fit models were selected following Pannekoek & van Strien (Citation2006a, Citation2006b). A model with a significance value greater than 0.05 indicates that the data do not differ significantly from a Poisson distribution. However, even where data do not follow a Poisson distribution, the indices, overall slope, and change points remain reliable (Pannekoek & van Strien Citation2006a, Citation2006b). The model takes any overdispersion into consideration and the significance of the slope is assessed using a Wald-test (Pannekoek & van Strien Citation2006a, Citation2006b). TRIM describes trends based on the mean annual rate of change and defines them as either a strong (>5% mean change per year) or moderate decrease (<5%). Trends are considered to be significantly divergent from stable when the 95% confidence interval of the slope does not contain zero ().
Table 1. Sample sizes and population trend estimates for the 12 species based on Shetland Breeding Bird Survey data collected between 2002 and 2019.
To compare the estimated trends calculated for species in Shetland to those in other parts of the UK, we obtained BBS trends (www.bto.org/bbs) of species for Scotland and the UK (). Trends derived from the SBBS and BBS in Scotland and UK were considered to differ if the 95% confidence intervals of the mean annual change over 2002–2019 did not overlap, to account for differences in periods covered.
Table 2. Species comparisons of population trend estimates, with 95% confidence intervals, for the Shetland Breeding Bird Survey (SBBS) and the Breeding Bird Survey (BBS) for Scotland and the UK.
Results
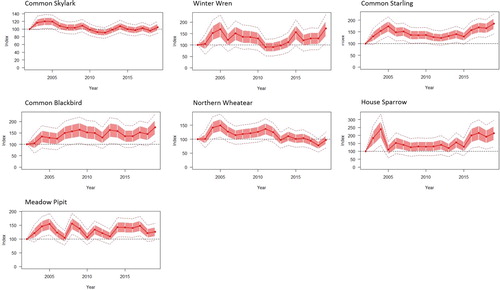
Twelve species were recorded in a mean of 30 or more SBBS survey squares between 2002 and 2019, enabling trends in breeding populations to be calculated. These included five wader species () and seven passerines (). Eurasian Oystercatcher Haematopus ostralegus, Northern Lapwing Vanellus vanellus, Common Redshank Tringa totanus, Common Skylark Alauda arvensis, and Northern Wheatear Oenanthe oenanthe generally declined across study squares over the survey period, whilst Common Blackbird Turdus merula, Common Starling, and House Sparrow Passer domesticus generally increased. No significant change in population size was detected for Eurasian Curlew Numenius arquata, Common Snipe Gallinago gallinago, Winter Wren, and Meadow Pipit Anthus pratensis (). The population trend estimates reported in show a constant population change rate for species over the whole survey period; however, there is variation in the annual indices between 2002 and 2019 for some species ( and ). This is particularly evident for the Common Snipe, which showed a significant decline between 2003 and 2011 (mean change [95% confidence intervals]: −0.09% [−0.017, 0.053], P = 0.02), followed by an increase between 2011 and 2019 (1.1% [0.02, 0.10], P < 0.001).
Figure 2. Changes in annual indices for the five wader species, with indices measured relative to that of the first survey year (2002) and standard error (red shaded areas) and 95% confidence intervals (dotted line) shown.
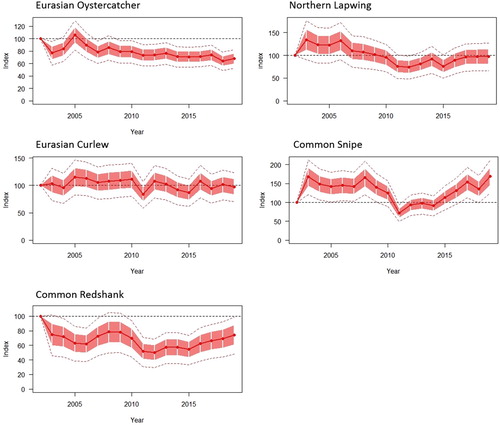
Figure 3. Changes in annual indices for seven passerine species with indices measured relative to that of the first survey year (2002), with standard error (red shaded areas) and 95% confidence intervals (dotted line).
For the waders, the rate of decline for the Eurasian Oystercatcher, Northern Lapwing, and Common Redshank in Shetland was comparable to that recorded in Scotland and across the UK over a similar period (). The Shetland trend for the Common Snipe was also comparable to that of Scotland, with no change being detected, in contrast to the UK as a whole, where Common Snipe has increased. For the Eurasian Curlew, no significant trend was detected in Shetland in contrast to the declines observed in this species in Scotland and the UK as a whole.
For passerines, the rate of decline for the Common Skylark and Northern Wheatear in Shetland was comparable to those observed in Scotland and the UK (). The increase in Common Blackbird in Shetland follows a similar increase in Scotland and the UK, whilst House Sparrow increased in both Shetland and Scotland, in contrast to the whole UK population for which the trend appears to be stable. Common Starling showed the greatest divergence in trends, with an increase in Shetland compared to declines in Scotland and the UK.
Discussion
This first analysis of Shetland BBS data delivers recent trends for 12 bird species and an opportunity for comparison with the rest of the UK. Changes in breeding wader and passerine populations in the UK have been linked to a suite of reasons, including changes in agricultural practices, the management of the wider landscape, and predator numbers (Fuller et al. Citation1995, Evans Citation2004, Fletcher et al. Citation2010, Franks et al. Citation2017). Some higher intensities of grazing and the simplification and intensification of agricultural systems have particularly been associated with the declines of some breeding waders elsewhere (Roodbergen et al. Citation2012, Calladine et al. Citation2014). The climate, topography and soil conditions, and distance from external markets on Shetland are unsuitable for the types of agricultural practices that have intensified most markedly elsewhere in the UK. Land is also largely managed under a traditional crofting system, characterized by less intensive mixed pasture and arable with light grazing by sheep and cattle, with a small-scale land ownership pattern and a cultural link to traditional practices. Shetland has also experienced much less of the new woodland development that has displaced waders from parts of mainland UK (Douglas et al. Citation2014, Calladine et al. Citation2018), although the small areas of plantation that do occur may still have an effect on ground-nesting birds. Shetland experienced agricultural changes during the 1970s, when there was a move from a regime dominated by traditional crofting to one in which widespread sheep grazing became more pronounced, due to the introduction of European Union subsidies that encouraged higher sheep stocking rates. Between 1987 and 2000, the Environmentally Sensitive Area agri-environment scheme supported the removal of sheep from the hills to reduce grazing intensity, which may have benefited Eurasian Curlew and Common Snipe through subsequent vegetation recovery. Since 2000, with a switch away from headage payments to basic or area payments, stock numbers have been reduced further. This has led to more ungrazed pasture and an increase in sward height in many fields. Leatherjacket (Tipulidae) numbers rise markedly when the grass grows longer and this could provide food for waders (Fraser, pers. comm.). However, this may not have benefited Northern Lapwings with their preference for foraging in short swards (McCallum et al. Citation2015, Citation2016). Farming in Shetland was further intensified when the Shetland Islands Council offered low-interest loans that allowed crofters to join the Crofting Agricultural Grant Scheme (Harvey, pers. obs.). This led to increased liming and re-seeding of hill land, agricultural improvement of in-bye, and some drainage. Many of these areas of ‘agriculturally improved’ hill are now reverting and can provide attractive habitat for waders. It is likely that the mosaic created by some re-seeding of hill land benefitted waders by providing higher prey densities in the agriculturally improved rough pasture than the heathland (Grant et al. Citation1992). Furthermore, the Scottish Agricultural Census shows a 27% decline in crop and fallow land use (2000–2012) in Shetland, particularly since 2008 (Martin Citation2005). Simplification of agricultural mosaics towards exclusive pasture or exclusive arable has been associated with declines in breeding waders (Bell & Calladine Citation2017).
Although the lower intensification of farming on Shetland, compared to mainland UK, has seemingly enabled some Shetland wader and passerine populations to remain stable, the collective changes in habitat and agricultural practices have likely negatively influenced other species. The decline in Northern Wheatear populations is likely to have been influenced by habitat changes in Shetland. UK declines in Northern Wheatear have been linked to losses of suitable grassland habitat and under-grazing, possibly due to declines in Rabbit Oryctolagus cuniculus abundance, though Rabbits are still relatively abundant on Shetland (Baillie et al. Citation2010, Kämpfer & Fartmann Citation2019). The decline in Common Skylark observed across Scotland and the rest of the UK, which is largely driven by marked declines in lowland and arable areas, is also reflected in the Shetland trend for this species. In Shetland, the majority of Common Skylarks tend to be associated with moorland and areas of grassland that have not been ‘agriculturally improved’ (Ellis Citation2004). An investigation of specific trends in these habitats compared to arable/agriculturally improved grassland habitats could help to explain the apparent decline in Common Skylark populations. In other parts of the UK, declines in populations of Common Skylark are likely due to changes in agricultural practices, such as the shift from spring- to autumn-sown cereals (Ellis Citation2004, Donald & Morris Citation2005). To identify drivers of population changes related to land use and farming practices, research into land cover and habitat change over the survey period is required. This would also provide evidence to target future environmental support payments and effectively incentivise beneficial land management practices on Shetland. Sensible fiscal incentives to crofters could be mutually beneficial to birds and people given the socio-economic importance of crofting on Shetland. Targeted agri-environment spend on Shetland could also help to support its nationally important wader populations. In particular, the Eurasian Curlew, a red-listed Bird of Conservation Concerns (BOCC) (Eaton et al. Citation2015), is faring better on Shetland than elsewhere in the UK.
Mammalian predators of ground-nesting birds found on Shetland include Polecat × Ferret Mustela putorius × Mustela putorius furo and Stoat Mustela erminea, plus Otter Lutra lutra, Brown Rat Rattus norvegicus, Hedgehog Erinaceus europaeus, and Domestic Cat Felis catus domesticus. All of these species are introduced and can pose a threat to breeding waders (Baines Citation1990, Grant et al. Citation1999, Macdonald & Bolton Citation2008). Hedgehogs, where these have been introduced onto islands, have also been shown to cause lower breeding success and breeding population declines in waders (Jackson et al. Citation2004, Calladine et al. Citation2015, Calladine et al. Citation2017). Shetland’s ground-nesting birds may benefit from a lack of Red Fox Vulpes vulpes and forest plantations. Foxes are one of the most important predators of breeding waders in the UK (Roos et al. Citation2018) and breeding waders may avoid suitable habitats near forest edges, due to higher levels of mammalian predator activity along these habitat interfaces (Douglas et al. Citation2014, Hancock et al. Citation2020). There is also the potential of predation by native avian species predators such as gulls, skuas, and Hooded Crow Corvus cornix that breed across Shetland. Given that Eurasian Oystercatcher, Northern Lapwing, and Common Redshank numbers have declined on Shetland it is important to understand whether, and to what extent, these declines are caused by predation, habitat change, or other factors.
Stoats were established on mainland Shetland before the late seventeenth century, whilst Polecat x Ferret hybrids were introduced in the 1980/1990s. An eradication project is in progress on Orkney to remove recently introduced non-native Stoats due to concerns about the risk they pose to ground-nesting birds and native mammals (Fraser et al. Citation2015). A lot could be learnt from this eradication attempt in terms of determining how feasible, and successful, a similar project might be on Shetland. The reduced suite of predators on Shetland (compared to mainland UK), together with influences of habitat and land use, may be responsible for the relatively healthy populations of some ground-nesting species, such as Eurasian Curlew and Common Snipe. The SBBS survey squares were not restricted to mainland Shetland but included nine smaller islands with different mammal faunas. A further study would benefit from comparing mainland trends to those on islands without particular introduced mammal species, such as Stoat. Among these islands, Foula, Fair Isle, Unst, Yell, Fetlar, Housay, and South Havra are all recommended as priority sites in the UK for invasive vertebrate eradications, based on eradication benefit feasibility and low reinvasion risk (Stanbury et al. Citation2017).
Common Snipe is most reliably detected outside the times and weather conditions when most breeding bird surveys are undertaken, with individual birds varying the frequency of their drumming displays according to wind speed, precipitation level, time of day, and season (Hoodless et al. Citation2006). This inflates the variance around the annual indices (wide confidence intervals), resulting in lower power to detect differences between years. However, given that the Scotland BBS detected no significant change, and the UK BBS an increase, it is expected that the stable trend for Common Snipe in Shetland is robust. Interestingly, the Common Snipe declined in numbers between 2008 and 2011, followed by an increase to 2019.
The Shetland trends for Winter Wren and Meadow Pipit were stable, and comparable to the trends for these species in Scotland and the UK. In contrast, populations of House Sparrow and the zetlandicus subspecies of Starling have increased in Shetland, against a background of UK-level declines for both species, and a Scottish decline for House Sparrow. Generally, resident insectivores may have benefitted from increased winter temperatures (1.2°C between 1947 and 2009, Dawson et al. Citation2011) and the availability of shrubs and trees in Shetland gardens. An increased interest in horticulture in Shetland since the 1990s has led to a large increase in the cover provided by gardens that could also explain the increasing Common Blackbird population (P. Harvey, pers. comm.). Poor first-year survival has been identified as a likely contributing factor to the UK-level decline of Starling (Freeman et al. Citation2007). As well as benefitting from milder winters and improvements in garden vegetation, survival rates of young Starlings in Shetland could also benefit from the foraging opportunities associated with less intensive farming and larger areas of intertidal habitat than are available to more southern populations. Starlings may also benefit from bird food provided in gardens and were recorded as the most abundant species on Shetland during the RSPB’s ‘Big Garden Birdwatch’ survey in 2019, with records in 96% of gardens surveyed (https://www.rspb.org.uk/get-involved/activities/birdwatch/results/). An increase in grant-aided woodland has also enabled European Robin Erithacus rubecula, Eurasian Blackcap Sylvia atricapilla, Willow Warbler Phylloscopus trochilus, Common Chiffchaff Phylloscopus collybita, Goldcrest Regulus regulus, Common Linnet Linaria cannabina, Eurasian Siskin Spinus spinus, and Common Redpoll Acanthis flammea to become regular breeders in Shetland since the 1990s (Shetland Bird Report Citation2019), with Eurasian Sparrowhawk Accipiter nisus breeding for the first time in 2020 (Shetland Bird Report Citation2020).
Although SBSS survey squares are well distributed across the Shetland Islands (), they were not randomly selected. As a result, selected survey squares may be unrepresentative of the wider Shetland countryside in some respects. Survey square selection has tended to be biased towards lowland squares, and towards squares that are relatively close to roads and built-up areas. If changes in bird populations vary with respect to distance to roads, houses, or between lowlands and uplands, SBBS trends may be skewed for some species. This is also the case for historic coverage in Scotland, with oversampling of lowland sites and those in close proximity to human populations due to the non-random uptake of randomly selected squares, meaning that the Scottish BBS data, at least in upland regions, is biased in a similar way to the Shetland census (Darvill et al. Citation2020). Increasing the representativeness of survey squares (e.g. through random square selection) would make trends based on SBBS data less biased and more reliable. It is important to note that SBBS also differs from UK and Scottish BBS in other aspects than square selection, in ways that could complicate comparison of findings derived from these two surveys. Trends from the UK BBS are derived using the maximum number of recorded registrations of individual adult birds on either of the two annual survey visits to each square (with some corrections to minimize the influence of non-breeding wader flocks on these counts). By contrast, SBBS trends are based on the estimated number of pairs found in each square. Although SBBS and BBS are likely subject to different biases, and it would be useful to understand the implications of these biases more fully, trends derived from either survey are still likely to be broadly representative of bird populations in the areas they represent. Comparisons between these trends are, therefore, likely to reveal any substantial differences in bird population change between Shetland and other parts of the UK.
To develop the trends further for Shetland, a two strata approach is recommended: (i) non-random selected squares in the SBBS and potentially other sources of site-specific monitoring such as for nature reserves and (ii) a random sample of remaining areas. Combining these two strata will improve the representativeness of surveyed squares and confidence that trends derived from these accurately reflect the changes in Shetland’s bird populations. In addition, comparisons between the two strata could determine the extent to which the non-random selection of 1-km squares affects the SBBS derived trends. For waders, this comparison could also include the 1993 and 1998/1999, 60 randomly selected lowland 1-km2 squares to assess changes in wader populations since these earlier surveys, although the survey methodology differs in timing, effort, and area (O'Brien & Smith Citation1992) to the SBBS. Since 2002, the numbers of SBBS participants and survey squares have increased, improving the precision and statistical power of survey outputs. Further increasing the number of surveyed squares could increase the number of breeding species for which there are sufficient data to model trends. Species that did not meet the criteria of being recorded in a minimum annual mean of 30 survey squares include Common Ringed Plover Charadrius hiaticula (21), European Golden Plover Pluvialis apricaria (18), Dunlin Calidris alpinae (8), Rock Pipit Anthus petrosus (15), Hooded Crow (9), and Twite Linaria flavirostris (7).
Conclusion
Understanding bird trends on a regional scale is important for many reasons. Without BBS coverage, incremental population change may go unnoticed for long periods. Drivers of population change can be local as well as global, and it cannot be assumed that bird populations on Shetland behave the same way as they do elsewhere in the UK. Differences between Shetland and other parts of Scotland and the wider UK, particularly in the suite of mammalian predators present, may lead to differences in population trends between these regions.
Targeted monitoring of breeding bird populations is therefore particularly important for archipelagos such as the Hebrides, Orkney, and Shetland, as well as for more remote – in terms of accessibility – parts of Scotland, where current BBS participation is low. This is particularly true where locations have important numbers of endemic subspecies, such as the Hebridean Dunnock Prunella modularis hebridium and St Kilda Winter Wren Troglodytes troglodytes hirtensis. Part of this can be achieved through initiatives such as Upland Rovers (www.bto.org) – an incentive to encourage BBS surveys in remote and under-sampled areas of Scotland, where only one survey visit is the minimum required, and local schemes such as SBBS. Improved understanding of local population changes can improve conservation decisions and management. Monitoring local breeding bird populations means that local drivers of change, such as habitat change, local disturbance, food abundance, and predator changes, can be identified earlier allowing for more rapid conservation responses.
Acknowledgements
We thank Graham Fraser for advice on agriculture in Shetland and Mark Hancock for comments on earlier drafts. Thanks to all surveyors, notably those that have partaken for a number of years: Richard Ashbee, Geoff and Donna Atherton, Gary Bell, Geoff Blackman, Paul Bloomer, Nick Brett, Robbie Brookes, Juan Brown, Graham Bundy, Mark Chapman, Andy Cook, David Cooper, Dennis Coutts, Wendy Dickson, Steve Duffield, Pat Dugard, Jon Dunn, Nick Dymond, Harry Edwards, Pete Ellis, Margaret Fiddy, Peter Flint, Dick Foyster, Paul Fisher, Rob Fray, Andy Freeman, Joyce Garden, Andy Gear, Sheila Gear, Beth & Tony Gerrard, Paul Goddard, Dave Hall, Phil Harris, Andrew Harrop, Derek Herning, Martin Heubeck, Sally Huband, John-Lowrie Irvine, Barbara Johnson, Carol Johnson, Logan Johnson, Laughton Johnston, Ray Johnston, Micky Maher, Brian Marshall, Karen Mackelvie, Mick Mellor, Will Miles, Steve Minton, Jim Nangle, Jim Nicolson, Rebecca Nason, Reinoud Norde, Kevin Osborn, Dave Okill, Mike Pennington, George Petrie, Jonathon Pinnick, Nathalie Pion, Dougie Preston, Ann Prior, Julie Redpath, Roger Riddington, Iain Robertson, Derek Rushton, Martin Schofield, Guy Smith, Simon Smith, Jenny Sutherland, Jonathan Swale, Roger Tait, Rory Tallack, Brydon Thomason, Howard Towll, Glen Tyler, John Uttley, Sue White, Kris Wilson, David Wood, and various staff from Fair Isle Bird Observatory, RSPB, and SNH. We also thank the two reviewers for their constructive comments which improved the manuscript.
References
- Baillie, S.R., Marchant, J.H., Leech, D.I., Joys, A.C., Noble, D.G., Barimore, C., Downie, I.S., Grantham, M.J., Risely, K. & Robinson, R.A. 2010. Breeding birds in the wider countryside: Their conservation status 2009. BTO Research Report 541. BTO, Thetford. http://www.bto.org/birdtrends.
- Baines, D. 1990. The roles of predation, food and agricultural practice in determining the breeding success of the Lapwing (Vanellus vanellus) on upland grasslands. J. Anim. Ecol. 59: 915–929.
- Baker, D.J., Grice, P.V. & Siriwardena, G.M. 2013. How has Environmental stewardship affected English farmland bird populations? Results and lessons from a national assessment. Aspects Appl. Biol. 118: 47–54.
- Bell, M.V. & Calladine, J. 2017. The decline of a population of farmland breeding waders: a twenty-five-year case study. Bird Study 64: 264–273.
- Bogaart, P., Van der Loo, M. & Pannekoek, J. 2018. rtrim: trends and indices for monitoring data. https://cran.rproject.org/package=rtrim.
- Calladine, J., Díaz, M., Reino, L., Jardine, D. & Wilson, M. 2018. Plantations of non-native tree species. In G. Mikusinski, J. M. Roberge & R. J. Fuller. (ed) Ecology and Conservation of Forest Birds, 350–386. Cambridge University Press, Cambridge.
- Calladine, J., Humphreys, E.M. & Boyle, J. 2015. Changes in breeding wader populations of the Uist machair between 1983 and 2014. Scott. Birds 35: 207–215.
- Calladine, J., Humphreys, E.M., Gilbert, L., Furness, R.W., Robinson, R.A., Fuller, R.J., Littlewood, N.A., Pakeman, R.J., Ferguson, J. & Thompson, C. 2017. Continuing influences of introduced Hedgehogs Erinaceus europaeus as a predator of wader (Charadrii) eggs four decades after their release on the Outer Hebrides, Scotland. Biol. Invasions 19: 1981–1987.
- Calladine, J., Pakeman, R.J., Humphreys, E., Huband, S. & Fuller, R.J. 2014. Changes in breeding wader assemblages, vegetation and land use within machair environments over three decades. Bird Study 61: 287–300.
- Darvill, B., Harris, S.J., Martay, B., Wilson, M. & Gillings, S. 2020. Delivering robust population trends for Scotland’s widespread breeding birds. Scott. Birds 40: 297–304.
- Dawson, A., Napier, I., Davies, N. & Mcllveny, J. 2011. A review of SOTEAG monitoring data and other long-term environmental series from Shetland. https://www.nafc.uhi.ac.uk/t4-media/one-web/nafc/research/document/environment/review-of-SOTEAG-monitoring-data.pdf.
- Donald, P.F. & Morris, T.J. 2005. Saving the Skylark. Br. Birds 98: 570–578.
- Douglas, D.J.T., Bellamy, P.E., Stephen, L.S., Pearce-Higgins, J.W., Wilson, J.D. Grant, M.C., & Fuller R. 2014. Upland land use predicts population decline in a globally near-threatened wader. J. Appl. Ecol. 51: 194–203.
- Eaton, M., Aebischer, N., Brown, A., Hearn, R., Lock, L., Musgrove, A., Noble, D., Stroud, D. & Gregory, R. 2015. Birds of Conservation Concern 4: the population status of birds in the UK, Channel Islands and Isle of Man. Br. Birds 108: 708–746.
- Ellis, P. 2004. Living Shetland Biodiversity Action Plan May, 2004. Species action plan “Skylark”. https://fliphtml5.com/zyki/xotr/basic.
- Evans, K.L. 2004. The potential for interactions between predation and habitat change to cause population declines of farmland birds. Ibis 146: 1–13.
- Fletcher, K., Aebischer, N.J., Baines, D., Foster, R. & Hoodless, A.N. 2010. Changes in breeding success and abundance of ground-nesting moorland birds in relation to the experimental deployment of legal predator control. J. Appl. Ecol. 47: 263–272.
- Franks, S.E., Douglas, D.J.T., Gillings, S. & Pearce-Higgins, J.W. 2017. Environmental correlates of breeding abundance and population change of Eurasian Curlew Numenius arquata in Britain. Bird Study 64: 393–409.
- Fraser, E.J., Lambin, X., McDonald, R.A. & Redpath, S.M. 2015. Stoat (Mustela erminea) on the Orkney Islands – assessing risks to native species. Scottish Natural Heritage Commissioned Report 871.
- Freeman, S.N., Noble, D.G., Newson, S.E. & Baillie, S.R. 2007. Modelling population changes using data from different surveys: the Common Birds Census and the Breeding Bird Survey. Bird Study 54: 61–72.
- Fuller, R.J., Gregory, R.D., Gibbons, D.W., Marchant, J.H., Wilson, J.D., Baillie, S.R. & Carter, N. 1995. Population declines and range contractions among lowland farmland birds in Britain. Conserv. Biol. 9: 1425–1441.
- Furness, R.W. & Greenwood, J.J.D. 1993.Birds as Monitors of Environmental Change. (R. W. Furness & J. J. D. Greenwood (eds)). Chapman & Hall, London.
- Grant, M.C., Chambers, R.E. & Evans, P.R. 1992. The effects of re-seeding heathland on breeding Whimbrel Numenius phaeopus in Shetland. III. Habitat use by broods. J. Appl. Ecol. 29: 516–523.
- Grant, M.C., Orsman, C., Easton, J., Lodge, C., Smith, M., Thompson, G., Rodwell, S. & Moore, N. 1999. Breeding success and causes of breeding failure of curlew Numenius arquata in Northern Ireland. J. Appl. Ecol. 36: 59–74.
- Hancock, M.H., Klein, D. & Cowie, N.R. 2020. Guild-level responses by mammalian predators to afforestation and subsequent restoration in a formerly treeless peatland landscape. Restor. Ecol. 28: 1113–1123.
- Harris, S.J., Massimino, D., Balmer, D.E., Eaton, M.A., Noble, D.G., Pearce-Higgins, J.W., Woodcock, P. & Gillings, S. 2020. The Breeding Bird Survey 2019. BTO Research Report 726. British Trust for Ornithology, Thetford.
- Holling, M. & the Rare Breeding Birds Panel, 2019. Rare breeding birds in the UK in 2017. Br. Birds 112: 697–772.
- Hoodless, A.N., Inglis, J.G. & Baines, D. 2006. Effects of weather and timing on counts of breeding Snipe Gallinago gallinago. Bird Study 53: 205–212.
- Jackson, D.B., Fuller, R.J. & Campbell, S.T. 2004. Long-term population changes among breeding shorebirds in the Outer Hebrides, Scotland, in relation to introduced Hedgehogs. Biol. Conserv. 117: 151–166.
- Kämpfer, S. & Fartmann, T. 2019. Breeding populations of a declining farmland bird are dependent on a burrowing, herbivorous ecosystem engineer. Ecol. Eng. 140: 105592.
- Macdonald, M.A. & Bolton, M. 2008. Predation on wader nests in Europe. Ibis 150: 54–73.
- Martin, P. 2005. Review of cereal growing on Shetland. http://croftingyear.org.uk/userfiles/file/cereals/shetland-cereals-report-apr-2015.pdf.
- Massey, K., Cosgrove, P., Massey, F., Jackson, D. & Chapman, M. 2016. Habitat characteristics of breeding Eurasian Whimbrel Numenius phaeopus on mainland Shetland, Scotland, UK. Bird Study 63: 500–508.
- Massimino, D., Johnston, A. & Pearce-Higgins, J.W. 2015. The geographical range of British birds expands during 15 years of warming. Bird Study 62: 523–534.
- McCallum, H.M., Park, K.J., O'Brien, M.G., Gimona, A., Poggio, L. & Wilson, J.D. 2015. Soil pH and organic matter content add explanatory power to Northern Lapwing Vanellus vanellus distribution models and suggest soil amendment as a conservation measure on upland farmland. Ibis 157: 677–687. doi: 10.1111/ibi.12286
- McCallum, H.M., Wilson, J.D., Beaumont, D., Sheldon, R., O'Brien, M.G. & Park, K.J. 2016. A role for liming as a conservation intervention? Earthworm abundance is associated with higher soil pH and foraging activity of a threatened shorebird in upland grasslands. Agriculture, Ecosystems & Environment 223: 182–189.
- Mitchell, P.I., Newton, S.F., Ratcliffe, N. & Dunn, T.E. 2004. Seabird Populations of Britain and Ireland. T. & AD Poyser, London.
- Morrison, C.A., Robinson, R.A., Clark, J.A. & Gill, J.A. 2010. Spatial and temporal variation in population trends in a long-distance migratory bird. Divers. Distrib. 16: 620–627.
- Morrison, C.A., Robinson, R.A., Clark, J.A., Leech, D.I. & Gill, J.A. 2015. Season-long consequences of shifts in timing of breeding for productivity in Willow Warblers, Phylloscopus trochilus. Bird Study 62: 161–169.
- O'Brien, M. & Smith, K. 1992. Changes in the status of waders breeding on wet lowland grasslands in England and Wales between 1982 and 1989. Bird Study 39: 165–176.
- Ockendon, N., Hewson, C.M., Johnston, A. & Atkinson, P.W. 2012. Declines in Afro-Palearctic migrant birds are linked to bioclimatic wintering zone, possibly via constraints in arrival time advancement. Bird Study 59: 111–125.
- Pannekoek, J. & van Strien, A. 2006a. TRIM Version 3.54 (Trends and Indices for Monitoring Data). Statistics Netherlands, Voorburg.
- Pannekoek, J. & van Strien, A. 2006b. [26 May 2020]. Frequently Asked Questions Updated 2006 (TRIM Version 3.51). Statistics Netherlands, Voorburg.
- Roodbergen, M., van der Werf, B. & Hotker, H. 2012. Revealing the contributions of reproduction and survival to the Europe-wide decline in meadow birds: review and meta-analysis. J. Ornithol. 153: 53–74.
- Roos, S., Smart, J., Gibbons, D.W. & Wilson, J.D. 2018. A review of predation as a limiting factor for bird populations in mesopredator-rich landscapes: a case study of the UK. Biol. Rev. 93: 1915–1937.
- Shetland Bird Report. 2019. Shetland Bird Club. (M. Pennington (ed)). Shetland.
- Shetland Bird Report. 2020. Shetland Bird Club. (M. Pennington (ed)). Shetland.
- Stanbury, A., Thomas, S., Aegerter, J., Brown, A., Bullock, D., Eaton, M. & Oppel, S. 2017. Prioritising islands in the United Kingdom and crown dependencies for the eradication of invasive alien vertebrates and rodent biosecurity. Eur. J. Wildl. Res. 63: 31.