Abstract
A new lignitised, slightly compressed pollen organ, Erdtmanitheca portucalensis, with affinities to extinct Erdtmanithecales from the Early Cretaceous (Aptian–Albian) of Vale de Água (Lusitanian Basin, western Portugal), is described. The pollen organ is composed of loosely arranged microsporophylls radiating from a central core. The estimated number of microsporophylls is about 100–150. The microsporophylls are sessile and ellipsoidal to barrel-shaped with a flattened or slightly apically depression containing about ten narrow sporangia. The sporangia enclose abundant well-preserved pollen grains of Eucommiidites-type. Pollen grains found in situ are elliptical in equatorial outline, about 16.0–27.2 μm long and 11.9–16.4 μm wide. The main (distal) colpus is long with expanded rounded ends. It is flanked by two subsidiary colpi in an almost equatorial position. The surface of the pollen wall is psilate and occasionally punctate. The ektexine is composed of a distinct tectum, granular infratectal layer and a thin foot layer. The endexine is thick and laminar. The new Early Cretaceous Portuguese pollen-organ is similar in several respects to that of Erdtmanitheca texensis described from the Late Cretaceous of Texas, USA. The new fossil species further documents the importance of the Bennettitales-Erdtmanithecales-Gnetales group in the Early Cretaceous floras of Portugal extending the stratigraphic and geographical distribution of the genus with regard to systematic and phylogenetic significance of the Eucommiidites-producing plants that may have been co-occurring with the Early Cretaceous diversification of angiosperms. It is ascertained that perforate tectum occurs in pollen grains with a well-developed foot layer as well as in pollen grains in which a foot layer is poorly developed or lacking, and that pollen features do not support a separation of the Erdtmanithecales seeds and pollen organs.
Lower Cretaceous strata from the Estremadura and Beira Litoral regions (Lusitanian Basin, western Portugal) have yielded rich fossil floras including exquisitely preserved angiosperm flowers. These floras are pivotal for understanding vegetational structure and changes in the region during the earliest phases of angiosperm diversification. Previous studies of plant fossils from this region focused on macrofossil floras mostly containing leaves (Heer, Citation1881; Saporta, Citation1894; Romariz, Citation1946; Teixeira, Citation1948, 1950, Citation1952) and microfossil floras containing dispersed palynomorphs (Hasenboehler, Citation1981; Pais & Reyre, Citation1981; Pais & Trincão, Citation1983; Leereveld et al., Citation1989; Trincão, Citation1990; Heimhofer et al., Citation2007). More recently, the discovery and study of well-preserved mesofossil floras with abundant angiosperm reproductive organs such as inflorescences, flowers, stamens, fruits and seeds, has greatly increased our knowledge on the reproductive biology and organisation of Cretaceous angiosperms (Friis et al., Citation1994, Citation1997, 1999, Citation2000, Citation2001, Citation2004, Citation2006). The same mesofossil floras also include well-preserved and informative remains of other plant groups. Most notably, the recognition of a new complex of seeds with a unique seed structure otherwise only found in Bennettitales, Erdtmanithecaes and Gnetales (the BEG group of Friis et al., Citation2007) is of importance for understanding the total vegetation structure and is of potentially great importance for understanding phylogenetic relationships among seed plants. Some of the seeds are closely related to extant Ephedra L. of the Gnetales (Rydin et al., Citation2004, Citation2006) and one seed type, Erdtmanispermum juncalense Mendes, Friis & Pais can be placed in the extinct order Erdtmanithecales (Mendes et al., Citation2008a ). The Erdtmanithecales are also represented in the Early Cretaceous floras of Portugal by the microsporangiate organ Eucommiitheca hirsuta Friis & Pedersen (Friis & Pedersen, Citation1996). Other seeds are of uncertain position in the BEG group and have been assigned to extinct genera such as Buarcospermum Friis, Pedersen & Crane, Lignierispermum Friis, Pedersen & Crane and Lobospermum Friis, Pedersen & Crane (Friis et al., Citation2007, 2009). A further seed type, Raunsgaardispermum Mendes, Pais & Friis, combines distinct ephedroid seed characters with bennettitalean and erdtmanithecalean pollen features (Mendes et al., Citation2008b ).
In this article, we describe a new Erdtmanithecales microsporangiate organ from Portugal that we assign to the genus Erdtmanitheca Pedersen, Crane & Friis first described from the Late Cretaceous (Early Cenomanian) of Texas, USA (Pedersen et al., Citation1989). This new finding further documents the importance of the BEG group in the Early Cretaceous floras of Portugal and adds to the diversity of the group.
Materials and methods
The fossil pollen organs described here were extracted from sediment samples collected from the Vale Farelo opencast clay pit complex, near the small village of Vale de Água (39° 37' 13.1'' N; 08° 51' 57.2'' W), in the Estremadura region, western Portugal ().
Figure 1. Map illustrating the studied area in western Portugal (square). The approximate location of the Vale Farelo clay pit complex where the specimens were collected is indicated by an asterisk.
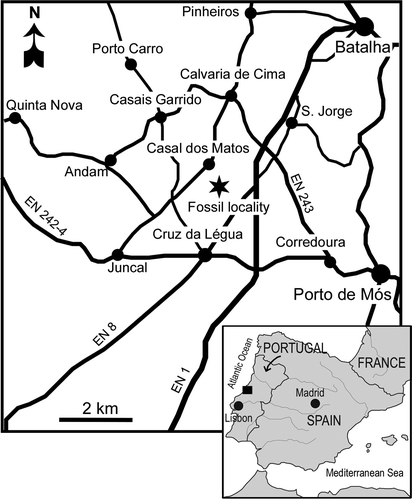
The fossil specimens were isolated from dark-grey mudstone deposits previously assigned to the “Complexos Gresosos da Nazaré e Cós-Juncal” (Carta Geológica de Portugal, Folha 26-B Alcobaça, Zbyszewski et al., Citation1961) and subsequently included in the Figueira da Foz Formation (Dinis, Citation1999, Citation2001). According to Dinis (pers. comm., 2009) the fossil-bearing horizons belong to the Famalicão Member, which is the basal part of the Figueira da Foz Formation (Dinis, Citation1999, Citation2001; Rey et al., Citation2006).
A late Aptian age was suggested by Dinis et al. (Citation2002) for the Figueira da Foz lower boundary, using mainly macrofossils, pollen and spores as well as sedimentological and lithofacial correlations (Teixeira, Citation1950; Dinis & Trincão, Citation1991; Friis et al., Citation1999; Heimhofer et al., Citation2005), and Rey et al. (Citation2006) indicated a late Aptian to early Albian age for the Famalicão Member of the Figueira da Foz Formation.
In general, the fossils are lignitised with the original three-dimensional form slightly compressed. Bulk matrix samples were dried in the laboratory, disaggregated in water and sieved using a shower through a 125 μm screen. The fossils described in this paper were isolated from one sediment sample (sample VF-141) collected by M. M. Mendes and J. L. Dinis in 2008. The species diversity is low, but the sample yielded some seeds, twigs and leaves of conifers, as well as several angiosperm fossils such as possible nymphaealean seeds, seeds assignable to the extinct genus Anacostia Friis, Crane and Pedersen, as well as other seeds and a variety of endocarps. A preliminary palynological analysis of the same sample has documented a palynoflora mainly characterised by fern spores, gymnosperm pollen, including pollen grains of Eucommiidites-type, and a few angiosperm pollen grains.
The fossils were cleaned in hydrofluoric (40% HF) and hydrochloric (10% HCl) acids, thoroughly rinsed in water and air dried. The specimens were observed with a Nikon SMZ800 stereomicroscope. The specimens selected for scanning electron microscopy were mounted on polished aluminium stubs and coated with gold for 60 seconds in a sputter coater, and examined using a Hitachi Field S-4300 scanning electron microscope (FE-SEM) at 2 kV, at the Swedish Museum of Natural History, Stockholm. Pollen for TEM was removed from specimen P0185 after SEM, embedded in Epon (TAAB 812) after dehydration in a grade of acetone and acetone/Epon solutions. Sections were cut with a diamond knife on a RMC MT7 Ultramicrotome, triple stained with lead citrate and uranyl acetate (following Daddow, Citation1983), and examined using a Leo 912 AB Omega transmission electron microscope (with a high voltage of 80 kV) at the Swedish Museum of Natural History. Pollen grains for light microscopy (LM) were removed from specimen P0185 and glycerine was used for mounting permanent slides. LM studies were performed using an ECLIPSE E600 Nikon microscope and specimens were photographed using a Nikon DS-L2 digital camera.
Dimensions of the pollen organs were measured under the stereomicroscope and the dimensions of the pollen grains embedded in glycerine were measured using LM; dry pollen grains were measured from SEM-micrographs, and the layers of the pollen wall were measured on TEM-micrographs.
LM, SEM- and TEM-micrographs were improved using Photoshop software to enhance contrast and remove stain from the background.
The specimens formally described in this paper are deposited in the Earth Sciences Department, Technology and Sciences College, New University of Lisbon, Portugal.
Results
Systematic description
Spermatophyta
Erdtmanithecales Friis & Pedersen, Citation1996
Erdtmanithecaceae Friis & Pedersen, Citation1996
Erdtmanitheca Pedersen, Crane & Friis, Citation1989
Erdtmanitheca portucalensis Mendes, Pais, Pedersen & Friis sp. nov. ()
Figure 2. Erdtmanitheca portucalensis sp. nov. from the Early Cretaceous of Portugal. LM-micrographs of Eucommiidites-type pollen grains isolated from the paratype (P0185). Scale bar – 10 μm.
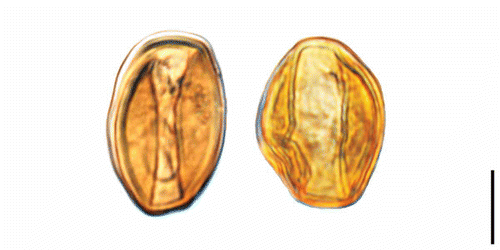
Figure 3. Erdtmanitheca portucalensis sp. nov. from the Early Cretaceous of Portugal. SEM-micrographs of pollen organs. A. Holotype showing sessile microsporophylls loosely arranged in a radial pattern from a central core (P0186). B, C. Detail of holotype showing the ellipsoidal to barrel-shaped microsporophyll (P0186). D. Detail of microsporophyll showing sporangia with pollen grains (P0186). E. Fragment from paratype showing microsporophyll arranged in radial pattern (P0185). F. Detail of paratype showing pollen grains in elongated pollen sacs (P0185). Scale bars – 1 mm (A, E); 500 μm (B); 250 μm (C); 50 μm (D); 100 μm (F).

Figure 4. Erdtmanitheca portucalensis sp. nov. from the Early Cretaceous of Portugal. SEM-micrographs of Eucommiidites-type pollen grains isolated from holotype and paratype. A, B & D. Group of pollen grains isolated from the paratype showing well-defined distal colpus flanked by two subsidiary colpi in the equatorial plane (P0185). C. Orbicules (Ubisch bodies) spaced on the sporangium wall (P0185). E. Pollen grain isolated from the holotype showing wall with perforations on the proximal surface between the lateral colpi. F–I. Pollen grains isolated from paratype showing psilate wall surface and margins of apertures (P0185). J. Pollen grain isolated from the holotype showing perforations along the margins of the main colpus (P0186). K. Pollen grain, isolated from the holotype, in equatorial view showing colpi (P0186). L. Fragmented pollen grain isolated from the holotype showing wall stratification and details of aperture structure (P0186). Scale bars – 20 μm (A, J); 25 μm (B, D); 5 μm (C, L, M); 10 μm (E–I).
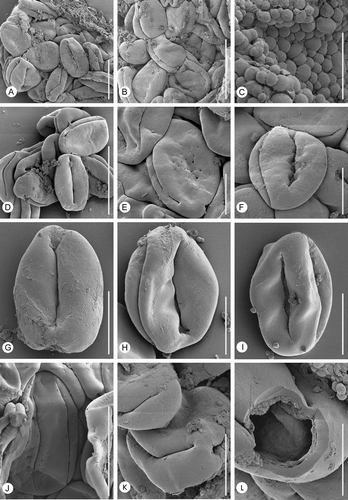
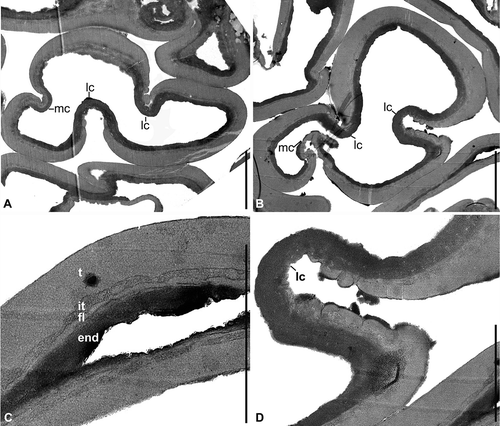
Figure 5. Erdtmanitheca portucalensis sp. nov. from the Early Cretaceous of Portugal. Eucommiidites-type pollen grains isolated from the paratype (P0185). TEM-micrographs of pollen wall ultrastructure. A, B. Transverse sections of pollen grains perpendicular to the main colpus (mc) and lateral colpi (lc). C. Detail of grain in showing stratification of pollen wall with thick homogenous tectum (t), thin granular infratectal layer (it), foot layer (fl) and thick laminar endexine (end). D. Detail of showing apertural region of one lateral colpus enlarged with thinning of tectum towards the aperture and tectal verrucae on the margins to the aperture. Scale bars – 3 μm (A, B); 2 μm (C, D).
Specific diagnosis
Pollen organ bearing loosely arranged microsporophylls radiating from a central core. Microsporophyll ellipsoidal to barrel-shaped with a flattened or slightly depressed peltate apical head; each microsporophyll with about ten narrow sporangia in a radial arrangement. Pollen of Eucommiidites-type with long main (distal) colpus flanked by two lateral colpi in almost equatorial position; tectum psilate, occasionally with small puncta proximally. Tectum thick, infratectal layer and foot layer thin. Pollen length (L) to width (W) ratio about 3:2.
Derivation of the specific name
The specific epithet portucalensis is derived from Portugal, where the fossils were collected.
Dimensions
Pollen organs about 1.9 mm long and 1.97–2.65 mm wide. Pollen grains about 20.5–21.8 μm long and 14.5–16.1 μm wide.
Holotype
P0186 (Vale de Água – Vale Farelo clay pit, sample VF-141) (–D, 4E, J–L).
Paratype
P0185 (Vale de Água – Vale Farelo clay pit, sample VF-141) (, 3E–F, 4A–D, F–I).
Type locality
Vale de Água – Vale Farelo opencast clay pit complex, Estremadura region, western Portugal (39° 37' 13.1'' N; 08° 51' 57.2'' W).
Stratigraphic position
Famalicão Member of the Figueira da Foz Formation.
Age
Early Cretaceous (late Aptian–early Albian).
Description and remarks on the species
The material studied here includes two dispersed lignitised and slightly compressed pollen organs. None of the specimens are complete and there are no remains of stalk or dehiscence scar. The shape of the fragments indicates that the pollen organ was originally spherical (best seen in specimen P0185), consisting of numerous microsporophylls borne in a radial arrangement around a central receptacle. The specimens are about 1.89 mm long and 1.97–2.65 mm wide (–F). The most complete specimen consists of about 20 microsporophylls and the other specimen consists of six microsporophylls. The estimated diameter of the pollen organs is about 2.0 mm and the estimated number of microsporophylls in each pollen organ is about 100–150. The microsporophylls are sessile and loosely arranged in a radial pattern from the central receptacle, separate from each other for most of their length. The microsporophylls are ellipsoidal to barrel-shaped, about 1.1 mm long and 0.6 mm wide, with a rounded peltate apical head that is flattened or with a shallow central depression.
The microsporophylls are apparently synangiate with about ten radially arranged sporangia embedded in a common tissue, but the number of sporangia in each microsporophyll and the exact organisation of the sporangia on the microsporophyll is unclear. Dehiscence is by longitudinal slits that extend for almost the full length of the sporangia from the base to the margin of the apical head (–D).
In situ pollen is of the Eucommiidites-type, elliptical in equatorial outline, rounded at both ends, and about 16 (20.5) 27.2 μm long and 11.9 (13.9) 16.4 μm wide (15 specimens measured). Length to width ratio is about 3:2. The pollen wall is typically invaginated in the aperture regions (, 4A, B, D–I) and details of the aperture region and membrane are only seen in a few specimens in SEM, but can also be observed in TEM. The invagination also indicates that the pollen grains were probably more spherical in hydrated, living state. The aperture arrangement is distinct. There is a well-defined distal colpus, with slightly expanded rounded ends (, B, D–I). The distal colpus is flanked by two lateral colpi placed more or less in the equatorial plane. They are slit-like with pointed ends and longer than the distal colpus and almost meet at either end (, B, D), although none of them merge to form a ring colpus. Margins of the apertures are irregular and the aperture membrane is verrucate-granular (, H, I, 5A, B, D).
The pollen wall is tectate and psilate. In some specimens, small holes (puncta), 0.1–0.4 μm in diameter, occur on the proximal surface between the lateral colpi (, E) or occasionally along the margins of the main colpus (, J). The puncta were not observed in the TEM-sections and were apparently not continuous through the tectum. Some pollen are slightly contracted over the proximal pole to form a faint regular pattern. We ascribe the differences in the appearance of the tectum surface to differences in preservation, where increased degradation of the pollen wall causes more prominent holes.
The pollen wall is about 1 μm thick in non-apertural regions and thinner over the apertures. Pollen wall stratification and details of aperture structure were observed by SEM in a fragmented pollen grain () and by TEM in ultrathin sections (). The ektexine (outer layer, lighter staining in ultrathin sections) is composed of a distinct tectum, a thin granular infratectal layer, and a thin foot layer. The tectum is homogenous, about 0.60 μm thick in non-apertural regions, but gradually thinning in the apertural regions and lacking over the aperture (A, B, D). It is granular to verrucate on the margins of the lateral colpi (D) and apparently smooth on the margins of the main colpus (A). The granular infratectal layer is about 0.1 μm thick. It is more distinct in SEM () than in TEM (C) and may be compressed during fossilisation. The foot layer is of approximately the same thickness as the infratectal layer, about 0.1 μm. It is also homogenous as the tectum, but with uneven outer and inner surface (C).
The endexine (inner layer, darker staining in ultrathin sections) is about 0.25 μm thick, laminar, and of more or less even thickness in apertural and non-apertural regions ().
Small orbicules (Ubisch bodies), about 0.5–1.5 μm in diameter, and with an almost smooth surface are densely spaced on the inner surface of the sporangia wall and scattered on the surface of the pollen grains (). They are solid in TEM.
Discussion
Eucommiidites and Erdtmanithecales
The organisation of the fossil pollen organs described here and their distinctive Eucommiidites-type pollen unequivocally place the fossils in the Erdtmanithecales, an extinct order established to accommodate fossil seeds and microsporangiate organs containing Eucommiidites-type pollen (Friis & Pedersen, Citation1996).
The pollen genus Eucommiidites Erdtman was established on the base of dispersed pollen from the Early Jurassic of Pålsjö, Sweden (Erdtman, Citation1948). The pollen is characterised by an unusual aperture configuration typically with a main colpus in a distal position flanked either by two subsidiary colpi or a single ring colpus in an almost equatorial position (Erdtman, Citation1948; Couper, Citation1956, Citation1958). Pollen grains of Eucommiidites-type are widespread in palynofloras in the Northern Hemisphere, from the Late Triassic to the Late Cretaceous (Erdtman, Citation1948; Kuyl et al., Citation1955; Couper, Citation1956, Citation1958; Groot & Penny, Citation1960; Hughes, Citation1961, Citation1994; Brenner, Citation1963, Citation1967; Reymanówna, Citation1968; Scheuring, Citation1970, Citation1978; Pedersen et al., Citation1989; Batten & Dutta, Citation1997).
Eucommiidites pollen found in situ in seeds and microsporangiate structures have a distinct tectate pollen wall with a homogenous, sometimes punctate-perforate tectum and a granular infratectal layer (Pedersen et al., Citation1989; Friis & Pedersen, Citation1996; Tekleva et al., Citation2006). The perforations are prominent in the pollen found in the micropyles of Erdtmanispermum balticum Pedersen, Crane and Friis and are also present in the pollen of Erdtmanitheca texensis Pedersen, Crane and Friis, although the perforation is less pronounced. A punctate tectum was also documented for dispersed pollen grains assigned to Eucommiidites troedssonii Erdtman from the Early Jurassic of Grojec, Poland (Scheuring, Citation1978) and from the Hettangian–Sinemurian of Sose Bugt, Bornholm, Denmark (Batten & Dutta, Citation1997); perforation is particularly pronounced in the latter.
The infratectal layer is interspaced either between tectum and endexine (Erdtmanitheca texensis) or more common between tectum and foot layer (Erdtmanispermum balticum, Eucommitheca hirsuta and Bayeritheca hughesii Kvacek & Pacltová). The foot layer, when present, is thin (Eucommiitheca hirsuta) or thicker (Bayeritheca hughesii, Erdtmanispermum balticum) (Pedersen et al., Citation1989; Friis & Pedersen, Citation1996; Tekleva et al., Citation2006). A granular infratectal layer was also reported for Eucommiidites troedssonii from Sose Bugt (Batten & Dutta, Citation1997), but it is unclear from the description and figures whether the foot layer is present or not. The ultrastructure of Eucommiidites troedssonii from Grojec is illustrated by line drawings only and is unclear for the infratectal layers.
A variety of other grains assigned to the Eucommiidites complex by Tekleva et al. (Citation2006) based on comparable aperture arrangement have a distinctly different wall structure and cannot be included in Erdtmanithecales as currently circumscribed.
The pollen organs have peltate microsporophylls, each with several elongate sporangia. The microsporophylls are arranged on elongated axes or in spherical heads (Pedersen et al., Citation1989; Friis & Pedersen, Citation1996; Kvaček & Pacltová, Citation2001). Seeds assigned to the Erdtmanithecales are characterised by having a nucellus enclosed by two layers; a thin membranous integument that is extended into a long narrow micropyle and a hard three-valved outer seed envelope that completely encloses the integument except for the apical micropylar opening (Pedersen et al., Citation1989).
New studies of Erdtmanithecales and other Early Cretaceous seed plants concluded that Erdtmanithecales are most closely related to Bennettitales and Gnetales, and the term BEG (Bennettitales-Erdtmanithecales-Gnetales) group was coined for this constellation of seed plants (Friis et al., Citation2007, Citation2009). Four new genera, Buarcospermum, Lignierispermum, Lobospermum and Rugonella Friis, Pedersen and Crane, were assigned to this complex, but their more precise systematic affinity within the group was not resolved (Friis et al., Citation2009).
Rothwell et al. (Citation2009) recently questioned the Erdtmanithecales concept. In their studies of the phylogenetic position of Erdtmanithecales seeds and pollen organs were analysed separately based on differences in morphology and ultrastructure between pollen in situ in seed micropyles (tectum punctate, foot layer distinct) and pollen in situ in the pollen organs (tectum typically psilate, foot layer absent or indistinct). In this analysis the seeds were resolved as most closely related to the Gnetales and the pollen organs most closely related to the Bennettitales. The new fossil described here shows, however, a range in pollen wall structure that encompasses the structure observed for pollen in situ in the seeds and that for pollen in situ in the pollen organs. Several grains have a punctate tectum, although the holes are not continuous, and there is a distinct foot layer. Dispersed Eucommiidites pollen grains also show distinctly punctate tectum combined with the absence or poorly developed foot layer. This together with the otherwise very similar pollen wall structure of pollen in seeds and pollen in sporangia indicate that seeds and pollen organs were produced by the same kind of plants.
Comparison with other Eucommiidites producing pollen organs
Three genera and species of pollen organs with Eucommiidites pollen have been included in the Erdtmanithecales: Erdtmanitheca texensis (Late Cretaceous of Texas, USA), Eucommiitheca hirsuta (Early Cretaceous of Beira Litoral, Portugal) and Bayeritheca hugesii (Late Cretaceous of Bohemia, Czech Republic). These pollen organs are united by having sporangia arranged in a radial pattern on a peltate microsporophyll and by their characteristic Eucommiidites pollen. Pollen grains resembling Eucommiidites were also found in situ in cones of Hastyostrobus muirii Van Konijnenburg-van Cittert from the Middle Jurassic of Hasty Bank, Yorkshire, England. Although the wall ultrastructures of these grains (Tekleva et al., Citation2006) show some similarities to other Eucommiidites grains, the infratectal layer described by Tekleva et al. (Citation2006) as spongy appears alveolar rather than granular. The pollen cone is poorly preserved and there are no details of sporangia or their organisation on the axis (Van Konijnenburg-van Cittert, Citation1972) and further comparison with this fossil is currently not possible.
Eucommiitheca hirsuta and Bayeritheca hughesii are distinguished from the new Portuguese fossil in having microsporangiate units arranged along an elongate axis. In E. hirsuta, the arrangement of microsporophylls is opposite and decussate; in B. hughesii, the arrangement is less clear. Pollen grains in B. hughesii are smaller than in the new Portuguese fossils and the foot layer slightly thicker. Pollen grains observed in E. hirsuta are also smaller (15.0–20.0 μm × 10.0–12.0 μm in contrast to 16.0–27.2 μm × 11.9–16.4 μm in the new fossil).
Erdtmanitheca texensis has microsporophylls arranged in spherical heads radiating from a central receptacle very similar to the arrangement in the new Portuguese fossils. Further similarities between our new fossils and E. texensis are the synangiate nature of the microsporophylls with about ten narrow, elongate sporangia extending from the margin of the peltate apex to the base of the microsporophyll, and dehiscence of the sporangia by longitudinal slits. A generic assignment of the Portuguese fossils to Erdtmanitheca is therefore well-supported. The new fossils differ, however, from E. texensis in several details and are therefore assigned to a new species, E. portucalensis. Most significantly, the Portuguese fossils have fewer microsporophylls (about 100–150 in contrast to about 300 in E. texensis) and the microsporophylls are more loosely arranged. Although none of the specimens is preserved completely, their organisation strongly suggests that they were spherical heads as in E. texensis. The mode of attachment on the plant is unknown for both the new fossil and for E. texensis.
The pollen grains of Erdtmanitheca texensis and E. portucalensis are very similar in general shape and size and in the distribution of apertures. The only difference is that the lateral furrows in E. portucalensis are slightly longer. Tectum perforations are more distinct in some grains of E. portucalensis, but not in all, and the variations observed in pollen wall surface between E. portucalensis and E. texensis are probably mostly due to variation in preservation. In pollen wall ultrastructure the two species are closely similar and both have distinct orbicules. There are, however, minor differences that also support assignment of the two fossils to different species. In E. texensis the pollen wall is markedly thinner on the proximal side, while in E. portucalensis does not show such thinning. Also in E. texensis the granular infratectal layer rests directly on the endexine, and foot layer is lacking, while in E. portucalensis the granular infratectal layer is interspaced between the tectum and foot layer with a distinct delimitation to both.
Erdtmanithecales in Portugal
Pollen grains of Eucommiidites-type have been reported from many Early Cretaceous palynofloras of western Portugal, such as Lagosteiros, Forte de Santa Susana, Forte da Crismina and S. Julião beach ranging in age from Berriasian to Albian (Trincão, Citation1990).
Recently, Eucommiidites pollen was also recovered in the Lower Cretaceous strata of the Nossa Senhora da Luz clay pit complex, the Juncal locality, and in the Vale Farelo clay pit complex, the Vale de Água locality, ranging in age from the Aptian to Albian (Mendes, work in progress). None of the dispersed grains was studied in greater detail using SEM or TEM and they may not all belong to the Erdtmanithecales. The presence of Erdtmanithecales in Portugal is, however, well-established for the Early Cretaceous.
From the Lusitanian Basin in western Portugal two different kinds of pollen organs with Eucommiidites pollen in situ are currently known from the younger part of the Early Cretaceous, including Eucommiitheca hirsuta (Friis & Pedersen, Citation1996) and Erdtmanitheca portucalensis (this work), and one kind of seeds, Erdtmanispermum juncalense (Mendes et al., Citation2008a ), has been reported from the earliest Cretaceous. No pollen organs were found associated with Erdtmanispermum juncalense and no pollen was found inside the micropyles of the seeds. However, the assignment of Erdtmanispermum juncalense to Erdtmanithecales is ascertained by the close similarity to the coeval Erdtmanispermum balticum from the earliest Cretaceous of Bornholm (Denmark) that has Eucommiidites pollen in situ (Pedersen et al., Citation1989).
Seeds that could be associated with Erdtmanitheca portucalensis have not yet been found from the Vale Farelo clay pit complex – Vale de Água locality, and no other seeds of the BEG group were identified from this locality.
Seeds of Raunsgardispermum from the Early Cretaceous of Portugal combine ephedroid seed characters with bennettitalean and erdtmanithecalean pollen features (Mendes et al., Citation2008b ) and are also assignable to the BEG group. The pollen grains are similar to those of Erdtmanitheca portucalensis in their psilate-perforate tectum, but they differ in being monocolpate and the perforation of the tectum is denser and occasionally coarser, although the size of the perforations varies from grain to grain and may also for this taxon be linked to preservation.
Conclusions
The new pollen organ Erdtmanitheca portucalensis and its in situ Eucommiidites-type pollen described here is clearly assignable to the extinct order Erdtmanithecales. The Portuguese Early Cretaceous pollen organ shares key characters with the Late Cretaceous Erdtmanitheca texensis. The new fossil findings provide more knowledge on the morphological variation in Eucommiidites-pollen-producing plants. This finding extends the geographical and stratigraphic range of Erdtmanitheca that was previously known only from the Early Cenomanian (Late Cretaceous) of eastern North America. The new finding also provides additional support for the presence of a diverse complex of plants, e.g. the Bennettitales-Erdtmanithecales-Gnetales group, in the Early Cretaceous vegetation co-occurring with the early diverging angiosperms. The presence of psilate pollen with more coarsely perforate grains in Erdtmanitheca portucalensis shows that this feature may be related to preservation and may not be a useful systematic character.
It is also clear that a punctate tectum occurs both in pollen with a distinct foot layer and in pollen with a foot layer that is poorly developed or lacking and that these pollen features (punctate tectum/distinct foot layer contra psilate tectum/absent or indistinct foot layer) cannot be used to place the Erdtmanithecales pollen organs and seeds in separate clades.
Acknowledgements
We thank Jorge Dinis, Coimbra, for valuable assistance in field work and for information on the geology of the Figueira da Foz Formation. We also thank Eduarda Ferreira, Caparica, for technical assistance in the laboratory, Yvonne Arremo, Stockholm, for technical assistance with the scanning electron microscope, Carina Svensson and Lena M. Gustavsson, Stockholm, for technical assistance with sections for ultrastructural studies and with the TEM investigations. Patrick Herendeen, Chicago, and Johanna van Konijnenburg-van Cittert, Leiden, are thanked for helpful comments on the manuscript. This work was supported by a fellowship from the “Fundação para a Ciência e a Tecnologia” (SFRH/BD/24849/2005) to Mário Miguel Mendes and a grant from the Swedish Natural Science Research Foundation (VR) to Else Marie Friis.
Related Research Data
References
- Batten , D. J. and Dutta , R. J. 1997 . Ultrastructure of exine of gymnospermous pollen from Jurassic and basal Cretaceous deposits in Northwest Europe and implications for botanical relationships . Rev. Palaeobot. Palynol. , 99 : 25 – 54 .
- Brenner , G. J. 1963 . The spores and pollen of the Potomac Group of Maryland , Vol. 27 , Baltimore : Md. Dep. Geol. Mines Water Resour. Bull .
- Brenner , G. J. 1967 . The gymnospermous affinity of Eucommiidites Erdtman, 1948 . Rev. Palaeobot. Palynol. , 5 : 123 – 127 .
- Couper , R. A. 1956 . Evidence of a possible gymnospermous affinity for Tricolpites troedssonii Erdtman . New Phytol. , 55 : 280 – 285 .
- Couper , R. A. 1958 . British Mesozoic microspores and pollen grains. A systematic and stratigraphic study . Palaeontographica B , 103 : 75 – 179 .
- Daddow , L. Y. M. 1983 . A double lead stain method for enhancing contrast of ultra thin sections in electron microscopy: A modified multiple staining technique . J. Microsc. , 129 : 147 – 153 .
- Dinis , J. L. 1999 . Estratigrafia e sedimentologia da formação da Figueira da Foz. Aptiano a Cenomaniano do sector norte da Bacia Lusitânica , Coimbra : Univ. Coimbra. Ph.D. Diss .
- Dinis , J. L. 2001 . Definição da Formação da Figueira da Foz – Aptiano a Cenomaniano do sector central da margem oeste ibérica . Comun. Inst. Geol. Min. , 88 : 127 – 160 .
- Dinis , J. , Rey , J. and de Graciansky , P. -C . 2002 . Le Bassin Lusitanien (Portugal) à l'Aptien supérieur-Albien: Organisation séquentielle, proposition de corrélations, évolution . C. R. Geoscience , 334 : 757 – 764 .
- Dinis , J. L. and Trincão , P. 1991 . Controlos deposicionais e biostratigrafia da base dos “grés belasianos” (Aptiano, Bacia Lusitaniana) . Comunic. Serv. Geol. Portugal , 77 : 89 – 102 .
- Erdtman , G. 1948 . Did dicotyledonous plants exist in early Jurassic time? . GFF , 70 : 265 – 271 .
- Friis , E. M. , Crane , P. R. and Pedersen , K. R. 1997 . Anacostia, a new basal angiosperm from the Early Cretaceous of North America and Portugal with trichotomocolpate/monocolpate pollen . Grana , 36 : 225 – 244 .
- Friis , E. M. , Crane , P. R. , Pedersen , K. R. , Bengtson , S. , Donoghue , P. C. J. , Grimm , G. W. and Stampanoni , M. 2007 . Phase-contrast X-ray microtomography links Cretaceous seeds with Gnetales and Bennettitales . Nature , 450 : 549 – 552 .
- Friis , E. M. and Pedersen , K. R. 1996 . Eucommiitheca hirsuta, a new pollen organ with Eucommiidites pollen from the Early Cretaceous of Portugal . Grana , 35 : 104 – 112 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 1994 . Angiosperm floral structures from the Early Cretaceous of Portugal . Plant Syst. Evol. , 8 : 31 – 49 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 1999 . Early angiosperm diversification: The diversity of pollen associated with angiosperm reproductive structures in Early Cretaceous floras from Portugal . Ann. Mo. Bot. Gard. , 86 : 259 – 296 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 2000 . Fossil floral structures of a basal angiosperm with monocolpate, reticulate-acolumellate pollen from the Early Cretaceous of Portugal . Grana , 39 : 226 – 239 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 2001 . Fossil evidence of water lilies (Nymphaeales) in the Early Cretaceous . Nature , 410 : 357 – 360 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 2004 . Araceae from the Early Cretaceous of Portugal: Evidence on the emergence of monocotyledons . Proc. Natl. Acad. Sci. , 101 : 16565 – 16570 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 2006 . Cretaceous angiosperm flowers: Innovation and evolution in plant reproduction . Palaeogeogr. Palaeoclimatol. Palaeoecol. , 232 : 251 – 293 .
- Friis , E. M. , Pedersen , K. R. and Crane , P. R. 2009 . Early Cretaceous mesofossils from Portugal and eastern North America related to the Bennettitales-Erdtmanithecales-Gnetales group . Am. J. Bot. , 96 : 252 – 283 .
- Groot , J. J. and Penny , J. S. 1960 . Plant microfossils and age of nonmarine Cretaceous sediments of Maryland and Delaware . Micropalaeontology , 6 : 225 – 236 .
- Hasenboehler , B. 1981 . Étude paléobotanique et palynologique de l'Albien et du Cénomanien du “Bassin Occidental Portugais” au sud de l'Accident de Nazaré (Province d'Estremadure, Portugal) , Paris : P. & M. Curie Univ. D. Sci. Diss .
- Heer , O. 1881 . Contributions à la flore fossile du Portugal , Lisboa : Comiss. Trab. Geol. Portugal .
- Heimhofer , U. , Hochuli , P. A. , Burla , S. and Weissert , H. 2007 . New records of Early Creteaceous angiosperm pollen from Portuguese coastal deposits: Implications for the timing of the early angiosperm radiation . Rev. Palaeobot. Palynol. , 144 : 39 – 76 .
- Heimhofer , U. , Hochuli , P. A. , Burla , S. , Dinis , J. M. L. and Weissert , H. 2005 . Timing of Early Cretaceous angiosperm diversification and possible links to major palaeoenvironmental change . Geology , 33 : 141 – 144 .
- Hughes , N. F. 1961 . Further interpretation of Eucommiidites Erdtman 1948 . Palaeontology , 4 : 292 – 299 .
- Hughes , N. F. 1994 . “ The enigma of angiosperms origins ” . Cambridge, , UK : Cambridge Univ. Press .
- Kuyl , O. S. , Muller , J. and Waterbolk , H. T. 1955 . The application of palynology to oil geology with special reference to Western Venezuela . Geol. Mijnbouw , 17 : 49 – 75 .
- Kvaček , J. and Pacltová , B. 2001 . Bayeritheca hughesii gen. et sp. nov., a new Eucommiidites-bearing pollen organ from the Cenomanian of Bohemia . Cret. Res. , 22 : 695 – 704 .
- Leereveld , H. , de Haan , P. J. and Juhász , M. 1989 . Stratigraphic evaluation of spore/pollen assemblages from the Lower Cretaceous of the Alpine-Mediterranean realm , Utrecht : Lab. Palaeobot. Palynol. Utrecht Univ. Sp. Serv. Rep. 89/07 .
- Mendes , M. M. , Friis , E. M. and Pais , J. 2008a . Erdtmanispermum juncalense sp. nov., a new species of the extinct order Erdtmanithecales from the Early Cretaceous (probably Berriasian) of Portugal . Rev. Palaeobot. Palynol. , 149 : 50 – 56 .
- Mendes , M. M. , Pais , J. and Friis , E. M. 2008b . Raunsgaardispermum lusitanicum gen. et sp. nov., a new seed with in situ pollen from the Early Cretaceous (probably Berriasian) of Portugal: Further support for the Bennettitales-Erdtmanithecales-Gnetales link . Grana , 47 : 211 – 219 .
- Pais , J. and Reyre , Y. 1981 . Problèmes posés par la population sporopollinique d'un niveau à plantes de la série de Buarcos (Portugal) . Bol. Soc. Geol. Portugal , 22 : 35 – 40 .
- Pais , J. and Trincão , P . 1983 . Contribuiç[otilde]es para o estudo estratigráfico e paleobotânico dos “grés grosseiros inferiores” (arenitos do Carrascal). I – A jazida de Almas do Juncal . Ciências Terra , 7 : 141 – 160 .
- Pedersen , K. R. , Crane , P. R. and Friis , E. M. 1989 . Pollen organs and seeds with Eucommiidites pollen . Grana , 28 : 279 – 294 .
- Rey , J. , Dinis , J. L. , Callapez , P. and Proença Cunha , P. 2006 . Da rotura continental à margem passiva. Composição e evolução do Cretácico de Portugal , Lisboa : Cadernos de Geologia de Portugal, Ed. INETI .
- Reymanówna , M. 1968 . On seeds containing Eucommiidites troedssonii pollen from the Jurassic of Grojec, Poland . Bot. J. Linn. Soc. , 61 : 147 – 152 .
- Romariz , C. 1946 . Estudo e revisão das formas portuguesas de Frenelopsis . Bol. Mus. Lab. Min. Geol. Fac. Ciên. Univ. Lisboa , : 135 – 149 .
- Rothwell , G. W. , William , L. C. and Stockey , R. A. 2009 . Is the anthophyte hypothesis alive and well? New evidence from the reproductive structures of Bennettitales . Am. J. Bot. , 96 : 296 – 322 .
- Rydin , C. , Pedersen , K. P. and Friis , E. M. 2004 . On the evolutionary history of Ephedra: Cretaceous fossils and extant molecules . Proc. Natl. Acad. Sci. USA , 101 : 16571 – 16576 .
- Rydin , C. , Pedersen , K. R. , Crane , P. R. and Friis , E. M. 2006 . Former diversity of Ephedra (Gnetales): Evidence from Early Cretaceous seeds from Portugal and North America . Ann. Bot. , 98 : 123 – 140 .
- Saporta , G. 1894 . Flore fossile du Portugal. Nouvelles contributions à la flore Mésozoïque, accompagnées d'une notice stratigraphique , Lisbon : Mem. Dir. Trav. Geol. Portugal .
- Scheuring , B. W. 1970 . Palynologische und palynostratigraphische Untersuchungen des Keuper im Bölchen-Tunnel (Solothurner Jura) , Vol. 88 , Basel : Komm. f. Schweiz. Paläontol. Abhandl. Abh .
- Scheuring , B. W. 1978 . Mikrofloren aus den Meridekalken des Mt. San Giorgio (Kanton Tessin) , Vol. 100 , Basel : Komm. f. Schweiz. Paläontol. Abhandl. Abh .
- Teixeira , C. 1948 . Flora Mesozóica Portuguesa, Parte I , Lisbon : Mem. Serv. Geol. Portugal .
- Teixeira , C. 1950 . Flora Mesozóica Portuguesa, Parte II , Lisbon : Mem. Serv. Geol. Portugal .
- Teixeira , C. 1952 . Notes sur quelques gisements des végétaux fossils du Crétacé des environs de Leiria . Rev. Fac. Ciên. Lisboa , 2 : 133 – 154 .
- Tekleva , M. V. , Krassilov , V. A. , Kvaček , J. and Van Konijnenburg-van Cittert , J. H. A. 2006 . Pollen genus Eucommiidites: Ultrastructure and affinities . Acta Palaeobot. , 46 : 137 – 155 .
- Trincão , P. 1990 . Esporos e pólenes do Cretácico Inferior (Berriasiano-Aptiano) de Portugal: Paleontologia e Biostratigrafia , Lisbon : Univ. Nova de Lisboa. Ph. D. Diss .
- Van Konijnenburg-van Cittert , J. H. A . 1971 . In situ gymnosperm pollen from the Middle Jurassic of Yorkshire . Acta Bot. Neerl. , 20 : 1 – 96 .
- Van Konijnenburg-van Cittert , J. H. A. 1972 . Some additional notes on male gymnosperm fructifications from the Jurassic flora of Yorkshire . Acta Bot. Neerl. , 21 : 95 – 98 .
- Zbyszewski , G. , França , J. C. and Ferreira , O. 1961 . Carta geológica de Portugal na escala 1:50 000. Notícia explicativa da folha 26-B, Alcobaça , Lisbon : Serv. Geol. Portugal .