Abstract
Pollen deposition during 1997–2004 was monitored for seven sites in northern Sweden, ranging from the boreal forest zone to above the birch forest-line. Fifty-three taxa were recorded in the traps. Of these, 22 are observed in the vegetation surrounding the trap sites. The remaining 31 taxa are most probably long distance transported. Taxa that strongly correlate with the site above the birch forest-line are Trientalis europaea, Asteraceae, Ericales and Linnaea borealis, while pollen of Pinus, Picea, Betula, Alnus and Juniperus correlates with sites within the boreal forest zone. A positive correlation is noted between pollen accumulation rates (PARs) of Pinus and Picea and mean July temperatures of the year prior to pollen release. A comparison between pollen counts retrieved from pollen traps, lake surface sediments and moss polster samples at one site shows similar pollen composition for the pollen trap and lake surface sediments, while the moss polster samples have higher percentage of Pinus pollen, lower percentage of Betula pollen and generally a lower diversity of pollen and spores.
Pollen accumulation rates (PARs) as well as pollen percentages are used to interpret fossil pollen data and to reconstruct past vegetation. PARs show the total pollen deposit per taxon, surface unit and year (grains/cm2/year) and may, therefore, more reliably reflect changes in pollen production and vegetation composition than the relative composition shown by pollen percentages. For northern Fennoscandia, PAR threshold values have been used in estimating absence, presence and dominance of the most common tree species, i.e. Pinus, Picea and Betula, in vegetation around any given site (Hicks, Citation2001; Seppä & Hicks, Citation2006). Threshold values have been provided by analysis of core-top samples from lake sediments (Hyvärinen, Citation1975, Citation1976; Seppä, Citation1996; Seppä et al., Citation2002) and from pollen trap data (Seppä & Hicks, Citation2006) and they seem to match especially well for Pinus (Seppä & Hicks, Citation2006). Several investigations have also been published for past vegetation in northern Scandinavia using PAR values in vegetational reconstructions (e.g. Barnekow, Citation1999a , Citation1999b ; Barnekow & Sandgren, Citation2001; Bjune et al., Citation2004; Giesecke, Citation2004, Citation2005; Bergman, Citation2005; Bjune, Citation2005; Karlsson et al., Citation2007; Karlsson, Citation2008). For interpretation of pollen records from forest-line ecotones and for pollen assemblages for which there is no modern analogue vegetation, the use of PARs is particularly valuable (Birks & Birks, Citation1980; Birks, Citation1984; Hicks & Hyvärinen, Citation1999; Birks et al., Citation2000; Rull et al., Citation2005; Hicks, Citation2006; Seppä & Hicks, Citation2006).
One problem in using PARs in reconstruction based on the fossil pollen record is, however, that detailed and reliable data on sedimentation rates are required (e.g., Jensen et al., Citation2002; Telford et al., Citation2004; Seppä & Hicks, Citation2006). When using PAR values from pollen traps instead of PARs from lake surface sediments, problems of soil erosion, catchment size and inflowing streams (e.g. Bonny, Citation1976, Citation1978; Bergman, Citation2005) are avoided and also allow more precise temporal and spatial resolution (Seppä & Hicks, Citation2006).
Understanding the connection between pollen production and deposition is important in all interpretations of pollen records. PAR data obtained by pollen traps are, therefore, particularly valuable since they show actual pollen deposition at a specific site with a yearly resolution. Comparison of PAR values from Finland, Sweden, Norway, Iceland and Svalbard (Hättestrand et al., Citation2008) showed that PARs can vary considerably in different regions within the same vegetation. The study also stressed the importance of modern analogue PAR data sets for local areas for interpretation of fossil PARs.
The aim of the present study is to provide a regional pollen monitoring dataset, including PARs and percentages, for northern Sweden that can be used as a modern analogue for studies of fossil pollen stratigraphy and vegetation reconstruction within the boreal–alpine transition. Furthermore, the study aims to increase our understanding of the relationship between pollen production and pollen deposition for different taxa under different environmental conditions.
The study is based on pollen monitoring data from 14 traps at seven sites in northern Sweden. Trap sampling was initiated in 1997 from sites within the boreal forest zone, where Weichselian interstadial sediments had been collected earlier. In 1998 and 1999, the collecting was extended to sites just below and above the forest-line of birch, and the monitoring continued at all sites until 2004. PAR values for individual pollen traps and site mean and median PARs are discussed. The correlation between PARs of Pinus, Picea and Betula and mean July temperatures are investigated through linear regression, and the general trends of the PAR data are explored through a principle component analysis (PCA)-ordination. The pollen trap data were also used to calculate percentages and results from one of the sites were compared with lake surface sediment samples and moss polster samples.
The pollen monitoring data presented here has been used as reference when interpreting interstadial pollen assemblages from the monitoring sites of Rissejauratj and Riipiharju (Hättestrand, Citation2007; Hättestrand & Robertsson, Citation2010). Part of the dataset (1997–2002) was also included in a study, where the relationship between pollen production and deposition at the north-western fringe of the European boreal forest was examined (Hättestrand et al., Citation2008).
Material and methods
Sites and fieldwork
Pollen monitoring was performed at seven sites in northern Sweden, from the boreal forest to above the present forest-line (, , ). Two modified Tauber traps (Tauber, Citation1967, Citation1974; Hicks & Hyvärinen, Citation1986; Hicks et al., Citation1996) were placed at each site. Sampling and laboratory procedures follow the standardised methods of the Pollen Monitoring Programme (Hicks et al., Citation1996, Citation1999). An exception is that the trap opening was larger than recommended (32 cm2 instead of 19.6 cm2) due to the use of trap lids with a pre-made circular knob with a diameter of 6.4 cm that could easily be sawed off when constructing the traps. The pollen traps passively collect airborne material at ground level. They were emptied in September–October after the pollen deposition season and the pollen year is the year of collection. The traps are given codes (e.g., 971A; 97 represents the year of collection, 1997, 1 the monitored site number and A the individual trap). The sampling of individual traps occasionally failed for various reasons, and hence, there are gaps in the time series for some sites, but most recorded successfully (). For traps, where the lid of the container was broken (through cracking), the content of the traps was used to calculate pollen percentages but not PAR values (within brackets in ). For site 5, the trap 5B was moved c. 3 m in 2003 to avoid destruction by terrain vehicles; the new location was maintained also for 2004.
Figure 1. Map of the sampling localities. Vegetation zonation is from Treter (Citation1984). Meteorological stations: Vm = Vajmat, Sv = Svartbyn, Ph = Paharova, St = Saittarova, Pk = Parkalompolo, NS = Nedre Soppero, Nm = Naimakka.
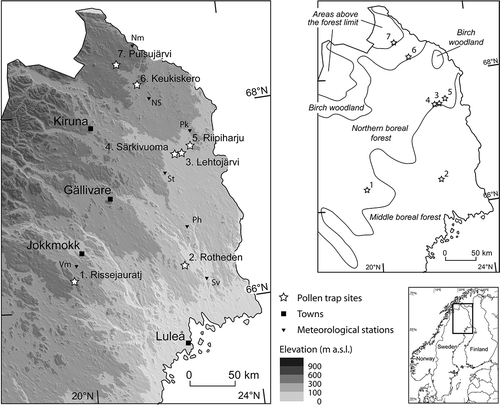
Figure 2. Aerial photographs of the pollen trap sites. All the photographs represent 3 × 3 km and north is to the top. The pollen trap sites are indicated by an inversed triangle and the circles around the sites mark an area with a radius of 500 m from each site. The main tree species referred to in the figure are Pinus sylvestris, Picea abies, Betula pubescens ssp. tortuosa, Betula pendula, Alnus incana and Populus tremula.

Table I. Location of the pollen traps
Table II. Recorded trap years
Detailed vegetation mapping was carried out in a 10 m radius circle around each trap. The vegetation was further surveyed in a 10 to 100 m radius circle, while the vegetation in a broader area (approximately 2 km) was studied through interpretation of aerial photographs and field observations. The aerial photographs used were infrared colour photographs at the scale 1:60 000.
At Rissjauratj-1 (site 1), two samples of moss polsters and two samples of lake surface sediments were collected in 2003 within the same mire surface that was monitored by the pollen traps. The sampled lakes (c. 3 and 20 m from the traps) are small and shallow (c. 4 × 10 m and 40 × 40 m in diameter, and 1–3 m deep), and were sampled from the margin of the water body using a long fishing rod with a cup at the end. Only the uppermost part (c. 0–5 cm) of the sediments was sampled. For the moss polster samples, the uppermost 1 cm of a 10 × 10 cm surface of Sphagnum moss was cut and put into a plastic bag and amalgamated into a single sample by mixing. One moss polster sample was sampled c. 0.2 m from the B-trap and the other was sampled c. 15 m away.
Temperature data
Meteorological data from seven stations in northern Sweden () was obtained from the Swedish Meteorological and Hydrological Institute (SMHI), Örebro. Site specific July temperatures were calculated from every day 24-hour mean values obtained at the meteorological stations closest to the pollen monitoring sites (). When the trap sites were situated between two meteorological stations, data from both stations was used. To compensate for decrease in temperature with increasing elevation, a correction of 0.65 °C per 100 m height was used when transposing temperatures from the meteorological stations to the trap sites.
Laboratory methods
The pollen trap content was sieved through an ordinary kitchen sieve and then through a 250 μm mesh to remove macroscopic objects such as leaves, insects and other small animals (rodents, frogs and in one case, a bird). The macro objects were washed with distilled water, so that pollen from their surface would return into the pollen trap before the objects were removed and annotated. A known amount of Lycopodium clavatum L. spores (3–6 tablets depending on site) were added to the pollen traps (Stockmarr, Citation1971, Citation1973) for calculation of pollen concentrations. Excess water was removed from the trap content by filtering using a suction funnel and filter paper that dissolve in acetolysis mixture. Thereafter, the filter papers were treated by acetolysis (Fægri & Iversen, Citation1989; Hicks et al., Citation1996, 1999) and samples with mineral material were further treated with hydrofluoridic acid (HF).
The lake surface sediment samples were treated by standard methods (Fægri & Iversen, Citation1989), while the amalgamated moss polsters were boiled in sodium hydroxide (NaOH) and washed through a 250 μm mesh before standard preparation (Fægri & Iversen, Citation1989).
The concentrated pollen samples were mounted on slides and analysed under light microscope at 400–1000× magnification. At least 500 grains were counted for each sample, except for some samples with low pollen content and/or high amount of minerogenic particles. Samples with very low pollen counts (< 100 grains) were excluded from the study. However, three monitored traps with fairly high pollen counts were included even though the number of grains was < 500 (006A/449 grains; 002A/473 grains; 027A/197grains). The keys of Fægri and Iversen (Citation1989) and Moore et al. (Citation1991) were used for identification of pollen grains together with the modern reference collection at Stockholm University. Most taxa were identified at species or genus level, but some taxa were only identified at the family or higher taxonomic level.
Numerical analysis of pollen monitoring data
PAR is defined as the number of pollen grains deposited on a 1 cm2 surface during one year. In pollen traps that are emptied every year, the PAR values are calculated by taking the total amount of pollen of each taxon in the trap divided by the size of the trap opening. The number of grains in the whole trap can be calculated through the relation of counted pollen to counted spores of added Lycopodium clavatum in the analysed pollen slides, and the relation of counted spores in the slides to the total amount of added Lycopodium clavatum spores to the trap. Mean and median PAR values for each taxon were calculated for all sites and all recorded traps (A + B). Since pollen deposition can vary very much between years, the occurrence of extreme years in the dataset will have a large effect on the mean values. Also problems in the sampling, like insects in the traps, can affect the mean values. Median values show the ‘middle’ year and therefore, they better represent the ‘normal’ amount of pollen deposited at a site each year. In addition, a comparison was made between plants growing at the sites of pollen monitoring and recorded pollen taxa in the traps.
Linear regression was performed to compare the relation between PAR values of Pinus, Picea and Betula and mean July temperatures of the year prior to pollen emission (the year of pollen formation). Two types of regression analysis were made, one based on PARs versus temperatures combining data from all trap sites and another one based on data from each site separately.
PCA was used to identify major trends within the PAR dataset. In selection of taxa for the ordination, all taxa that in at least one pollen trap sample exceeded 50 grains/cm2/year were used. PCA, a linear ordination type, was selected after running a detrended correspondence analysis (DCA) showing that the gradient length was too short [< 2 standard deviation (SD)] to represent unimodal response (Ter Braak & Prentice, Citation1988). In the PCA ordination, the scaling was focused on inter-sample distances, the species scores were divided by standard deviation, the species data were log-transformed and the species data table was centred by species. The ordinations were made with the software package Canoco v4.5 (Ter Braak & Smilauer, Citation2002). The statistical significance of the first and second axis in the PCA-ordination were tested in Psimpoll v4.25 (Bennett, Citation2005), by a method based on exceeding eigenvalues generated by a broken-stick model of the distribution of variance amongst the various components (Legendre & Legendre, Citation1983; Jackson, Citation1993).
Mean pollen percentages of all monitored years were also calculated for each trap. Raw pollen counts were first used to calculate percentages for each single trap (e.g. for 971A). The percentages from single traps were thereafter used to calculate mean percentages for all recorded years (e.g. mean for 97-041A). The basic sum (ΣP) for the percentage calculations includes pollen of trees, shrubs, dwarf shrubs and herbs. The percentages of aquatic plants, spore plants and varia are based on (ΣP) + (sum of aquatic plants alt. spores alt. varia). For the lake surface sediment and moss polster samples from Rissejauratj, pollen percentages were also calculated, using the same basic sums as for the pollen trap samples.
Results
Meteorological parameters
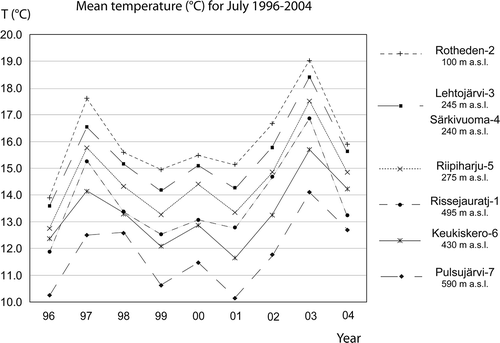
The transferred July mean temperatures for 1996 to 2004 at the pollen monitoring sites show that the temperatures at the sites generally follow each other and that elevation and latitude are important parameters for the temperatures (; Table I). For all years, the July temperatures at Lehtojärvi, Särkivuoma and Riipiharju are higher than at Rissejauratj even though the sites are situated c. 150 km north of Rissejauratj. This is explained by Rissejauratj being located at a higher elevation [495 m compared to 240, 245 and 275 m above sea level (a.s.l.), respectively]. However, the July temperatures at Keukiskero, c. 250 km north of Rissejauratj, are for most of the years lower than at Rissejauratj even though Keukiskero is situated 65 m lower in elevation. In the presentation of recorded PARs and percentages, the pollen monitoring sites are sorted according to their mean July temperatures, from the coldest to the warmest site (Pulsujärvi-7, Keukiskero-6, Rissejauratj-1, Riipiharju-5, Särkivuouma-4, Lehtojärvi-3 and Rotheden-2).
Figure 3. Mean July temperatures 1996–2004 for the pollen trap sites. The July temperature for Pulsujärvi in 1998 is deviating from the trend for the other sites, since some of the colder days of July in 1998 were not recorded at the Naimakka meteorological station.
PAR values
PAR values were calculated for all taxa present in five or more traps (). Consistently high accumulation rates are recorded for pollen from the dominant trees Pinus (mean PAR value of 405–5110), Picea (17–616) and Betula (977–3086), from shrubs of Salix (8–92) and Juniperus (3–892), dwarf shrubs of Ericales (19–522) and the herbs of Gramineae (16–360) and Cyperaceae (25–242). The PARs of Pinus, Betula and Picea vary substantially from year to year, while the PARs for Ericales and Gramineae are more equal from year to year than values of most other taxa.
Figure 4. Annual PAR alues of trees (A), shrubs and dwarf shrubs (B) and herbs (C), in the monitored traps. All herb taxa that occur in five or more traps are shown. Where the PAR bar is hardly visible, dots are used to illustrate presence of a taxon in a trap. Herb taxa presented in the PCA-ordination, that occur in less than five traps, are listed here with trap name and PAR value: Saxifraga cernua (007A:150); Potentilla type (006A/7: 015A/78: 995A/44: 972B/6); Caryophyllaceae (047B/84: 001A/8: 991A/6: 971B/7); cf. Stachys sylvatica-type (006B/51: 995A/3: 974B/61); Potamogeton-type (986A/11: 021B/14: 011B/43).

At the boreal forest sites, mean PARs for Pinus range between c. 2800 and 5100 and for Picea the values range between c. 140 and 620 (). For sites above the Pinus and Picea forest-lines (Keukiskero-6 and Pulsujärvi-7), PARs are about 400–720 for Pinus and 15–65 for Picea. Betula PARs range between 1100 and 3100 where Betula trees grow (site 1–6), while just above the birch forest-line (at Pulsujärvi-7), PARs are c. 980. Except for Pinus, Picea and Betula, mean PARs higher than 50 are only recorded for taxa growing within 100 m of the trap sites (). For Salix, Juniperus, Ericales, Gramineae and Cyperaceae, mean PARs usually range between 10 and 520. These taxa are all very common within the area of pollen monitoring. High PARs for Ericales are generally recorded for the sites situated above the forest-line of Pinus and Picea (). At Riipiharju, a site with high mean PARs for Juniperus and Salix (c. 892 and 92, respectively), stands of these taxa are growing within few metres of the traps. Pulsujärvi-7 and Riipiharju-5 are sites close to grassland surfaces and in these traps Gramineae PARs are relatively high (mean PAR of 158–360 compared to16–67 for the other sites). For the herbaceous taxa Trientalis, Rubus chamaemorus L., Linnaea borealis L. and Trollius europaeus L., the mean PAR values are related to specific sites, so that some sites have high mean PARs (28–177), while the values for other sites are generally zero. Of these taxa, only Trollius europaeus has not been found growing close to any of the monitoring sites, although plants have been observed some kilometres away from Pulsujärvi-7 and Keukiskero-6. Plants of Rubus chamaemorus commonly have a high percentage of surface coverage at the mire sites (c. 10–25%), even though the mean PARs of the taxa in the traps are 32 or less.
Table III. Observed PAR values for presence and absence of Pinus, Picea and Betula trees at the pollen monitoring sites presented in this article (northern Sweden), compared with threshold values derived from pollen traps and lake sediment core-tops in northern Finland and Norway (Hicks, Citation2001; Seppä & Hicks, Citation2006)
Table IV. Mean and median PARs, based on A-trap and B-trap values combined, for all recorded years at each site
Table IV. (Continued)
Pollen of Urtica, Filipendula, Artemisia, Chenopodiaceae, Rumex and Ambrosia-type are deposited at all trap sites in about the same quantities, even though plants of these taxa have not been observed close to any of the trap sites (). Asteraceae pollen is also deposited at all trap sites, although PARs of the taxa vary largely between sites. This variation can be explained by plants of the taxon Solidago virgaurea L. being present at some sites ().
Linear regression
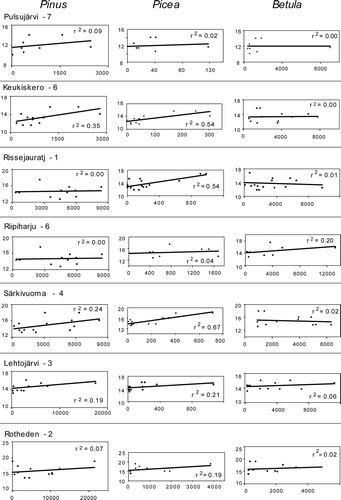
Linear regression analyses based on total counts for all monitored traps shows a correlation between Pinus and Picea PARs, and mean July temperatures, respectively (). A similar correlation was not found for Betula. These analyses include both the variations of mean July temperatures at each site and the vegetation and temperature gradient for the total dataset (Pulsujärvi-7 to Rotheden-2). In the regression analysis of individual sites (), a correlation between Picea PARs and mean July temperatures is clear, while for Pinus this correlation is weakly positive or zero at the site level, and for Betula weakly positive or sometimes negative ().
Figure 5. Linear regression of PAR values versus mean July temperatures for all recorded traps.
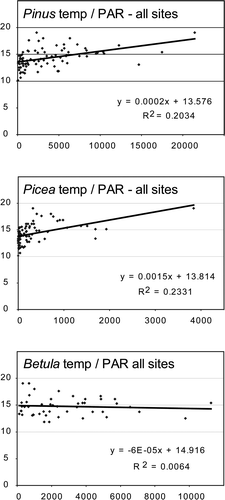
Figure 6. Linear regression of PAR values versus mean July temperatures, for all sites separately.
Ordination analysis
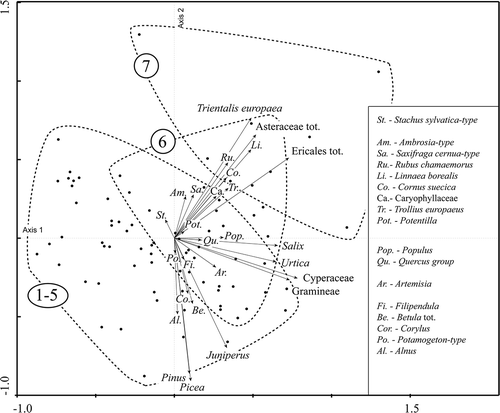
The PCA-ordination outcome for the traps situated above the forest-line at Pulsujärvi-7 does not overlap with the outcome of the traps within the boreal forest zone (traps 1–5; ). However, monitoring data for Keukiskero-6, situated within the birch forest zone but above the forest-line of pine and spruce, does overlap with both the data from Pulsujärvi-7 and the boreal sites (traps 1–5). Ordination axis 1 shows the gradient between pollen trap samples with low PARs and samples with high PARs (). This gradient is largely dependent on PAR variation in herbaceous taxa such as Cyperaceae, Gramineae and Urtica. Axis 2 is primarily displaying the differences between sites with high tree pollen PARs (Picea, Pinus, Alnus, Betula, Corylus) and sites with low tree pollen PARs but high values for some herb taxa (Trientalis europaea, Asteraceae, Linnaea borealis, Rubus chamaemorus, Cornus suecica L., etc.). Ordination axis 1 and 2 explain 22% and 15% of the total variance of the dataset, respectively.
Figure 7. PCA-ordination showing all monitored traps. Lines are drawn to visualise the ordination outcome for the traps of each site. Trap site 7 (Pulsujärvi) is located above the birch forest limit, site 6 (Keukiskero) is placed within the birch forest zone, the trap sites 1–5 are situated within the boreal forest zone (1 Rissejauratj, 2 Rotheden, 3 Lehtojärvi, 4 Särkivuoma, 5 Riipiharju).
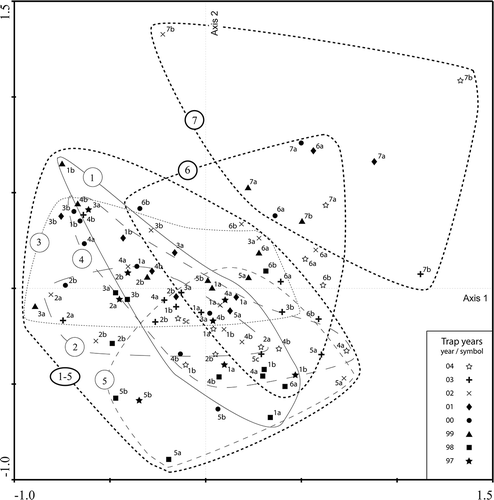
Figure 8. PCA-ordination showing the relation of recorded pollen taxa to the individual samples of the traps. All taxa with PARs > 50 in at least one trap are used in the ordination. Also the ‘Quercus group’ is included, with pollen from trees growing in southern Sweden (Quercus, Tilia, Acer, Larix, Carpinus and Ulmus), even though the PARs of the group are < 50 in all traps.
Percentage data
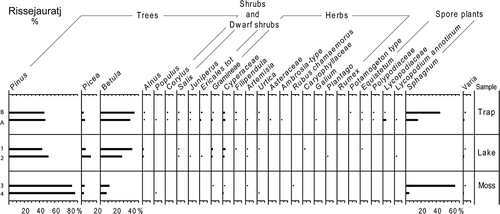
The highest Pinus percentages, about 65%, are recorded for Rotheden-2 within the boreal forest (, ). For the other sites within the boreal forest zone, the Pinus percentages range between 35% and 53%, while traps above the forest-line have Pinus percentages between 12% and 23%. The percentage values for Picea range between 0.5% and 9% for all sites. Betula percentage values range from 15% to 50% at the boreal sites, from 45% to 70% at Keukiskero-6, and from 30 to 40% at Pulsujärvi-7. The percentage values of other recorded tree pollen (except Pinus, Picea and Betula) are mostly below 1%. The Ericales percentage values are highest at Keukiskero-6 and Pulsujärvi-7 (c. 5–30%). Herbaceous taxa with percentages above 1% at any of the trap sites are Gramineae, Cyperaceae, Urtica, Trientalis europaea, Rubus chamaemorus, Linnaea borealis, Asteraceae and Filipendula (, ). High Sphagnum percentages are recorded at the mire sites Rissejauratj-1 and Särkivuoma-4.
Figure 9. Pollen monitoring data calculated into percentages. The calculations are performed on raw pollen counts for the A and B traps separately. Percentages between 0.1 and 0.9% are illustrated as dots, while taxa with lower percentages are not presented in the figure. High Ericales percentages for trap 4A are probably caused by insects.

Table V. Pollen monitoring data calculated into percentages
Percentages from pollen traps, lake surface sediments and moss polsters
Percentage values for Pinus and Betula are similar for pollen trap and lake surface sediment samples (c. 45–50% for Pinus and c. 25–45% for Betula; ). In contrast, the moss polster samples from the mire surface have much higher percentages of Pinus (c. 85%) and markedly lower percentages for Betula (c. 10%). Percentage values of Picea are low, about 3–12%, in all samples. Grains from shrubs, dwarf shrubs, herbs, water plants and spore plants are recorded most commonly in the pollen trap samples, and more commonly in the lake surface sediment samples than in the mire surface samples. However, Sphagnum spores reach high percentage values in the pollen trap samples and in the mire surface samples, but they do not occur at all in the lake surface samples.
Figure 10. Pollen percentages from the Rissejauratj pollen traps (mean for 1997 to 2004 for the A-trap and mean for 1997 to 1998 and 2000 to 2004 for the B-trap) compared to samples of lake surface sediments from two small lakes and moss polsters retrieved from the same site in 2003. Lake surface sediment sample 1 is retrieved from a small water body (4 × 10 m) c. 3 m from the pollen traps, while lake surface sediment sample 2 is collected c. 20 m away in a larger water body (c. 40 × 40 m). Moss polster sample 3 is taken from a Sphagnum tussock c. 0.2 m from the B-trap and moss polster sample 4 is retrieved c. 15 m from the traps, on a lower and wetter Sphagnum surface.
Discussion
PARs and vegetation
The pollen deposition at a site reflects the composition of the surrounding vegetation as well as the background pollen component, i.e. pollen transported over longer distances from beyond the surveyed area (Andersen, Citation1970). In the present study, the total pollen monitoring data set includes 22 taxa that are known from the vegetation around all trap sites and 31 pollen types that represent plants not occurring close to the traps. Included in the background pollen component are pollen types representing plants distribution only in southern Scandinavia (as e.g. Quercus and Ambrosia) as well as plants that grow also in northern Scandinavia (e.g. Urtica and Filipendula), but not at the trap sites. The mean PARs for these taxa are generally low (0–10) with the exception of Urtica that has relatively high (6–47) mean PAR values. Pollen of Gramineae and Cyperaceae is the most common type of herbaceous pollen recorded in the traps followed by pollen of Urtica.
Several of the long distance transported herbaceous taxa in the pollen traps (Artemisia, Chenopodiaceae, Rumex and Caryophyllaceae) are abundant in the Weichselian interstadial cold records of northern Sweden (Lagerbäck & Robertsson, Citation1988; Hättestrand & Robertsson, Citation2010). It is possible that the occurrence of these taxa in the interstadial pollen samples indicate extremely low local pollen production rather than high frequency of the taxa in the vegetation surrounding the investigated sites.
Since presence of forest forming trees close to the forest-line is closely related to mean July temperatures, past distribution of forest trees from fossil pollen records can be used as a proxy in reconstructing palaeotemperatures. Particularly useful is the comparison of fossil PARs to modern analogue PAR data from sites within the forest-line ecotone, where the taxa grow at present. Pollen monitoring has been used to delimit PARs for vegetation close to tree- and forest-lines in northern Finland and Norway (Hicks, Citation2001; Jensen et al., Citation2007; Seppä & Hicks, Citation2006).
PARs for absence and presence of Pinus, Picea and Betula in the vegetation surrounding the trap sites were calculated () and compared to threshold values for crossing the tree-lines of the taxa (Hicks, Citation2001; Seppä & Hicks, Citation2006). The definition of ‘absence’ in the present study is that the traps are situated above the forest-line of the taxa and that no trees (plants taller than 2 m) of the taxa occur within a 300 m radius (or more) of the traps. The definition of tree-line [as used in Hicks (Citation2001) and Seppä and Hicks (Citation2006)] is the ‘limit for the extreme tree-like occurrences of a tree species’, while the definition of forest-line is the ‘limit of continuous forest’ (Tuhkanen, Citation1999). Reasons that explain why the PARs of the northern Swedish traps are not related to tree-line positions [as in Hicks (Citation2001) and Seppä and Hicks (Citation2006)] are that the tree-line is difficult to define in the pollen monitoring area since the boreal/alpine transition zone here is very wide including single scattered trees of especially Pinus and Picea, and that the position of the tree-line is less important for the deposition of tree pollen at the sites than distance to the nearest trees and distance to the forest-line. However, these differences in definition between datasets are probably of minor practical importance for the present comparison.
PARs for sites with absence of Pinus, Picea and Betula trees are in the present study higher (for the sites closest to the forest-lines of the taxa) than the PAR threshold values for tree-lines of the same taxa obtained by Hicks (Citation2001) and Seppä and Hicks (Citation2006) (). Differences between the data sets are found in sampling period, definition (absence of taxa versus relation to tree-line position) and geographical location. The sampling period for the north Swedish sites (6–8 years) is shorter than for the pollen trap samples presented in Hicks (Citation2001; 13–18 years). However, when comparing PARs for northern Sweden over the period 1997–2004 with those from 1982–1999 in Hicks (Citation2001), it can be concluded that the north Swedish record probably does not have extremely high mean PARs for Pinus, Picea and Betula. Different background pollen components due to a more southern location for the Swedish traps can be part of the explanation to higher PARs for sites without trees in the north Swedish record. The sub-alpine birch forest inland in northern Norway, sampled by pollen traps 1996–2004, is characterised by tree-Betula PARs of c. 2000 (Jensen et al., Citation2007). At Keukiskero-6, sampled within the birch forest zone of northern Sweden, the Betula PARs (including both tree and dwarf birch) are c. 2400. For open birch woodland at the coast of northern Norway, the tree-Betula PARs are only about 80–350 (Jensen et al., Citation2007). As expected, the mean PARs for Pinus (2900–5100), Picea (140–620) and Betula (1100–3100) at the northern Swedish trap sites within the boreal zone are higher than the threshold values delimiting forest vegetation described by Hicks (Citation2001) and Seppä and Hicks (Citation2006) (). It is worth noting that the site Rotheden-2, which is situated within a mixed forest, records higher PARs for Pinus and Picea, but lower values for Betula than the open and semi-open sites. In northern Norway, mean PARs of Pinus are as low as 200–400 for sites in the open coastal lowland, while at inland sites with mixed birch-pine forest, mean PARs of Pinus are 200–400 and for dense pine-dominated forest inland PARs are ≥ 2000 (Jensen et al., Citation2007).
PARs and mean July temperatures
July mean temperature is commonly used when reconstructing palaeotemperatures (e.g. Rosén et al., Citation2001; Seppä & Birks, Citation2001; Andreev et al., Citation2003). Comparison between climate proxy records are facilitated if palaeoecological records can be related to July mean temperatures or to other commonly reconstructed parameters such as summer mean temperature or temperature of the warmest month. Autio and Hicks (Citation2004) showed that PARs for Pinus and Picea relate to July temperature data for the year previous to pollen release. In Hättestrand et al. (Citation2008), a relation between absolute temperature of the warmest month and PAR values for the most common pollen taxa was observed for sites within the north-western fringe of the European boreal forest. In the present dataset, July was the warmest summer month for all years, except for 1996 and 2002.
In the present paper, a correlation between mean July temperatures of the year previous to pollen release and PAR data for Pinus and Picea is shown (, ). For Pinus, the correlation is best when relating PARs from all pollen trap sites to the temperature data, while correlation for single sites are weak or lacking. This shows a true relation between the vegetation gradient of the dataset and temperature: There are more Pinus trees and higher PARs at sites with higher July temperatures and less Pinus trees and lower PARs at sites with lower temperatures. For Keukiskero-6 and Pulsujärvi-7, the summer temperatures are too low for pine forests to grow. However, the regression of all trap sites is also based on Rotheden-2, which is the site with the highest temperatures monitored within a closed forest. The fact that Rotheden-2 is sampled within a forest results in higher PARs for some forest trees, than would have been the case if sampling would have been performed at an open or semi-open site nearby. In the regression, where all trap sites are used, this exaggerates the increase of PARs with temperature for these taxa, since Rotheden-2 also is the site with the highest summer temperatures. Strongest correlation between mean July temperatures and PARs is found for Picea at single sites (Keukiskero-6, Rissjauratj-1 and Särkivuoma-4: r 2 of 0.54–0.67), i.e. correlation can also be found at sites just above the Picea forest-line. For Betula, no correlation to mean July temperature is found, which is in accordance with the observations by Autio and Hicks (Citation2004). The Picea regression () is strongly affected by one data with extremely high PAR value. For Picea, the r 2 values are low for sites with trees very close to the traps or sites within the forest vegetation (i.e. Riipiharju and Rotheden; ). High Picea values at these sites could perhaps be due to other factors than high pollen production, e.g. pollen from nearby trees could be transported to the traps by local wind and rain. The R values are probably more reliable for sites with a larger open area around the traps.
The correlation between meteorological parameters and annual pollen deposition was investigated at four sites on a transect across an altitudinal timberline at Aakenustunturi in northern Finland (Autio & Hicks, Citation2004). The correlation of PARs and meteorological parameters was strongest for Pinus sylvestris L. for which there was a correlation of the July mean temperature, July effective temperature sum and total effective temperature sum, for the year previous to spore/pollen release. Pollen deposition of Betula pubescens Ehrh. and Picea abies (L.) H.Karst. were related to thermal factors for early June and early July, respectively (also for the year previous to pollen emission). However, the results of Picea might have been affected by low pollen deposition of the taxon and infection of spruces in the studied region by the rust fungus Chrysomyxa ledi (Alb. et Schwein.) de Bary in two years (Autio & Hicks, Citation2004).
General trends in the PAR data
In the ordinations of the PAR data, Trientalis, Asteraceae, Linnaea borealis and Ericales are dominant for the alpine site of Pulsujärvi-7 (). These taxa have markedly higher values for this site than for the other trap sites (). Keukiskero-6 traps with high Betula PARs group together with traps from the boreal forest, while Keukiskero-6 traps with low Betula PARs group with traps from above the birch forest-line at Pulsujärvi-7 (, ). At Riipiharju, Juniperus and Salix stands grow close to the pollen traps, which can explain the very high Juniperus PARs (> 1000) for some samples (traps 975B, 985A, 985B, 005B) and also high PARs for Salix (, ). Traps with high Juniperus PARs at Riipiharju-5 also have intermediate to high PARs of Pinus, Betula and Picea. The traps placed within the boreal forest (Rotheden-2) are generally grouped in the left part of the ordination () indicating lower correlation with Salix, Urtica, Cyperaceae and Gramineae for traps at this site than for traps of other sites (all other, except for Pulsujärvi-7, are placed at or close to mires). One reason for the relatively low explanatory power of the performed PCA is probably the high variation in the biological data. In the PCAs, all single years are plotted for all sites. Since the PARs vary highly from year to year and different taxa vary in different ways, it is expected that the data should be scattered. However, the PCA-plots show that there is still some clear distinction of data from different sites.
Percentage data
The percentage data based on pollen counts for all monitored years () show that, although pollen deposition differs much from year to year in the traps, mean values based on several years are showing a more consistent picture of pollen deposition at specific sites. At some sites, a small difference in the actual position of the A and B traps have influenced the deposition in the traps. For example, for 6A, there is generally more Ericales pollen deposited than in 6B, which is affecting the percentage data (, ). The traps are placed only c. 2 m apart, but 6A is positioned on a tussock, while 6B is placed on a lower mire surface, and 6B was commonly water filled when emptied.
Comparison between pollen trap percentages at Rissejauratj and percentages from lake surface sediment samples and mire surface samples show that moss samples from the mire surface have markedly higher Pinus percentages and lower Betula percentages than samples from the traps and lakes. Also the number of recorded pollen taxa is highest in the pollen traps and lowest in the mire surface samples. Other studies have shown that Pinus pollen is more abundant in moss samples than in pollen trap samples (e.g. Spieksma et al., Citation1994; Vermoere et al., Citation2000; Räsänen et al., Citation2004; Pardoe et al., Citation2010). It has been proposed that mosses preserve large robust pollen grains better than small grains sometimes with thin exine (e.g. Juniperus) and that small grains may be easily washed out or degraded from mosses (Spieksma et al., Citation1994; Vermoere et al., Citation2000).
Measurements of Sphagnum fuscum (Schimp.) H.Klinggr. growth on two mires in subarctic environments at Abisko and Stordalen in northern Sweden give a length increment of 0.45 cm per season and show that favourable conditions for growth are enhanced by precipitation and increased temperature (Sonesson et al., Citation2002). At Rissejauratj, the climate is warmer than at the studied mires at Abisko and Stordalen, and it is likely that the growth of Sphagnum at Rissejauratj is about 0.45 cm or more per season. Because only the uppermost centimetre was cut off from the moss, probably 1–3 years of pollen deposition was collected in the mire surface samples. Since the sampling was done when the traps were collected at the end of the season of 2003, the pollen monitoring year 2003 is included in the moss samples, but also probably the monitoring year of 2002 and maybe also part of 2001. However, the years of 2003 and 2002 are not characterised by extremely high Pinus and low Betula PARs in the pollen traps (). It is likely that surface water can wash away small grains from the mire surface, while they are retained in the water body of the small lakes as well as in the pollen traps. However, it seems less likely that large amount of Betula grains could be totally destroyed in only 1–3 years. Possibly, pollen grains washed off from the mire surface could be accumulated in the pools/lakes of the mire. However, since the lake samples are close to the samples of the pollen traps, both in percentages of Betula and Pinus and in the presence of other pollen taxa, this assumption is not confirmed in the present study. The fact that the total amount of recorded taxa is lower in the moss polster samples than in the samples of the traps and lakes may reflect the short period of pollen deposition in the moss polster samples (1 cm, c. 1–3 years). If a longer time period had been represented (through sampling to a larger depth), it is likely that a higher amount of rare taxa would have been found in the moss polster assemblages. However, the lower amount of recorded taxa in the samples could also indicate that pollen grains have been washed out from the moss or that they have degraded.
Giesecke and Fontana (Citation2008) compared pollen deposition in modified Tauber traps and pollen deposition in lakes and observed that especially Pinus pollen grains were lost on their way through the water column and thus are underrepresented in the lake sediments. This process was not observed in the present study indicating that other factors may also play a role in pollen deposition such as lake type and where in the lake the sediment sample was taken. Pardoe et al. (Citation2010) suggested that modern pollen data sets as reference for interpretation of fossil pollen data is best collected using a single collecting medium only (e.g. either lake sediments or moss polsters) as the different media clearly seem to vary. The results from the present study are in accordance with this conclusion.
Pollen monitoring and the use of PARs
PAR threshold values for presence and absence of Pinus, Picea and Betula in the vegetation are easy to use when comparing to fossil pollen data. However, general PARs of the most common taxa for sites with different vegetation are also valuable since they add information that can be used in the interpretation of the vegetation surrounding a site.
PARs for single years can show great variation in absolute values () and large differences in ordination outcome (). Still, the mean PARs for the pollen trap sites () and the pollen percentage data () are relatively robust and show general patterns reflecting vegetation differences at the sites studied. The monitoring period is fairly short (6–8 years) in the present study but the record still seems to be long enough to include years of high and low PARs, relate PARs to meteorological data and reveal trends in pollen deposition in the area. The use of double traps is valuable in pollen monitoring, since the possibility of recording PAR data for every year at each site increases. Also, when double traps are recorded for the same year, they can be compared to each other and differences and similarities between the traps can be analysed.
In trap 44A, an extreme value of 7040 is recorded for Ericales pollen (). In the parallel trap 44B, the PAR value for Ericales was only 298. Some of the species included in the Ericales pollen taxon are insect pollinated and a bee was found in the 44A trap. It is likely that the extreme Ericales value in the trap is caused by the insect, influencing both the mean PAR for Ericales at Särkivuoma-4 and the percentage value for Ericales in the 4A-trap (, Tables IV, V).
In the present paper, mean PAR values are used, while median PARs were used in Hättestrand et al. (Citation2008). Both studies investigated the use of PAR values as a reference for fossil material. Mean PARs include years of high and low pollen deposition and are in this paper used to produce long-term mean PARs for the sites. This is comparable to PARs of fossil samples since they generally include both years of high and low accumulation rates (if the samples are based on several years). When trying to show the general trend of PAR values for many sites within a large region, median PARs might be better to use since sites with extreme PARs then are totally excluded and the most common PAR values for the region are shown.
In the present study and in the study of Autio and Hicks (Citation2004), a correlation of Pinus and Picea PARs and July mean temperatures is demonstrated. Barnekow et al. (Citation2007) found a strong correlation between PARs of Pinus, Picea and Betula ssp. deposited in peat and July–August mean temperatures. Kuoppamaa et al. (Citation2009) also observed a correlation between PARs in peat and July mean temperatures; however, they found a stronger correlation for Betula than for Pinus. In future studies, it might be of interest to investigate PARs of Pinus, Picea and Betula spp. in annually laminated sediments and relate them to meteorological data.
Conclusions
This study presents eight years of pollen monitoring in northern Sweden. It includes years of high and low pollen deposition, data on PARs related to meteorological parameters and mean PARs sufficiently robust to reveal trends of pollen deposition related to vegetation. Only 22 of the 53 taxa recorded in the pollen traps are observed in the vegetation around any of the trap sites, while 31 taxa most probably are long distance transported. Except for Pinus, Picea and Betula, site mean PARs higher than 50 are only recorded for taxa growing within 100 m of the trap sites. PARs of Pinus, Picea and Betula trees vary markedly from year to year, while PARs of the dwarf shrub and herb taxa Ericales and Gramineae are characterised by relatively low annual variations. A correlation between PARs of Pinus and Picea and mean July temperatures of the year prior to pollen release is present in the data. PAR values for presence and absence of Pinus, Picea and Betula trees in the surrounding vegetation have been recorded. Values of absence are obtained from traps above the forest-line of the studied taxa, at sites with 300 m or more to the closest tree of the taxa. Pinus PARs are 140–620 for absence and 2800–5100 for presence, Picea PARs are 15–65 for absence and 400–720 for presence and Betula PARs are 980 for absence and 1100–3100 for presence. PARs of Trientalis, Rubus chamaemorus, Linnaea borealis and Trollius europaeus are related to specific sites, while PARs of Urtica, Asteraceae, Filipendula, Artemisia, Chenopodiaceae, Rumex, Umbelliferae and Ambrosia-type are recorded in similar quantities at most trap sites.
A comparison between pollen percentages retrieved from pollen traps, lake surface sediments and moss polsters shows accordance between pollen assemblages from pollen traps and lake surface sediments. The moss polster samples deviate from the other two sample types by having higher percentage of Pinus pollen, lower percentage of Betula pollen and a lower amount of totally recorded pollen taxa.
Acknowledgements
Sheila Hicks and Ann-Marie Robertsson are thanked for introducing the author to pollen monitoring. The author is also thankful to Robert Lagerbäck, who joined the author in 1996 on the first tour to place traps at sites where Weichselian interstadial sediments had earlier been retrieved. The following pollen monitoring was performed with special help from Clas Hättestrand who joined the author in the field and also was responsible for trap collection in 1999 and 2000. Other colleagues/friends that have assisted with the trap collection are: Ola Fredin, Kirk Lurvey, Arjen Stroeven, Gunhild Rosqvist, Sheila Hicks, Ann-Marie Robertsson and Hanna Karlsson. Vegetation mapping at the trap sites was performed with assistance of Ann Karlsson. Laboratory work on the traps collected from 1998 to 2000 was performed by Anders Borgmark and Per Westman, and some of the traps collected in 1997 and 1998 were counted by Raija-Liisa Huttunen, Oulu University, Oulu, Finland. Ann-Marie Robertsson, Clas Hättestrand and Stefan Wastegård kindly provided useful comments on the manuscript and Clas Hättestrand made the map in . In a later stage, also Thomas Giesecke, Else Marie Friis and one anonymous reviewer provided helpful comments improving this work and Christian Pott helped with the illustrations. Financial support was given from the Swedish Natural Science Research Council (NFR), Swedish Society for Anthropology and Geography, Carl Mannerfelts fond, De Geers stipendium and Helge Ax:son Johnsons stiftelse.
Related Research Data
References
- Andersen , S. T. 1970 . The relative pollen productivity and representation of north European trees and correction factors for tree pollen spectra . Danmarks Geologiske Undersøgelse II , 96 : 1 – 99 .
- Andreev , A. A. , Tarasov , P. E. , Siegert , C. , Ebel , T. , Klimanov , V. A. , Melles , M. , Bobrov , A. A. , Yu , Dereviagin, A. , Lubinski , D. J. and Hubberten , H.-W. 2003 . Late Pleistocene and Holocene vegetation and climate on the northern Taymyr Peninsula, Arctic Russia . Boreas , 32 : 484 – 505 .
- Autio , J. and Hicks , S. 2004 . Annual variations in pollen deposition and meteorological conditions on the fell Aakenustunturi in northern Finland: Potential for using fossil pollen as a climate proxy . Grana , 43 : 31 – 47 .
- Barnekow , L. 1999a . Holocene vegetation dynamics and climate change in the Torneträsk area, northern Sweden , Lund : University of Lund, PhD Diss .
- Barnekow , L. 1999b . Holocene tree-line dynamics and inferred climatic changes in the Abisko area, northern Sweden, based on macrofossil and pollen records . The Holocene , 9 : 253 – 265 .
- Barnekow , L. , Loader , N. J. , Hicks , S. , Froyd , C. A. and Goslar , T. 2007 . Strong correlation between summer temperature and pollen accumulation rates for Pinus sylvestris, Picea abies and Betula spp. in a high-resolution record from northern Sweden . Journal of Quaternary Science , 22 : 653 – 658 .
- Barnekow , L. and Sandgren , P. 2001 . Palaeoclimate and tree-line changes during the Holocene based on pollen and plant macrofossil records from six lakes at different altitudes in northern Sweden . Review of Palaeobotany and Palynology , 117 : 109 – 118 .
- Bennett , K. D. 2005 . Documentation for psimpoll 4.25 and pscomb 1.03. C programs for plotting diagrams and analysing data , Software manual , Uppsala : Uppsala University .
- Bergman , J. 2005 . Tree-limit ecotonal response to Holocene climate change in the Scandes Mountains of west-central Sweden , Lund : University of Lund, PhD Diss .
- Birks , H. H. 1984 . “ Late-Quaternary pollen and plant macrofossil stratigraphy at Lochnan and Druim, north-west Scotland ” . In Lake sediments and environmental history , Edited by: Haworth , E. Y. and Lund , J. W. G. 377 – 405 . Leicester : Leicester University Press .
- Birks , H. H. , Battarbee , R. W. and Birks , H. J. B. 2000 . The development of the aquatic ecosystem at Kråkenes Lake, western Norway, during the late glacial and early Holocene – a synthesis . Journal of Paleolimnology , 23 : 91 – 144 .
- Birks , H. J. B. and Birks , H. H. 1980 . Quaternary palaeoecology , London : Arnold .
- Bjune , A. E. 2005 . Holocene vegetation history and tree-line changes on a north-south transect crossing major climate gradients in southern Norway – evidence from pollen and plant macrofossils in lake sediments . Review of Palaeobotany and Palynology , 133 : 249 – 275 .
- Bjune , A. E. , Birks , H. J. B. and Seppä , H. 2004 . Holocene vegetation and climate history on a continental – oceanic transect in northern Fennoscandia based on pollen and plant macrofossils . Boreas , 33 : 211 – 223 .
- Bonny , A. P. 1976 . Recruitment of pollen to the seston and sediment of some Lake District lakes . Journal of Ecology , 64 : 859 – 887 .
- Bonny , A. P. 1978 . The effect of pollen recruitment processes on pollen distribution over the sediment surface of a small lake in Cumbria . Journal of Ecology , 66 : 385 – 416 .
- Fægri , K. and Iversen , J. 1989 . Textbook of pollen analysis , Chichester : John Wiley & Sons .
- Giesecke , T. 2004 . The Holocene spread of spruce in Scandinavia , Uppsala : Uppsala University, PhD Diss .
- Giesecke , T. 2005 . Moving front or population expansion: How did Picea abies (L.) Karst. become frequent in central Sweden? . Quaternary Science Reviews , 24 : 2495 – 2509 .
- Giesecke , T. and Fontana , S. 2008 . Revisiting pollen accumulation rates from Swedish lake sediments . The Holocene , 18 : 293 – 305 .
- Hättestrand , M. 2007 . Weichselian interstadial pollen stratigraphy from a Veiki plateau at Rissejauratj in Norrbotten, northern Sweden . GFF , 129 : 287 – 294 .
- Hättestrand , M. , Jensen , C. , Hallsdóttir , M. and Vorren , K.-D. 2008 . Modern pollen accumulation rates at the north-western fringe of the European boreal forest . Review of Palaeobotany and Palynology , 151 : 90 – 109 .
- Hättestrand , M. and Robertsson , A.-M. 2010 . Weichselian interstadials at Riipiharju, northern Sweden – interpretation of vegetation and climate from fossil and modern pollen records . Boreas , 39 : 296 – 311 .
- Hicks , S. 2001 . The use of annual arboreal pollen deposition values for delimiting tree-lines in the landscape and exploring models of pollen dispersal . Review of Palaeobotany and Palynology , 117 : 1 – 29 .
- Hicks , S. 2006 . When no pollen does not mean no trees . Vegetation History and Archaeobotany , 15 : 253 – 261 .
- Hicks , S. , Ammann , B. , Latalowa , M. , Pardoe , H. and Tinsley , H. 1996 . European Pollen Monitoring Programme: Project description and guidelines , Oulu : University of Oulu .
- Hicks , S. and Hyvärinen , H. 1999 . Pollen influx values measured in different sedimentary environments and their palaeoecological implications . Grana , 38 : 1 – 15 .
- Hicks , S. and Hyvärinen , V.-P. 1986 . Sampling modern pollen deposition by means of “Tauber traps”: Some considerations . Pollen et Spores , 28 : 219 – 242 .
- Hicks , S. , Tinsley , H. , Pardoe , H. and Cundill , P. 1999 . European Pollen Monitoring Programme: Supplement to the guidelines , Oulu : University of Oulu .
- Hyvärinen , H. 1975 . Absolute and relative pollen diagrams from northernmost Fennoscandia . Fennia , 142 : 1 – 23 .
- Hyvärinen , H. 1976 . Flandrian pollen deposition rates and tree-line history in northernmost Fennoscandia . Boreas , 5 : 163 – 175 .
- Jackson , C. A. 1993 . Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches . Ecology , 74 : 2204 – 2214 .
- Jensen , C. , Kunzendorf , H. and Vorren , K.-D. 2002 . Pollen deposition rates in peat and lake sediments from the Pinus sylvestris L. forest-line ecotone of northern Norway . Review of Palaeobotany and Palynology , 121 : 113 – 132 .
- Jensen , C. , Vorren , K.-D. and Mørkved , B. 2007 . Annual pollen accumulation rate (PAR) at the boreal and alpine forest-line of north-western Norway, with special emphasis on Pinus sylvestris and Betula pubescens . Review of Palaeobotany and Palynology , 144 : 337 – 361 .
- Karlsson , H. 2008 . Vegetation changes and forest-line positions in the Swedish Scandes during Late Holocene – anthropogenic impact vs. climate , Umeå : Swedish University of Agricultural Sciences, PhD Diss .
- Karlsson , H. , Hörnberg , G. , Hannon , G. and Nordström , E.-M. 2007 . Long-term vegetation changes in the northern Scandinavian forest limit: A human impact-climate synergy? . The Holocene , 17 : 37 – 49 .
- Kuoppamaa , M. , Goslar , T. and Hicks , S. 2009 . Pollen accumulation rates as a tool for detecting land-use changes in a sparsely settled boreal forest . Vegetation History and Archaeobotany , 18 : 205 – 217 .
- Lagerbäck , L. and Robertsson , A.-M. 1988 . Kettle holes – stratigraphic archives for Weichselian geology and palaeoenvironment in northernmost Sweden . Boreas , 17 : 439 – 468 .
- Legendre , L. and Legendre , P. 1983 . Numerical ecology , Amsterdam : Elsevier .
- Moore , P. E. , Webb , J. A. and Collinson , M. E. 1991 . Pollen analysis , Oxford : Blackwell .
- Pardoe , H. S. , Giesecke , T. , Van der Knaap , W. O. , Svitavská-Svobodová , H. , Kvavadze , E. V. , Panajiotidis , S. , Gerasimidis , A. , Pidek , I. A. , Zimny , M. , Swieta-Musznicka , J. , Latalowa , M. , Noryskiewicz, A. M., Bozilova , E. , Tonokov , S. , Filipova-Marinova , M. V. , Van Leeuwen , J. F. N. and Kalnina , L. 2010 . Comparing pollen spectra from modified Tauber traps and moss samples: Examples from a selection of woodlands across Europe . Vegetation History and Archaeobotany , 19 : 271 – 283 .
- Räsänen , S. , Hicks , S. and Odgaard , B. V. 2004 . Pollen deposition in mosses and in a modified “Tauber trap” from Hailuoto, Finland: What exactly do the mosses record? . Review of Palaeobotany and Palynology , 129 : 103 – 116 .
- Rosén , P. , Segerström , U. , Eriksson , L. , Renberg , I. and Birks , H. J. B. 2001 . Holocene climatic change reconstructed from diatoms, chironomids, pollen and near-infrared spectroscopy at an alpine lake (Sjuodjijaure) in northern Sweden . The Holocene , 11 : 551 – 562 .
- Rull , V. , Abbot , M. B. , Polissar , P. J. , Wolfe , A. P. , Bezada , M. and Bradley , R. S. 2005 . 15 000-yr pollen record of vegetation change in the high altitude tropical Andes at Laguna Verde Alta, Venezuela . Quaternary Research , 64 : 308 – 317 .
- Seppä , H. 1996 . Post-glacial dynamics of vegetation and tree-lines in the far north of Fennoscandia . Fennia , 174 : 1 – 96 .
- Seppä , H. and Birks , H. J. B. 2001 . July mean temperature and annual precipitation trends during the Holocene in the Fennoscandian tree-line area: Pollen-based climate reconstructions . The Holocene , 11 : 527 – 539 .
- Seppä , H. and Hicks , S. 2006 . Integration of modern and past pollen accumulation rate (PAR) records across the arctic tree-line: A method for more precise vegetation reconstructions . Quaternary Science Reviews , 25 : 1501 – 1516 .
- Seppä , H. , Nyman , M. , Korhola , A. and Weckström , J. 2002 . Changes of tree-lines and arctic-alpine vegetation in relation to post-glacial climate dynamics in northern Fennoscandia based on pollen and chironomid records . Journal of Quaternary Science , 17 : 287 – 301 .
- Sonesson , M. , Carlsson , B. Å , Callaghan , T. V. , Halling , S. , Björn , L. O. , Bertgren , M. and Johanson , U. 2002 . Growth of two peat-forming mosses in subarctic mires: Species interactions and effects of simulated climate change . Oikos , 99 : 151 – 160 .
- Spieksma , F. T. M. , Nikkels , B. H. and Bottema , S. 1994 . Relationship between recent pollen deposition and airborne pollen concentration . Review of Palaeobotany and Palynology , 82 : 141 – 145 .
- Stockmarr , J. 1971 . Tablets with spores used in absolute pollen analysis . Pollen et Spores , 13 : 615 – 621 .
- Stockmarr , J. 1973 . Determination of spore concentration with an electronic particle counter , Årbog : Danmarks Geologiske Undersøgelse 1972 .
- Tauber , H. 1967 . Investigations of the mode of transfer of pollen in forested areas . Review of Palaeobotany and Palynology , 3 : 277 – 286 .
- Tauber , H. 1974 . A static non-overload pollen collector . New Phytologist , 73 : 359 – 369 .
- Telford , R. J. , Heegaard , E. and Birks , H. J. B. 2004 . All age-depth models are wrong, but how badly? . Quaternary Science Reviews , 23 : 1 – 5 .
- Tuhkanen , S. 10–11 May 1999 . “ The northern tree-line in relation to climate ” . In Sustainable development in northern tree-line Forests. Proceedings of the tree-line workshop Edited by: Kankaanpää , S. , Tasanen , T. and Sutinen , M.-L. Vol. 1998 , 10–11 May , 29 – 61 . Whitehorse , , Canada
- Ter Braak , C. J. F. and Prentice , I. C. 1988 . A theory of gradient analysis . Advances in Ecological Research , 18 : 271 – 313 .
- Ter Braak , C. J. F. and Smilauer , P. 2002 . Canoco reference manual and Canodraw for Windows user's guide. Software for vanonical community ordination (version 4.5) , Ithaca , NY : Microcomputer Power .
- Treter , U. 1984 . Die Baumgrenzen Skandinaviens – ökologische und dendroklimatische Untersuchungen , Wiesbaden : Franz Steiner Verlag .
- Vermoere , M. , Vanhecke , L. , Waelkens , M. and Smets , E. 2000 . A comparison between modern pollen spectra of moss cushions and Cundill pollen traps: Implications for the interpretation of fossil pollen data from southwest Turkey . Grana , 39 : 146 – 158 .