Abstract
The aim of this study was to estimate the polleniferous potential of the most common plant communities and crops across the landscape through the study of pollen loads collected by Apis mellifera. We analysed pollen load samples from hives located in Mallorca (western Mediterranean) for one year and compared the results with a phytosociological study of the foraging area. We also analysed meteorological data to evaluate the influence of some environmental factors on the pollen production phenology. We identified 74 different pollen types (66.2% from native flora, 25.7% from exotic flora, and 8.1% from crops) belonging to 33 families. The highest polleniferous potentials in the landscape were attained by sclerophyllous forests (Quercetea ilicis) in spring, crops in autumn, wild annual rangelands (Stellarietea mediae) in winter and gardens in late summer. Our results suggest that the higher the coverage present in a vegetation class, the higher the quantity and diversity of pollen that may be provided to pollinators; therefore, its polleniferous potential may be higher. Some meteorological conditions (rainfall, temperature and daily hours of sunlight) may affect this potential through their influence on flowering phenology.
Most plants in Mediterranean ecosystems are pollinated by insects, whose main floral reward is pollen (Herrera, Citation1985; Petanidou & Vokou, Citation1990). Most abundant pollinator groups in studies made in the Mediterranean basin are Hymenoptera and Diptera, with the honeybee (Apis mellifera L.) being dominant (Herrera, Citation1988; Petanidou & Vokou, Citation1990; Potts et al., Citation2006). It is estimated that the honeybee is responsible for 80% of insect pollination (Ortega, Citation1987), maintaining his loyalty to collect the pollen of one species (Butler, Citation1945; Ribbands, Citation1949; Singh, Citation1950; Ortiz, Citation1994) until it has accumulated the loads on his third pair of legs. These loads have a morphology, colour and size depending on the pollen and nectar types used during the pollen collecting process (Sá-Otero et al., Citation2002). Botanical origin can be determined through microscopic analysis of the pollen grains or by direct observation of the physical characteristics of the loads (Hidalgo et al., Citation1990). The palynological composition of the pollen loads is a reflection of the local flora surrounding the beehive (Díaz-Losada et al., Citation1998). Moreover, the use of pollen traps is more accurate than field observations regarding the blooming periods and gives information about the available pollen sources and their contribution to the colony (Dimou & Thrasyvoulou, Citation2007). Nevertheless, weather and seasonal variations affect pollen collection by A. mellifera (Andrada & Tellería, Citation2005; Dimou et al., Citation2006).
Mallorca, one of the 5000 islands and islets of the Mediterranean basin, is the largest island of the archipelago of the Balearic Islands (Spain). These islands provide a large variety of different sizes, geologies and altitudes resulting in the existence of a great diversity of habitats. Mallorca has an area of 3800?km2 with a maximum altitude of 1445?m above sea level, including two mountain formations, the Sierra de Tramuntana and the Sierra de Llevant, with a vast central plain between them. The native vegetation of the island is similar to that of other Mediterranean islands and coastal systems, with certain characteristics in common, such as the predominance of sclerophyllous evergreen species, which have water saving devices and mechanisms to prevent herbivory. These have been adapted for the Mediterranean climate, which is characterised by the alternation of two main seasons. The essential features of this climate are hot and dry summers, which results in a significant water stress for the plants, and another wet and cool season, during which unpredictable and heavy rains can occur from fall to spring. The relative duration of summer drought and the amount and timing of rainfall in the wet season allow a large spatial variation in the entire Mediterranean basin. The alternation of these two seasons integrates each region, its landscape and its flora (Thompson, Citation2005).
The most common classes of forest and garrigue of Majorca are Quercetea ilicis, Rosmarinetea officinalis and Rhamno-Prunetea. These vegetation classes are the mature stages of the vegetation series, the climax of which are the oak woodlands (Quercion ilicis). The class Quercetea ilicis is the island’s most abundant vegetation type and includes two different landscape formations: Quercion ilicis (oak forests) and Oleo-Ceratonion (shrubby Mediterranean garrigue). The most characteristic species of Quercion ilicis is Quercus ilex with other secondary shrubs like Osyris alba, Smilax aspera or Viburnum tinus. By contrast, Oleo-Ceratonion has a higher diversity of character species (Olea europaea, Pistacia lentiscus, Rhamnus alaternus and Myrtus communis), because it presents a higher variety of forest and garrigue formations. Rosmarinetea officinalis is represented by low shrubby formations located in calcareous shallow soils. Some of its character species are low shrubs, such as Rosmarinus officinalis, Cistus spp., or Erica spp. The class Rhamno-Prunetea includes thorny shrub formations and little forests, which are often located at the boundaries of farming zones. Some of its character species are Crataegus monogyna, Rubus ulmifolius, Rosa sempervirens and Prunus spinosa.
Some of the most common herbaceous classes of Mallorca are Artemisietea vulgaris, Festuco-Brometea, Galio-Urticetea, Helianthemetea guttati, Lygeo-Stipetea and Molinio-Arrhenatheretea. The class Artemisietea vulgaris is a hemicryptophyte nitrophilous vegetation associated with livestock uses. Some of its character species are Centaurea calcitrapa, Convolvulus arvensis, Foeniculum vulgare and Silybum marianum. Festuco-Brometea is a dry grassland vegetation dominated by perennial gramineous plants with mesophyll character, such as Brachypodium phoenicoides or Hypericum perforatum. The class Galio-Urticetea is a nitrophilous perennial vegetation with mesophytic character, which is associated with humid or disturbed soils. Some of its character species are Allium triquetrum, Calystegia sepium and Smyrnium olusatrum. Helianthemetea guttati is dry grassland vegetation dominated by terophyte or geophyte plants such as Merendera filifolia, Trifolium campestre and Medicago minima. Lygeo-Stipetea is a dry grassland vegetation dominated by perennial gramineous plants such as Brachypodium retusum or Dactylis hispanica. It additionally includes some orchid species such as Ophrys spp. or Serapias spp. Finally, the class Molinio-Arrhenatheretea is a hemicryptophyte vegetation with a slightly nitrophilous character that is associated with temporarily flooded soils. Some of its character species are Mentha suaveolens and Ranunculus spp. (Rivas-Martínez et al., 2001).
Human activity has created massive changes in the spatial configuration of Mediterranean habitats and ecological conditions (Thompson, Citation2005). As a result of these changes, most forests, garrigues and calcareous dry grasslands (Festuco-Brometea and Helianthemetea guttati) have been restricted to non-agricultural areas of the island. In contrast to such restrictions, annual grasslands (Artemisietea vulgaris, Polygono-Poetea annuae and Stellarietea mediae) are associated with dry fruit crops, so they have a changing variety of blooming species throughout the year due to the influence of farming management and seasonal changes. The most abundant dry fruit crops in Mallorca are carob (Ceratonia siliqua) and almond (Prunus dulcis).
We are, at least in Spain, not aware of any palynological studies, in which species are attributed to plant communities of the natural or anthropic landscape that act as pollen sources. In fact, Boi et al. (Citation2008) identified more than 200 pollen taxa in Mallorca, but they did not take the vegetation class, in which those taxa were a characteristic, into account. In our view, the combination of palynological and phytosociological studies could be a new way to identify the most interesting pollen sources of a landscape. We think that our study could be applied to all landscapes of the Mediterranean basin, and possibly to other Mediterranean landscapes from around the world. Thus, it could help beekeepers, at least from Mediterranean climate zones, to ensure an adequate yield of their hives. The main objective of this study is to estimate the polleniferous potential of different plant communities of a Mediterranean landscape throughout the year by combining an analysis of the pollen collected by Apis mellifera with a phytosociological study of the area. The secondary objective is to assess the possible influence of environmental factors (climatic, meteorological and anthropogenic) on the vegetation potential as a pollen source.
Material and methods
Study area
We defined a circular area of a 2?km radius (1256.6 hectares) around the apiary, because it is the most probable area frequented by Apis mellifera, according to the method proposed by Hidalgo et al. (Citation1990). The apiary was located on a farm, in the west of Mallorca, the coordinates being 39° 40′ 0.72″ N, 2° 38′ 52.32″ E (). The farm of 100 hectares cultivates the same ratio of almond trees (Prunus dulcis) and carob trees (Ceratonia siliqua), together with annual grasslands, which are harvested as pasture for sheep. This is the typical farm management system on the island. The rest of the foraging area includes other farms, native forests dominated by sclerophyllous scrubs and urban areas with gardens. The location was chosen as being representative of the central plain of the island, characterised by a landscape mosaic of natural vegetation, agricultural and urban areas.
Figure 1. Topographic map of Mallorca showing the location of the apiary. The white square in the lower box indicates the position of the island in the western Mediterranean.
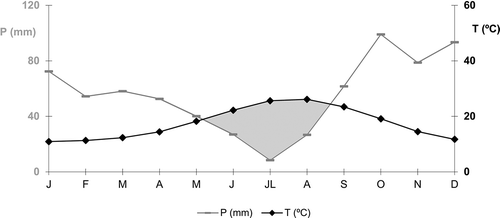
According to the bioclimatic model CLIBA2 (Guijarro, Citation1986–2012), the climate within coordinates of the apiary is Mediterranean, with a thermo-Mediterranean thermotype and a humid ombrotype. The average annual rainfall is 672?mm, the average annual temperature is 17.5?°C and there is a rainfall shortage from May to late August (). The terrain is mostly flat, interrupted in the north by hills of the neighbouring Sierra de Tramuntana, where the altitude varies between 100 and 447?m above sea level.
Figure 2. Ombrothermal diagram of the study area: P, average monthly rainfall (in millimetres); T, average monthly temperature (in °C). Grey area indicates water deficit.
Sampling and palynological analysis
Pollen load samples were collected from two Langstroth hives between October 2009 and September 2010, using a standard pollen trap placed at the entrance of each hive. A one yearlong study is sufficient to determine the polleniferous flora of an area and four or less hives are sufficient for a detailed study (Dimou et al., Citation2006). The collection of the samples was originally planned every two weeks, from dawn to dusk, according to Díaz-Losada et al. (Citation1998), but in the case of bad weather for the bee foraging activity (hard rain or wind), the collection was postponed to the next suitable day.
The preparation of microscopic pollen samples and their botanical identification were made according to the method proposed by Dimou and Thrasyvoulou (Citation2007). The variations are detailed later: pollen loads from both hives were collected separately and the impurities were eliminated. The subsequent drying process was performed at 40?°C for eight hours. Then, the sample from each hive was homogenised and separated into representative subsamples of 12?g (dry weight). When the weight of the total sample was lower than this amount, we analysed the totality. Pollen loads from each subsample were manually separated according to colour tone, texture and morphology, and each load type was identified and assigned to a plant taxon through its microscopic analysis. At least five pollen loads of each colour tone were mounted in glycerine jelly without acetolysis, to identify the botanical origin of each separated colour fraction. Then, we counted at least 300 pollen grains of each slide to verify the homogeneity of the pollen type for each colour tone.
The most general taxonomic level assigned to each load type was family, followed by pollen type (marked ‘Type’), then by genus and finally species. The pollen atlases consulted to assign the botanical level were Reille (Citation1999), Bucher et al. (Citation2004) and Boi and Llorens (Citation2007). To identify the species level, we compared hive samples with pollen obtained directly from flowering species recorded near the hives on sampling days. We also consulted the pollen reference collection of the Department of Botany at the University of the Balearic Islands. For taxonomic classification of the families of pollen types, we used as a reference the Flora Ibérica (Castroviejo, Citation1986–2012).
Finally, the preparations of each identified taxon were labelled with the sampling date and hive, and were then included in the pollen collection index of the study. This was used in addition to the reference pollen collection of the Department of Botany of the University of the Balearic Islands.
Phyto-sociological study
Many field visits and observations were made to identify the different vegetation classes that made up the plant landscape of the foraging area; subsequently, the vegetation coverage of each one was calculated. The vegetation class is generally accepted as the highest category in vegetation taxonomy (Pignatti et al., Citation1995). Previously, we made a vegetation map on a 1:20?000 scale from an aerial photograph, in which we identified different vegetation patches, which then needed to be verified in the field. This landscape analysis was based on the phyto-sociological Sigmatist method proposed by Braun-Blanquet (Citation1979). Each patch on the map included various vegetation classes with different coverages. The sum of all the different coverages of the map had a total value higher than the real surface of the foraging area. This occurs because the coverage of different vegetation classes often overlaps. Garden vegetation was treated as though its vegetation coverage was the entire surface of all the corresponding urban area, due to the difficulty in calculating the real area of the garden.
Subsequently, we assigned each pollen taxon identified in the hives to a vegetation class, applying the criterion that the taxon had to be characteristic of the respective class. Vegetation classes, following the tradition, are defined mainly on the basis of character species (Pignatti et al., Citation1995). The study of vegetation series of Spain by Rivas-Martínez (Citation1987) and the study of the vegetation of Mallorca by Llorens et al. (Citation2007) were taken as a reference, adapting the study with field observations when necessary. In the case of the pollen loads formed by two combined taxa, we considered if both taxa were character of the same vegetation class or not. In the second case, the criterion adopted was to assign half of the dry weight of the total load to each taxon, because it was impossible to separate accurately both pollen types.
We statistically treated crops and garden vegetation as true vegetation classes, although they are not. This allowed us to compare qualitative and quantitative data from the native and the anthropic vegetation of the area.
Meteorological study
To assess the influence of the weather on the polleniferous potential of the studied vegetation, we statistically analysed how the weather influenced flowering. The meteorological data used were: accumulated rainfall between samplings, average daily temperature, daily hours of sun, daily minimum relative humidity and daily average wind speed. These data corresponded to a station situated at the University of the Balearic Islands (2?km from the hives) and were provided by the Meteorological Agency (AEMET). Knowing the accumulated rainfall before taking the first sample, we calculated the average number of days between all sampling of the study (21.4 days) and used its corresponding rainfall values before the start of the study.
Statistical analyses
The variables ‘number of taxa’ and ‘dry weight of pollen collected’ were correlated with the meteorological variables. After checking the normality of all variables using the Kolmogorov–Smirnov test, we applied a Pearson correlation, whose results were indicated by the Pearson correlation coefficient (r), the number of variables analysed (n) and the level of significance (p). Statistical analyses of the data were performed using the SPSS software package, version 19.0.
Results
Results about flora and vegetation are presented separately to allow for different conclusions.
Polleniferous flora
In total, we collected 1039.7?g of pollen (dry weight): 412.4?g from hive A, coming from 60 pollen types, and 627.3?g from hive B, coming from 58 pollen types. By summing up both hives, 74 different pollen types were identified, out of which 44 were found in both colonies and the rest only in one. Furthermore, 66.2% of total pollen types were derived from native flora, 21.6% from exotic flora (gardens), 8.1% from crops and 4.1% from undetermined taxa. Moreover, 0.6% of the total weight (6.2?g) was coming from pollen loads formed by two combined taxa, either mixed (intermediate colour) or separated in the same load. We recorded the proportion of each pollen type in respect to the total weight, the botanical family, the flowering period and the vegetation class (). Only three of the 74 collected taxa were anemophilous (Olea eropaea, Pistacia lentiscus and Quercus ilex), reaching a 4.1% of the total pollen weight. The rest of the pollen (95.9%) came from entomophilous taxa.
Only eight pollen types individually provided more than 1% of the total weight of the study (104?g), accounting for 19.5% and being common to both hives. They were, in descending order: Ceratonia siliqua (6.7%), Prunus dulcis (3.9%), Quercus ilex (2.4%), Diplotaxis erucoides (1.5%), Cistus albidus (1.4%), Rhamnus alaternus (1.3%), Olea europaea (1.2%) and Oxalis pes-caprae (1.1%). Furthermore, the analysis of samples revealed that the botanical families Compositae and Rosaceae were the most represented with ten and eight taxa, respectively, followed by Liliaceae and Labiatae with five and four taxa. These were the four families with the highest taxonomic diversity, accounting for 36.5% of the total pollen types. It is also noted that 19 of the 33 families were represented by a single pollen type, five families by two types and five families by three types. However, when we analysed the importance of these families by the amount of pollen they provided, Leguminosae was the most polleniferous because it included Ceratonia siliqua as a character species, despite the fact that it was only represented by three taxa ().
The pollen weight in both hives held a statistically significant relation (r?=?0.898, n?=?2, p?=?0), as well as the number of pollen types (r?=?0.868, n?=?2, p?=?0). Despite this, both parameters followed different trends in each hive, so there was no statistically significant relation between them (). However, when we studied the relation of these two parameters with the main meteorological variables, there was a statistically significant relation between the dry weight of pollen and the accumulated rainfall (hive A: r?=?0.777, n?=?17, p?=?0; hive B: r?=?0.652, n?=?17, p?=?0.005).
Figure 3. Weight of pollen in grams and number of taxa collected in both hives A and B during the year (above). Accumulated daily rainfall (mm), daily hours of sun and daily mean temperature (°C) per sampling (below).
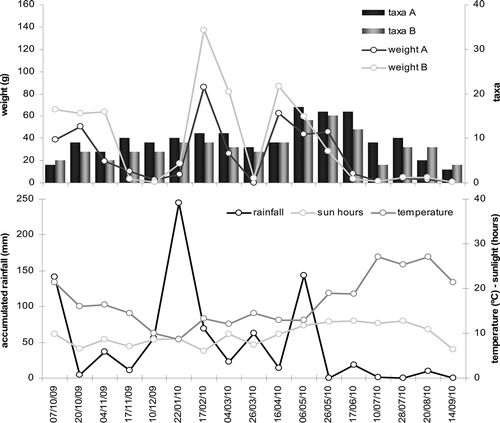
A significant relationship between the number of taxa and the hours of sun was revealed (hive A: r?=?0.487, n?=?17, p?=?0.047; hive B: r?=?0.475, n?=?17, p?=?0.054) as well as significant negative relation between the average daily temperatures and the accumulated rainfall (r?=?–0.480, n?=?17, p?=?0.051) throughout the year. However, there were no other relations among the rest of the meteorological variables.
The majority of the pollen (63.6%) was collected at the end of the winter and in spring (February–May). Three great increases of weight were observed in the annual pollen fluctuation (). Taking both hives into account, the greatest amount of pollen was recorded in February 2010 (17/02/10) with 223.4?g (21.5% of total), of which 84.3% corresponded to Prunus dulcis. Another remarkable value was recorded at the beginning of spring 2010 (16/04/10) with 148.5?g (14.3% of total). In October 2009 (20/10/09), 112.9?g (10.9%) of pollen were collected, of which 85.4% belonged to Ceratomia siliqua. In contrast, the lowest values of the year were recorded in September 2010 (14/09/10) with 2.1?g (0.2% of total) and in December (10/12/09) with 3.4?g (0.3% of total). Between 17/06/10 and 14/09/10 was a long period of scarcity of pollen, coinciding with the summer drought. Regarding the number of pollen types () and considering both colonies, the highest peak of the year was recorded in May 2010 (06/05/10 and 26/05/10) with 31 species (41.9% of total). In contrast, seven taxa (4.9% of total) was the minimum, recorded in September 2010 (14/09/10).
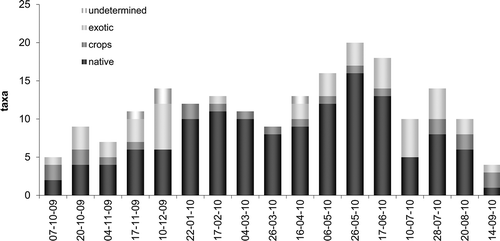
We furthermore calculated the annual curve of collected pollen taxa in relation to their origin (native, crops, exotic and undetermined) by adding up both hives (). On the one hand, it shows the maximum of native taxa in late spring, with a secondary peak in late autumn and a remarkable lack of native pollen in late summer and early autumn. On the other hand, it shows how pollen of exotic species appeared irregularly throughout the year, while pollen types from crops were collected more regularly.
Figure 4. Pollen annual curve based in taxa origin (native, exotic, crop and undetermined) of the pollen types collected in both hives.
Polleniferous vegetation
The vegetation study included the elaboration of a vegetation map of the bee foraging area superimposed on an aerial photograph, in which we identified 27 vegetation patches. Subsequently, we went to check these results in the field and we identified different vegetation classes, which were combined in each patch. Thus, we identified 12 different vegetation classes within the foraging area (namely Stellarietea mediae, Quercetea ilicis, crops, gardens, Lygeo-Stipetea, Rosmarinetea officinalis, Helianthemetea guttati, Artemisietea vulgaris, Festuco-Brometea, Galio-Urticetea, Molinio-Arrhenatheretea and Rhamno-Prunetea). Finally, we calculated the coverage of the identified classes of each patch, so we obtained a total coverage value for each class. The total vegetation coverage of the foraging area was 1634.3 hectares, which included the following coverages (decreasing order): Stellarietea mediae (31.5%), Quercetea ilicis (25.5%), crops (18.6%), gardens (7.6%), Lygeo-Stipetea (6.4%), Rosmarinetea officinalis (5.5%), Helianthemetea guttati (2.7%), Artemisietea vulgaris (1.8%), Festuco-Brometea (0.1%), Galio-Urticetea (0.1%), Molinio-Arrhenatheretea (0.1%) and Rhamno-Prunetea (0.1%) ().
Table I. Pollen types collected during the year, according to the model of Dimou and Thrasyvoulou (Citation2007) with the addition of the vegetation classes
To assess the polleniferous potential of the different vegetation classes, we compared the dry weight of the pollen values and the number of pollen types of each class with the respective vegetation coverage (). The weight of each class was calculated by adding the total weight of its taxa. Thus, the dry weight values of the pollen were assigned to the different classes. They were, in decreasing order: 298.9?g of crops, 248.4?g of Quercetea ilicis, 246.2?g of Stellarietea mediae, 143.2?g of gardens, 43.5?g of Rosmarinetea officinalis, 37.0?g of Rhamno-Prunetea, 18.5?g of Artemisietea vulgaris, 3.0?g of Molinio-Arrhenatheretea and 0.9?g of Galio-Urticetea. Pollen types that could not be taxonomically identified accounted for less than 0.1% of the total weight. Furthermore, we calculated the number of pollen types that belonged to each vegetation class. They were, in descending order: 20 of Stellarietea mediae, 16 of gardens, nine of Quercetea ilicis, seven of Rosmarinetea officinalis, six of Artemisietea vulgaris, six of crops, three of Molinio-Arrhenatheretea, three of Rhamno-Prunetea and one of Galio-Urticetea. Three other pollen types could not be identified accounting for 4.1% of the whole study ().
In addition, we analysed the statistical correlation between the vegetation coverage, the dry weight of pollen and the number of pollen types for the nine vegetation types that provided pollen to hives. We observed a positive relationship between coverage and pollen weight (r?=?0.908, n?=?9, p?=?0.01). The same relationship existed also among coverage and number of pollen taxa (r?=?0.637, n?=?9, p?=?0.065).
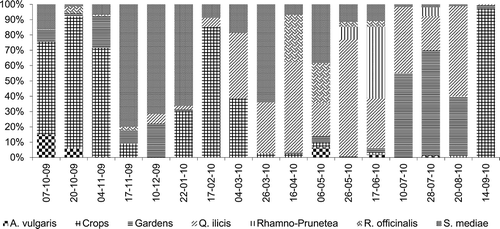
We also studied the relative importance of a polleniferous source of the different vegetation classes throughout the year, excluding classes that provided less than 1% of the total weight of pollen (Galio-Urticetea and Molinio-Arrhenatheretea). There was a predominance of pollen from species that belonged to Quercetea ilicis in spring and summer, and from crops in autumn (September–November) and in February. Pollen from the Stellarietea mediae appeared during most of the year, but peaked from November to January and in March. Pollen from gardens predominates in late summer (July–August), which was a period of low pollen collection. To a lesser extent, Rosmarinetea officinalis provided pollen in late spring as did Rhamno-Prunetea in early summer. Finally, the least abundant pollen was from Artemisietea vulgaris, which to some extent was present in October and early summer ().
Figure 5. Pollen proportion (%) of the vegetation classes that provided more than 1% of the total weight.
Discussion
Polleniferous flora
Both hives showed a similar pattern of pollen collection throughout the year, represented in dry weight and number of pollen types, in accordance with Dimou et al. (Citation2006). Nevertheless, both parameters followed different trends. Honeybees gathered a high diversity of pollen types during the year, especially in May and June, which were the richest months in terms of pollen taxa, in accordance with Dimou and Thrasyvoulou (Citation2007). In this respect, Kummerow (Citation1983) and Herrera (Citation1988) reported that spring is the time of greatest diversity of flowering species and of pollinating insects in the Mediterranean ecosystems. In addition, Thompson (Citation2005) argued that Mediterranean plant communities are characterised by an annual pattern of flowering phenology with a very remarkable peak in spring and another one in autumn. These two peaks were also observed in the annual pollen collection curve obtained in our study, so that we can say that the studied flora had a typical Mediterranean flowering curve. This fact is clear in the native flora pollen curve, while pollen types from crops and gardens follow a different collection pattern, supposedly because they were managed by humans, so their flowering did not depend only on the weather conditions. However, both peaks appear a few weeks later than expected, possibly because they were prompted by great rainfalls. Nevertheless, these results suggest that the analysis of pollen collected by Apis mellifera during the year could be a good method to study the entomophilous flora of an area, thus providing us with reliable knowledge about plant diversity and the annual flowering pattern.
Table II. Inventory table with the relative coverage (%) and vegetation classes identified in each patch on the vegetation map
A proof of the richness of species collected in our study is the fact that more than half of the 33 identified botanical families were represented by only one pollen taxon. In addition, the four most diverse botanical families (Compositae, Rosaceae, Labiatae and Liliaceae) together provided only 36.5% of the total pollen types. However, despite the high diversity of pollen collected, only a few species were collected intensively, as other authors described before (Percival, Citation1965; Louveaux, Citation1968; Hidalgo et al., Citation1990; Andrada & Tellería, Citation2005; Dimou & Thrasyvoulou, Citation2007). The species collected most intensively in our study have also been highlighted in previous studies: Ceratonia siliqua and Prunus dulcis (Ortega, Citation1987; Ortiz et al., Citation1996), which produce large amounts of pollen grains per flower and per area (Hidalgo et al., Citation1996); the genus Quercus (Ortiz & Fernández, Citation1992; Ortiz, Citation1994; Hidalgo et al., Citation1996; Andrés et al., Citation2004) and specifically Q. ilex (Ortega, Citation1987); Olea europaea and Rhamnus alaternus (Andrés et al., Citation2004); Cistus albidus, both as a genus (Talavera et al, Citation1988; Ortiz, Citation1994; Hidalgo et al., Citation1996) and as a species (Bonet et al., Citation1986; Ortega, Citation1987; Talavera et al., Citation1988, Hidalgo et al., Citation1990, 1996); Diplotaxis erucoides and Oxalis pes-caprae (Ortega, Citation1987).
The most polleniferous species were Ceratonia siliqua and Prunus dulcis, which are widely farmed around the hives. Moreover, in field observations we observed a large number of flowers per stem or branch and stamens per flower in both of them. In addition, they both belong to families of special polleniferous interest (Ortega, Citation1987). In relation to this, Freitas and Silva (Citation2006) noted that species of interest usually have intense and abundant blooms both in plant and population terms. The other prominent species (Quercus ilex, Diplotaxis erucoides, Cistus albidus, Rhamnus alaternus, Olea europaea and Oxalis pes-caprae) are part of the flora and belong to phyto-sociological classes with high coverage in the area. Díaz-Losada et al. (Citation1998) and Dimou and Thrasyvoulou (Citation2007) confirmed our results affirming that the pollen types collected in higher percentages correspond to abundant plants of the local flora or are the most cultivated in the area and Bilisik et al. (Citation2008) asserted that most of them were crops. Meanwhile, De Sá et al. (Citation2004) noted that Apis mellifera collect greater amounts of pollen from species with large populations, especially if they offer abundant and continuous blooms. In our case, this happened particularly in autumn with Ceratonia siliqua and in winter with P. dulcis, which prevailed over the pollen of native species, either by their high coverage near the hives or by their dense flowering or both features at once. Thus, our results suggest that the existence of species with large population and abundant bloom near the hives influence the variation of the amount and taxonomic diversity of pollen collected during the year.
However, there was an annual fluctuation of pollen collected from beehives both quantitatively and qualitatively, which we mainly attribute to the effect of weather conditions on the flowering phenology. The pollen weight had considerable peaks at various times during the study and we think that it was due to the increase of the flowering in the area in response to rainfall. Although that response occurred with a few weeks delay, we observed that pollen peaks coincided approximately with the next sampling day. Moreover, there was a prolonged shortage of entry of pollen in the hives between June and September of 2010, which we attribute to a summer drought, characteristic of the Mediterranean climate (Thompson, Citation2005). This shortage correlated with the highest temperatures and the lowest rainfalls of the year. Thus, our results suggest that accumulated rainfall and temperature, together with summer drought, have influence on the amount of pollen collected by Apis mellifera throughout the year by affecting the flowering phenology.
The fact that the number of pollen types collected was directly related to sun hours seems to be another evidence of the influence of weather on the polleniferous flora. On the one hand, pollen types diversity increased during the spring, as other authors previously described for the Mediterranean basin (García-García et al., Citation2001; Dimou & Thrasyvoulou, Citation2007). Moreover, the highest diversity was collected in late spring, coinciding with the longest photoperiod of the year. On the other hand, from July to September 2010 was the lowest collected diversity period, which seemed to be caused by the summer drought. In relation to this, Thompson (Citation2005) stated that summer drought imposes a restriction on the flowering period on plants. This could also explain the low diversity of pollen types collected in late summer, coinciding with Bilisik et al. (Citation2008) and Novais et al. (Citation2009). Thus, our results suggest that the diversity of pollen types collected during the year is conditioned by the effect of daily sun hours and the summer drought on flowering phenology.
Table III. Vegetation types of the study area, arranged in order of decreasing vegetation cover (%), with the absolute and relative dry weight values of pollen and the number of taxa harvested
Polleniferous vegetation
Vegetation classes with the greatest coverage were also the most polleniferous and, in general, the richest in pollen diversity. This let us to believe that the more extensive vegetation units may provide higher amounts and diversity of pollen to pollinators. The selection of the polleniferous sources made by Apis mellifera is conditioned by the composition of the vegetation of the area, both in quantitative and diversity terms, as discussed by Dimou and Thrasyvoulou (Citation2007). So we conclude that, despite the selection of the available polleniferous sources made by honeybees, the analysis of the pollen loads provides vegetation data of the area where the hives are situated in, as also concluded by Díaz-Losada et al. (Citation1998).
Wild vegetation was harvested as a pollen source during the year, although the maximum quantity and diversity values were recorded in the spring. The class Quercetea ilicis was the most polleniferous of the wild vegetation classes, together with Stellarietea mediae providing about half the total pollen amount. Quercetea ilicis pollen was especially present in spring and summer due to some highly polleniferous species (Quercus ilex and Olea europaea) and other accompanying shrubs (Chamaerops humilis, Myrtus communis and Osyris alba). They also provided pollen to the hives in the autumn and winter, although in small quantities, especially through Rhamnus alaternus and other less polleniferous species (Viburnum tinus and Smilax aspera). However, Quercetea ilicis had low pollen diversity in relation to its vegetation coverage. Thus, this class seems to present the highest polleniferous potential of the landscape in spring and a higher one than most of the other classes in summer. In contrast, the potential was quite low, or absent, during the rest of the year.
Stellarietea mediae was the only vegetation class that provided pollen throughout the year and produced the greatest diversity, so it seems to be very important as a polleniferous source. We think that it is due to the fact that it is a class represented by annual herbaceous species with blooms that can last for several months, so they overlap during the year. Unlike other wild vegetation, Stellarietea mediae is associated with crops, so its coverage and taxa composition may vary according to farming management (tillage, pasture, fallow, etc.). Its pollen had a strong presence in late fall, winter and spring, while in summer it appeared briefly. Thus, its potential as a pollen source seems to be limited by the summer drought. We identified Diplotaxis erucoides as a character species of this vegetation class, which stood out as one of the most polleniferous taxon, probably due to its abundance in the area and to its polleniferous capacity (Ortega, Citation1987). Therefore, this seems to present the highest potential of the landscape in winter, being moderate or quite low for the rest of the year.
The class Rosmarinetea officinalis showed high taxa diversity in relation to its coverage, so it seems to be a good polleniferous resource of wild vegetation. It was also a main pollen source in spring, especially due to Cistus albidus, which was one of the most polleniferous taxa. It has a long flowering period and a high pollen production per flower (Ortega, Citation1987; Hidalgo et al., Citation1996). These results suggest that this is a class with higher polleniferous potential than most of the other classes in spring, and is low or absent for the rest of the year.
Artemisietea vulgaris class was a minor polleniferous resource that provided only a small amount of pollen according to its limited coverage. However, it was primarily represented by summer flowering species, which gives importance to this vegetation class, because wild flowering is scarce in summer. The class Rhamno-Prunetea presented a much smaller coverage, because it is associated to hygrophilous terrains. Despite this, its pollen was predominant in early summer, giving importance to this vegetation class, because it provides pollen when spring flowering has finished. It is probably due to the fact that it is represented by taxa of the family Rosaceae, which have a high polleniferous capacity (Ortega, Citation1987). Thus, these classes seem to present a moderate polleniferous potential in comparison with the rest of the classes in late spring and summer, and are low or absent for the remainder of the year.
The classes Galio-Urticetea and Molinio-Arrhenatheretea also provided pollen to the hives, although they did it in very little amounts and at specific times, according to their low coverage, so they may be considered minor pollen sources. Alternatively, we identified other three vegetation classes (Lygeo-Stipetea, Helianthemetea guttati and Festuco-Brometea) that were not harvested as polleniferous sources at all, despite the fact that together they exceeded 9.2% of the total vegetation coverage. These results suggest that wild vegetation classes with predominance of anemophilous taxa do not provide sources of pollen, so its polleniferous potential is zero.
However, both vegetation classes managed by man (crops and gardens) together provided more than 40% of the total amount of pollen and almost a third of the pollen diversity during the year, despite the fact that the sum of their coverage just exceeded a quarter of the total. This indicates the great influence of human activity on the plant landscape of the area, especially through the introduction of dry farming (Ceratonia siliqua and Prunus dulcis), which stood out as important pollen sources in autumn and winter. The high polleniferous potential of these classes seems to be due to its high coverage near the hives and because they are species with a high pollen production (Ortega, Citation1987; Hidalgo et al., Citation1996). However, they are monocultures, so provided low pollen diversity compared with their high coverage. Despite this, our results suggest that this vegetation class seems to present the highest polleniferous potential of the landscape in autumn, and one of the highest in winter.
Other evidence of human influence on vegetation composition is the high presence of pollen from gardens, especially in late summer, which is a time when there is a scarcity of wild flowers. We attribute this to the fact that this vegetation is not affected by summer drought, because it is basically located in urban areas with watering systems. Thus, most of its taxa did not follow the typical Mediterranean flowering curve. Another characteristic of the garden vegetation class was the high taxa diversity that it provided compared to its coverage, which we attribute to the fact that it was represented by plants with very different origin from around the world. For all these reasons, gardens were an interesting pollen source, especially in periods of wild flowering scarcity. Thus, the polleniferous potential of this class seems to be the highest of the landscape in summer, and is quite low for the rest of the year.
Conclusions
Apis mellifera collected a quantity and diversity of pollen throughout the year that can be considered a reliable representation of the polleniferous flora of the study area. During the year, there appears to be different pollen sources from natural landscapes or agricultural and urban areas that overlap or follow one another. Our results suggest that when a vegetation class presents a higher coverage than other classes, it may provide more quantity and diversity of pollen to pollinators, and therefore its polleniferous potential may be higher. Our results also suggest that weather and summer drought may condition the polleniferous potential of vegetation by affecting its flowering phenology. Nevertheless, we believe that further studies may help us to understand more deeply the role of the different vegetation classes as pollen sources in the landscape of Mediterranean climate zones from around the world.
Specimens investigated
Artemisietea vulgaris Lohmeyer, Preising & Tüxen ex von Rochow 1951
Festuco-Brometea Br.-Bl. & Tüxen ex Br.-Bl. 1949
Galio-Urticetea Passarge ex Kopecky 1969
Helianthemetea guttati (Br.-Bl. in Br.-Bl., Roussine & Nègre 1952) Rivas Goday & Rivas-Martínez 1963 em. Rivas Martínez 1978
Lygeo-Stipetea Rivas-Martínez 1978 nom. conserv.
Molinio-Arrhenatheretea Tüxen 1937
Quercetea ilicis Br.-Bl. ex A. & O. Bolòs 1950
Rhamno-Prunetea Rivas Goday & Borja ex Tüxen 1962
Rosmarinetea officinalis Rivas-Martínez, T.E. Díaz, F. Prieto, Loidi & Penas classis nova
Stellarietea mediae Tüxen, Lohmeyer & Preising ex von Rochow 1951
Acknowledgements
The authors are grateful to Gregori Lladó for his generous collaboration as beekeeping advisor, to José María Castro for the realisation of , and to Blaize Benson for the correction of the English language.
Related Research Data
References
- Andrada, A. C. & Tellería, M. C. (2005). Pollen collected by honey bees (Apis mellifera L.) from south of Caldén district (Argentina): Botanical origin and protein content. Grana, 44, 1–8.
- Andrés, C., Díez, M. J. & Terrab, A. (2004). Análisis polínico de mieles de los Parques Naturales Sierra Norte de Sevilla y Sierras Subbéticas. Lazaroa, 25, 125–133.
- Bilisik, A., Cakmak, I., Bicakci, A. & Malyer, H. (2008). Seasonal variation of collected pollen loads of honeybees (Apis mellifera L. anatoliaca). Grana, 47, 70–77.
- Boi, M., Lladó, G. & Llorens, L. (2008). Estudio de la flora melífera, mieles y producción polínica de Majorca. Palma: Mallorca Rural, Leader?+?.
- Boi, M. & Llorens, L. (2007). Atlas polínico de las Baleares. Flora Endémica. Palma: Associació Jardí Botànic de Palma.
- Bonet, A., Sebastia, M. T. & Rita, J. (1986). Principales floraciones melíferas de la provincia de Barcelona. Actas del II Congreso Nacional de Apicultura, Gijón, Spain, 5–9 December 1984.
- Braun-Blanquet, J. (1979). Fitosociología. Bases para el estudio de las comunidades vegetales. Madrid: Blume.
- Bucher, E., Kofler, V., Vorwohl, G. & Zieger, E. (2004). Lo spettro pollinico dei mieli dell’Alto Adige. Laives: Laboratoro Biologico, Agenzia Provinciale per la Protezione dell’Ambiente e la Tutela del Lavoro.
- Butler, C. G. (1945). The influence of various physical and biological factors of the environment on honeybee activity. An examination of the relationship between activity and nectar concentration and abundance. Journal of Experimental Biology, 21, 5–12.
- Castroviejo, S. (1986–2012). Flora Ibérica. Madrid: Real Jardín Botánico, CSIC.
- De Sá, M. P., Díaz Losada, E. & Armesto Baztán, S. (2004). Flower election by honeybee and floral morfology. Lazaroa, 25, 113–123.
- Díaz-Losada, E., Ricciardelli-d’Albore, G. & Saa-Otero, M. P. (1998). The possible use of honeybee pollen loads in characterising vegetation. Grana, 37, 155–163.
- Dimou, M. & Thrasyvoulou, A. (2007). Seasonal variation in vegetation and pollen collected by honeybees in Thessaloniki (Greece). Grana, 46, 292–299.
- Dimou, M., Thrasyvoulou, A. & Tsirakoglou, V. (2006). Efficient use of pollen traps to determine the pollen flora used by honey bees. Journal of Apicultural Research and Bee World, 45, 42–46.
- Freitas, B. M. & Silva, E. M. S. (2006). Potencial apícola da vegetação do semi-árido brasileiro. In F. A. R. Santos (Ed.), Apium Plantae (Vol. 3, pp. 19–32). Recife: IMSEAR.
- García-García, M. C., Ortiz, P. L. & Díez, M. J. (2001). Pollen collecting behaviour of Apis mellifera during one day. Grana, 40, 205–209.
- Guijarro, J. A. (1986–2012). Contribución a la bioclimatología de Baleares. Palma: Universidad de les Illes Balears, PhD Diss.
- Herrera, J. (1985). Nectar secretion patterns in southern Spanish Mediterranean shrublands. Israel Journal of Botany, 34, 47–58.
- Herrera, J. (1988). Pollination relationships in southern Spanish Mediterranean shrublands. Journal of Ecology, 76, 274–287.
- Hidalgo, M. I., Bootello, M. L. & Pacheco, J. (1990). Origen floral de las cargas de polen recogidas por Apis mellifera L. en Alora (Málaga, España). Acta Botánica Malacitana, 15, 33–44.
- Hidalgo, M. I., Recio, M. & Cabezudo, B. (1996). Producción de polen en un matorral del sur de España. Acta Botánica Malacitana, 21, 49–55.
- Kummerow, J. (1983). Mediterranean type ecosystems. The role of nutrients. In F. J. Kruger, D. T. Mitchell & J. V. M. Jarvis (Eds), Comparative phenology of Mediterranean-type plant communities (pp. 300–317). Berlin: Springer.
- Llorens, L., Gil, L. & Tébar, F. J. (2007). La vegetació de l’illa de Mallorca. Bases per a la interpretació i gestió d’hàbitats. Palma: Associació Jardí Botànic de Palma.
- Louveaux, J. (1968). Étude expérimentale de la récolte du pollen. Traité de biologie de l’abeille. París: R. Chauvin.
- Novais, J. S., Lima, L. C. & Ribeiro, F. A. (2009). Botanical affinity of pollen harvested by Apis mellifera L. in a semi-arid area from Bahia (Brazil). Grana, 48, 224–234.
- Ortega, J. L. (1987). Flora de interés apícola y polinización de cultivos. Madrid: Mundi-Prensa.
- Ortiz, L. (1994). El polen recogido por Apis mellifera L. en Hinojos (Huelva) durante la primavera. Acta Botánica Malacitana, 19, 115–122.
- Ortiz, L., Arista, M. & Talavera, S. (1996). Producción de néctar y frecuencia de polinizadores en Ceratonia siliqua L. (Caesalpinaceae). Anales del Jardín Botánico de Madrid, 54, 540–646.
- Ortiz, L. & Fernández, I. (1992). Estudio microscópico de miel y polen apícola de la provincia de Sevilla. Acta Botánica Malacitana, 17, 183–193.
- Percival, M. (1965). Floral biology. London: Pergamon Press.
- Petanidou, T. & Vokou, D. (1990). Pollination and pollen energetics in Mediterranean ecosystems. American Journal of Botany, 77, 986–992.
- Pignatti, S., Oberdorfer, E., Schaminée, J. H. J. & Westhoff, V. (1995). On the concept of vegetation class on phytosociology. Journal of Vegetation Science, 6, 143–152.
- Potts, S. G., Petadinou, T., Roberts, S., O’Toole, C., Hulbert, A. & Willmer, P. (2006). Plant-pollinator biodiversity and pollination services in a complex Mediterranean landscape. Biological Conservation, 129, 519–529.
- Reille, M. (1999). Pollen et spores d’Europe et d’Afrique du nord. Marsella: Labotatoire de Botanique Historique et Palynologie.
- Ribbands, C. R. (1949). The foraging method of individual honey-bees. Journal of Animal Ecology, 18, 47–66.
- Rivas-Martínez, S. (1987). Memoria del mapa de series de vegetación de España. Madrid: ICONA, Miniserio de Agricultura, Pesca y Alimentación.
- Rivas-Martínez, S., Fernández-González, F., Loidi, J., Lousa, M. & Penas, A. (2001). Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobotanica, 14, 5–341.
- Sá-Otero, M. P., Marcial-Bugarín, S., Armesto-Baztán, S. & Díaz-Losada, E. (2002). Método de determinación del origen geográfico del polen apícola comercial. Lazaroa, 23, 25–34.
- Singh, S. (1950). Behavior studies of honeybees in gathering nectar and pollen, Cornell University. Agricultural Experiment Station Memoirs, 288, 1–57.
- Talavera, S., Herrera, J., Arroyo, J., Ortiz, P. L. & Devesa, J. A. (1988). Estudio de la flora apícola de Andalucía Occidental. Lagascalia, 15, 567–591.
- Thompson, J. D. (2005). Plant evolution in the Mediterranean. Montpellier: Centre d’Ecologie Fonctionnelle et Evolutive.