
Abstract
Methane emissions from ruminants are the major single source of greenhouse gases in New Zealand and considerable effort has gone into the measurement and prediction of these emissions. This paper analyses methane emissions measured in respiration chambers from adult sheep and lambs fed fresh-cut pasture of different quality. Results show that the major driver of methane production (pCH4, g/d) is dry matter intake (DMI, kg/d). Pasture quality affected methane emissions only in animals less than 1 year of age. A single equation to be used for all sheep based on DMI alone (ln(pCH4) = 0.792(0.0275) ln(DMI) + 3.1(0.03)) or a two-equation approach with an equation for sheep >1 year based on DMI (ln(pCH4) = 0.826(0.0438) ln(DMI) + 3.15(0.017)) and one for sheep <1 year based on DMI and the metabolisable energy content (ME, MJ/kg) of the diet (ln(pCH4) = 0.749(0.0190)ln(DMI) + 0.051(0.0064)ME + 2.45(0.069)) is proposed.
Introduction
Agriculture in New Zealand accounts for 47% of all national CO2-equivalent greenhouse gas (GHG) emissions, and within the agricultural sector, enteric methane (CH4) emissions from ruminants, predominantly cattle and sheep, contribute more than two-thirds of the emissions (MfE Citation2014). To obtain accurate estimates of emissions from New Zealand ruminants, local conditions need to be taken into account, in particular the fact that the majority of New Zealand sheep consume a pasture-based diet 365 days a year with little supplementary feed. Currently, annual emissions from adult sheep >1 year in the New Zealand national inventory are estimated using a constant methane yield factor (Ym) (20.9 g CH4 kg dry matter intake [DMI]−1) multiplied by daily estimates of DMI. The current Ym value was the mean value obtained from a summary of New Zealand data where CH4 was estimated from experiments on grazing animals. Methane was measured using the sulphur hexafluoride (SF6) tracer technique and intake was estimated using indirect techniques (Clark et al. Citation2003). Methane measurements obtained using the SF6 tracer technique on groups of animals have generally been found to be more variable than those obtained using respiratory chambers (Vlaming et al. Citation2007; Pinares-Patino & Clark Citation2008) although group mean values generally compare well (Grainger et al. Citation2007; Pinares-Patino et al. Citation2008a; Pinares-Patiño et al. Citation2011). Despite many different techniques being available for estimating intake in grazing animals, the reliability of these techniques is questionable (Undi et al. Citation2008) meaning that there is large uncertainty around the Ym value currently used in the New Zealand inventory for adult sheep.
New Zealand—as required by all Annex 1 parties to the UN Framework Convention on Climate Change—is reporting its emissions using guidance from the International Panel on Climate Change (IPCC Citation1996, Citation2000). New Zealand has also adopted country-specific emissions methodology with a lower Ym value for emissions from animals <1 year of age, based on Clark et al. (Citation2003). In New Zealand, the value used for young sheep is 16.8 g CH4 kg DMI−1, 20% lower than the older sheep value. A crucial issue around the studies used to arrive at the New Zealand and IPCC (2006) values for young sheep, which are based on New Zealand data, is that they were not derived from studies where emissions from young sheep were directly compared with older sheep under the same experimental conditions. Rather, the values were obtained by comparing data from a small number of independent experiments on young animals. In common with data from sheep >1 year old, experiments on animals <1 year old used the SF6 tracer technique to measure methane and indirect methods to estimate intake (Lassey et al. Citation1997; Ulyatt et al. Citation2002). Given the limited number of studies with young sheep, the uncertainty around both the SF6 tracer technique and intake estimates, plus the findings from other studies that animal age does not influence emissions (Knight et al. Citation2008), there is an urgent need to verify under more controlled conditions that Ym values for sheep <1 year of age are lower than those for sheep >1 year of age.
Although methane emissions from animals are primarily driven by DMI (Grainger et al. Citation2007; Shibata & Terada Citation2010), the composition of the diet and the level of intake has an effect on methane emissions (Blaxter & Clapperton Citation1965; Reynolds et al. Citation2011; Hammond et al. Citation2013). The methane prediction equation of Blaxter & Clapperton (Citation1965), derived from a series of studies where animals were fed different diets at varying levels of intake above maintenance, relates methane yield to both level of intake and feed digestibility. Dietary chemical composition is also included in the prediction equations of Dijkstra et al. (Citation1992) and Ellis et al. (Citation2009). However, all these prediction equations have been derived from mixed forage:concentrate diets. To date, very little evidence exists to show that pasture chemical composition has a major impact on methane emissions from ruminants consuming fresh forage diets (Ulyatt et al. Citation2005; Molano & Clark Citation2008; Hammond et al. Citation2009; Hart et al. Citation2009).
As part of the effort to improve the New Zealand national agricultural GHG emissions inventory, a series of four experiments were conducted in which sheep of contrasting ages were fed fresh-cut pasture varying in quality at a range of feeding levels. These trials were all carried out indoors where methane emissions were determined in respiration chambers and DMI was accurately measured. The aim of these trials was to develop an updated methane prediction algorithm(s) suitable for use in the national inventory that accounts for the influence of factors such as DMI, pasture quality, level of feeding and animal age. Although it is convention under IPCC good practice guidelines (IPCC Citation2006) to estimate annual emissions using a separately estimated Ym factor and intake (DMI or energy intake), it is also possible to estimate emissions directly in a single step from intake and other variables. An advantage of using a Ym factor is that a common methodology can be adopted for estimating intake and a separate Ym factor adopted to adjust for different feeding situations. In New Zealand sheep, where the diet is primarily fresh forage, it may be simpler to predict methane emissions directly from intake and other variables, for example diet quality, without developing a separate Ym value. Both of these approaches were used when analysing the results obtained from the four trials undertaken.
Materials and methods
A total of four animal trials were carried out to assess the effect of level of feeding (Trials 1 and 2) and the effect of diet quality (Trials 3 and 4) in sheep <1 year old (Trials 1 and 4) and sheep >1 year old (Trials 2 and 3), on methane emissions from sheep fed fresh-cut pasture. All trials were approved by the AgResearch Grasslands Animal Ethics Committee (AE no. 11501, 11441, 11981 and 12502 for Trials 1 to 4, respectively) in accordance with the Animal Welfare Act 1999.
Diet preparation, animal handling and methane measurements
The diet offered to the sheep in all trials was ryegrass-dominant pasture harvested at AgResearch Aorangi Research Farm, Palmerston North, New Zealand. Pasture was cut daily at around 1000 h, transported to the AgResearch Grasslands Ulyatt Reid Animal Facility 1 h after harvesting. The pasture was divided into equal individual feed portions for afternoon feeding and morning feeding the next day. The pasture portions were stored in a cold room at 4 °C and offered to the animals at 1600 h the same day and 0800 h the next day. Fresh water was available throughout the trial.
Feed allocation was calculated based on multiples (m) of maintenance energy requirements (MEr) of the animals (Australian Agricultural Council Citation1990) calculated according to the following equation:(1) where W = liveweight (kg), A = age (y) and M/D = energy density (MJ/kg) of the diet. From each cut of pasture a representative sample was collected and dry matter (DM) content was determined by drying the sample at 105 °C for 24 h in a ventilated oven. A separate sub-sample was dried at 65 °C for 48 h and then ground to pass through a 1 mm sieve using a Wiley mill (MF 16, IKA Werke, Germany). Chemical composition was analysed using near infrared spectroscopy (NIR) by FeedTech (Palmerston North, New Zealand). NIR analysis provided values for organic matter digestibility (OMD), metabolisable energy (ME), soluble sugars and starch (SSS), crude protein (CP), lipids (L), neutral detergent fibre (NDF) and ash.
At the beginning of each trial the animals were weighed and randomly allocated to their respective treatments stratified for body weight, except for Trial 2 where dry and pregnant/lactating sheep were compared. In each trial, the sheep were acclimatised to indoor housing in group pens (n = 6–8 per pen) and fed with fresh-cut pasture for 10–14 days at 0800 h and 1600 h. Eating behaviour was observed to identify any animals not eating; animals not eating were removed from the group and replaced with spare animals which were always carried in each trial. The sheep were then acclimatised to metabolic crates for 3 days before being transferred into the respiration chambers (Pinares-Patiño et al. Citation2008b). Methane emissions were measured for 2 consecutive days in the respiration chambers. Feed refusals were collected prior to each feeding when the animals were either in metabolic crates or respiration chambers. The refusals or representative sub-samples, where these were large, were dried at 65 °C for 48 h. Dry matter intake and the achieved level of feeding above maintenance (mMEr) was calculated from the difference between the feed offered and refused, corrected for DM. At the end of the 2-day methane emission measurement periods, the animals were weighed and transferred back on to pasture at AgResearch Grasslands.
Trial 1: Effect of level of feeding on methane emissions in sheep <1 year of age
The aim of this experiment was to determine the effect of intake (DMI or mMEr) on methane production (pCH4) and methane yield (Ym) of sheep aged <1 year fed fresh-cut pasture.
The experiment was carried out in June 2008. Twelve Romney wether- and 12 Romney ewe-lambs, 8–9 months of age, were randomly allocated to four groups of six, with equal numbers of female and male lambs. The groups were offered fresh-cut pasture at 0.8, 1.2, 1.6 or 2.0 MEr and methane emissions were measured in respiration chambers from six animals per group.
Trial 2: Effect of level of feeding on methane emissions from ewes >1 year old
The aim of this experiment was to determine the effect of intake (mMEr and DMI) on pCH4 and Ym on animals >1 year old. To create a wider range of DMI and mMEr than in Trial 1, emissions from non-pregnant, pregnant and lactating ewes were measured throughout a production cycle. A control group of dry ewes was measured in parallel to aid in the interpretation of any differences in methane emissions arising from time-dependent changes in dietary composition of the pasture occurring during the 7-month experiment (May–December 2008).
Twelve ewes were selected from a flock of 60 synchronised pregnant Romney ewes at AgResearch Ballantrae farm and transported to AgResearch Grasslands. A second group of 12 dry ewes were also transported from Ballantrae farm for methane measurements. The quantity of feed offered to the pregnant/lactating ewes group varied with stage of pregnancy/lactation and ranged from 1.2–3.6 MEr, while the quantity offered to the dry ewes remained constant throughout at 1.2 MEr ().
Table 1 Feeding levels of the intake group (dry, pregnant and lactating ewes) and control (dry) ewes during Trial 2.
The pregnant ewes lambed on 8 August (±3 days) and the lambs were weaned on 27 October. Methane measurements of pregnant and control dry ewes were carried out on 23 May, 21 June and 14 July (Periods P1 to P3, respectively). Post-lambing methane measurements on both groups were undertaken on 31 August and 15 October (P4 and P5) and a final measurement taken on both groups after weaning on 4 December (P6). A second group of 12 dry ewes were also transported from Ballantrae farm for methane measurements. For the lactating ewes, when indoors either in group pens, metabolic crates or respiration chambers, the lambs were allowed to suckle from their mothers for 30 min at 0800 h and 1600 h when refusals were collected and before new pasture was offered to the ewes.
Trial 3: Effect of level of feeding and diet quality on methane emissions from ewes >1 year old
The aim of this experiment was to determine the effect of pasture quality on pCH4 and Ym at a range of feeding levels in sheep >1 year old.
A group of 24 adult male sheep were selected from the Grasslands Animal Facility, Palmerston North, New Zealand and randomly allocated to one of four feeding levels (0.8, 1.2, 1.6 and 2.0 MEr). Animals were offered fresh-cut pasture harvested at Aorangi Farm. The experiment consisted of three methane measurement periods in November 2009, January 2010 and April 2010 representing spring, summer and autumn, respectively. The pastures from which feed was taken were not managed specifically to manipulate diet quality but changes in diet quality were anticipated due to the natural seasonal variation.
Trial 4: Effect of feeding level and diet quality on methane emissions from lambs aged 15–40 weeks
The aim of this experiment was to determine the effect of both feeding level and diet quality on pCH4 and Ym in lambs between 15 and 36 weeks of age. This provided a direct comparison with the results obtained from Trials 2 and 3 using sheep >1 year old.
A group of 48 lambs (24 male, 24 female) were selected from the flock at AgResearch Grasslands, Palmerston North, at 13 weeks of age (10 days post-weaning) and allocated to one of four treatments. The treatments were two different pasture qualities cut from differently managed paddocks (irrigated/fertilised versus unmanaged) offered at two levels (1.4 MEr and ad libitum) of feeding. Measurements of methane emissions were carried out at 16, 20, 25, 30, 35 and 40 weeks of age. To minimise any post-weaning stress, only pasture from the irrigated/fertilised paddocks (managed) was offered to the lambs in the first measurement period.
Meta-analysis
A total of 510, 2-day measurements of methane emissions in respiration chambers from 115 animals fed fresh-cut pasture were carried out across the four experiments (). The dataset covered a range of DMI from 0.3–1.7 kg/d corresponding to 0.5–3.3 MEr. It also covered a range of pasture quality with OMD for example varying from 551–870 g/kg DM. In total, 312 measurements from sheep <1 year and 198 measurements from sheep >1 year were carried out. The age structure of the older sheep, however, was limited, with the maximum age being 2.3 years. This limited age range precluded using age as an independent variable in any analysis of the complete dataset. In order to remove trends in the residuals when comparing the four trials together, DMI, pCH4 and OMD had to be log-transformed.
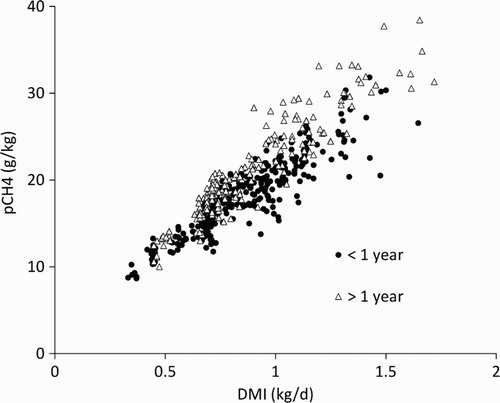
Figure 1 Methane production (pCH4) and dry matter intake (DMI) measured for animals <1 year (n = 311) and >1 year (n = 199).
Statistical analysis
Data from individual trials were analysed first using by simple linear regression with methane as the independent variable (Ym and pCH4) and DMI or achieved feeding level (mMEr) being the explanatory variable. Multiple regression analysis was then carried out to explore the effects of the various diet quality parameters in addition to DMI or feeding level. In Trial 2, an analysis of parallelism using linear regression with groups was performed to assess the effects of physiological status on emissions. All the above analyses were carried out using GenStat® v. 13 (VSNi, Hemel Hempstead, UK; Payne et al. Citation2010). A t-test was performed to compare intake and methane emissions at different feeding levels in Trials 1, 3 and 4 and physiological status in Trial 2 using GenStat.
A meta-analysis was conducted on the whole dataset and for this the response variables (Ym and pCH4) and the dominant explanatory covariate (DMI) needed to be transformed using natural logarithms in order to de-trend the residuals, linearise and stabilise variances in the data.
An analysis of the whole dataset using age as an explanatory variable alongside other variables such as DMI was not undertaken due to the very limited range of ages used in the trials, for example, the age range for mature sheep was only 0.3–2.3 years. To examine the effects of age a further analysis was conducted on the data from young and old animals (i.e. with 1 year of age being the separating value). For each combined dataset (Trials 1 and 4 for sheep <1 year; Trials 2 and 3 for sheep >1 year), an analysis was performed using a linear mixed effects model and REML method using GenStat v. 13 (Payne et al. Citation2010). The optimum set of variables to include in relationships derived from these analyses was assessed using a combination of criteria such as the deviance and the Akaike Information Criterion 1.
Results
Trial 1: Effect of level of feeding on methane emissions in sheep <1 year old
The aim of this experiment was to determine the effect of DMI and mMEr on pCH4 and Ym of growing sheep fed fresh-cut pasture. The trial was carried out in June 2008 with a group of 24 mixed sex lambs (8–9 months old) which were offered four levels of fresh-cut pasture. The average live weight of the lambs at the start of the experiment was 38 ± 1.5 kg.
Ash, CP, lipid, sugar and NDF content of the pasture were 34, 157, 31, 161 and 445 g/kg DM, respectively, the OMD was 806 g/kg DM and the ME content 11.9 MJ/kg DM. Refusals in this experiment were minimal as the animals usually achieved the intended feeding level (). As feeding level increased from 0.76 (0.35 kg DMI day−1) maintenance to 1.82 (0.87 kg DMI day−1) maintenance, pCH4 increased from 9.3 to 18.3 g/d (). However, since feeding level (mMEr) and DMI increased at a faster rate than pCH4, Ym decreased as MEr and DMI increased ().
Table 2 Achieved feeding level relative to maintenance energy requirements, dry matter intake, methane production and yield from lambs offered four levels of fresh-cut pasture.
Regression analysis was used to obtain relationships between Ym and DMI, Ym and mMEr, pCH4 and DMI, and pCH4 and mMEr. The strongest relationship found was between DMI (kg/d) and pCH4 (Equation [2]) where DMI alone explained 91% of the variation in pCH4. The relationship between pCH4 and mMEr was also very strong and explained 88% of the variation in pCH4 (Equation [3]). The relationship between DMI and Ym was negative and explained 60% of the variance in Ym (Equation [4]). Replacing DMI with mMEr resulted in a relationship (Equation [5]) that explained 60% of the variation in Ym.(2)
(3)
(4)
(5)
Trial 2: Effect of level of feeding on ewes >1 year old
The primary aim of this experiment was to examine the relationship between methane emissions (pCH4 and Ym) and level of feeding. To obtain the full range of feeding levels found in practice, dry, pregnant and lactating ewes were used in the trial, allowing the effect of physiological state to be included in the analysis. Finally, because the experiment ran over a whole reproductive cycle, the impact of the seasonal variation in pasture quality could also be examined.
The chemical composition of the pasture offered during the trial differed markedly with the OMD ranging from 693–811 g/kg and the ME from 10.4–12.0 MJ/kg (). Sugar content was relatively high in Periods 4 and 5 when animals were lactating and NDF contents were lower than in other periods. The protein content of the pasture was satisfactory for the first four periods but dropped to a concentration below that generally recommended for ruminants in the last two periods (McNeill et al. Citation1997).
Table 3 Chemical composition, organic matter digestibility and metabolisable energy content of the pasture fed to ewes in Trial 2 while pregnant (P1–3), lactating (P4 and 5) and after weaning (P6).
During the first and the last period, when all animals were offered the same amount of feed, no differences in DMI, Ym and pCH4 were observed between the pregnant/lactating and control ewes (). As mMEr and DMI increased to meet the increasing demand of the pregnant/lactating ewes, pCH4 increased (). In common with Trial 1, regression analysis revealed that DMI alone was a very good predictor of pCH4, accounting for 86% of the variation (Equation [6]). The relationship between pCH4 and mMEr was also strong (r2
= 82%), while those between Ym and DMI and between Ym and mMEr were weaker, accounting for only 33% and 29% of the variance, respectively (data not shown).(6)
Table 4 Dry matter intake and methane production from pregnant, lactating and dry ewes fed fresh-cut pasture in Trial 2 while pregnant (P1–3), lactating (P4 and 5) and after weaning (P6).
At the same level of DMI, lactating ewes in Period 5 produced 15% more methane than when they were pregnant in Period 3 (). Consequently, Ym was also increased in Period 5 compared with values found in Period 3. Although DMI was similar in these two periods, mMEr increased in Period 5 compared with Period 3 because of changes in both animal age and weight of the ewes between the two periods. Differences in diet quality, however, did not affect the control group, which had relatively constant Ym across the trial at equal levels of intake irrespective of changes in diet quality. Adding OMD, ME and other dietary components in addition to DMI and mMEr into the analysis of the whole dataset from Trial 2, either singly or in combination, made little difference to the amount of variation explained, increasing it at best from 86% to 87% for DMI and from 82% to 84% for mMEr (data not shown).
Trial 3: Effect of level of feeding and diet quality on methane emissions from ewes >1 year old
The aim of this trial was to determine the effect of diet quality in addition to level of feeding on Ym and pCH4 in adult sheep. The trial was carried out in three periods (spring, summer and autumn). The chemical composition of the pasture changed with the season (). The pasture in spring was characterised by low NDF, high protein and high sugar content, whereas the composition of the summer pasture was characterised by a high NDF content accompanied by low sugar and protein contents. Values in autumn were intermediate between those recorded in the summer and spring. In all periods, the animals were offered feed at up to twice their maintenance energy requirements, but only in the spring period was this level of intake achieved (). In summer and autumn, when the diet was of much poorer quality, intakes of only 1 and 1.4 mMEr, respectively, were achieved.
Table 5 Pasture chemical composition, organic matter digestibility and metabolisable energy content of diets fed to the adult sheep.
Table 6 Dry matter intake, achieved feeding level and methane production and methane yield of sheep fed fresh-cut pasture of different quality.
In common with Trials 1 and 2, pCH4 was strongly related to DMI, explaining 91% of the variation in pCH4 (Equation [7]). In contrast to Trials 1 and 2, the relationship between pCH4 and mMEr was much weaker (r2
= 0.59) than the relationship between pCH4 and DMI (data not presented). Only weak relationships between Ym and both DMI and mMEr were found. Only the relationship between Ym and DMI was significant and DMI explained only 9.3% of the variation in Ym (data not shown).(7)
Including feed quality variables in the relationship between pCH4 and DMI improved the prediction equation; the biggest improvement came from the inclusion of either lipids and OMD or NDF and OMD. The improvement, however, was comparatively small with the r2 increasing from 0.91 to 0.94 in both cases (data not presented). Including quality parameters in the relationship between pCH4 and mMEr did, however, significantly improve the relationship. For example, adding OMD changed the proportion of variance explained from 58% to 87%, a value still below the proportion of variance explained using DMI alone (data not shown). In contrast, adding a quality parameter into the relationship between Ym and DMI/mMEr had little effect (data not shown).
Trial 4: Effect of feeding level and diet quality on methane emissions from lambs aged 15–40 weeks
The hypothesis for this trial was that methane emissions (pCH4 and Ym) in sheep <1 year old have a different relationship between intake and herbage quality compared with older sheep, but that differences would disappear over time as the young animals approach 1 year of age.
In order to achieve a good range of DMI and feed quality, the lambs were offered the fresh pasture at two feeding levels, either 1.4 or 2.5 mMEr which was effectively ad libitum for the higher feeding level (). With increasing age, the pCH4 increased but no trend in Ym was observed (). Over the course of the experiment, DMI varied from 0.4–1.6 kg/d. Pasture quality was directly manipulated via management in addition to the expected seasonal changes.
Table 7 Dry matter intake, achieved feeding level and methane production and methane yield of lambs fed fresh-cut pasture of different quality.
As the trial took place over a 6-month period, pasture quality differed both within periods, due to deliberate management effects, and between periods due to seasonal effects. OMD varied from 666–793 g/kg in the unmanaged pastures and from 654–764 g/kg in the managed pastures (). With sugar, component absolute values were low in general, ranging from 54–132 g/kg. Pasture management did not change sugar concentration of the pasture ().
Table 8 Diet composition of the pasture fed to lambs in six periods after weaning from differentially managed paddocks to yield good and average quality within each period.
As with the previous three trials, a strong relationship between DMI and pCH4 was found (Equation [8]) although less of the variance (80%) was explained than in the other trials. A feature of the relationship was also that the slope parameter was less than in other DMI:pCH4 relationships. The relationship between pCH4 and mMEr explained less of the variance (60%) than the pCH4 and DMI relationship (data not shown). Relationships between Ym and DMI and between Ym and mMEr were also much weaker, explaining 36% and 21% of the variance, respectively (data not shown).(8)
Including feed characteristics of the diet in the models improved model predictions significantly. For example, adding OMD alone into the relationship between pCH4 and DMI improved the percentage of variance explained from 81% to 84% (Equation [9]). More complex models that included multiple dietary components did not significantly improve on this relationship (data not shown). Adding quality parameters into the relationships between pCH4 and mMEr, Ym and DMI, and Ym and mMEr also improved the predictions significantly (maximum r2
values of 0.72, 0.54 and 0.59, respectively; data not shown).
Meta-analysis
As observed in the analysis of the individual trials, the REML analysis showed that the simple relationship between DMI and pCH4 (Equation [10]) was superior to the relationship between mMEr and pCH4 and the negative relationships between Ym and either DMI or mMEr. The relationship between pCH4 and mMEr (Equation [11]), although strong, is less accurate (lower deviance and Akaike coefficient) than the pCH4 and DMI relationship:(10)
Similar to the results found in the individual trials for Ym, the regression using DMI (Equation [12]) is the better model compared with the regression of Ym and mMEr (Equation [13]):(12)
(13) Including dietary components in addition to DMI in the equations improved the predictions slightly and OMD was a significant component in each model. However, the standard error of the difference (SED) values increased compared with the relationship between pCH4 or Ym with DMI alone. For relationships of pCH4 and Ym with mMEr, the SED did not improve and these equations describe the dataset poorly compared with DMI relationships alone. In Equation (14), the best of the equations for the relationship of pCH4 and DMI is shown. Both DMI and OMD have a positive influence on pCH4.
(14) Including age in the analysis had a significant effect on the relationship of pCH4 and DMI. However, because of the limited age structure of the experimental animals, the datasets were split into two sets with one dataset representing sheep <1 year and the other sheep >1 year, similar to the distinction made in the current New Zealand inventory ().
For young sheep (<1 year) in Trials 1 and 4, the relationship between DMI and pCH4 is described in Equation (15) and the equation for old animals (>1 year) is described in Equation (16):(15)
(16)
The slope of the old animals in this relationship is much steeper than for the young animals, while the intercept is not significantly different. Including dietary composition in the analysis had a significant effect on methane emissions from young sheep but not older sheep. Of the dietary parameters tested, the inclusion of ME content resulted in the best fit for the young animals (Equation [17]) followed by log-transformed OMD values (Equation [18]). None of the other dietary components or the combination of them improved the relationship over the one shown in Equation (17) or Equation (18):(17)
(18)
Discussion
Our results confirm that the principal driver of methane emissions in ruminants is feed intake. Dry matter intake alone explained 80%–91% of the variation in the methane production across the four studies presented here. Expressing intake as multiples of maintenance energy explained less of the variance but using mMEr is an accurate way to predict methane emissions in sheep consuming the type of forage diets commonly found on New Zealand sheep farms. The relationships between intake and pCH4 are characterised by a significant intercept, meaning that Ym decreases as intake increases. Such a negative relationship between DMI and Ym, which is probably associated with an increased rate of passage and/or reduced digestibility, has also been described earlier in sheep fed fresh-cut pasture or white clover (Molano & Clark Citation2008; Hammond et al. Citation2013). The confirmation here of a negative relationship between pCH4 and intake indicates that a single yield factor describing methane emissions in sheep fed fresh-cut pasture does not adequately reflect the underlying biology. This behaviour of Ym was also described for sheep fed roughage and concentrate diets (Blaxter & Clapperton Citation1965) indicating that this relationship is diet independent.
Relationships between intake and Ym were much weaker than those found using pCH4 and intake across all four experiments. This indicates that when predicting pCH4 for sheep using the type of diets predominating in New Zealand, adopting the IPCC convention of a methane yield (Ym) factor multiplied by an intake factor is less satisfactory than directly predicting pCH4 from feed intake.
A primary objective of this study was to examine whether animals <1 year old produce less methane at equal levels of intake than animals >1 year old. Evidence for there being a different relationship comes from the work of Lassey et al. (Citation1997) and Ulyatt et al. (Citation2002) and, based largely on their work, the New Zealand inventory adopted Ym values of 21.6 and 16.8 g CH4/kg DMI for sheep <1 year and >1 year, respectively. A separate evaluation of trials conducted in New Zealand between 2002 and 2005 (Kelliher et al. Citation2009) also found that the Ym values for sheep >1 and sheep <1 year were different, although their values of 21.6 and 19.6 g CH4/kg DMI, respectively, are much closer than those adopted in the national methane inventory. A possible explanation for a lower methane emission from younger animals is that, at the same level of intake, young animals have a smaller body size and therefore smaller rumens, meaning a faster rate of passage of ingested feed and a faster fermentation rate. It would follow from this reasoning that as young sheep became closer in body size to adult ewes, the effect would reduce and eventually disappear. The results from the four individual trials reported here do not support the simple hypothesis that emissions from younger animals would be generally lower than those from older animals but that these differences would disappear over time as the young animals approached 1 year of age. In fact the relationship between DMI and pCH4 in animals <1 year had similar intercepts but flatter slopes than the equivalent relationship found for animals >1 year, meaning that at lower levels of intake, which will often be associated with younger animals, differences between younger and older animals will be minimal. Our results are in contrast to those of Knight et al. (Citation2008) who showed that early in a lamb's life, Ym values are low but increase until they reach 6 months of age. However, the authors state that seasonal variation in pasture quality confounded their results.
Dietary composition is often a confounding effect with intake when working with fresh pastures because it is impossible to maintain a constant herbage quality over time. However, most methane prediction algorithms have a dietary composition component (Blaxter & Clapperton Citation1965; Moe & Tyrrell Citation1979; Mills et al. Citation2003). In general, it is recognised that increased non-fibre sugar content in ruminant feeds leads to a more propionate-based fermentation pattern which in turn decreases the amount of hydrogen produced (van Nevel et al. Citation1974; Janssen Citation2010; Archimède et al. Citation2014) and consequently leads to lower methane emissions. However, in the four trials reported here, including diet chemical composition to the regression analyses had little impact on the overall variation explained by the model. Digestibility is a common measure of feed quality and incorporating feed quality had the greatest impact in Trial 4, when the inclusion of OMD increased the variance explained from 81% to 84%. A similar pattern was observed using individual components such as sugar or NDF. For example, OMD ranged from 551–870, soluble sugars from 48–202 and NDF from 420–652 g/kg, values that cover the range found in practice on sheep farms in New Zealand.
The lack of any large effect of chemical composition on measured CH4 emissions is a common finding. In a review, Pelchen & Peters (Citation1998) found no clear effect of energy density or fibre content on methane emissions in sheep. Even when roughages with completely different dietary composition such as white clover and forage chicory—where plant cell wall concentrations are very low and soluble sugars reach up to 170 g/kg DM—were fed to sheep, no difference in methane emissions was observed (Hammond et al. Citation2011; Sun et al. Citation2012). These recent studies confirm some earlier New Zealand work where differences in pasture composition could not be linked to methane emissions in sheep (Ulyatt et al. Citation2005; Molano & Clark Citation2008; Hammond et al. Citation2009; Hart et al. Citation2009). Higher digestibility in forages would increase methane emissions per kg DMI, but at the same time higher digestibility is usually associated with less fibre and more sugar and protein content. These components lead to more propionogenic fermentation pathways which decrease methane emissions. High amounts of sugars are needed to reduce pCH4 in ruminants and the effect may not be driven only by the changed sugar fermentation pathways but by a decrease in pH. Methanogens are sensitive to low pH (Hook et al. Citation2011) but in order to decrease pH below 5.5, large amounts of soluble sugars are needed (Lettat et al. Citation2010). It is possible that the concentrations of sugar needed to lower the pH to a level where it reduces pCH4 are not generally present in forages.
Although our results suggest that for sheep consuming fresh forages diet quality does not strongly influence CH4 emissions, Trial 4 does indicate that young sheep are more sensitive to diet quality than older sheep. For example, the inclusion of OMD into the regression model in addition to DMI significantly improved the relationship and, since the OMD term is positive, this indicates that for any given level of DMI in sheep <1 year, emissions increase as OMD increases. This effect is a feature of the Blaxter & Clapperton (Citation1965) prediction equation and is likely to be caused by the fact that high-quality material provides more fermentable substrate for rumen micro-organisms. Why this mechanism appears to have less influence in older sheep is unclear but might be due to a more efficient fermentation of plant fibre.
Meta-analysis: recommended inventory equations
The REML analysis conducted on the entire dataset confirmed that DMI alone was the best single predictor of pCH4 as described in Equation (10):
Adding herbage quality to the whole dataset had no significant effect and resulted in an increased standard error of prediction and is therefore less suitable for use in an inventory than a relationship incorporating DMI alone. Due to limitations in the range of ages studied, it is not possible to capture the effect of age, and any interactions with diet quality, in a single equation that can be applied to all sheep.
The separate meta-analyses of the trials involving sheep <1 year and those >1 year do however suggest that emissions from those <1 year are different from those from animals >1 year at the same level of intake. Intake is the dominant explanatory variable in both cases but the slopes of the relationships are significantly different for the two age categories. Because other New Zealand studies also suggest that emissions differ with age (Clark et al. Citation2003), it would appear appropriate to assume that age does have an influence on emissions.
In older sheep, herbage composition had no influence on pCH4 for sheep >1 year fed fresh pasture. The most appropriate equation to describe methane production in older sheep is Equation (16) which includes only DMI:
Methane emissions from sheep <1 year are influenced by herbage quality with the ME content of the pasture being the best parameter to include alongside DMI. The relationship that best predicts methane emissions from young sheep is Equation (17):
On average, the standard errors of the predicted methane emission using the separate Equations (16) and (17) are lower than those obtained using a single simple Equation (10) (). The predictive accuracy of each model was compared using cross-validation. The sum of squared differences between model prediction and actual measurement was calculated for each model under each cross-validation. For the simpler model the mean predictive error was 0.0441, while for the model with separate parts for old and young animals the mean predictive error was 0.0271 representing a 60% greater accuracy when the two separate Equations (16) and (17) were used.
Table 9 Predictions and prediction errors of the three equations representing the simple model (Equation [10]) and the young-old model (Equations [16] and [17]).
Impact of new equations on estimated emissions from ewes and lambs in New Zealand 1990 and 2012
To quantify the impact that these new methane prediction equations have on estimated emissions from New Zealand sheep, emissions from adult ewes and growing lambs were estimated for the years 1990 and 2012 using the national agricultural inventory accounting software. Equations (16) and (17) were used separately to estimate emissions for ewes and lambs and compared with: (a) estimates obtained using Equation (10) for both ewes and lambs; and (b) estimates obtained using the current fixed Ym values of 20.9 g/kg DMI for ewes and 16.8 g/kg DMI for lambs ().
Table 10 Changes in New Zealand methane emissions (tonnes CO2 equivalent) for ewes and lambs from 1990 to 2012 using the simple Equation (10) or the age split Equations (16) and (17) compared with the current inventory.
The use of a single equation, as opposed to the two separate equations, made little difference to combined ewe and lamb total emissions in 1990. However, in 2012, using separate equations for ewe and lamb emissions resulted in a smaller increase of methane emissions over the current method than using a single combined equation (). The impact of using a combined ewe and lamb equation (Equation [10]) to predict ewe emissions, rather than a separate ewe equation, differed between 1990 and 2012. In 1990, using a single equation resulted in higher emissions than the two-equation approach, but the situation was reversed in 2012; in percentage terms, however, these differences were small (1.6% and −0.8%, respectively). In contrast, using a specific lamb prediction equation rather than a combined ewe and lamb equation resulted in lower estimated emissions from lambs in both 1990 and 2012 (−10.9% and −18.5%, respectively).
Using the new algorithms (Equation [10] alone or Equations [16] and [17] combined) obtained from the experiments described in this paper resulted in higher estimated combined ewe and lamb emissions (both per head and in total) in 1990 and 2012 compared with the current estimation method (). The increase was greater in 1990 than in 2012, meaning that although the new algorithms predict higher emissions than the current method, the estimated decrease in combined ewe and lamb emissions between 1990 and 2012 is also greater: current, 166,525 tonnes; Equation (10), 191,854 tonnes; Equations (16) and (17), 179,081 tonnes.
The calculations undertaken using the national inventory software clearly demonstrate that using the new algorithms obtained from the experiments described here make a difference to New Zealand's estimated annual methane emissions. Using a single equation for all sheep irrespective of age increases estimated lamb emissions by 31%–33% over the current method while using a specific equation for sheep <1 year old increases them by 20% in 1990 and 11% in 2012; this simply reflects the fact that the results from these series of experiments do not support previous studies showing an approximate 20% decrease in emissions from sheep <1 year old compared with those >1 year old. The impact of the new algorithms, however, is not restricted to reduced effect of age on emissions since the new algorithms obtained from the experiments described here predict greater emissions from older sheep (). They also capture the well-documented phenomenon of a decrease in emissions per unit of intake as the level of intake increases (eg, Blaxter & Clapperton Citation1965); this is demonstrated by the larger predicted decrease in emissions between 1990 and 2012 in response to improved animal performance and higher levels of intake found in New Zealand sheep over this period (MfE Citation2014).
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
The authors gratefully acknowledge funding from the Ministry for Primary Industries, New Zealand.
References
- Archimède H, Martin C, Periacarpin F, Rochette Y, Etienne TS, Anais C, Doreau M 2014. Methane emission of Blackbelly rams consuming whole sugarcane forage compared with Dichanthium sp. hay. Animal Feed Science and Technology 190: 30–37. doi: 10.1016/j.anifeedsci.2014.01.004
- Australian Agricultural Council. 1990. Feeding standards for Australian livestock—ruminants. Sydney, Australia, CSIRO Publications.
- Blaxter KL, Clapperton JL 1965. Prediction of the amount of methane produced by ruminants. British Journal of Nutrition 19: 511–522. doi: 10.1079/BJN19650046
- Clark H, Brookes I, Walcroft A 2003. Enteric methane emissions from New Zealand ruminants 1990–2001 calculated using an IPCC Tier 2 Approach. Report prepared for the Ministry of Agriculture and Forestry (March 2003). Wellington, Ministry of Agriculture and Forestry.
- Dijkstra J, Neal HDSC, Beever DE, France J 1992. Simulation of nutrient digestion, absorption and outflow in the rumen: Model description. The Journal of Nutrition 122: 2239–2256.
- Ellis JL, Kebreab E, Odongo NE, Beauchemin K, McGinn S, Nkrumah JD et al. 2009. Modeling methane production from beef cattle using linear and nonlinear approaches. Journal of Animal Science 87: 1334–1345. doi: 10.2527/jas.2007-0725
- Grainger C, Clarke T, McGinn SM, Auldist MJ, Beauchemin KA, Hannah MC et al. 2007. Methane emissions from dairy cows measured using the sulfur hexafluoride (SF6) tracer and chamber techniques. Journal of Dairy Science 90: 2755–2766. doi: 10.3168/jds.2006-697
- Hammond KJ, Muetzel S, Waghorn GC, Pinares-Patino CS, Burke JL, Hoskin SO 2009. The variation in methane emissions from sheep and cattle is not explained by the chemical composition of ryegrass. Proceedings of the New Zealand Society of Animal Production 69: 174–178.
- Hammond KJ, Hoskin SO, Burke JL, Waghorn GC, Koolaard JP, Muetzel S 2011. Effects of feeding fresh white clover (Trifolium repens) or perennial ryegrass (Lolium perenne) on enteric methane emissions from sheep. Animal Feed Science and Technology 166–167: 398–404. doi: 10.1016/j.anifeedsci.2011.04.028
- Hammond KJ, Burke JL, Koolaard JP, Muetzel S, Pinares-Patiño CS, Waghorn GC 2013. Effects of feed intake on enteric methane emissions from sheep-fed fresh white clover (Trifolium repens) and perennial ryegrass (Lolium perenne) forages. Animal Feed Science and Technology 179: 121–132. doi: 10.1016/j.anifeedsci.2012.11.004
- Hart KJ, Martin PG, Foley PA, Kenny DA, Boland TM 2009. Effect of sward dry matter digestibility on methane production, ruminal fermentation, and microbial populations of zero-grazed beef cattle. Journal of Animal Science 87: 3342–3350. doi: 10.2527/jas.2009-1786
- Hook SE, Steele MA, Northwood KS, Wright ADG, McBride BW 2011. Impact of high-concentrate feeding and low ruminal pH on methanogens and protozoa in the rumen of dairy cows. Microbial Ecology 62: 94–105. doi: 10.1007/s00248-011-9881-0
- IPCC. 1996. Houghton JT, Meira Filho LG, Lim B, Treanton K, Mamaty I, Bonduki Y, Griggs DJ, Callender BA eds. IPCC/OECD/IEA. Revised 1996 IPCC Guidelines for National Greenhouse Gas Inventories. Bracknell, United Kingdom Meteorological Office.
- IPCC. 2000. Penman J, Kruger D, Galbally I, Hiraishi T, Nyenzi B, Emmanul S et al. eds. Good Practice Guidance and Uncertainty Management in National Greenhouse Gas Inventories. IPCC National Greenhouse Gas Inventories Programme. Japan, Institute for Global Environmental Strategies for IPCC.
- IPCC. 2006. Eggleston HS, Buendia L, Miwa K, Ngara T, Tanabe K et al. eds. 2006 IPCC Guidelines for National Greenhouse Gas Inventories. Volume 4. Agriculture, Forestry and Other Land Use. IPCC National Greenhouse Gas Inventories Programme. Published for the IPCC by the Institute for Global Environmental Strategies: Japan.
- Janssen PH 2010. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Animal Feed Science and Technology 160: 1–22. doi: 10.1016/j.anifeedsci.2010.07.002
- Knight TW, Molano G, Clark H, Cavanagh A 2008. Methane emissions from weaned lambs measured at 13, 17, 25 and 35 weeks of age compared with mature ewes consuming a fresh forage diet. Australian Journal of Experimental Agriculture 48: 240–243. doi: 10.1071/EA07258
- Kelliher FM, Clark H, Smith MH, Lassey KR, Sedcole R 2009. Reducing uncertainty of the enteric methane emissions inventory. Report for the Ministry of Agriculture and Forestry. Wellington, Ministry of Agriculture and Forestry.
- Lassey KR, Ulyatt MJ, Martin RJ, Walker CF, Shelton DI 1997. Methane emissions measured directly from grazing livestock in New Zealand. Atmospheric Environment 31: 2905–2914. doi: 10.1016/S1352-2310(97)00123-4
- Lettat A, Nozière P, Silberberg M, Morgavi DP, Berger C, Martin C 2010. Experimental feed induction of ruminal lactic, propionic, or butyric acidosis in sheep. Journal of Animal Science 88: 3041–3046. doi: 10.2527/jas.2010-2926
- McNeill DM, Slepetis R, Ehrhardt RA, Smith DM, Bell AW 1997. Protein requirements of sheep in late pregnancy: partitioning of nitrogen between gravid uterus and maternal tissues. Journal of Animal Science 75: 809–816.
- MfE 2014. New Zealand's Greenhouse Gas Inventory 1990–2012. Wellington, Ministry for the Environ-ment. http://www.mfe.govt.nz/publications/climate/greenhouse-gas-inventory-2014/index.html (accessed 20 October 2015).
- Mills JAN, Kebreab E, Yates CM, Crompton LA, Cammell SB, Dhanoa MS et al. 2003. Alternative approaches to predicting methane emissions from dairy cows. Journal of Animal Science 81: 3141–3150.
- Moe PW, Tyrrell HF 1979. Methane production in dairy cows. Journal of Dairy Science 62: 1583–1586. doi: 10.3168/jds.S0022-0302(79)83465-7
- Molano G, Clark H 2008. The effect of level of intake and forage quality on methane production by sheep. Australian Journal of Experimental Agriculture 48: 219–222. doi: 10.1071/EA07253
- van Nevel CJ, Prins RA, Demeyer DI 1974. On the inverse relationship between methane and propionate in the rumen. Zeitschrift fur Tierphysiologie, Tierernahrung und Futtermittelkunde 33: 121–125.
- Payne RW, Murray DA, Harding SA, Baird DB, Soutar DM 2010. GenStat for Windows, 13th edition. Introduction. Hemel Hempstead, VSN International.
- Pelchen A, Peters KJ 1998. Methane emissions from sheep. Small Ruminant Research 27: 137–150. doi: 10.1016/S0921-4488(97)00031-X
- Pinares-Patiño CS, Clark H 2008. Reliability of the sulfur hexafluoride tracer technique for methane emission measurement from individual animals: an overview. Australian Journal of Experimental Agriculture 48: 223–229. doi: 10.1071/EA07297
- Pinares-Patiño CS, Holmes CW, Lassey KR, Ulyatt MJ 2008a. Measurement of methane emission from sheep by the sulphur hexafluoride tracer technique and by the calorimetric chamber: failure and success. Animal 2: 141–148. doi: 10.1017/S1751731107000857
- Pinares-Patiño CS, Lovejoy P, Hunt CS, Martin R, Molano G, Waghorn GC, Clark H 2008b. Brief communication: A versatile sheep respiration chamber system for measurement of methane emissions. Proceedings of the New Zealand Society of Animal Production 68: 29–30.
- Pinares-Patiño CS, Lassey KR, Martin RJ, Molano G, Fernandez M, MacLean S et al. 2011. Assessment of the sulphur hexafluoride (SF6) tracer technique using respiration chambers for estimation of methane emissions from sheep. Animal Feed Science and Technology 166–167: 201–209. doi: 10.1016/j.anifeedsci.2011.04.067
- Reynolds CK, Crompton LA, Mills JAN 2011. Improving the efficiency of energy utilisation in cattle. Animal Production Science 51: 6–12. doi: 10.1071/AN10160
- Shibata M, Terada F 2010. Factors affecting methane production and mitigation in ruminants. Animal Science Journal 81: 2–10. doi: 10.1111/j.1740-0929.2009.00687.x
- Sun XZ, Hoskin SO, Zhang GG, Molano G, Muetzel S, Pinares-Patiño CS et al. 2012. Sheep fed forage chicory (Cichorium intybus) or perennial ryegrass (Lolium perenne) have similar methane emissions. Animal Feed Science and Technology 172: 217–225. doi: 10.1016/j.anifeedsci.2011.11.007
- Ulyatt MJ, Lassey KR, Shelton ID, Walker CF 2002. Seasonal variation in methane emission from dairy cows and breeding ewes grazing ryegrass/white clover pasture in New Zealand. New Zealand Journal of Agricultural Research 45: 217–226. doi: 10.1080/00288233.2002.9513512
- Ulyatt MJ, Lassey KR, Shelton ID, Walker CF 2005. Methane emission from sheep grazing four pastures in late summer in New Zealand. New Zealand Journal of Agricultural Research 48: 385–390. doi: 10.1080/00288233.2005.9513671
- Undi M, Willson C, Ominski KH, Wittenberg KM 2008. Comparison of techniques for estimation of forage dry matter intake by grazing beef cattle. Canadian Journal of Animal Science 88: 693–701. doi: 10.4141/CJAS08041
- Vlaming JB, Brookes IM, Hoskin SO, Pinares-Patiño CS, Clark H 2007. The possible influence of intra-ruminal sulphur hexafluoride release rates on calculated methane emissions from cattle. Canadian Journal of Animal Science 87: 269–275. doi: 10.4141/A06-056