
ABSTRACT
The plant growth regulator trinexapac-ethyl (TE) is widely used to increase seed yield by reducing lodging in many grass species, including perennial ryegrass (Lolium perenne L.) and tall fescue (Festuca arundinacea Schreb.). However, the seed yield responses of annual ryegrass (Lolium multiflorum Lam.) to TE has not been widely investigated. In four field experiments, different rates of TE (between 0 and 800 g ha−1) were applied and the plant response measured. Seed yield was significantly (P < 0.05) increased above the control by all TE treatments with an average response of 51% and 65% for the 200 g and 400 g TE ha−1 treatments, respectively. The seed yield response was associated with reduced lodging and shorter stems contributing to 1.1 more seeds per spikelet attaining saleable weight resulting in 72% more seeds m−2. The plant growth regulator TE can improve seed yield in well-managed stands of annual ryegrass.
Introduction
Annual ryegrass (Lolium multiflorum Lam.) is the second most important grass species grown for seed in New Zealand, second only to perennial ryegrass (Lolium perenne L.). The species consists of two types or subgroups: the Westerwolds type that are true annuals with no vernalisation requirements; and the Italian types that have a range of vernalisation requirements (Cooper Citation1957; Warnke et al. Citation2004). Italian types will survive more than 1 year if summer moisture is adequate and temperatures are mild. In New Zealand, more seed is produced from the Italian type than the Westerwolds type. In seed crops of many temperate grass species, including perennial ryegrass, Italian ryegrass and tall fescue (Festuca arundinacea Schreb.), stem lodging occurs during reproductive development (Rolston et al. Citation2003; Borm & van den Berg Citation2008; Rijckaert Citation2010). Lodging is the permanent bending of stems from a vertical orientation to a more horizontal orientation. Lodging is a major factor in reducing grass seed yields, especially when it occurs before or during anthesis and early seed development. Seed yield reductions depend on the severity of lodging and have been recorded as high as 95% (Hampton et al. Citation1985; Rolston et al. Citation1997; Griffith Citation2000; Chynoweth et al. Citation2014). These losses are associated with reduced mid-canopy light interception (Rolston et al. Citation2007a) and therefore reduced availability of photosynthate during seed fill. Other adverse effects of lodging include reduced pollination and seed set (Burbidge et al. Citation1978; Wright & Hebblethwaite Citation1979; Warringa & Marinissen Citation1996) and increased vegetative tillering (Hampton & Hebblethwaite Citation1985). Crop inputs such as irrigation and nitrogen (N) fertiliser also influence the incidence and severity of lodging (Rolston et al. Citation1994; Simic et al. Citation2009).
The plant growth regulator trinexapac-ethyl (TE) reduces stem elongation in many grass species including perennial ryegrass and tall fescue (Chastain et al. Citation2003; Rolston et al. Citation2004, Citation2007b, Citation2010a; Borm & van den Berg Citation2008). As an acyl cyclohexanedione inhibitor of gibberellic acid (GA) biosynthesis, TE prevents hydroxylation of the growth inactive form of GA to the active form (Rademacher et al. Citation1992) resulting in stem shortening and, potentially, reduced lodging. Reduced stem length in perennial ryegrass is associated with increased seed yields (Chynoweth et al. Citation2014). Previous Northern Hemisphere studies suggest seed yield responses of Italian ryegrass to TE are generally small, averaging 10% (Mellbye et al. Citation2007; Rijckaert Citation2010; Macháč Citation2012); however, there is no published research on responses in New Zealand seed production environments. The research objective was to evaluate the seed yield responses to different rates of TE in New Zealand annual ryegrass seed production.
Materials and methods
Site management
Four experiments were undertaken within commercial growers’ fields in the Canterbury region of the South Island, New Zealand at locations between 43.7–44.5°S and 171.2–172.4°E at 10–20 m above sea level, with annual ryegrass cultivars of the Italian ryegrass type. The growers applied weed control, fertiliser, fungicides and winter/spring defoliation inputs, with the research team applying the TE to the fields and the experimental plots within it. All sites were irrigated in late spring/early summer with approximately 200 mm of water. The TE (as Moddus® containing 250 g TE L−1) treatments were applied at Zadoks growth stage 32 (Zadoks et al. Citation1974) using a backpack plot sprayer with an electric pressure pump powering a handheld boom fitted with six XR 11002 flat fan nozzles and applying 250 L ha−1. Plots were 9 × 3.2 m, with treatments replicated four times in a randomised complete block design.
Experimental management and application
In Experiment 1 (E1), the diploid cultivar ‘Crusader’ was sown at 12.5 kg ha−1 near Southbridge in early autumn (early March) 2002. Nitrogen application (108 kg N ha−1) was split between autumn (20%) and spring (80%). The field was cut for silage in mid spring (15 October) and closed for seed production. On 26 November, TE was applied at rates ranging from 0–800 g active ingredient (ai) ha−1 in 200 g increments.
In Experiment 2 (E2), the tetraploid cultivar ‘Nourish’ was sown at 12 kg ha−1 near Irwell in early autumn (late March) 2007. Nitrogen (100 kg N ha−1) was applied in a split spring application. The field was grazed by sheep during late winter and early spring, and was closed for seed production in mid spring (25 October). TE was applied at rates ranging from 0–800 g ai ha−1 in 200 g increments on 26 November.
In Experiment 3 (E3), the tetraploid cultivar ‘Feast II’ was sown at 16.5 kg ha−1 near Irwell in mid March 2008. Nitrogen (175 kg N ha−1) was applied in a split spring application. The field was grazed by sheep during late winter and early spring and was closed for seed production on 28 October. TE was applied at rates ranging from 0–800 g ai ha−1 in 200 g increments in late spring (21 November).
In Experiment 4 (E4), ‘Crusader’ was sown at 10 kg ha−1 near Temuka in mid March 2008. Nitrogen (175 kg N ha−1) was applied in a split spring application. The field was grazed by sheep during late winter and early spring. In mid spring (30 October), the trial area was further mown to a height of 70 mm and closed for seed production. TE was applied at rates ranging from 0–800 g ai ha−1 in 200 g increments in late spring (28 November).
Components of seed yield
Approximately 1 week prior to swathing, a 0.25 m2 quadrat was cut at ground level from each plot and weighed. Crop mass was assessed by drying a weighed subsample of approximately 100 g fresh weight at 80 °C for 16 h and calculating dry matter (DM) ha−1. Head density was counted from a weighed subsample and calculated as seed heads m−2. Stem length was measured as the average stem length (mm) from a weighed subsample. Crop mass, head density and stem length were assessed in Experiments E2, E3 and E4. Spikelets were counted on 15 heads per plot, and florets (total, fertile and sterile) were classified and counted on the mid-spikelet from 10 heads. Seed set was calculated as the percentage of fertile florets compared with total florets. Thousand seed weight (TSW) was assessed from the processed and cleaned sample by weighing four replicates each of 50 seeds per treatment. Harvest index was calculated from the ratio of seed weight to total seed and straw weight at harvest. Lodging percentage was visually assessed on a 0% (no lodging) to 100% (fully lodged) linear scale from early anthesis in Experiments E1, E3 and E4.
At harvest, when seed moisture content (SMC) was approximately 42%, a 1.7 m wide swathe the length of the plot was cut with a modified plot windrower, sun dried and threshed approximately 1 week later at 12% SMC with either a ‘Wintersteiger’ Elite or ‘Sampo’ plot combine harvester. The field-dressed seed was cleaned on a small-scale ‘Westrup’ air-screen cleaner to a First Generation Seed Certification standard (98% purity, AsureQuality Citation2008) to calculate machine-dressed seed yield. The number of seeds m−2 was determined from the seed yield and TSW, and seeds per spikelet were calculated by dividing number of seeds m−2 by the number of heads and spikelets per head.
Statistical analysis was conducted with GenStat® (version 15, VSN International) using a general ANOVA model, with treatment effects assessed using least significant difference (LSD) tests (α = 0.05).
Results
Seed yield
Seed yield was increased (P < 0.05) by all rates of TE tested in the four experiments, with seed yield increases ranging from 51%–66% (). The average seed yield increase with 200 g TE ha−1 was 650 kg ha−1 (range 460–960 kg ha−1) across experiments; a 51% increase above the control. At 400 g TE ha−1, the average seed yield increase was 820 kg ha−1 (range 670–1140 kg ha−1); a 65% increase above the control. Further significant increases in seed yield were not achieved at TE rates above 400 g ha−1 with yield increases of 65% and 66% for the 600 g TE and 800 g TE treatments, respectively.
Table 1. Seed yield (kg ha−1) response to five rates of trinexapac-ethyl (TE) on annual ryegrass for Experiments 1 to 4 (E1–4) grown in Canterbury, New Zealand during the 2002–09 growing seasons.
Stem response
Stem length
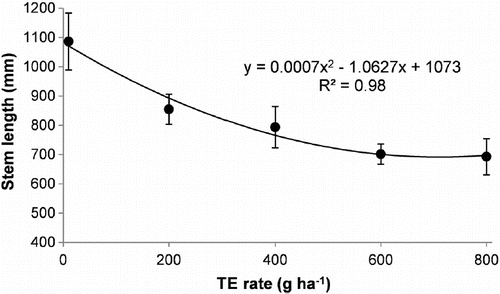
Stem length response to increasing TE application (R2 = 0.98) can be described using a polynomial curve (). At 400 g TE ha−1 (optimum rate for seed yield), stems were on average 25% shorter than the untreated control. This equates to 35 kg ha−1 of seed yield increase for every 1 cm reduction in stem length.
Figure 1. Reproductive stem length of annual ryegrass following treatment with five trinexapac-ethyl (TE) rates. Individual points are means of four replicates from each of three experiments (E2, E3 and E4) grown in Canterbury, New Zealand from 2007–09. Bars are the standard error of the mean.
Lodging
TE significantly (P < 0.05) delayed the onset of lodging and, in particular, kept the crops standing through anthesis. All rates of TE reduced lodging at early anthesis in mid December from 77% in the control treatment to 14% or less ().
Table 2. Lodging percentage of annual ryegrass at early anthesis following treatment with trinexapac-ethyl (TE) for three experiments (E1, E3 and E4) grown in Canterbury, New Zealand during the 2002–08 growing seasons. Note; 0% = reproductive stems upright, 100% = fully lodged.
Harvest mass and harvest index
Harvest mass for all treatments averaged 11.6 T DM ha−1. There was no effect on harvest mass with increasing TE rates. Harvest index (seed weight at harvest/seed weight at harvest + straw weight) increased as TE application rate increased to 400 g ha−1 in E2, and 200 g ha−1 in E3 and E4 ().
Table 3. Harvest index (%) of three annual ryegrass cultivars following treatment with five trinexapac-ethyl (TE) rates in three field experiments (E2–E4) when grown in Canterbury, New Zealand during the 2007–09 growing seasons.
Change in seed yield components
The components of seed yield for the optimum TE rate of 400 g ha−1 and the untreated control from E2–E4 are compared in . The application of 400 g TE had no effect on seed head density, spikelet numbers per head or florets per spikelet. Although TE did not increase the number of fertile florets with developing seeds, the number of saleable seeds harvested was increased by 72% from 35,100 m−2 at 0 g TE ha−1 to 60,300 m−2 at 400 g TE ha−1, driven by an increase in the number of saleable seeds per spikelet from 1.4 to 2.5 (). Individual seed weight (expressed as thousand seed weight) was reduced by 7%.
Table 4. Mean of harvest mass, reproductive stem length, lodging percentage and components of seed yield for three annual ryegrass cultivars when treated with trinexapac-ethyl (TE) at 0 or 400 g ai ha−1 in three field experiments (E2–E4) grown in Canterbury, New Zealand during the 2007–08 growing season.
Discussion
Trinexapac-ethyl had a significant effect on the seed yield of annual ryegrass with increases greater than 50% over the control treatment. At the economic optimum of 400 g TE ha−1, seed yield increased by an average of 820 kg ha−1. With a seed price of NZ$2.20 kg−1 and trinexapac-ethyl price of NZ$282 ha−1, the net profit of NZ$1804 ha−1 represents a cost:benefit ratio of 1:6.4.
The seed yield increases of >50% reported in the present study were considerably higher than previous Northern Hemisphere trials have reported for annual ryegrass (Mellbye et al. Citation2007; Rijckaert Citation2010; Macháč Citation2012). Seed crop management of annual ryegrass in New Zealand differs from that in the Northern Hemisphere experiments, particularly in the crop management during spring. In New Zealand, annual ryegrass is defoliated in spring until approximately 100 days after the shortest day. If the final defoliation is less than 100 days after the shortest day, significantly lower seed yields occur (Rolston et al. Citation2010b). These late spring defoliations remove growing points associated with early elongating reproductive tillers and delay the time for the crop to reach the growth stage when TE application is made. In the present study, TE application was between 133 and 140 days after the shortest day. In contrast, in Oregon in the United States, Mellbye et al. (Citation2007) applied TE at or beyond the same growth stage, which was reached at 110 days after the shortest day.
In Europe, seed responses of perennial ryegrass to TE were lower than results from New Zealand trials (Chynoweth et al. Citation2010, Citation2014; Macháč Citation2012). In contrast, investigations on the effect of TE on perennial ryegrass seed yield components from Oregon were similar to the response observed in New Zealand trials. Chastain et al. (Citation2014) found that stem length of perennial ryegrass was reduced with increasing rates of TE. Similar to previous New Zealand trials, Chastain et al. (Citation2014) showed an average 43% increase in seed yield at 400 g TE ha−1 with no effect on spikes m−2, spikelets spike−1 and florets spikelet−1. An improved understanding of the physiology of TE-generated seed yield increases in annual and perennial ryegrass is needed. Understanding the associated loss of aborted, undersized or shattered seed will lead to significant advances in seed science and to higher seed yields in the future. A better knowledge of the relationship between seed yield responses to stem shortening and soluble carbohydrate partitioning between the seed head and stem (Trethewey & Rolston Citation2009) is needed. This will provide information on the importance of source–sink relationships and the distance between photosynthetic tissue and the developing seed as a driver of seed yield in grass seed crops.
Seed yield response in the present study was associated with shorter stems, with a 25% reduction in stem length at 400 g TE ha−1, the economic optimum rate for seed yield. Shorter stems and reduced lodging resulted in a higher conversion of fertile florets to saleable seed and more seeds per unit area. Lodging can adversely affect seed yield, with losses recorded between 60% and 95% (Hampton et al. Citation1985; Rolston et al. Citation1997; Griffith Citation2000; Chynoweth et al. Citation2014). Reduced mid-canopy light interception due to lodging (Rolston et al. Citation2007a), especially during anthesis and early seed development, can reduce photosynthate and seed set (Trethewey et al. Citation2010). In the present study, all application rates of TE reduced lodging at anthesis, and resulted in increased seed yields. These results are similar to those previously reported for perennial ryegrass (Trethewey et al. Citation2010; Chynoweth et al. Citation2014) and annual ryegrass (Macháč Citation2012) where increased seed yields from the application of TE were associated with delayed or reduced lodging and improved number of saleable seeds per head. The percentage of seed set in our experiments was similar for all TE application rates suggesting pollination was not a TE response component. In the present study, TE had no effect on harvest mass, whereas a greater number of seeds m−2 increased harvest index at 400 g TE ha−1 by approximately 6%.
Although application of TE significantly increased seed yields in the present study, only 41% of seeds set were converted to saleable seeds. These results are similar to previous results in perennial ryegrass that showed less than 50% of fertile florets were recovered as saleable seed (Rolston et al. Citation2007b). Identifying why the remaining seeds failed to develop or to be captured as saleable seeds requires further investigation. To optimise management in ryegrass seed production crops throughout New Zealand, the relationship between the timing of the last spring defoliation (closing date), TE application rate, and genotype by environment interactions on seed yield, requires further investigation.
Conclusion
The results of this study showed that applications of plant growth regulator increased seed yields of annual ryegrass by at least 50% under New Zealand conditions. At 400 g TE ha−1, seed yield increased by an average of 820 kg ha−1 giving a net profit of NZ$1804 ha−1 which represents a cost:benefit ratio of 1:6.4. Harvest mass and the number of reproductive heads m−2 were unchanged. The seed yield response was associated with reduced lodging and shorter stems, resulting in 72% more seeds m−2, increased conversion of fertile florets to saleable seed, more seeds per head and 74% more saleable seeds per spikelet. To optimise ryegrass seed production throughout New Zealand, genotype by environment interactions on seed yield requires further investigation, including the relationship between the timing of the last spring defoliation (closing date) and TE application rate.
Acknowledgements
The authors wish to thank the growers who provided the trial sites: Experiment 1, Ron Winchester of Altonbrook Farm; Experiments 2 and 3, Derek Tod of Irwell; Experiment 4, Nick Ward of Temuka.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- AsureQuality. 2008. MAF Biosecurity New Zealand export certification standard: technical requirements—seed varietal certification 2008–2009. Lincoln: AsureQuality Ltd.
- Borm GEL, van den Berg W. 2008. Effects of the application rate and time of the growth regulator trinexapac-ethyl in seed crops of Lolium perenne L. in relation to spring nitrogen rate. Field Crop Res. 105:182–192. doi: 10.1016/j.fcr.2007.10.001
- Burbidge A, Hebblethwaite PD, Ivins JD. 1978. Lodging studies in Lolium perenne grown for seed. 2. Floret site utilization. J Agr Sci. 90:269–274. doi: 10.1017/S0021859600055349
- Chastain TG, Young WC, Garbacik CK, Silberstein TB. 2003. Seed partitioning and yield responses to trinexapac-ethyl in perennial ryegrass. Proceedings of the 5th International Herbage Seeds Conference, Gatton, Australia. p. 104–108.
- Chastain TG, Young WC III, Silberstein TB, Garbacik CJ. 2014. Performance of trinexapac-ethyl on Lolium perenne seed crops in diverse lodging environments. Field Crop Res. 157:65–70. doi: 10.1016/j.fcr.2013.12.002
- Chynoweth RJ, Rolston MP, McCloy BL. 2010. Plant growth regulators: a success story in perennial ryegrass seed crops. Seeds for Futures. Agronomy Society of New Zealand Special Publication 13/Grassland Research and Practice Series. 14:47–58.
- Chynoweth RJ, Trethewey JAK, Rolston MP, McCloy BL. 2014. Reduced stem length increases perennial ryegrass seed yield. Agronomy New Zeal. 44:61–70.
- Cooper JP. 1957. Developmental analysis of populations in the cereals and herbage grasses: II. Response to low-temperature vernalization. J Agric Sci. 49:361–383. doi: 10.1017/S002185960003834X
- Griffith SM. 2000. Changes in dry matter, carbohydrate and seed yield resulting from lodging in three temperate grass species. Ann Bot-London. 85:675–680. doi: 10.1006/anbo.2000.1125
- Hampton JC, Clemence TGA, McCloy GL. 1985. Chemical manipulation of grass seed crops. In: Hare MD, Brock JL, editors. Producing herbage seeds. Grassland research and practice series 2. Palmerston North: New Zealand Grassland Association; p. 9–14.
- Hampton JG, Hebblethwaite PD. 1985. The effect of the growth regulator paclobutrazol (PP333) on the growth, development and yield of Lolium perenne grown for seed. Grass Forage Sci. 40:93–101. doi: 10.1111/j.1365-2494.1985.tb01724.x
- Macháč R. 2012. Effects of trinexapac-ethyl (Moddus) on seed yields and its quality of eleven temperate grass species. In: Barth S, Milbourne D, editors. Breeding strategies for sustainable forage and turf grass improvements. Dordrecht, New York: Springer Science Business Media; p. 359–364.
- Mellbye ME, Gingrich GA, Silberstein TB. 2007. Use of plant regulators on annual ryegrass: the Oregon experience. In: Aamlid TS, Havstad LT, Boelt B, editors. Proceedings of the 6th International Herbage Seed Conference; Gjennestad, Norway, Bioforsk Fokus; p. 236–238.
- Rademacher W, Temple-Smith KE, Griggs DL, Hedden P. 1992. The mode of action of acylcyclohexanediones—a new type of growth retardant. Current Plant Science and Biotechnology in Agriculture. 13:571–577. doi: 10.1007/978-94-011-2458-4_68
- Rijckaert GA. 2010. Effects of plant growth regulation in seed crops of Italian ryegrass (Lolium multiflorum L.). In: Smith GR, Evers GW, Nelson LR, editors. Proceedings of the 7th International Herbage Seed Conference; Dallas, Texas, USA; p. 211–216.
- Rolston MP, Archie WJ, McCloy BL. 2003. Forage tall fescue seed yields enhanced by plant growth regulators. In: Loch DS, editor. Proceedings of the 5th International Herbage Seed Conference; Gatton; p. 109–112.
- Rolston MP, McCloy BL, Pyke NB. 2004. Grass seed yields increased with plant growth regulators and fungicides. Proc New Zeal Grassland Assoc. 66:127–132.
- Rolston MP, McCloy BL, Trethewey J. 2007a. Nitrogen—moddus interaction in perennial ryegrass seed production. Agronomy New Zeal. 37:37–44.
- Rolston MP, McCloy BL, Trethewey JAK, Chynoweth RJ. 2010b. Removing early spring emerged reproductive growing points enhances seed yield of Italian ryegrass. Agronomy New Zeal. 40:133–139.
- Rolston MP, Rowarth JS, DeFilippi JM, Archie WJ. 1994. Effects of water and nitrogen on lodging, head numbers and seed yield of high and nil endophyte perennial ryegrass. Proc Agron Soc New Zeal. 24:91–94. doi: 10.1080/03014223.1994.9517457
- Rolston MP, Rowarth JS, Young WC III, Mueller-Warrant GW. 1997. Grass seed crop management. In: Fairey DT, Hampton JG, editors. Forage seed production, volume 1: temperate species. Wallingford, UK: CAB International; p. 105–126.
- Rolston MP, Trethewey JAK, Chynoweth R, McCloy B. 2010a. Trinexapac-ethyl delays lodging and increases seed yield in perennial ryegrass seed crops. New Zeal J Agr Res. 53:403–406. doi: 10.1080/00288233.2010.512625
- Rolston P, Trethewey J, McCloy B, Chynoweth R. 2007b. Achieving forage ryegrass seed yields of 3000 kg ha−1 and limitations to higher yields. In: Aamlid TS, Havstad LT, Boelt B, editors. Proceedings of the 6th International Herbage Seed Conference; Gjennestad, Norway; p. 100–106.
- Simic A, Vučkovič S, Maletič R, Sokolovič D, Djordjevič N. 2009. The impact of seeding rate and inter-row spacing on Italian ryegrass for seed in the first harvest year. Turk J Agric For. 33:425–433.
- Trethewey J, Rolston MP, Chynoweth R, McCloy B. 2010. Light, lodging and flag leaves-what drives seed yield in ryegrass? In: Smith GR, Evers GW, Nelson LR, editors. Proceedings of the 7th International Herbage Seed Conference; Texas; p. 104–108.
- Trethewey JAK, Rolston MP. 2009. Carbohydrate dynamics during reproductive growth and seed yield limits in forage perennial ryegrass. Field Crop Res. 112:182–188. doi: 10.1016/j.fcr.2009.03.001
- Warnke S, Barker RE, Jung G, Sim S-C, Rouf Mian MA, Saha MC, Brilman LA, Dupal MP, Forster J. 2004. Genetic linkage mapping of an annual × perennial ryegrass population. Theor Appl Genet. 109:294–304. doi: 10.1007/s00122-004-1647-3
- Warringa JW, Marinissen MJ. 1996. The effect of light intensity after anthesis on dry matter distribution and seed yield of Lolium perenne L. Grass Forage Sci. 51:103–110. doi: 10.1111/j.1365-2494.1996.tb02043.x
- Wright D, Hebblethwaite PD. 1979. Lodging studies in Lolium perenne grown for seed. 3. Chemical control of lodging. J Agr Sci. 93:669–679. doi: 10.1017/S0021859600039083
- Zadoks JC, Chang TT, Konzak CF. 1974. A decimal code for the growth stages of cereals. Weed Res. 14:415–421. doi: 10.1111/j.1365-3180.1974.tb01084.x