Abstract
We measured secondary succession in permanent plots for 15 years after a lightning-induced fire in a 90-year-old frost flat heathland dominated by Dracophyllum subulatum at Rangitaiki, North Island, New Zealand. We placed this succession in the longer term context provided by earlier published data from the same site. Species re-establishment was rapid, with all of the consistently present species of mature heathland present by 15 years. The new Dracophyllum population has reached its pre-burn density, although cover is still minimal. Vascular plant cover began to decline 11 years after fire, but non-vascular cover continued to increase. Poa cita declined dramatically in abundance after 11 years. Earlier studies at the site demonstrate that nearly the entire species complement of mature heathland is present by 25 years, indication of a direct succession. With substantial resilience to fire, frost flat heathland is a fire-adapted ecosystem in a country where such ecosystems are rare.
Introduction
Until recent widespread destruction for agriculture and forestry, frost flat heathlands, a historically rare ecosystem (Williams et al. Citation2007) comprising short sclerophyllous shrublands dominated by the ericaceous shrub Dracophyllum subulatum (monoao) on well-drained, infertile volcanic soils, were characteristic of shallow basins on the North Island Volcanic Plateau (Smale Citation1990). Such sites are mantled by deep deposits of infertile rhyolitic Taupo (and earlier) tephra (200 AD). These are mostly flow-tephras, but even more infertile water-sorted tephras occur in places. Despite climates that are generally amenable for plant growth, the most biologically stressed sites are subject to year-round frosts resulting from cold air ponding. Nutrient deficiency and frostiness severely restrict the complement of species capable of occupying the most extreme sites, which are dominated by species with traits such as small leaves, slow growth rates and long lifespans that are characteristic of stress-tolerators (sensu Grime Citation1977). Droughtiness because of excessively drained soils has been suggested as another factor maintaining heathland vegetation on frost flats but, despite the empirically determined drought-tolerance of one characteristic species, Poa cita (silver tussock) (Scott Citation1969), convincing evidence for this influence is lacking, even at the most extreme sites (Smale Citation1990).
Although fire has played a major role in shaping vegetation patterns in drier regions since human arrival in New Zealand (Wardle Citation1991), there have been relatively few studies of its impacts on woody ecosystems or of their resilience to it. Beech (Nothofagus spp.) forests show little resilience to fire, with very slow recovery (Wiser et al. Citation1997) and extensive formerly forested areas in drier regions remain in induced tussock grasslands for centuries afterwards. Conifer/broadleaved forests appear more resilient, with re-establishment through protracted teatree (Leptospermum/Kunzea) successions occurring commonly in wetter regions (Esler & Astridge Citation1974; Payton et al. Citation1984; Bray et al. Citation1999; Perry et al. Citation2010) and components of the mature forest reaching the canopy by 100 years on some sites (Atkinson Citation2004). Nevertheless, they can hardly be considered ‘fire-adapted’. Few of the dominant species survive fire or burn easily and replace themselves readily after it; their re-establishment requires amelioration of site conditions by preceding seral vegetation (Wardle Citation1991).
Unusually for wetter regions, the central North Island has a long history of Polynesian burning. Early (1839) European travellers on the plateau recorded evidence of ubiquitous recent fire (see Nicholls Citation1978) with secondary shrubland and grassland communities widespread (Nicholls Citation1963). Dracophyllum subulatum, the archetypal shrub of frost flats, is highly flammable, hence the popular generic name of ‘turpentine scrub’. There is also contemporary evidence of natural fires ignited by lightning on a much smaller scale at Rangitaiki, the best-known extant frost flat (Smale Citation1990). There, elliptical patches of younger heathland occur in a matrix of older vegetation, comparable with mosaics in Calluna vulgaris (ling) heathlands in Europe (Gimingham Citation1972). The most recent fire occurred during an electrical storm in February 1994, completely razing a~ 0.3 ha of a 90 year-old shrubland before being extinguished by rain. At least five such fires occurred in the 20th century at Rangitaiki, although the total area of vegetation affected is < 1% of the total. The 1994 burn presented an opportunity to examine secondary succession after fire using permanent plots, the most robust way of documenting vegetation change (Mueller-Dombois & Ellenberg Citation1974). This data was combined with a re-appraisal of vegetation data from previous fires at the site (Smale Citation1990) to ask: In this ecosystem, characterized by slow-growing, short-statured, stress-tolerant species, how quickly does the species complement return following fire, in the presence of the ongoing management regime? That is, how resilient (sensu Pimm Citation1984) is this ecosystem to disturbance by fire?
Materials and methods
Site description
The study site in Rangitaiki Conservation Area lies at at 740 m.a.s.l at the southern end of the Kaingaroa Plateau, central North Island, New Zealand (38.91° N, 176.39° S). It consists of a gently undulating plain of some 1000 ha overlain by Taupo pumice colluvium at least 0.6–1 m deep (Pullar & Birrell Citation1973). Underlying the colluvium is ~ 6–8 m of earlier rhyolitic tephras, particularly Waimihia ash (Pullar & Birrell Citation1973). Strongly leached pumice soils derived from water-sorted pumice predominate, with coarse sandy upper horizons that are excessively drained (Rijkse Citation1986). Topsoils are moderately to strongly acidic (pH 4.9–5.5), with low to moderate carbon content (23–94 mg cm−3) with a high C:N ratio (16–25), low to moderate total nitrogen (1.2–4.5 mg cm−3) and phosphorus (0.15–0.39 mg cm−3), moderate cation exchange capacity (14), and low base saturation (22%) and exchangeable potassium (0.44 cmol(+) kg−1), calcium (2.2 cmol(+) kg−1), magnesium (0.45 cmol(+) kg−1) and sodium (0.04 cmol(+) kg−1) (M. McLeod, personal communication; Blakemore et al. Citation1987; Yeates et al. Citation2004). Climate is cool and humid, with a mean annual rainfall of 1200–1600 mm. Rainfall is relatively constant and well distributed through the year (New Zealand Meteorological Service Citation1985). Mean annual temperature is 9.4 °C with a January mean (midsummer) temperature of 14.5 °C and a July mean (midwinter) temperature of 4.1 °C. Up to 230 ground frosts per year have been recorded at nearby Iwitahi (Washbourn Citation1978); the coldest frost recorded between 1959 and 1980 was -15.8 °C in winter (June) and -7.5 °C in summer (December). Snow falls on about four days per year (New Zealand Meteorological Service Citation1980), but does not lie for long. The area experiences relatively high solar radiation, with a daily mean of 14–15 MJ m−2 (NIWA 2003; http://www.niwa.co.nz/news-and-publications/publications/all/wa/13-4/solar). The predominant vegetation cover is Dracophyllum subulatum-dominant shrubland. The site was extensively grazed until about a century ago and introduced hares (Lepus europaeus) are present in small numbers, but their current impact appears minimal. Control of environmental weeds has been undertaken by the managing agency since 1989, with mechanical removal of Pinus contorta (lodgepole pine) and Calluna vulgaris and attempted chemical control of Pilosella officinarum Vaill. (mouse-ear hawkweed).
Vegetation sampling
One week after the 1994 fire, five 2 × 2 m plots were established randomly across the burn and remeasured annually between 1994 and 2000, then less frequently. Frost flat heathland is homogeneous in terms of both vegetation and site, with little variation over wide areas (Smale Citation1990), so relatively few plots can suffice for reporting general trends. Numbers of dead Dracophyllum individuals initially present in each plot were counted, and new seedlings recorded to measure seedling density. Point intercepts of the first plant reached by a pin at 104 points in a grid were summed per species per plot and expressed as a percentage of the total number of intercepts per plot to provide a measure of percentage cover. Species recorded as present in a plot but not intercepted were given an arbitrary score of 0.01% sum cover per plot. Environmental weeds that appeared on the burn inside (Pilosella officinarum) or outside (Pinus contorta) plots were removed manually, in accordance with the ongoing management regime. Data are archived in the National Vegetation Survey (NVS) databank (http://nvs.landcareresearch.co.nz/).
Analyses
Mean percentage cover per species was averaged across the five plots in each measurement year. Likewise, mean species richness was averaged across plots in each year. To test whether successional change in composition was convergent among the five plots, we used ordination to arrange optimally each measurement (i.e. each plot in each year) in multidimensional space, based on measured composition. We included compositional data from 25-year-old lichenfield and 80-year-old shrubland in the ordination, because these represent older successional stages that were useful for gauging the extent to which our plots had recovered over 15 years. We used non-metric multidimensional scaling (NMS) in PC-Ord v.5.0 (MjM Software Design) with default settings for the ‘slow and thorough’ procedure to select the optimum number of axes (2) and to avoid local minima and minimize final stress (final stress = 15.4; McCune et al. Citation2002). The stress value obtained (15.4) is acceptable and typical for ecological datasets so long as the solution is used for general interpretation of patterns (McCune et al. 2002). We allowed the ordination to select the number of axes and ran a large number of randomizations to minimize the risk of settling on a local minima.
Cover scores were transformed before analysis using the approach recommended by McCune et al. (2002) that allows for a large number of zero values. We calculated the mean axis 1 and 2 plot scores for each year since burning, and for lichenfield and shrubland, and plotted these to illustrate the successional trajectory and the relative amount of compositional change (i.e. distance among mean plot scores) between years. We overlaid selected species scores (centroids) from the ordination to highlight the position of key species during the succession.
Species dominance in each plot at each measurement was calculated as the reciprocal of Simpson's diversity index (Lepš Citation2005); this was estimated using the vegan library as part of R v. 2.8.1. (R Development Core Team Citation2008). Data on cover abundance and Dracophyllum density from Smale (Citation1990) was used to add two later successional communities to our study.
Results
After a brief lag, total vegetation cover increased rapidly after fire (Fig. 1A initially mostly comprised of vascular species, but increasingly comprising non-vascular species. Vascular cover began to decline 11 years after fire, but non-vascular cover has continued to increase (A).
Figure 1 Changes in mean percentage cover and species richness for all species, vascular species and non-vascular species in the 15 years after fire in Dracophyllum subulatum frost flat heathland at Rangitaiki. (A) Change in mean percentage cover (± 1 SE). (B) Change in mean species richness (± 1 SE).
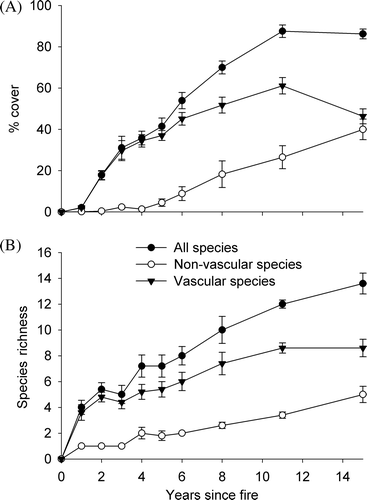
Twenty-four species, including two adventive annual herbs and Pilosella officinarum, an environmental weed here, were recorded within plots, and another 12 (half of them annual adventive herbs) on the wider burn (; B). Only two years after fire, over half of the 12 species consistently present in frost flat heathland (Smale Citation1990) had re-established, and all of them had re-established by 15 years. Total species richness and vascular species richness increased rapidly immediately after fire, and subsequently at a slower rate. Non-vascular species richness has increased at a slower, more consistent rate (B).
Table 1 Mean per cent cover by species over the 15 years after a natural fire in Dracophyllum subulatum frost flat heathland at Rangitaiki.
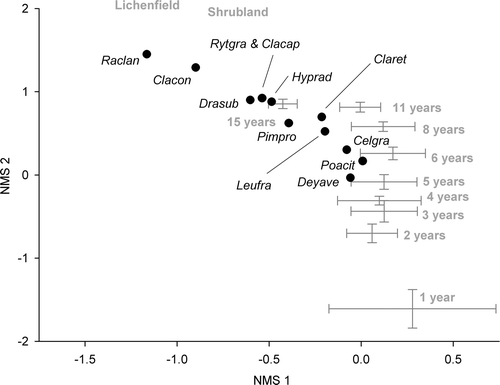
Two main patterns are apparent in the behaviour of species populations. One group, comprising Dracophyllum subulatum, Leucopogon fraseri, Pimelea prostrata, Celmisia gracilenta, Cladia retipora and Cladonia capitellata, has increased. A second, smaller group, comprising Stackhousia minima, Carex breviculmis and Poa cita, increased initially, then decreased. Deyeuxia avenoides showed an anomalous pattern of an early increase, then decreasing, but increasing again after 11 years. The remaining species showed no consistent pattern. The direction of compositional change tracked consistently until 2005, after which it has changed direction with the rapid recent decline in P. cita(). Composition has converged among the plots, with errors around means becoming smaller.
Figure 2 Compositional changes in the 15 years after fire in Dracophyllum subulatum frost flat heathland at Rangitaiki, summarized using non-metric multidimensional scaling (NMS) ordination. The optimal NMS solution had two axes and we calculated the mean axis 1 and 2 plot scores (± 1 SE) for each year since burning and for lichenfield (~ 25 years) and shrubland (~ 80 years) and plotted these to illustrate the successional trajectory. We overlaid selected species scores (centroids) from the ordination to highlight the position of key species during the succession. Mean plot scores for each year after fire and for the two older communities are labelled. Lichenfield and shrubland data are from Smale (Citation1990).
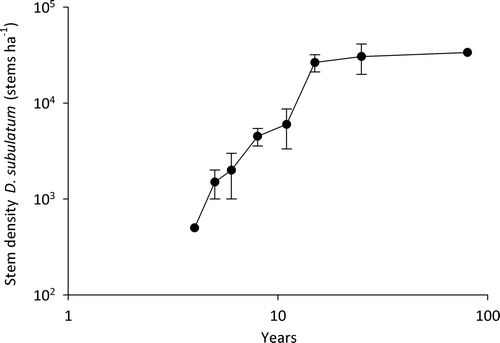
Dracophyllum seedlings, already present 3 years after the burn, increased dramatically between 11 and 15 years. Their mean density of 26 500 seedlings ha−1 is now approaching the preburn density of 32 500 individuals ha−1 (), although mean cover (1.4%) is still minimal compared with that of mature heathland (42.5%: Smale Citation1990).
Figure 3 Density of Dracophyllum subulatum (stems ha−1) over the 15 years after a natural fire in frost flat heathland at Rangitaiki. Values for ~ 25 and ~ 80 years are from Smale (Citation1990); the value for 100 years is the measured preburn density.
Discussion
The vascular flora complement of mesic frost flat heathland of some 60 species, 10 of them adventive (Smale Citation1990), is similar to that of northern gumland heaths on very infertile soils but under much milder climates (Clarkson et al. 2010). Rapid re-establishment of vegetation occurs after fire (A), with 40% of species present and Dracophyllum nearing its preburn density by 15 years (Table 1 and B) when the area is dominated by Cladia retipora (B). Virtually all species are present by ~ 25 years (Smale Citation1990), indicating direct succession (i.e. where the initial colonizers are the final dominants; Whittaker & Levin Citation1977). Re-establishment occurs mostly from newly recruited individuals; only two species, P. cita and Celmisia gracilenta, partly survived the fire, with the remainder re-establishing entirely from newly recruited individuals. Dracophyllum seedlings appeared within 4 years of the fire in the plot nearest the surrounding intact shrubland, and in all plots within 7 years. Thus most of the regeneration ‘wave’ of Dracophyllum occurred within the first 15 years after fire on this very small (0.3 ha) burn. This is much faster than on more extensive areas (tens to hundreds of hectares), where population age structures span many decades (Smale Citation1990), a reflection of the close proximity of the seed source – mature Dracophyllum plants – to the recent burn. With its inability to survive fire, feeble replacement in its absence (Smale Citation1990) and elimination by too-frequent fire, Dracophyllum may be described as an ‘obligate seeder’ (Keeley Citation1986), dependent upon a particular fire frequency for rejuvenation and population expansion. Mesic heathland of several ages is present at Rangitaiki, representing a succession of mostly anthropogenic fires of different scales. Along with the most recent (1994) natural burn, there is a substantial tract of 35-year-old lichenfield and extensive tracts of ~ 45-year-old lichenfield and ~ 100-year-old shrubland (Smale Citation1990). These communities of different ages on a uniform site give an indication of the likely development over the first 80 years after fire in mesic heathland. All consistently present species are already present in lichenfield 15 years after fire, and all of them are still widespread, although in many cases much less prominent, in mature to senescent shrubland, ~ 65 years later. A further 12 species are present by 15 years and an additional 29 species by ~ 25 years. By ~ 80 years, only nine additional species are present, of which several (Coprosma cheesemanii, Gleichenia dicarpa, Oreobolus pectinatus) are confined to damper, peaty sites and are not components of normal successions on mesic frost flats.
Figure 4 Frost flatheathland burnt by a lightning-induced natural fire at Rangitaiki, central North Island, New Zealand. A, March 1995, one year after fire; loamfield. B, March 2009, 15 years after fire; lichenfield dominated by Cladia retipora.

Floristic richness eventually declines. The mean number of species recorded in 4-m2 plots averaged 14 after 15 years (this study), 17 after ~ 25 years, but only 11 in ~ 80-year-old shrubland (Smale Citation1990). Fifteen species present in the ~ 25-year-old lichenfield were not recorded from the older shrubland. Apart from four accidentals, they are all characteristic species of open ground: three orchids (Microtis oligantha, Prasophyllum colensoi, Thelymitra longifolia), three native herbs (Raoulia aff. australis, Hypericum japonicum, Acaena microphylla) and five adventive herbs (Smale Citation1990). The decline in floristic richness with age is due primarily to autogenic influences, with nearly 20% of the ground, on average, smothered by dense Racomitrium mats by age ~ 80 years, and > 40% of it shaded beneath Dracophyllum crowns (Smale Citation1990), whose dense acidic leaf litter on the soil surface appears to inhibit the establishment of other species. Thus two-thirds of available growing space in the oldest community is, on average, effectively occupied by just two species. Conversely, the high average cover of Cladia retipora – one-third after 15 years – does not reduce species richness. Many herbaceous and semi-woody species grow in intimate mixture with this lichen, whose mats shrink regularly upon drying to expose the underlying soil surface. Changes in the relative cover of major species over time also reflect autogenic influences. For example, ground-hugging Leucopogon fraseri and Pimelea prostrata are particularly susceptible to smothering by Racomitrium and eventually decline, whereas the cover of Dracophyllum increases as its crowns expand.
In common with other extreme environments, heathland vegetation is unable to modify the environment sufficiently to facilitate the entry of other species. Frost levels beneath tall, dense Dracophyllum stands on more fertile sites (Yeates et al. 2004) are only marginally lighter than outside them (Smale Citation1990). Although Cockayne (Citation1928) described D. subulatum as having ‘deeply-descending roots’, it is shallow-rooted in the coarse sandy Otamatea soils (Rijkse Citation1986) at Rangitaiki, occupying only the uppermost 3–4 cm of the profile (MC Smale, personal observation). Deeper rooted species of more fertile soils such as Coprosma propinqua (Wardle Citation1991) are virtually absent from heathland here, so that recycling of nutrients within the profile is confined to the humus-rich uppermost horizon, which contains most of the available nutrients.
The small complement of frost-hardy species that comprise the ‘grey scrub’ (Wardle Citation1991) in dongas (dry gullies formed by water erosion) and the Rangitaiki River gorge constitute possible successors to the existing heathland, but are absent from Rangitaiki. Some (e.g. Coprosma propinqua, Olearia virgata, Aristotelia fruticosa) are probably excluded by infertility (see Wardle Citation1991), others (e.g. fire-sensitive Halocarpus bidwillii and Phyllocladus alpinus) by the very limited seed sources left after a long fire history. Common forest precursors such as Leptopermum scoparium, Kunzea ericoides var. ericoides (kanuka), Pseudopanax crassifolius and Pittosporum tenuifolium are excluded from Rangitaiki by summer frost (Reitsma Citation1994). Thus, except in the very long-term, no replacement by species other than the existing ones is likely at Rangitaiki.
The question inevitably arises of the antiquity of frost flat heathlands. Rhyolitic euptions have intermittently infilled valleys on the Volcanic Plateau to produce the characteristically frosty, infertile landscapes of ‘frost flats’ over at least the past 50 000 years (Froggatt & Lowe Citation1990). All the consistently present species of frost flat heathland are adapted to the occurrence of at least infrequent fire, by survival or by rapid establishment from wind-blown seed on bare ground. The present-day vegetation pattern at Rangitaiki indicates a long fire history, with extensive seral shrublands on adjacent ranges indicating large-scale fires continuing until early last century. Extremely cold- and infertility-tolerant Halocarpus bidwillii and Phyllocladus alpinus – the main components of shrub-heaths on frost flats from which grey scrub is believed to derive (Wardle Citation1991) – are very sensitive to fire, and the tiny extant populations of these species at Rangitaitki are relictual. Open shrubland of Halocarpus and Phyllocladus with Dracophyllum and its characteristic associates may have dominated sites like Rangitaiki before fires became more frequent with the advent of humans (McGlone and Wilmshurst Citation1999).
Southeastern Australian heathlands are also resilient to fire, with all species of the mature community present within the first season or two, although later dominants may be unimportant initially (Purdie & Slatyer Citation1976). A temporary increase in species richness after fire, comprising mostly fire and disturbance ephemerals, is also characteristic (McFarland Citation1988), eventually decreasing autogenically as long-lived woody dominants re-establish more-or-less complete canopies (Specht & Specht Citation1989). Important components of all fire regimes include intensity, frequency and season (Gill & Groves Citation1981), and the effects of these on secondary succession remain to be investigated in frost flat heathlands.
Some aggressive adventive species have the capacity to deflect frost flat successions along quite different pathways. Pinus contorta forms dense forest that ousts heathland species completely (MC Smale, personal observation); successional pathways through Pilosella officinarum are unknown. In their absence, and for the foreseeable future while the proportions of major species on frost flats, particularly Dracophyllum, may wax and wane depending on disturbance, the total floristic assemblage is unlikely to change perceptibly. Cycles similar to those in northern hemisphere Calluna heaths (Gimingham Citation1972) appear to be operating but remain to be properly demonstrated. Ground exposed by fires of natural or human origin will continue to be colonized by the same suite of species that has colonized bared ground in the past and comprises existing vegetation. Under it, soil fertility is likely to decline inexorably, if slowly, with continuing leaching from upper horizons of nutrients released by weathering and no means of rejuvenation. Biomass and productivity are likely to decline and, in this sense, frost flat heathland represents a long-term retrogressive succession, or regression. With its ability to recover its initial structure or character following fire (Pimm Citation1984), frost flat heathland is an example of an ecosystem that is resilient to fire in a country where fire-adapted ecosystems are rare.
Acknowledgements
The Department of Conservation granted permission to conduct the study. Data collection and analysis was funded by the New Zealand Foundation for Research, Science and Technology through the Sustaining and Restoring Biodiversity Outcome-Based Investment (C09X0503). PT Whaley assisted with data collection and M McLeod provided information on soils. Dr SK Wiser and anonymous referees made helpful comments on earlier drafts.
Related Research Data
References
- Atkinson , IAE . 2004 . Successional processes induced by fires on the northern offshore islands of New Zealand . New Zealand Journal of Ecology , 28 ( 2 ) : 181 – 193 .
- Blakemore LC , Searle PL , Daly BK 1987 . Methods for chemical analysis of soils. New Zealand Soil Bureau Science Report 80 . New Zealand Soil Bureau , Lower Hutt .
- Bray , JR , Burke , WD and Struik , GJ . 1999 . Propagule dispersal and forest regeneration in Leptospermum scoparium (manuka)–L. ericoides (kanuka) forests following fire in Golden Bay, New Zealand . New Zealand Natural Sciences , 24 : 35 – 52 .
- Clarkson BR , Smale MC , Williams PA , Wiser SK , Buxton RP 2011 . Infertility and fire maintain biodiversity of gumland heaths in northern New Zealand . New Zealand Journal of Ecology (in press)
- Cockayne L 1928 . Die Vegetation der Erde XIV. The vegetation of New Zealand Leipzig . Engelmann , Germany .
- Esler , AE and Astridge , SJ . 1974 . Tea tree (Leptospermum) communities of the Waitakere Range, Auckland, New Zealand . New Zealand Journal of Botany , 12 ( 4 ) : 485 – 501 .
- Froggatt , PC and Lowe , DJ . 1990 . A review of the late Quarternary silicic and some other tephra formations in New Zealand: their stratigraphy, nomenclature, distribution, volume, and age . New Zealand Journal of Geology and Geophysics , 33 ( 1 ) : 89 – 109 .
- Gill AM , Groves RH 1981 . Fire regimes in heathlands and their plant-ecological effects . In : Specht RL Heathlands and related shrublands. Ecosystems of the world 9B . Elsevier , Amsterdam, , The Netherlands . Pp. 61 84 .
- Gimingham , CH . 1972 . Ecology of heathlands , London, , UK : Chapman & Hall .
- Grime , JP . 1977 . Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory . American Naturalist , 111 ( 982 ) : 1169 – 1194 .
- Keeley JE 1986 . Resilience of Mediterranean shrub communities to fires . In : Dell B , Hopkins AJM , Lamont BB , Resilience in Mediterranean-type ecosystems . Junk , Dordrecht, , The Netherlands . Pp. 95 112 .
- Lepš , J . 2005 . “ Diversity and ecosystem function ” . In Vegetation ecology , Edited by: van der Maarel , E . 199 – 237 . Oxford, , UK : Blackwell Science .
- McCune B , Grace JB , Urban DL 2002 . Analysis of ecological communities. Gleneden Beach , OR, MjM Software Design .
- McFarland , DC . 1988 . Fire and the vegetation composition and structure of subtropical heathlands in south-eastern Queensland . Australian Journal of Botany , 36 ( 5 ) : 533 – 546 .
- McGlone , MS and Wilmshurst , JM . 1999 . Dating intitial Mäori environmental impact in New Zealand . Quaternary International , 59 ( 1 ) : 5 – 16 .
- Mueller-Dombois D , Ellenberg H 1974 . Aims and methods of vegetation ecology . New York , Wiley .
- New Zealand Meteorological Service 1980 . Summaries of climatological observations to 1980 . New Zealand Meteorological Service miscellaneous publication 177. Wellington, New Zealand Meteorological Service .
- New Zealand Meteorological Service 1985 . Climatic map series 1:2 000 000. New Zealand Meteorological Service, miscellaneous publication 175 . New Zealand Meteorological Service , Wellington .
- Nicholls , JL . 1963 . Vulcanicity and indigenous vegetation in the Rotorua district . Proceedings of the New Zealand Ecological Society , 10 : 58 – 65 .
- Nicholls JL 1978 . Vegetation In : Vucetich CG , Wells N , Soils, agriculture and forestry of Waiotapu region, central North Island. Department of Scientific and Industrial Research Soil Bureau Bulletin 31 .
- Payton , IJ , Allen , RB and Knowlton , JE . 1984 . A post-fire succession in the northern Urewera forests North Island, New Zealand . New Zealand Journal of Botany , 22 ( 2 ) : 207 – 222 .
- Perry GLW , Ogden J , Enright NJ , Davy LV 2010 . Vegetation patterns and trajectories in disturbed landscapes, Great Barrier Island, northern New Zealand . New Zealand Journal of Ecology 34 : 311 323 .
- Pimm , SL . 1984 . The complexity and stability of ecosystems . Nature , 307 : 321 – 326 .
- Pullar WA , Birrell KS 1973 . Age and distribution of late Quaternary pyroclastic and associated deposits of the Rotorua and Taupo area, North Island, New Zealand . New Zealand Soil Survey report no. 1 .
- Purdie , RW and Slatyer , RO . 1976 . Vegetation succession after fire in sclerophyll woodland communities in south-eastern Australia . Australian Journal of Ecology , 1 : 223 – 236 .
- R Development Core Team , 2008 . R: a language and environment for statistical computing . R Foundation for Statistical Computing , Vienna, , Austria . http://www.R-project.org .
- Reitsma , L . 1994 . Frost resistance of some native plants from the central Volcanic Plateau, North Island, New Zealand, in relation to plant succession . New Zealand Journal of Botany , 32 ( 2 ) : 217 – 226 .
- Rijkse WC 1986 . Soils of the Taupo region: interim soil maps and legend. New Zealand Soil Bureau District Office report . New Zealand Soil Bureau , Lower Hutt .
- Scott , D . 1969 . Effect of soil moisture tension on leaf elongation in Poa caespitosa . New Zealand Journal of Botany , 7 ( 1 ) : 103 – 105 .
- Smale , MC . 1990 . Ecology of Dracophyllum subulatum-dominant heathland on frost flats at Rangitaiki and north Pureora, New Zealand . New Zealand Journal of Botany , 28 ( 3 ) : 225 – 248 .
- Specht , RL and Specht , A . 1989 . Species richness of sclerophyll (heath) plant communities in Australia – the influence of overstorey cover . Australian Journal of Botany , 37 ( 4 ) : 337 – 350 .
- Wardle , P . 1991 . Vegetation of New Zealand , Cambridge : Cambridge University Press .
- Washbourn , RW . 1978 . Establishment practice on frost-prone sites at Kaingaroa Forest . New Zealand Journal of Forestry , 23 ( 1 ) : 107 – 120 .
- Whittaker , RH and Levin , SA . 1977 . The role of mosaic phenomena in natural communities . Theoretical Population Biology , 12 : 117 – 139 .
- Williams , PA , Wiser , SK , Clarkson , BR and Stanley , MC . 2007 . New Zealand's historically rare terrestrial ecosystems set in a physical and physiognomic framework . New Zealand Journal of Ecology , 31 ( 2 ) : 119 – 128 .
- Wiser , SK , Allen , RB and Platt , KH . 1997 . Mountain beech forest succession after a fire at Mount Thomas Forest, Canterbury, New Zealand . New Zealand Journal of Botany , 35 ( 4 ) : 505 – 515 .
- Yeates , GW , Schipper , LA and Smale , MC . 2004 . Site condition, fertility gradients, and soil biological activity in a New Zealand frost flat heathland . Pedobiologia , 48 ( 2 ) : 129 – 137 .