Abstract
The influences of geology and land use on macroinvertebrate communities were investigated in the Motueka River catchment, New Zealand. Comparisons of functional feeding groups, multivariate community composition and biotic indices (MCI, QMCI, %EPT) were made between native forest stream reaches that differed in subcatchment geology (ultramafic, hard sedimentary, granite, gravel, karst) and between stream reaches surrounded by different land uses (native forest, exotic forest and pasture) within two of these geologies (granite, gravel). Differences in invertebrate community density and taxon richness were greater across the three land uses in the same geology than among the five geologies. Macroinvertebrate communities in streams flowing through native forest had greater numbers of shredders, and higher MCI scores than pastoral streams. Exotic forest macroinvertebrate communities were similar to the native forest communities in granite geology, but distinct from both native and pastoral communities in sites with underlying gravel geology. The results demonstrate that there is potential for underlying geology to influence the magnitude of land use impacts on invertebrate communities. The geologies within a catchment should be accounted for when considering the consequences of land development on river and stream communities.
Introduction
With increasing human demand on water resources, finding the balance between maximising economic development (gained through various land use activities) while maintaining instream ecosystem integrity has become increasingly challenging. The decision making process for the best management of water resources is difficult, as the impact of differing land uses or activities on a waterway will depend on the large-scale catchment features that contribute to a waterway's natural state. In the absence of the impacts of human activities, geology, climate and vegetation are arguably the three main controlling factors that will most influence stream ecosystems, through large-scale controls on chemistry, sediment delivery and hydrology (Allan & Johnson Citation1997; Richards et al. Citation1997; Townsend et al. Citation1997, Citation2003; Davies et al. Citation2000). Human activity and associated land use practices have become a fourth controlling factor on stream ecosystem functioning (Townsend et al. Citation1997). Geology influences stream geomorphology, hydrology (e.g. groundwater storage capacity), sedimentation and water chemistry (e.g. through the release of minerals and trace elements from weathered rocks) at a catchment scale (Leopold Citation1994; Gordon et al. Citation1992; Mosley & Pearson Citation1997). Geology has been included in studies of catchment-scale influences on macroinvertebrate community structure (Quinn & Hickey Citation1990a, Citationb; Richards et al. Citation1997; Davies et al. Citation2000) and of biomass and productivity, the latter in relation water physicochemistry (e.g. Eggert & Burton Citation1994; Huryn et al. Citation1995).
The effect of land use on macroinvertebrate communities has been well documented, particularly the degradation of stream communities when native forest is converted to pasture (Quinn & Hickey Citation1990a; Quinn Citation2000; Huryn et al. Citation2002). Stream macroinvertebrate communities in mature exotic forest can be similar to those found in native forest (Harding & Winterbourn Citation1995) but, during and for several years following logging, these communities more closely resemble those of pastoral streams (Graynoth Citation1979; Death et al. Citation2003; Reid et al. Citation2010).
Allan & Johnson (Citation1997) stressed the importance of quantifying land use and geological influences separately so there is a separation of natural landscape influences versus those related to human activities. They cautioned that studies quantifying only land use impacts on macroinvertebrate communities risk attributing causation to land use that is in fact associated with the underlying geology. With increasing diversification and intensification of land use worldwide, understanding the links between geology and land use on aquatic communities is becoming an increasingly important consideration. The magnitude of impact related to land use intensification may differ depending on the underlying geology. A change in land use to pasture may affect invertebrate communities in a stream regardless of geology type, for example through a reduction in stream shade, increases in water temperature and nutrient input. However, potential sedimentation impacts would be expected to be less severe in a pastoral stream flowing through a stable geology than one draining an unstable, or highly erodable, geology.
In this study we examined the influence of geology and land use within a single river catchment by comparing macroinvertebrate communities in: (1) five different geology types within a single land use (native forest), and (2) three contrasting land uses within two of those geology types.
Methods
Study area
Situated in the north of the South Island of New Zealand (), the Motueka River drains a catchment of 2180 km2 and flows for approximately 110 km from the headwaters to Tasman Bay. The catchment is geologically complex, with ultramafic and sedimentary rock (primarily argillite) in the southeastern headwaters, a complex array of sedimentary rocks underlying the western tributaries including marble and limestone (karst), a band of granitic rocks (Separation Point granite) down the western centre of the catchment and a large band of thick (up to 0.7 km) alluvial gravel and clay (Moutere gravels) down the eastern centre of the catchment (Basher Citation2003).
Figure 1 Location of study sites in the Motueka River catchment. Geologies: SG, Separation Point granite; MG, Moutere gravel; K, karst; HS, hard sedimentary; UM, ultramafic. Land use sites indicated by: N, native forest; E, exotic forest; P, pasture. a, b, and c refer to the replicate streams for each geology and geology×land use combination ().
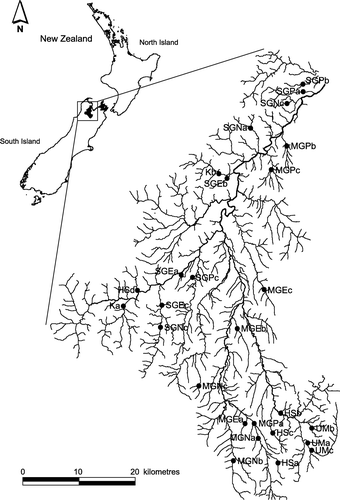
Land cover in the catchment is dominated by native forest (35%), exotic forest (25%), pastoral grassland (19%), with lesser areas in scrub (12%) and tussock grassland (7%) (NZ Landcover Database 2, Terralink International). The largest areas of native forest (dominated by Nothofagus spp.) are found in the western and southern headwaters, whereas exotic forest (primarily Pinus radiata) occupies part of the Separation Point granite and Moutere gravel terrains. Pastoral grassland generally occupies the valley floors and is mainly used for sheep and beef cattle grazing, although there are small areas where dairy farming occurs. Mean annual rainfall is about 1600 mm. However, there are strong rainfall gradients across the catchment with the highest rainfall (3500 mm/year) occurring in the western headwaters. In contrast, the low-elevation eastern parts of the catchment receive <1000 mm/year of rainfall (Basher Citation2003). The first part of this study was conducted in five main geology types draining native forest subcatchments. These were the igneous rock geologies; ultramafic and Separation Point granite, and the sedimentary rock geologies; karst, hard sedimentary and Moutere gravel (, ). The main vegetative cover in these geologies is beech forest (Nothofagus sp.), but high levels of magnesium and nickel levels are found in the soils formed from ultramafic rock, which appear to be toxic to plants resulting in sparse, stunted vegetative cover (Robinson et al. Citation1996). The River Environment Classification (REC) system (Snelder & Biggs Citation2002; Snelder et al. Citation2004) was used to classify the geology types of streams, and three streams within each geology type were chosen for sampling. However, after sampling, we found that one of the karst streams was misclassified, so it was included in the hard sedimentary group in subsequent analyses (). Three land use categories were chosen for consideration of the interaction between geology and land use. These were: (1) native (N), which included native forest, tussock and scrub; (2) exotic pine forest (E) consisting mainly of mature Pinus radiata that had not been harvested for 25 + years; and (3) pastoral land (P), which was predominately planted in exotic grass species and grazed by sheep and cattle. An assessment of the interaction between geology and land use was restricted to Moutere gravel and Separation Point granite geologies since exotic forestry and pastoral land use were rare or absent on the other geologies (, ). Pasture is also relatively rare in Separation Point granite geology and despite choosing sites with the highest available proportion of the catchment in pasture land cover, these sites ranged from only 17% to 37% pasture. Three streams were sampled in each land use type in both geologies ().
Table 1 Location and physical characteristics of the sampling sites.
Sampling in each stream was conducted in autumn and involved collection of macroinvertebrate, water chemistry and physicochemical information during baseflow conditions.
Biological measurements
Triplicate Surber samples (0.0625 m2; 0.5-mm mesh) were collected at randomly chosen locations within riffle habitats in each stream to assess macroinvertebrate community density and composition. Samples were preserved in 70% alcohol, and later macroinvertebrates were identified to the lowest possible taxonomic level, counted and recorded in the laboratory. Taxa were classified to functional feeding groups based on Cowie (Citation1980), Winterbourn et al. (Citation1984), Chadderton (Citation1988), Lester et al. (Citation1994) and Jaarsma et al. (Citation1998). Some taxa span functional feeding groups, e.g. Olinga (shredder/collector-browser) and Austroperla (shredder/collector-browser).
Factors affecting macroinvertebrate communities
Using GIS, the land use types in the subcatchments upstream of each sampling site were determined from the New Zealand Land Cover database (Terralink International Limited, Wellington, New Zealand), which has a minimum mapping unit of 1 ha and was derived from SPOT satellite images taken in 1997. The geological compositions of the subcatchments were similarly determined from the New Zealand Land Resource Inventory, which has a minimum mapping unit of 20 ha (Newsome Citation1995). Several water chemistry parameters were measured in the field during macroinvertebrate collection to help understand any patterns in the results. Specific conductivity at 25 °C (µS/cm) was measured at each site using a YSI85 hand-held field probe (Yellow Springs Instruments Incorporated, Yellow Springs OH, USA.). Turbidity (NTU) was measured using a HACH 2100P turbidity meter (Hach Company, Loveland CO, USA). Water samples were also collected for analysis of nitrate-nitrogen (NO3-N, g/m3), ammoniacal nitrogen (NH4-N, g/m3 ), total nitrogen (TN, g/m3), dissolved reactive phosphorus (DRP, g/m3), total phosphorus (TP, g/m3) and faecal indicator bacteria (Escherichia coli (E. coli), cfu/100 ml). Concentrations of NO3-N, NH4-N and DRP were determined using cadmium reduction (APHA Citation1998; 4500-NO3 I) and ammonium molybdate–ascorbic acid (APHA Citation1998; 4500-P G) flow injection methods, respectively.
In order to provide a measure of periphyton biomass for benthic browsing invertebrates, epilithic chlorophyll a was assessed by collecting 15-cm2 scrapings from each of five randomly selected stones in the vicinity of the Surber sample locations. Chlorophyll a concentrations were determined using the method of Biggs (Citation1995).
Data analysis
Macroinvertebrate densities, taxon richness, percentage Ephemeroptera, Trichoptera and Plecoptera (%EPT) taxa, functional feeding group proportions and two biotic indices, the Macroinvertebrate Community Index (MCI; Stark Citation1985) and Quantitative Macroinvertebrate Community Index (QMCI; Stark Citation1993) were calculated for each stream site. The MCI and QMCI are widely used in New Zealand as indicators of invertebrate community health (Boothroyd & Stark Citation2000). These indices allocate scores to macroinvertebrate taxa based on their relative sensitivities to pollution from 1 (pollution tolerant) to 10 (pollution sensitive). The MCI is calculated from presence/absence data and gives community sensitivity scores that theoretically range from 20 (indicating extremely poor water quality conditions) to 200 (indicating extremely good water quality conditions). The QMCI is calculated in a similar way but accounts for the presence and percentage community composition of taxa in a sample, with community sensitivity scores ranging from 1 to 10.
Histogram plots of the invertebrate data were inspected prior to analysis. Taxa richness and density were log10-transformed, and functional feeding groups arcsine√x transformed to improve normality. Percentage EPT, MCI and QMCI were not transformed. Statistical analyses were undertaken using STATISTICA version 8 (StatSoft, Incorporated, Tulsa, OK, USA).
Geological comparisons were made among the ultramafic, hard sedimentary, karst Moutere gravel and Separation Point granite native forest sites. A nested analysis of variance (ANOVA) was used to determine differences in macroinvertebrate communities (MCI, QMCI,%EPT, density, taxa richness, functional feeding group composition) among geologies and between streams nested within each geological category. Post-hoc contrasts using Tukey's Honestly Significant Differences (HSD) test were made, where ANOVA indicated that significant main effects differences existed.
The influence of land use (native forest, exotic forest, pasture) and geology (Moutere gravel and Separation Point granite) and any interactions on macroinvertebrate community characteristics were assessed using a full-factorial two-way ANOVA. The geology×land use interaction term was the main model component of interest. Post-hoc contrasts were also calculated between land uses using Tukey's HSD test.
Multivariate analysis of macroinvertebrate communities at all sites was also performed by canonical correspondence analysis (CCA) using R (version 2.10.0, http://www.r-project.org).
Results
Comparisons among native streams draining different geological types
Only five of a total of 71 taxa were found in common among all geologies: the mayflies Deleatidium spp. and Nesameletus spp., the stonefly Zelandoperla decorata (Tillyard), the dobsonfly Archichauliodes diversus (Walker) and the net-spinning caddisfly Aoteapsyche spp. An additional 20 taxa were found in common among all geologies except ultramafic.
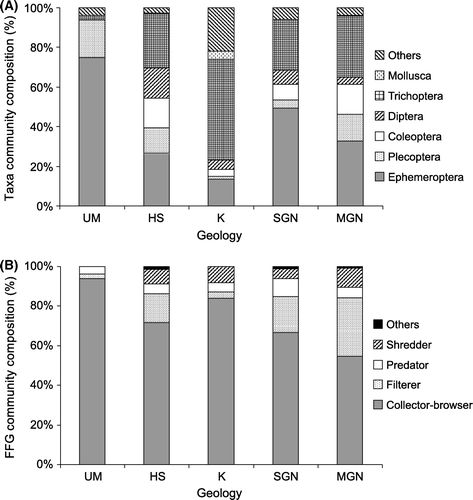
Ephemeroptera and Trichoptera dominated the macroinvertebrate community compositions in the native streams draining almost all geologies (a). The exception was the ultramafic group of sites where Plecoptera contributed a higher percentage of the community composition than Trichoptera (a). In native forest, EPT taxa represented over 65% of the communities in all of the geology types and over 95% at ultramafic sites (a).
Figure 2 Percentage community composition of orders (A) and functional feeding groups (B) for geology. The order group ‘Others’ includes the following taxonomic groups: Megaloptera, Odonata, Annelida, Hirudinea, Nematoda, Nemertea, Platyhelminthes, Crustacea, Acarina, Collembola and Coelenterata. The functional feeding group ‘Others’ includes the following: Detritivores, Piercer and Omnivores.
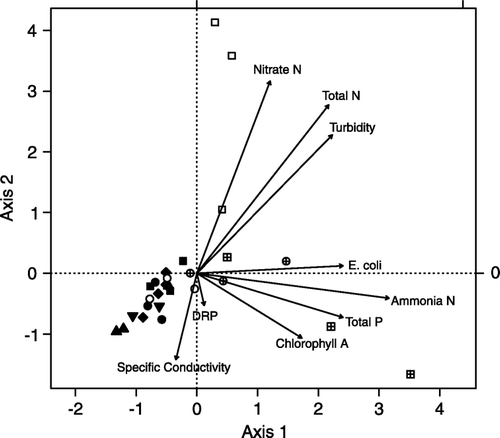
A CCA showed that the invertebrate communities of native forest sites were relatively similar across geology types, in comparison with those communities in exotic forest and pasture ().
Figure 3 Canonical Correspondence Analysis for macroinvertebrate assemblages plotted with physico-chemical parameters for each site: ▴ UM, ♦ HS, ▾ K, • SGN, ▪ MGN. Filled symbols denote native forest, clear symbols exotic forest, and crossed symbols pastoral sites.
There were significant differences among geologies for percentage composition of collector-browsers and filterers in native forest streams (). The percentage composition of collector-browsers in the ultramafic streams was significantly higher than in the hard sedimentary, Moutere gravel, Separation Point granite and karst sites (P<0.05) (b). Conversely, the percentage composition of filterers was significantly lower in ultramafic sites than hard sedimentary and Moutere gravel sites (b).
Table 2 Comparison of macroinvertebrate communities from different geologies in the Motueka river catchment using nested analysis of variance (ANOVA).
Very low densities of invertebrates were collected in the ultramafic streams compared with streams in the other geologies (a). Taxon richness was also lower in the ultramafic streams (b). To test whether this was a geological difference, or an artefact of the low densities of invertebrates collected, we performed a rarefaction analysis using EcoSim (Gotelli & Entsminger Citation2001), comparing the average taxon richness and density data of the ultramafic streams to each of the other geologies in turn. This analysis indicated that species richness in the ultramafic sites was still significantly lower than might have been expected had a similar density of invertebrates been collected from sites in the other geology types.
Figure 4 Mean (±standard error) density (A), taxa richness (B), percentage EPT (C), MCI (D), and QMCI (E) for geology. Site groups that share the same letter are not significantly different.
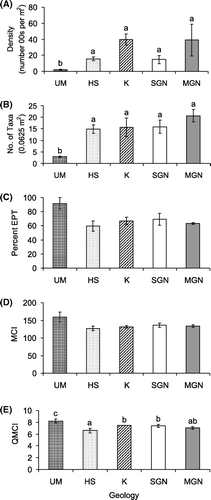
MCI and QMCI scores and % EPT taxa were all somewhat higher at the ultramafic sites (c-e). However, this difference was only statistically significant for QMCI values (, e). Post-hoc tests indicated that QMCI scores were significantly higher in the ultramafic geology than all the other geologies, and that the hard sedimentary QMCI values were also significantly lower than those from the karst and Separation Point granite geologies.
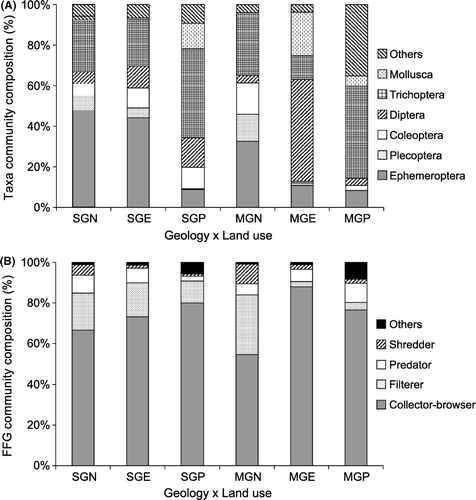
Figure 5 Percentage community composition of orders (A) and functional feeding groups (B) for geology×land use. The order group ‘Others’ is explained in .
Interactions between land use and geology
Moutere gravel and Separation Point granite were the two geologies examined for interaction with land use. A CCA showed a distinct separation of invertebrate communities between pastoral and native forest streams along Axis 1 (). The only exception was a Moutere gravel pastoral stream, where invertebrate communities more closely resembled those of the native and exotic forest streams communities (). This stream had the lowest proportion of the catchment in pasture of all the pastoral streams (). Pastoral stream invertebrate communities were associated with high concentrations of phosphorus, ammonia, total N, nitrate N and E. coli, high turbidity and high levels of chlorophyll a on the substrate (). Separation Point granite exotic forest stream communities were most similar to the native forest stream grouping of both geologies (). On the other hand, Moutere gravel exotic forest stream communities plotted closer to pastoral streams on Axis 1, but grouped separately from all other sites along Axis 2. The Moutere gravel exotic forest streams were dominated by Diptera [Orthocladiinae, Tanytarsus vespertinus (Hutton)] and molluscs [Potamopyrygus antipodarum (Gray)], whereas mayflies (in particular, Deleatidium spp.) were a major component of the invertebrate communities in exotic forest streams draining Separation Point granite (a).
There were no significant interactions between geology and land use for any of the functional feeding groups or invertebrate community metrics (), despite a tendency towards a smaller percentage of filter feeders in the exotic forest and pasture streams draining Moutere Gravel geology than streams with the same land use draining Separation Point Granite geology (P=0.054). Land use had a significant effect on the percentage of shredders in the invertebrate community (), with native forest sites having a higher percentage of shredders than the exotic forest and pasture sites (b). There was also a significant effect of land use on MCI (, d), with scores generally highest in native forest and lowest in pasture, whereas exotic forest sites were generally intermediate. MCI scores for Moutere gravel streams draining exotic forest were close to scores obtained in the pastoral streams in both geologies, whereas the scores from Separation Point granite streams draining exotic forest were closer to those for native forest streams (d). Land use appeared to influence %EPT taxa and QMCI scores with the native forest sites generally scoring more highly (c, e). However, statistically the %EPT taxa and QMCIs between the land uses were similar (P=0.068 and 0.083, respectively) ().
Figure 6 Mean (±standard error) density (A), taxa richness (B), percentage EPT (C), MCI (D) and QMCI (E) for geology×land use.
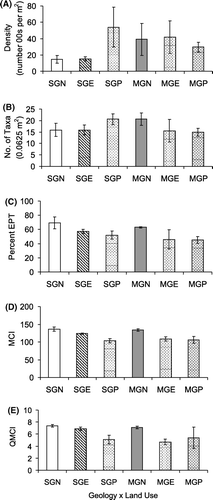
Table 3 Comparison of macroinvertebrate communities from different geology and land uses in the Motueka River catchment using two-way analysis of variance (ANOVA)
Discussion
Relationship between macroinvertebrate community structure and geology
We found differences in macroinvertebrate community structure among different geological types within native forest subcatchments, in particular the ultramafic sites. The generally higher altitude of the ultramafic streams (>610 m a.s.l.; ) may account for the differences in invertebrate diversity and density compared with sites in the other geologies. Jacobsen et al. (Citation1997) and Monaghan et al. (Citation2000) found that altitude influenced invertebrate community structure and taxa richness, but the differences in site elevations in our study were much smaller than those in their studies (100–4000 m and 780–3940 m). In addition, in Rocky Creek, a hard sedimentary stream that was also sampled above 600 m a.s.l., species diversity and density were similar to that of other streams in its geology group. Furthermore, the density and diversity of invertebrates in the ultramafic streams were also lower than has been found in other New Zealand rivers and streams with similar elevations (e.g. Quinn and Hickey Citation1990b; Harding et al. Citation1997). This suggests other features of the stream environment were a primary reason for the low diversity and density of macroinvertebrate communities in the ultramafic streams.
As mentioned earlier, the sparse, stunted vegetative cover in the ultramafic geology has been attributed to high magnesium and nickel levels in the soils formed from ultramafic rock, which appear to be toxic to plants (Robinson et al. Citation1996). Riparian vegetation can be an important source of energy for stream food webs (Maridet et al. Citation1998; Tabacchi et al. Citation1998). The absence of some invertebrate taxa such as shredders in the ultramafic streams may be related to the sparse riparian and subcatchment vegetation near the streams. Although we did not measure the levels of trace elements in the ultramafic streams, it is possible that toxic effects of these trace metals may be responsible for the low diversity and density of macroinvertebrates in these streams (see also CitationGillespie et al. this issue). The two most prominent orders that we found in ultramafic streams (Ephemeroptera and Plecoptera) are considered to have a higher tolerance to trace elements and metal toxicants than those of other invertebrate groups (Hickey & Vickers Citation1992; Hickey & Clements Citation1998; Beasley & Kneale Citation2003).
Biotic indices (such as the MCI and QMCI) are generally designed to detect the effects of organic enrichment (e.g. Chessman Citation1985; Hilsenhoff Citation1987; Stark Citation1985, Citation1993, Citation1998) on stream communities. It was, therefore, not surprising that MCI scores were relatively similar among the geologies, given that all of the samples were deliberately taken in native forest streams. In all cases, the MCI and QMCI scores for the geologies were high (>120) and indicative of clean water environments (Stark Citation1998), and typical of headwater streams with little catchment development, low nutrient levels and very low periphyton biomass (Quinn & Hickey Citation1990b). Although macroinvertebrate densities and diversity were much lower in the ultramafic sites compared with hard sedimentary and Moutere gravel sites, the macroinvertebrates that were present generally had high sensitivity scores and/or belonged to one of the EPT groups, which resulted in QMCI and%EPT values being higher at the ultramafic sites than at sites draining the other geology types.
Relationship between geology and land use on macroinvertebrate communities
In native forest systems, we demonstrated that geology was a factor influencing macroinvertebrate community structure. However, we found the influences of gravel and granite based geologies were small in relation to those of land use, which appeared to have an overriding influence on invertebrate community structure, particularly in streams that flowed through pastoral farmland. The percentage of shredders, and biotic indices (in particular MCI) were predictably higher in the native forest streams than the pastoral streams in both the Moutere gravel and Separation Point granite geologies, as has been found elsewhere (Quinn & Hickey Citation1990a; Scott et al. Citation1994, Harding & Winterbourn Citation1995; Quinn et al. Citation1997; Townsend et al. Citation1997; Collier et al. Citation2000; Quinn Citation2000; Hall et al. Citation2001).
In the CCA, the composition of invertebrate communities in pastoral streams was distinct from native forest streams. Invertebrate communities in the exotic forest streams appeared to overlap with the native forest stream on Separation Point granite, but for the Moutere gravel sites the exotic forest communities appeared to be distinct from the native and pasture streams. Several studies have found that macroinvertebrate communities in pine forested streams more closely resemble those in neighbouring native forest streams than pastoral streams, as was the case in the Separation Point granite streams (Harding & Winterbourn Citation1995; Friberg et al. Citation1997; Quinn et al. Citation1997).
Although the parameters we measured and resulting ANOVAs do not support the hypothesis that invertebrate communities in the Moutere gravel and Separation Point granite exotic forest streams were influenced by an interaction between geology and land use, underlying geological features may still provide an explanation for the separation of the exotic forest macroinvertebrate communities seen in the CCA. Susceptibility to erosion and water storage capacity are two geological features that are known to differ between the Moutere gravel and Separation Point granite geologies (Basher Citation2003). No data was collected in this study that assessed erosion or water storage of these geologies, but information on these features in the existing literature can provide some insights on how geology may have influenced exotic forest stream macroinvertebrate communities. Separation Point granite is deeply weathered and prone to erosion. In comparison, Moutere gravels are composed of greywacke and sandstone clasts in a silt/clay matrix that cements the clasts together making them less prone to erosion (Basher Citation2003). Since there had been no major disturbance events (e.g. logging, flooding) prior to sampling, it was most likely that any sediment yields to streams varied according to the resilience of rocks and soils to weathering and erosional processes (Hicks & Griffiths Citation1992).
Differences in water storage capabilities of the two geologies may have affected flow permanence in the exotic forest streams. Separation Point granite is deeply fractured allowing storage of water that is slowly released to sustain stream flow throughout the year. In contrast, Moutere gravels store limited amounts of water and surface flows in streams tend to become intermittent in summer (Duncan Citation1995; Basher Citation2003). By way of comparison, in the Moutere gravel streams specific discharge during dry periods ranged from 0.3 to 0.7 l/s/km2 and for the Separation Point granite streams from 8 to 13 l/s/km2 (Basher Citation2003). Furthermore, water uptake by mature exotic forest in the Moutere gravels has been shown to lower flood peaks dramatically and create longer low flow periods (compared with the previous land use of pasture and gorse) in response to the higher interception and evaporation rates from forest canopies (Pearce & Rowe Citation1979; Fahey & Rowe Citation1992; Duncan Citation1995). Low or intermittent flows can have slight to severe effects on macroinvertebrate communities, depending on the frequency, extent and timing of the low flow, flow permanence in the hyporheos, and on the recolonising abilities of macroinvertebrates when normal flows resume (Iversen et al. Citation1978; Sagar Citation1983; Lake Citation2003; Datry et al. Citation2007; Dewson et al. Citation2007). Macroinvertebrate communities in intermittent flow environments are likely to have less diversity than in perennial environments and especially so if sub-surface flow permanence is low (Datry et al. Citation2007; Storey & Quinn Citation2008). When flow resumes in intermittent environments, the main sources of invertebrate colonisation will be through aquatic invertebrate drift, aerial colonisation, upstream migration and vertical migration through the hyporheos (Williams & Hynes Citation1976; Sagar Citation1983; Fowler Citation2002, Citation2004). The lower diversity and density of invertebrates in one of the Moutere gravel exotic forest streams and dominance of particular groups of invertebrates (Diptera, Potamopyrgus) that are often associated with ephemeral streams in the other two streams could be an indication of invertebrate communities in the process of recovery from a period of flow intermittence (Storey & Quinn Citation2008).
Functional feeding groups in relation to land use
The paucity of shredders in invertebrate communities is a well known feature of New Zealand streams and rivers (Winterbourn et al. Citation1981; Ryder & Scott Citation1988; Quinn & Hickey Citation1990b). Although shredders were never a dominant part of any of the communities in our study, there were noticeably more shredders in the native forested subcatchments than in exotic forested or pasture streams. The only exception to this was the lack of shredders found at the ultramafic sites, most likely related to the stunted vegetative growth resulting in lower inputs of organic material into the streams. The influence of land use on shredder abundance has been alluded to in other New Zealand studies. Harding & Winterbourn (Citation1995) and Townsend et al. (Citation1997) found a greater abundance of facultative shredders and higher standing stocks of organic materials such as wood and leaves in beech and pine forest streams compared with pastoral streams. In this study, shredders formed a greater proportion of the community composition in native forest sites than pastoral sites. However, in contrast to Harding & Winterbourn's (1995) findings, shredders were not a major part of the exotic forest communities in our study.
Inputs of terrestrial organic materials are often significantly reduced in pastoral streams compared with pine and beech forest streams, and would explain the greater abundance of shredders in streams with native forest compared with pastoral streams (Harding & Winterbourn Citation1995; Quinn et al. Citation1997; Townsend et al. Citation1997). The caddis fly Olinga was the numerically dominant facultative shredder taxon that accounted for the significantly higher proportion of shredders found in the native forest than exotic forest streams. This is consistent with the findings of Harding & Winterbourn (Citation1995) who reported that Olinga feredayi was more common in NZ native forest than exotic forest streams.
Relative importance of geology and land use
Our results indicate that underlying geology has a small influence on the structure of macroinvertebrate communities relative to the overlying land use. These findings are consistent with studies that have shown changes in land use can mask geological influences on macroinvertebrate communities (e.g. Quinn & Hickey Citation1990b; Richards et al. Citation1997; Townsend et al. Citation1997, Citation2003). However, in our study, there was evidence to suggest macroinvertebrate communities in streams flowing through mature pine forests differed between two geologies under baseflow conditions. This provides support for the thesis that there can be an underpinning geological influence on macroinvertebrate communities in one land use type that may not be apparent in others (such as native forest and pastoral settings).
Many of the effects of land use modification on aquatic communities are well known for exotic forestry and pastoral development. However, the potential interplay between geology and land use is often not accounted for in the land development decision making process. Our study provides evidence that the geologies within a catchment should not be overlooked when considering the consequences of land development on river and stream communities.
Acknowledgements
We thank Renée Strickland and Aaron Quarterman for assistance in the field and Judy McKenzie for processing the macroinvertebrate samples. We also thank the Department of Conservation and land owners who allowed access to sites through their land. Some of the water chemistry results were obtained from Tasman District Council. Eric Goodwin provided technical assistance with multivariate analysis. Constructive comments by John Hayes, Dean Olsen and two anonymous reviewers helped to improve this manuscript. This project was funded by the New Zealand Foundation for Research, Science and Technology as part of the Motueka Integrated Catchment Management (ICM) Project (Contract No. C09X0214 and C09X0305).
Related Research Data
References
- Allan , JD and Johnson , LB . 1997 . Catchment-scale analysis of aquatic ecosystems . Freshwater Biology , 37 : 107 – 111 .
- APHA , 1998 . Standard methods for the examination of water and wastewater. 20th edition . Clesceri LS , Greenberg AE , Eaton AD . American Public Health Association, the American Water Works Association and the Water Environment Federation .
- Basher LR 2003 . The Motueka and Riwaka catchments: a technical report summarising the present state of knowledge of the catchments, management issues and research needs for integrated catchment management . Landcare Research, Lincoln .
- Beasley , G and Kneale , PE . 2003 . Investigating the influence of heavy metals on macroinvertebrate assemblages using Partial Canonical Correspondence Analysis (pCCA) . Hydrology and Earth System Sciences , 7 : 221 – 233 .
- Biggs , BJF . 1995 . The contribution of flood disturbance, catchment geology and land use to the habitat template of periphyton in stream ecosystems . Freshwater Biology , 33 : 419 – 438 .
- Boothroyd I , Stark J 2000 . Use of invertebrates in monitoring . In : Collier KJ , Winterbourn MJ . New Zealand stream invertebrate ecology and implications for management . Caxton Press , Christchurch , . Pp. 344 – 373 .
- Chadderton , WL . 1988 . Faunal and chemical characteristics of some Stewart Island streams . New Zealand Natural Sciences , 15 : 43 – 50 .
- Chessman , BC . 1985 . Rapid assessment if river using macroinvertebrates: A procedure based on habitat-specific sampling, family level identification and a biotic index . Australian Journal of Ecology , 20 : 122 – 129 .
- Collier , KJ , Smith , BJ , Quinn , JM , Scarsbrook , MR , Halliday , NJ , Croker , GF and Parkyn , SM . 2000 . Biodiversity of stream invertebrate faunas in a Waikato hill-country catchment in relation to land use . New Zealand Entomologist , 23 : 9 – 22 .
- Cowie B 1980 . Community dynamics of the benthic fauna in a West Coast stream ecosystem . PhD thesis , University of Canterbury , Christchurch .
- Datry , T , Larned , ST and Scarsbrook , MR . 2007 . Responses of hyporheic invertebrate assemblages to large-scale variation in flow permanence and surface–subsurface exchange . Freshwater Biology , 52 : 1452 – 1462 .
- Davies , NM , Norris , RH and Thoms , MC . 2000 . Prediction and assessment of local stream habitat features using large-scale catchment characteristics . Freshwater Biology , 45 : 343 – 369 .
- Death , RG , Baillie , B and Fransen , P . 2003 . Effect of Pinus radiata logging on stream invertebrate communities in Hawke's Bay, New Zealand . New Zealand Journal of Marine and Freshwater Research , 37 : 507 – 520 .
- Dewson , ZS , James , ABW and Death , RG . 2007 . A review of the consequences of decreased flow for instream habitat and macroinvertebrates . Journal of the North American Benthological Society , 26 : 401 – 415 .
- Duncan , MJ . 1995 . Hydrological impacts of converting pasture and gorse to pine plantation, and forest harvesting, Nelson, New Zealand . Journal of Hydrology (NZ) , 3 : 15 – 41 .
- Eggert , SL and Burton , TM . 1994 . A comparison of Acroneuria lycorias (Plecoptera) production and growth in northern Michigan hard and soft water streams . Freshwater Biology , 32 : 21 – 32 .
- Fahey BD , Rowe LK 1992 . Land-use impacts . In : Mosely M . Waters of New Zealand . New Zealand Hydrological Society , Wellington . Pp. 264 – 284 .
- Friberg , N , Winterbourn , MJ , Shearer , KA and Larsen , SE . 1997 . Benthic communities of forest streams in the South Island, New Zealand: effects of forest type and location . Archiv für Hydrobiologie , 138 : 289 – 306 .
- Fowler , RT . 2002 . Relative importance of surface and subsurface movement on benthic community recovery in the Makaretu River, North Island, New Zealand . New Zealand Journal of Marine and Freshwater Research , 36 : 459 – 469 .
- Fowler , RT . 2004 . The recovery of benthic invertebrate communities following dewatering in two braided rivers . Hydrobiologia , 523 : 17 – 28 .
- Gillespie PA , Forrest RW , Peake BM , Basher LR , Clement DM , Dunmore RA , Hicks DM in press . Spatial delineation of the depositional footprint of the Motueka River outwelling plume in Tasman Bay, New Zealand . Zealand Journal of Marine and Freshwater Research
- Gordon ND , McMahon TA , Finlayson BL 1992 . Stream Hydrology: an introduction for Ecologists . John Wiley & Sons Ltd , England .
- Gotelli NJ , Entsminger GL 2001 . EcoSim: Null models software for ecology. Version 7.0 . Acquired Intelligence Inc. & Kesey-Bear. http://garyentsminger.com/ecosim/index.htm (June 2010)
- Graynoth , E . 1979 . Effects of logging on stream environments and faunas in Nelson . New Zealand Journal of Marine and Freshwater Research , 13 : 79 – 109 .
- Hall , MJ , Closs , GP and Riley , RH . 2001 . Relationships between land use and stream invertebrate community structure in a South Island, New Zealand, coastal stream catchment . New Zealand Journal of Marine and Freshwater Research , 35 : 591 – 604 .
- Harding , JS and Winterbourn , MJ . 1995 . Effects of contrasting land use on physico-chemical conditions and benthic assemblages of streams in a Canterbury (South Island, New Zealand) river system . New Zealand Journal of Marine and Freshwater Research , 29 : 479 – 492 .
- Harding , JS , Winterbourn , MJ and McDiffett , WF . 1997 . Stream faunas and ecoregions in South Island, New Zealand: do they correspond? . Archiv für Hydrobiologia , 140 : 289 – 307 .
- Hickey , CW and Clements , WH . 1998 . Effects of heavy metals on benthic macroinvertebrate communities in New Zealand streams . Environmental Toxicology and Chemistry , 17 : 2338 – 2346 .
- Hickey , CW and Vickers , ML . 1992 . Comparison of the sensitivity to heavy metals and pentachlorophenol of the mayflies Deleatidium spp. and the cladoceran Daphnia magna . New Zealand Journal of Marine and Freshwater Research , 26 : 87 – 93 .
- Hicks DM , Griffiths GA 1992 . Sediment load . In : Mosely MP . Waters of New Zealand . Wellington , New Zealand Hydrological Society . Pp. 229 – 248 .
- Hilsenhoff , WL . 1987 . An improved biotic index of organic stream pollution . The Great Lakes Entomologist , 20 : 31 – 39 .
- Huryn , AD , Benke , AC and Ward , GM . 1995 . Direct and indirect effects of geology on the distribution, biomass and production of the freshwater snail Elimia . Journal of the North American Benthological Society , 14 : 519 – 534 .
- Huryn , AD , Butz , VM and Arbuckle , CJ . 2002 . Catchment land-use, macroinvertebrates and detritus processing in headwater streams: taxonomic richness versus function . Freshwater Biology , 47 : 401 – 415 .
- Iversen , TM , Wiberg-Larsen , P , Hansen , SB and Hansen , FS . 1978 . The effect of partial and total drought on the macroinvertebrate communities of three small Danish streams . Hydrobiologia , 60 : 235 – 242 .
- Jaarsma , NG , de Boer , SM , Townsend , CR , Thompson , RM and Edwards , ED . 1998 . Characterising food-webs in two New Zealand streams . New Zealand Journal of Marine and Freshwater Research , 32 : 271 – 286 .
- Jacobsen , D , Schultz , R and Encalada , A. 1997 . Structure and diversity of stream invertebrate assemblages: the influence of temperature with altitude and latitude . Freshwater Biology , 38 : 247 – 261 .
- Lake , PS . 2003 . Ecological effects of perturbation by drought in flowing waters . Freshwater Biology , 48 : 1161 – 1172 .
- Leopold LB 1994 . A view of the river . Harvard University Press , London .
- Lester , PJ , Mitchell , SF and Scott , D . 1994 . Effects of riparian willow trees (Salix fragilis) on macroinvertebrate densities in two small central Otago, New Zealand streams . New Zealand Journal of Marine and Freshwater Research , 26 : 267 – 276 .
- Maridet , L , Wasson , JG , Philippe , M , Amoros , C and Naiman , RJ . 1998 . Trophic structure of three streams with contrasting riparian vegetation and geomorphology . Archiv für Hydrobiologia , 144 : 61 – 85 .
- Monaghan , KA , Peck , MR , Brewin , PA , Masiero , M , Zarate , E , Turcotte , P and Ormerod , SJ . 2000 . Macroinvertebrate distribution in Ecuadorian hill streams: the effects of altitude and land use . Archives für Hydrobiologie , 149 : 421 – 440 .
- Mosley MP , Pearson CP 1997 . Introduction: hydrological extremes and climate in New Zealand . In : Mosley MP , Pearson CP . Floods and Droughts: the New Zealand experience . New Zealand Hydrological Society , Wellington . Pp. 1 – 14 .
- Newsome PJF 1995 . Directory of geographic databases . Manaaki Whenua–Landcare Research , Palmerston North .
- Pearce , AJ and Rowe , LK . 1979 . Forest management effects on interception, evaporation, and water yield . Journal of Hydrology (NZ) , 18 : 73 – 87 .
- Quinn JM 2000 . Effects of pastoral development . In : Collier KJ , Winterbourn MJ . New Zealand stream invertebrates: ecology and implications for management . Christchurch , Caxton Press . Pp. 208 – 229 .
- Quinn , JM , Cooper , AB , Davies-Colley , RJ , Rutherford , JC and Williamson , RB . 1997 . Land use effects on habitat, water quality, periphyton, and benthic invertebrates in Waikato, New Zealand, hill-country streams . New Zealand Journal of Marine and Freshwater Research , 31 : 579 – 597 .
- Quinn , JM and Hickey , CW . 1990a . Magnitude of effects of substrate particle size, recent flooding, and catchment development on benthic invertebrates in 88 New Zealand rivers . New Zealand Journal of Marine and Freshwater Research , 24 : 411 – 427 .
- Quinn , JM and Hickey , CW . 1990b . Characterisation and classification of benthic invertebrate communities in 88 New Zealand rivers in relation to environmental factors . New Zealand Journal of Marine and Freshwater Research , 24 : 387 – 409 .
- Reid , DJ , Quinn , JM and Wright-Stow , AE . 2010 . Response of stream macroinvertebrate communities to progressive forest harvesting: influences of harvesting intensity, stream size and riparian buffers . Forest Ecology and Management , 260 : 1804 – 1815 .
- Richards , C , Haro , RJ , Johnson , LB and Host , GE . 1997 . Catchment and reach-scale properties as indicators of macroinvertebrate species traits . Freshwater Biology , 37 : 219 – 230 .
- Robinson , BH , Brooks , RR , Kirkman , JH , Gregg , PEH and Gremigni , P . 1996 . Plant-available elements in soils and their influence on the vegetation over ultramafic (‘serpentine’) rocks in New Zealand . Journal of the Royal Society of New Zealand , 26 : 457 – 468 .
- Ryder , GI and Scott , D . 1988 . The applicability of the river continuum concept to New Zealand streams . Verhandlungen der Internationale Vereinigung für theoretische und angewandte Limnologie , 23 : 1441 – 1445 .
- Sagar , PM . 1983 . Invertebrate recolonisation of previously dry channels in the Rakaia River . New Zealand Journal of Marine and Freshwater Research , 17 : 377 – 386 .
- Scott , D , White , JW , Rhodes , DS and Koomen , A. 1994 . Invertebrate fauna of three streams in relation to land use in Southland, New Zealand . New Zealand Journal of Marine and Freshwater Research , 28 : 277 – 290 .
- Snelder , TH and Biggs , BJF . 2002 . Multiscale river environment classification for water resources management . Journal of the American Water Resources Association , 28 : 1225 – 1239 .
- Snelder , TH , Cattanéo , F , Suren , AM and Biggs , BJF . 2004 . Is the River Environment Classification an improved landscape-scale classification of rivers? . Journal of the North American Benthological Society , 23 : 580 – 598 .
- Stark JD 1985 . A macroinvertebrate community index of water quality for stony streams . Water & Soil Miscellaneous Publication 8 .
- Stark , JD . 1993 . Performance of the Macroinvertebrate Community Index: effects of sampling method, sample replication, water depth, current velocity and substratum on index values . New Zealand Journal of Marine and Freshwater Research , 27 : 463 – 478 .
- Stark , JD . 1998 . SQMCI: a biotic index for freshwater macroinvertebrate coded-abundance data . New Zealand Journal of Marine and Freshwater Research , 32 : 55 – 66 .
- Storey , RG and Quinn , JM . 2008 . Composition and temporal changes in macroinvertebrate communities of intermittent streams in Hawke's Bay, New Zealand . New Zealand Journal of Marine and Freshwater Research , 42 : 109 – 125 .
- Tabacchi , E , Correll , DL , Hauer , R , Pinay , G , Planty-Tabacchi , AM and Wissmar , RC . 1998 . Development, maintenance and role of riparian vegetation in the river landscape . Freshwater Biology , 40 : 497 – 516 .
- Townsend , CR , Arbuckle , CJ , Crowl , TA and Scarsbrook , MR . 1997 . The relationship between land use and physicochemistry, food resources and macroinvertebrate communities in tributaries of the Taieri River, New Zealand: a hierarchically scaled approach . Freshwater Biology , 37 : 177 – 191 .
- Townsend , CR , Dolédec , S , Norris , R , Peacock , K and Arbuckle , CR . 2003 . The influence of scale and geography on relationships between stream community composition and landscape variables: description and prediction . Freshwater Biology , 48 : 768 – 785 .
- Williams , DD and Hynes , HBN . 1976 . The ecology of temporary streams I. The faunas of two Canadian streams . Internationale Revue gesamten Hydrobiologie , 61 : 761 – 787 .
- Winterbourn , MJ , Cowie , B and Rounick , JS . 1984 . Food resources and ingestion patterns of insects along a West Coast, South Island, river system . New Zealand Journal of Marine and Freshwater Research , 27 : 197 – 215 .
- Winterbourn , MJ , Rounick , JS and Cowie , B . 1981 . Are New Zealand ecosystems really different? . New Zealand Journal of Marine and Freshwater Research , 15 : 321 – 328 .