Abstract
The overlap and conflict between marine mammals and fisheries is widespread and increasing globally. This study used satellite telemetry to examine the foraging patterns of lactating female New Zealand (NZ) sea lions Phocarctos hookeri from the Auckland Islands (50°50′S, 166°28′E), over two consecutive austral winters, 1996 and 1997. Overall, all individuals tagged were characterised as benthic foragers and were found to be diving beyond their calculated aerobic dive limits on 65% of all dives. They exhibited preferential use of the continental shelf and its edge, as they do in summer (59% overlap between winter and summer foraging areas). This study confirms that NZ sea lion foraging behaviours and areas are similar between seasons, and within and between years. Female winter foraging areas spatially overlapped by 38% with the operation areas of sub-Antarctic trawl fisheries. These findings are important because: female NZ sea lions show extreme foraging behaviour year round; and overlap with trawl fisheries is likely to occur throughout the entire lactation period of NZ sea lions. These results need to be accounted for in the management of this nationally critical, declining species.
Introduction
The New Zealand (NZ) sea lion (Phocarctos hookeri) is one of the world's rarest otariids (eared seals), with a pup production in significant decline. It is classified as Nationally Critical under the New Zealand Threat Classification System (Baker et al. Citation2010) and Vulnerable in decline by the International Union for Conservation of Nature (IUCN Citation2008). New Zealand sea lions breed predominately (99.9%) on the Auckland and Campbell/Motu Ihupuku islands in the New Zealand sub-Antarctic between latitudes 48°S and 53°S (Gales & Mattlin Citation1997; Chilvers et al. Citation2007). Pup production of NZ sea lions at the Auckland Islands (73% of the species; Maloney et al. Citation2012) has declined by more than 40% since 1998. Fisheries interactions both direct (by-catch) and indirect (resource competition) are considered the most plausible hypotheses for this decline (Robertson & Chilvers Citation2011; Chilvers Citation2012). There is extensive spatial and temporal overlap of the foraging area of lactating females from all Auckland Island breeding areas with fisheries operations during austral summers (Chilvers et al. Citation2005, Citation2011; Chilvers Citation2009).
Lactating female NZ sea lions have been satellite-tracked at the Auckland Islands during summer since 1994 (Gales & Mattlin Citation1997; Chilvers et al. Citation2011). These results have demonstrated that females forage over the entire area of the Auckland Islands shelf, predominantly foraging on the shelf and over the shelf edges. They have the longest foraging trips of any sea lion (Chilvers et al. Citation2005) despite being constrained as central place foragers with a dependent pup. Female NZ sea lions are the deepest and longest diving otariid and dive almost continuously while at sea with an extraordinarily large proportion of dives (e.g. on average 68%) exceeding their calculated aerobic dive limits (cADL; Gales & Mattlin Citation1997; Costa & Gales Citation2000; Chilvers et al. Citation2006). These factors combined show that lactating females are operating at the extreme of their physiological limits during early lactation (austral summers) and that they have done so consistently for more than a decade. This foraging behaviour makes them highly susceptible to external impacts, such as fisheries impacts, and any local environmental changes (i.e. possible effects from the Southern Ocean Oscillation) because they have limited ability to adapt their foraging behaviour to compensate (Gales & Mattlin Citation1997; Costa & Gales Citation2000; Chilvers et al. Citation2006).
In contrast to summer foraging, nothing has been published on the winter foraging behaviour of females, although data were collected during the winters of 1996 and 1997.
The aim of the present research was to use the winter foraging data from 1996 and 1997 to: 1) describe the foraging behaviour and location of the areas where lactating females from Enderby Island forage during the austral winter (July–Sept); 2) to determine the overlap with fisheries operations during the winter 1996–97 seasons; and 3) to investigate whether the high degree of site fidelity to foraging locations observed within and between years during summer (Chilvers Citation2008) also occurs during winter. This third aim will provide valuable information for species management by characterising the likely overlap between breeding females and fisheries operations during the autumn period when NZ sea lions moult and instruments cannot be attached.
Materials and methods
Capture and deployment
The research was conducted during the austral winter/spring (July–Sept) of 1996 and 1997. Study animals were selected on Enderby Island (50°30′S, 166°17′E; ), Auckland Islands, New Zealand sub-Antarctic. Enderby Island is the second largest breeding site of NZ sea lions (300–400 pups/year; Robertson & Chilvers Citation2011). All captured individuals were females observed to be nursing healthy pups. Captures were made using a specially designed hoop net (Furhman Diversified, Texas, USA). Netted animals were physically restrained by two people and anaesthetised using an isoflourane (2%–5%) oxygen mix delivered by a mask from a portable vaporising system (Gales & Mattlin Citation1997).
Figure 1 Auckland Islands showing the main breeding islands for NZ sea lions: Enderby, Dundas and Figure of Eight. Inset: New Zealand's sub-Antarctic area.
Once anaesthetised, the seals were weighed (200 kg capacity scale±0.5 kg) and measured (length, axillary girth) before instruments were attached. Prior to deployment, each instrument was glued or strapped to a piece of neoprene material cut to the same size as the unit's base. This neoprene base was then glued to the dorsal pelage just below the shoulder blades of the sea lion using two-part epoxy glue. Once the instruments were adequately attached to the sea lion (e.g. 8–10 min after glue application), the flow of anaesthetic was stopped and the animal allowed to recover and return to its pup. Each animal was observed after restraint until they were fully conscious and had returned to the group or location where captured.
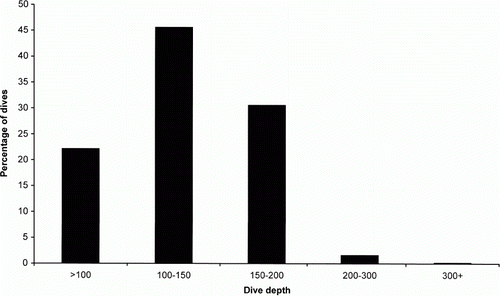
Figure 2 Frequency distribution of mean percentage of dives in each depth category for eight female NZ sea lions, winter foraging.
The instruments attached to the animals consisted of a micro-processor data logger, programmed to record wet/dry state, time and depth recorder (TDR), a satellite-linked platform transmitting terminals (PTTS) (Telonics 300 mW ST6, potted in epoxy, 130 mm×35 mm×15 mm, 175 g, Telonics, Mesa, Arizona, USA) and a VHF radio transmitter. A variety of TDRs were used: MK3e; MK5; and MK6 (Wildlife Computers, Redmond, Washington, USA). The MK3e TDRs (149 mm long by 26.5 mm diameter titanium tube weighing 136 g) are able to record depths of up to 1000 m. The MK5 TDRs are potted in a single epoxy block 65 mm×38 mm×13 mm×50 g (length×width×height×weight) and can record depth to 500 m. The MK6 TDRs are encased in an ovoid plastic housing 69 mm×57 mm×35 mm×80 g and can measure depth to 500 m. The TDRs were programmed to record depth every 5 s when wet (i.e. at sea) and to record the number and time of ‘dry’ records (i.e. onshore).
Analysis of dive data
Dive analysis programs from Wildlife Computers (e.g. zero offset correction, dive analysis and strip chart) were used to analyse the records from each TDR. Dives of less than 6 m were considered to be non-foraging dives primarily associated with travel and were not analysed, similar to previous NZ sea lion dive analysis (Gales & Mattlin Citation1997; Chilvers et al. Citation2006). As the accuracy resolution of the TDRs in the 0–500 m depth range was 2 m, by removing dives < 6 m, this eliminated any potential resolution errors around shallow dives. Bottom time was defined as the time the individual spent at depths exceeding 85% of the maximum depth for that dive. Data were analysed by individual animal. Times at which the animal left and entered the water were recorded on the TDR and were used to quantify the foraging cycle durations. The theoretical or calculated ADL for each female within this study was estimated based on a usable oxygen store of 47.4 ml O2 kg−1 (Costa et al. Citation1998) and assuming a diving metabolic rate of 58.7 ml O2 kg−0.75 min−1 (Costa & Gales Citation2000). All results are presented as means±1 SED. Statistical analysis comparing winter foraging parameters with previously collected summer foraging parameters was conducted using SPSS (2004, SPSS Inc.) using Pearson correlation analysis and ANOVAs.
Analysis of spatial data
Spatial data were analysed using the same methods as female NZ sea lion foraging data from Enderby during summer for comparability (Chilvers et al. Citation2005). Sea lion locations were calculated by reference to three satellites and assigned by Argos to six classes on the basis of their accuracy (i.e. most accurate to least accurate: 3, 2, 1, A, B, Z). All location categories (except Z which were excluded from analysis) were filtered using a maximum swimming speed parameter of 2.5 m s−1 (Crocker et al. Citation2001) and locations with a running mean speed greater than this value were excluded as outliers. All locations on land were also excluded. Although more accurate filtering methods are available, these methods were repeated here for comparability with past research. The accuracy of locations provided by Argos is classified as follows: class 3 accurate to 150 m; class 2 accurate to 350 m; class 1 accurate to 1 km; class 0 accurate to ≥ 1 km; and classes A and B have limited accuracy assigned.
Filtered locations were used to estimate the distance from Enderby Island and the total distance travelled. Total distance travelled resulted from the interpolation of all filtered locations. Due to the errors related to the accuracy of satellite locations, all distance calculations are presented to the nearest km. Calculations of mean distance travelled per trip, maximum distance from Enderby Island and Kernel ranges (Worton Citation1989) were restricted to complete trips (i.e. those defined by locations at the colony or within 10 km from the colony while the animals were travelling towards it). Locations from incomplete trips are represented in figures and tables but were not used in trip statistics. Kernel ranges for 50% and 65% of all locations per animal were created using the Animal Movement Extension of ARCVIEW (Hooge et al. Citation2000). Kernel ranges representing 50% and 65% of all locations, are useful representative ranges of foraging locations and aid in the description of the concentrated foraging area (Chilvers et al. Citation2005). These figures were calculated using smoothing factors calculated via least-square cross-validation (Seaman & Powell Citation1996).
All winter trip variables derived from location data were analysed and compared with previously collected summer foraging parameters using Microsoft Excel (2003, Microsoft, Washington, USA), SPSS (2004, SPSS Inc. Chicago, USA) and ARCVIEW (1998, ESRI Inc., California, USA). Since NZ sea lions dive almost continuously while at sea (Gales & Mattlin Citation1997), all trips and satellite locations are assumed to be part of a foraging trip and to represent foraging locations. The trawl fisheries operational location data for 1996 and 1997 were supplied by the Research Data Management section of the Ministry of Fisheries, New Zealand. Fisheries data represent all start/stop locations for arrow squid (Nototodarus sloanii; January to April 1996 and 1997, 8175 tows), scampi (Metanephrops challengeri; year round 1996–97, 2545 tows) and all other trawl fisheries (which predominantly include hoki [Macruronus novaezelandiae], orange roughy [Hoplostethus atlanticus] and ling [Geypterus blacodes], year round 1996 and 1997, 1671 tows) undertaken in the area of the Auckland Island shelf. Kernel ranges representing 50% and 95% of all trawler activity were created to show areas of highest activity. These figures were calculated using the same methods as described above for the sea lion foraging areas.
Results
Analysis of dive data
Dive data were collected from eight individuals over the two austral winters between July and September in 1996 and 1997. and show a summary of dive data from lactating NZ sea lions from the winter of 1996–97 and summer of 1995, 1997 and 2003–04 (Gales & Mattlin Citation1997, Costa & Gales Citation2000, Chilvers et al. Citation2006, Chilvers & Wilkinson Citation2009). Over 43,000 dives,>6 m were recorded. Dive depth, duration and percentage of time spent in at the deepest 85% depth of the dive were all significantly positively correlated. Deeper dives had longer dive durations and longer bottom time (Pearson correlation = 0.831, P<0.001; Pearson correlation = 0.635, P<0.001 respectively). shows the dive depth distribution across all dives for all individuals, with 97% of all dives being less than 200 m and the majority (46%) being between 100 to 150 m. Overall, these data, along with the satellite locations showing over 90% of locations are on the Auckland Island shelf in water less than 250 m in depth (A), indicate that all of the females from this research were benthic foraging individuals (consistently diving to the benthos as described in Chilvers & Wilkinson Citation2009). Overall, means from this dataset were compared with means from summer foraging data from 1995, 1997 and 2003–04 and the means of summer foraging data (split into individuals with benthic and mesopelagic foraging strategies) are given at the bottom of the tables (comparison data from Gales & Mattlin Citation1997; Costa & Gales Citation2000; Chilvers et al. Citation2006; Chilvers & Wilkinson Citation2009; Chilvers unpublished data, respectively). The only significant differences in diving behaviour were that the average winter dive duration and percentage bottom time was significantly longer than any summer diving behaviour (ANOVA P<0.001, F=8.2 and P<0.001, F=9.6 respectively; ). Similarly, average time spent ashore and at sea were significantly longer during winter than during summer foraging in either 1995 or 2003–04 (ANOVA P<0.001, F=7.0, P=0.003, F=3.9; )
Figure 3 Kernel range of all filtered satellite locations for lactating NZ sea lions from: A, Enderby Island, July to September 1996–1997 (n=6) and B, Enderby Island, January and February 2001–2004 (n = 26). Intensity of shading of kernel ranges represents percentage of time spent in the area displayed as highest concentration of locations (darkest) to lowest concentration (lightest). Bathymetric contours are shown as thin black lines, with the Auckland Islands shelf represented by the 500 m bathymetric boundary. Arrow squid Nototodarus sloanii trawl fishery effort (50% and 95% kernel ranges) from: A, January to April 1996–1997 and B, February to April 2001–2007 are represented by thick black solid lines. Scampi Metanephrops challengeri trawl fishery effort (50% and 95% kernel ranges): A, all year 1996–1997 and B, all year 2001–2007 are represented by dashed black lines. A, all other trawl fisheries activities (50% and 95% kernel ranges), all year 1996–1997 are represented by dotted black lines.
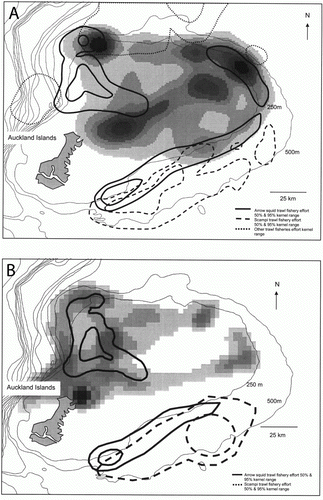
Table 1 Summary of statistics from the diving records of eight female NZ sea lions while at sea during the austral winters 1996 and 1997. Dives are defined as submersions > 6 m. All means presented±SE.
Table 2 Summary of foraging cycle and behaviour for eight female NZ sea lions tracked during winter 1996–97. Dives are defined as submersions > 6 m. Calculated aerobic dive limits (cADL) was estimated based on a useable oxygen store of 47.4 ml O2 kg−1 (Costa et al. Citation1998) and assuming a diving metabolic rate of 58.7 ml O2 kg−0.75 min−1 (Costa & Gales Citation2000).
Analysis of spatial data
Satellite tracking data were collected from six individuals between 39 to 58 days in the two winter seasons July–September, 1996 and 1997. shows a summary of foraging behaviour data from lactating NZ sea lions from the winter of 1996–97 and summer of 2003–04 (Chilvers et al. Citation2005; Chilvers & Wilkinson Citation2009). There is no summer 1995 or 1997 satellite data, only dive data as displayed in and . The percentage of each Argos location quality category after filtering for 1160 locations was: 1) classes 2 and 3 (11%); 2) class 1 (20%); 3) class 0 (34%); 4) A&B (35%). Filtering removed approximately 8% of all recorded locations across all location classes. The percentage of location classes are similar to the 2003–04 summer location classes which after filtering for 9200 locations was: 1) classes 2 and 3 (8%); 2) class 1 (24%); 3) ≤ class 0 (68%) (Chilvers et al. Citation2005) and are similarly proportioned to other Agros pinniped tracking studies analysed in a similar way (i.e. Beauplet et al. Citation2004; Robson et al. Citation2004). Overall, the six females winter foraging locations were similar distances from Enderby Island and were spread across the continental shelf and shelf edges in the area north and northeast of Enderby Island. Kernel range (KR) sizes showed variability between individuals but were not significantly different from each other, with overall mean areas of 491±61.2 km2 for 50% KR and 960±138.1 km2 for 65% KR (one-way ANOVA: unweighted F=0.87, d.f.=5, P=0.5; ). Chilvers et al. (Citation2005) showed there was no significant difference between KR and the number of complete foraging trips observed for each individual, indicating that KR is not a function of the number of foraging trips and that KR areas represent a clear fidelity to foraging areas. Comparisons of overall means from summer foraging data (n=26 animals from 2001 to 2004) and benthic and mesopelagic summer foraging data (n=10 and n=8 each respectively) are given at the bottom of the tables (i.e. comparison data from Chilvers et al. Citation2005; Chilvers & Wilkinson Citation2009; Chilvers unpublished data). The only significant differences between summer and winter foraging locations were that the 65% KR and mean distances between Enderby Island and the centre to the KR were both significantly larger in winter than summers (P=0.04, F=3.1, P=0.02, F=3.4 respectively; ). A shows the distribution of all foraging locations after filtering of individuals and the KR (50% and 95%) of fisheries activities data for the 1996 and 1997 years. B shows the comparable summer foraging location data for 26 lactating NZ sea lions from Enderby Island and the fisheries activity overlap for the years 2001 to 2007 (Chilvers Citation2009). Overall, there was a 59% overlap between filtered foraging locations between summer and winter locations. If only benthic summer foraging locations are used for comparison the overlap increases to 86%. Female winter foraging areas spatially overlapped by 38% with the operation areas of sub-Antarctic trawl fisheries.
Table 3 Deployment year, number, durations of foraging trips and kernel range (KR) sizes for six NZ sea lions satellite tracked from Enderby Island (EI), Auckland Islands during the austral winters of 1996 and 1997. All means presented±SE.
Discussion
The winter foraging and diving behaviour of female NZ sea lions has never previously been published and although the data for this research was collected in 1996 and 1997, winter foraging studies have not been repeated.
Perhaps the most striking feature of the winter data is that all females tracked were benthic foragers, as determined from their foraging behaviour (, ; , A; Chilvers & Wilkinson Citation2009). However, given the small sample size, it is not clear whether this result is due to a possible selectivity bias in our sampling (e.g. only benthic foragers were captured), changes in environmental variables (e.g. changes in prey distribution) or if, in fact, all females may change their foraging behaviour to exclusively benthic patterns during winter. Given that all summer foraging traits of individuals have been shown to be consistent within and across years (i.e. over a decade), despite presumed differences in environment and prey availability over this same period (Chilvers 2008), we believe that the most likely explanation is that the sampling of females was not representative with only benthic foraging females captured. This is consistent with a lack of difference in NZ sea lion diet between summer and winter (Childerhouse et al. Citation2001). Also, satellite studies undertaken at Enderby Island during summer have captured up to 12 lactating females at random and also had non-representative captures with, in one season, 11 out of 12 females captured being benthic foragers, while in other seasons the captures have been nine out of 12 mesopelagic females (Chilvers unpublished data). Given that all females from this research appeared to be benthic foragers, we compared their winter foraging behaviour with those females who had benthic foraging behaviour during summer (Tables ; Chilvers & Wilkinson Citation2009).
The summer foraging behaviour of female NZ sea lions is well described, showing they are the deepest, longest duration diving otariids, travelling the greatest distance from breeding areas with a significantly high proportion of their dives exceeding their cADL (Gales & Mattlin Citation1997; Costa & Gales Citation2000; Chilvers et al. Citation2006). It has been shown that during summer females show two distinct foraging styles, being either benthic (i.e. consistently diving to the bottom) or mesopelagic (i.e diving in deeper water depths with consecutive dives varying in depth by as much as 400 m, but off the bottom; Chilvers & Wilkinson Citation2009). All of these foraging traits have been shown to be consistent in individuals (Chilvers 2008) and all female NZ sea lions tracked from Enderby Island during summer also show extensive overlap both spatially and temporally with fisheries activities (Chilvers Citation2009).
Overall, the foraging patterns of benthic feeding females during winter and summer are similar. Fletcher (Citation2001), in a preliminary analysis of the at-sea distribution of NZ sea lions from 1996 to 1997 focusing on animals latitude and longitude locations, reported a significant difference in the mean foraging location of lactating females between summer and winter 1996–97. However, this analysis did not determine if the data analysed in summer came from mesopelagic or benthic foraging females, and this difference alone could result in the significant differences reported. Fletcher (Citation2001) did not report any significant difference between the 1996 and 1997 winter locations and the 1998 summer locations. There was also no evidence of differences in mean foraging location between year in either summer or winter (Fletcher Citation2001). Winter and summer benthic foragers utilised the entire north and northeast Auckland Island shelf area and have similar foraging and dive characteristics (i.e. distance from Enderby Island: 120 km vs. 122 km; mean dive depth: 118 m vs. 121 m; dive duration: 4.6 min vs. 4.2 min; , ). In addition, they both spent a high proportion of their time underwater while at sea (48% vs. 58%), spent a similar proportion of their foraging cycle at sea (57% vs. 64%) and both have a very high proportion of their dives exceeding cADL (65 vs. 82; ).
What differs between winter and summer is that females have longer foraging cycles in winter (132 h vs. 98 h), with both mean time ashore (73 h vs. 63 h) and mean time at sea (57 h vs. 38 h) being significantly longer than during summer. Additionally, foraging occurred over a larger foraging area than during any summer foraging trips recorded (65% KR: 960 km2 vs. 643 km2 or 622 km2 for benthic foragers) and a mean greater distance from Enderby Island (108 km vs. 64 km or 73.5 km for benthic foragers in summer; ). Longer winter foraging cycles are to be expected of lactating female pinnipeds because their pups in winter are 5–6 months older than pups in summer and are therefore more independent (e.g. can survive longer between feeds). This research indicates that these longer foraging cycles appear to lead to a less intense diving strategy over a larger foraging area and longer time. One potential explanation is that prey availability may decrease or become more dispersed during winter (e.g. spawning for arrow squid is only during January–May) which may result in this difference in benthic foraging behaviour. Studies of the seasonal diet of Auckland Island sea lions conducted contemporaneously with this study have shown they target a broad range of benthic and demersal prey with no obvious seasonal shift in prey types; however, this diet study did not distinguish between individuals with differing foraging strategies (Childerhouse et al. Citation2001). Similar increases in foraging ranges have also been reported in other otariids and have generally been attributed to factors relating to the cost of lactation and feeding dependent young (Merrick & Loughlin Citation1997; Boyd et al. Citation1998; Campagna et al. Citation2001).
Limitations
There are some obvious limitations to this research, the main one being a limited sample size and the time elapsed since data was collected. Undertaking winter research on NZ sea lions is a costly and logistically difficult exercise; consequently winter tagging has never been undertaken again since 1996–97. However, by comparing the summer diving behaviours of females across 1995, 1997, 2003 and 2004 (), it is clear that over that time period their summer diving behaviours have not changed significantly and, given we know that NZ sea lions summer foraging traits are consistent in individuals within and between years (Chilvers 2008), there is no reason to expect winter foraging behaviour to have changed over this time period either. However, it is a factor to be taken into consideration when interpreting these results.
Fisheries overlap
Data on sea lion diet (Childerhouse et al. Citation2001; Meyneir et al. Citation2010), the overlapping locations of summer sea lion foraging areas with fisheries operations (Chilvers et al. Citation2005; Chilvers Citation2009), and the fact that NZ sea lions are regular by-catch mortalities in fishing operations (Robertson & Chilvers Citation2011), all show that the trawl fisheries around the Auckland Islands have the potential to considerably impact NZ sea lions. This study shows that the winter foraging ranges of lactating NZ sea lions from Enderby Island is also likely to overlap with fisheries activities. This overlap indicates that both direct and indirect fisheries effects have the potential to impact on the population year round. Although fisheries intensity varies from year to year, the area where fisheries activities operate varies little across years (; Chilvers et al. Citation2005). There is currently a 12 nautical mile (22 km) marine reserve and marine mammal sanctuary surrounding the Auckland Islands group in which all fishing is banned. This area only contains a little over 30% of all winter foraging locations, and does not provide protection for the entire winter foraging area of any female tracked. These results are the same as is found in the summer foraging studies (Chilvers et al. Citation2005; Chilvers Citation2009; Chilvers et al. Citation2011).
Conclusion
This is the first published research on the winter foraging and dive behaviours of lactating NZ sea lions. The foraging patterns of benthic foragers during winter and summer only differ slightly, with females utilising similar foraging areas summer and winter and showing the same extreme foraging behaviour (i.e. 65% of all dives exceeding their cADLs). However, females do have longer foraging cycles, spend longer periods both ashore and at sea and cover larger foraging ranges during winter. The foraging locations of these female NZ sea lions show that there is likely to be overlap of winter foraging locations with fisheries activities, and this overlap occurs year round. This likely year round overlap, combined with the NZ sea lion winter and summer physiologically extreme foraging behaviour, needs to be considered in the management of this Nationally Critical and declining species as it strongly suggests they will be susceptible to this external impact. It is recommended that this research should be repeated to get a greater understanding of the differences between more recent summer and winter foraging patterns and current fisheries overlap.
Acknowledgements
Data presented in this paper were collected under funding from the New Zealand Department of Conservation (DOC). Approval for all work was obtained from the DOC Animal Ethics Committee (Approvals AEC52 1 Jun 2002). Thanks to I. Debski, S. Cooper, A. Todd and two anonymous reviewers all provided helpful, critical reviews of the manuscript.
References
- Baker , CS , Chilvers , BL , Constantine , R , DuFresne , S , Mattlin , R , van Helden , A and Hitchmough , R . 2010 . Conservation status of New Zealand marine mammals (suborders Cetacea and Pinnipedia), 2009 . New Zealand Journal of Marine and Freshwater Research , 44 : 101 – 115 . doi: 10.1080/00288330.2010.482970
- Beauplet , G , Dubroca1 , L , Guinet , C , Cherel1 , Y , Dabin , W , Gagne , C and Hindell , M . 2004 . Foraging ecology of subantarctic fur seals Arctocephalus tropicalis breeding on Amsterdam Island: seasonal changes in relation to maternal characteristics and pup growth . Marine Ecology Progress Series , 273 : 211 – 225 . doi: 10.3354/meps273211
- Boyd , IL , McCafferty , DJ , Reid , K , Taylor , R and Walker , TR . 1998 . Dispersal of male and female Antarctic fur seals (Arctocephalus gazella) . Canadian Journal of Fisheries & Aquatic Science , 55 : 845 – 852 . doi: 10.1139/f97-314
- Campagna , C , Werner , R , Karesh , W , Marin , MR , Koontz , F , Cook , R and Koontz , C . 2001 . Movements and location at sea of South American sea lions (Otaria flavescens) . Journal of Zoology , 257 : 205 – 220 . doi: 10.1017/S0952836901001285
- Childerhouse , S , Dix , B and Gales , NJ . 2001 . Diet of New Zealand sea lions (Phocarctos hookeri) at the Auckland Islands . Wildlife Research , 28 : 291 – 298 . doi: 10.1071/WR00063
- Chilvers , BL . 2008 . Foraging site fidelity of lactating New Zealand sea lions . Journal of Zoology , 276 : 28 – 36 . doi: 10.1111/j.1469-7998.2008.00463.x
- Chilvers , BL . 2009 . Foraging locations of a decreasing colony of New Zealand sea lions (Phocarctos hookeri) . New Zealand Journal of Ecology , 33 : 106 – 113 .
- Chilvers , BL . 2012 . Population viability analysis of New Zealand sea lions, Auckland Islands, New Zealand's sub-Antarctics: assessing relative impacts and uncertainty . Polar Biology , 35 : 1607 – 1615 . doi: 10.1007/s00300-011-1143-6
- Chilvers , BL , Amey , JM , Huckstadt , LA and Costa , DP . 2011 . Partitioning of breeding area foraging locations in New Zealand sea lions . Polar Biology , 34 : 565 – 574 . doi: 10.1007/s00300-010-0915-8
- Chilvers , BL and Wilkinson , IS . 2009 . Divers foraging strategies in lactating New Zealand sea lions . Marine Ecology Progress Series , 378 : 299 – 308 . doi: 10.3354/meps07846
- Chilvers , BL , Wilkinson , IS and Childerhouse , S . 2007 . New Zealand sea lion, Phocarctos hookeri, pup production: 1995 to 2005 . New Zealand Journal of Marine and Freshwater Research , 41 : 205 – 213 . doi: 10.1080/00288330709509909
- Chilvers , BL , Wilkinson , IS , Duignan , PJ and Gemmell , NJ . 2005 . Identifying the distribution of summer foraging areas for lactating New Zealand sea lions, Phocarctos hookeri . Marine Ecology Progress Series , 304 : 235 – 247 . doi: 10.3354/meps304235
- Chilvers , BL , Wilkinson , IS , Duignan , PJ and Gemmell , NJ . 2006 . Diving to extremes: are New Zealand sea lions (Phocarctos hookeri) pushing their limits in a marginal habitat? . Journal of Zoology , 269 : 233 – 241 .
- Costa , DP and Gales , NJ . 2000 . Foraging energetic and diving behaviour of lactating New Zealand sea lions, Phocarctos hookeri . Journal of Experimental Biology , 203 : 3655 – 3665 .
- Costa , DP , Gales , NJ and Crocker , DE . 1998 . Blood volume and diving ability of the New Zealand sea lions, Phocarctos hookeri . Physiological Zoology , 71 : 208 – 213 .
- Crocker , DE , Gales , NJ and Costa , DP . 2001 . Swimming speed and foraging strategies of New Zealand sea lions (Phocarctos hookeri) . Journal of Zoology , 254 : 267 – 277 . doi: 10.1017/S0952836901000784
- Fletcher D 2001 . Assessing seasonal differences in Hooker's sea lion locations . DOC Science Internal Series 33 . Wellington , , New Zealand , Department of Conservation . 9
- Gales , NJ and Mattlin , RH . 1997 . Summer diving behaviour of lactating New Zealand sea lions, Phocarctos hookeri . Canadian Journal of Zoology , 75 : 1695 – 1706 . doi: 10.1139/z97-796
- Hooge PN , Eichenlaub WM , Solomon EK 2000 . Using GIS to analyse animals' movements in the marine environment . Gustavus , AK , Alaska Biological Science Center. US Geological Survey .
- IUCN (International Union for Conservation of Nature) 2008 . 2008 IUCN Red List of Threatened Species 2008 . www.iucnredlist.org (accessed 28 January 2013) .
- Maloney , A , Chilvers , BL , Muller , CG and Haley , M . 2012 . Increasing pup production of New Zealand sea lions at Campbell Island/Motu Ihupuku: can it continue? . New Zealand Journal of Zoology , 39 : 19 – 29 . doi: 10.1080/03014223.2011.614265
- Merrick , RL and Loughlin , TR . 1997 . Foraging behaviour of adult female and young-of-the-year Stella sea lions in Alaskan waters . Canadian Journal of Zoology , 75 : 776 – 786 . doi: 10.1139/z97-099
- Meynier , L , Morel , PCH , Chilvers , BL , Mackenzie , DDS and Duignan , PJ . 2010 . Quantitative fatty acid signature analysis on New Zealand sea lions: model sensitivity and diet estimates . Journal of Mammalogy , 91 : 1484 – 1495 . doi: 10.1644/09-MAMM-A-299.1
- Robertson , BC and Chilvers , BL . 2011 . New Zealand sea lions Phocarctos hookeri possible causes of population decline . Mammal Review , 41 : 253 – 275 . doi: 10.1111/j.1365-2907.2011.00186.x
- Robson , BW , Goebel , ME , Baker , JD , Ream , RR , Loughlin , TR , Francis , RC , Antonelis , GA and Costa , DP . 2004 . Separation of foraging habitat among breeding sites of a colonial marine predator, the northern fur seal (Callorhinus ursinus) . Canadian Journal of Zoology , 82 : 20 – 29 . doi: 10.1139/z03-208
- Seaman , DE and Powell , RA . 1996 . An evaluation of the accuracy of kernel density estimators for home range analysis . Ecology , 77 : 2075 – 2085 . doi: 10.2307/2265701
- Worton , BJ . 1989 . Kernel methods for estimating the utilization distribution in home-range studies . Ecology , 70 : 164 – 168 . doi: 10.2307/1938423