
Abstract
Soil water storage in grassland is critical to regulate herbage yield while it may be threatened by continuous land use without plowing because of the progress of soil compaction associated with worsening soil hydraulic properties. This study aimed at contrasting the quantity and the availability of soil water in a meadow which had not been renovated for 13 years. We monitored matric potentials and mass soil water contents to 100 cm depth from autumn to winter in which plant transpiration was dormant. Soil water capacities were determined with soil water characteristics. The measurements were made in both a harvesting area in which agricultural vehicles had been operated, and a tree cover area which had experienced almost no vehicle loads. The soil layer in the tree cover area had a larger capacity for readily available moisture than that in the harvesting area. The matric potentials in the tree cover area varied in time between 0 and -1000 cm regardless of depth while those in the harvesting area were rather steady. These suggested better pore water continuity in the tree cover area. In the subsoil layers in both the harvesting and the tree cover areas, the soil water contents in terms of actually stored water did not reach as high a level as those expected from the soil moisture characteristics of the matric potential of -1000 cm. On the other hand, the measured matric potentials were consistently readily available for plants during the entire period of measurement. The apparent discrepancy between the matric potentials in readily available vs. actually stored water implied that the subsoil layers had become drier than observed during the study period, and that soil water hysteresis had prevented the full recovery of the water storage.
INTRODUCTION
Soil moisture is so essential a factor to regulate grassland yield that dry matter yield has been explained primarily by the amount of water transpired by plants. For instance, plant yield is often proportional to the amount of transpired water with a linearity coefficient called the water use efficiency (WUE). The typical WUEs for grassland species with evapotranspiration ranging from 200 to 800 mm are likely to be found around 20 to 25 kg of dry matter yield (DM) per hectare per millimeter of water (e.g. McKenzie et al. Citation1990; Kramer and Boyer Citation1995; Saeed and El-Nadi Citation1998) while a lower amount of evapotranspiration may cause the grassland species to have a WUE even smaller than 15 kg DM ha−1 mm H2O−1 (e.g. Liang et al. Citation1989; Martiniello Citation1999), suggesting the importance of maintaining soil moisture condition at an adequate level.
On the other hand, continuous land use can damage the soil moisture condition in grassland through soil compaction. A topsoil layer in grassland has a significantly lower chance of plowing across ordinary management schedules than in other types of agricultural land use which inevitably experience the operatation of agricultural vehicles for such practices as mowing, weeding or fertilizing. Even grazing animals can impose very localized but severe mechanical loads on the land surface. Soil compaction in a grassland can be represented by the increase in soil bulk density (Schwartz et al. Citation2003; Martinez and Zinck Citation2004; Bormann and Klaassen Citation2008), the lowering in infiltrability of rainfall (Alegre and Cassel Citation1996; Martinez and Zinck Citation2004) or the lowering in permeability of soil (Schwartz et al. Citation2003; Zimmermann et al. Citation2006; Bormann and Klaassen Citation2008). These unfavorable changes in soil physical conditions can result ultimately in a decline of herbage yield (Martinez and Zinck Citation2004).
Even Japanese grasslands, most of which can reap the benefits of plentiful annual precipitation, cannot avoid experiencing such severe seasonal or annual variations in precipitation that soil moisture deficits can threaten the stability of the harvest of forbs and grasses. In fact, many studies in Japan have dealt with the reduction in herbage yield due to a shortage of soil water (e.g. Hotta and Uchiyama Citation1964; Taniguchi et al. Citation1979; Miki et al. Citation1986; Fujii et al. Citation2011). More specifically, dry conditions seem to be harmful to the growth of legume species (e.g. Hashimoto et al. Citation1962; Iuchi Citation2008). Dry conditions can also promote the acidification of grassland soil, triggering a decline in herbage yields (Hojito and Nishimune Citation1990).
These types of decline in herbage yield can require grassland renovation with plowing. Ido et al. (Citation1978) gave questionnaires to 266 private and 116 public farms across all regions of Japan and clarified that 57% of the private and 36% of the public farms had to renovate their grasslands, for the farms had been troubled by a decline in grassland productivity. One can suggest that farms should renovate grassland within 10 years to maintain herbage yields since most of the grasslands in Ido et al. (Citation1978) had been used for less than 10 years after their establishment.
For stabilizing herbage yield, soil moisture conditions in the subsoil may have to be probed as well as those in the topsoil, since plants are likely to extend their roots into deeper layers to seek steadily available water when they encounter soil moisture deficits in a topsoil (e.g. Eissenstat Citation1991; Klepper Citation1991; Green and Clothier Citation1995). Herbaceous plants are also likely to evolve their root systems into subsoil layers (e.g. Garwood and Williams Citation1967; Gallardo et al. Citation1994; Huang et al. Citation1997; Grieu et al. Citation2001). So far, however, only a few analyses of soil moisture dynamics have taken into account the shift of plant-rooting depth in relation to the progress of drying in a topsoil (e.g. Lai and Katul Citation2000), mainly because of the shortage in number of observed facts to model the behaviors of soil water storage interacting with the growth of grassland species.
From the past studies mentioned above, it is hypothesized that continuous grassland use makes the water resource in subsoil vulnerable to depletion year by year because of the progress of the soil compaction associated with degrading the capacity and the conductivity of soil water. Thus, one should be concerned about not only a topsoil but a subsoil layer when one monitors the states and the quantity of soil water to estimate the sustainability in growing and harvesting grassland species.
The objective of this study was to contrast the quantity and the plant availability of soil water in a meadow which had not been renovated for 13 years. As a comparison, we observed the same sets of measurements both in a harvesting area in which agricultural vehicles had repeatedly been operated and in an adjacent tree cover area which had experienced almost no vehicle loads. To minimize the influences of root water uptake on the soil moisture condition, we monitored the transition of both the soil moisture condition and tree water uptake after a grassland cutting in autumn.
Table 1 Annual schedule of grassland practices in the study site. F: fertilization, M: mowing, W: weeding, H: herbicide application
Figure 1 Study site description. The three sampling locations were set at 1, 2 and 4 m away from the larch tree row to represent the tree cover area, the intermediate area and the harvesting area in this meadow site, respectively.
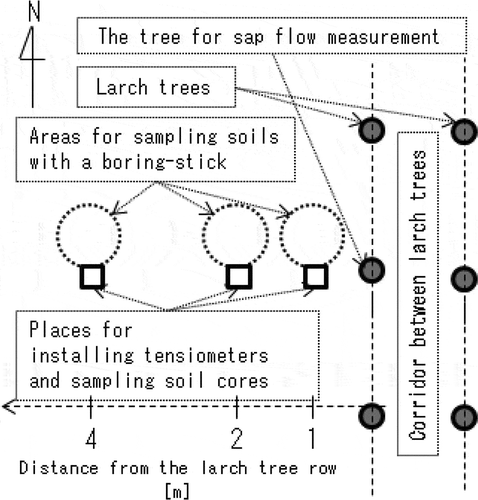
Figure 2 The depth profiles of (a–c) soil particle density, (d–f) soil bulk density and (g–i) soil hardness obtained at the tree cover area, the intermediate area and the harvesting area. Each plot indicates the mean value of triplicate (for the bulk densities) or five replicates (for the particle density and the soil hardness) samplings with the error bar denoting the variation between the minimum and the maximum.
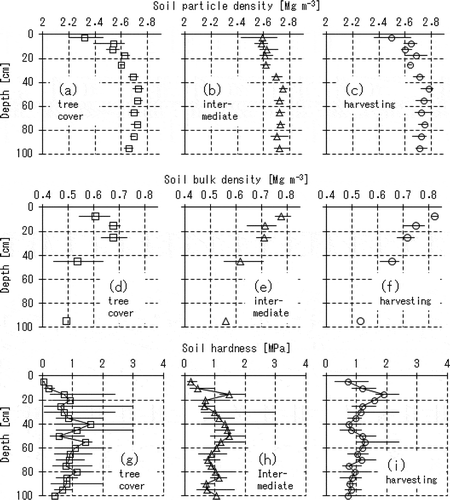
Figure 3 Comparisons of the soil water retention curves among the three distances from the tree row. The notation on each sub-graph indicate the sampling depth (cm), and the plots with squares (□), triangles (△) and circles (○) on each sub-graph denote the data sets obtained in the tree cover, the intermediate and the harvesting areas, respectively. Each plot on the sub-graphs indicates the mean value with the error bar denoting the variation between the minimum and the maximum among the triplicate measurements. At 95 cm depth in the three sampling locations, we obtained only one data set each.
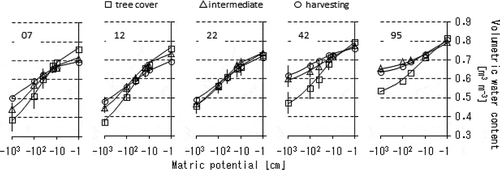
Figure 4 The time series data of the matric potentials of soil water at (a) the tree cover, (b) the intermediate and (c) the harvesting areas, (d) the daily precipitation and the atmospheric temperature, (e) the sap flow velocities in the outer-most 2 cm along the radius of the larch tree trunk, and (f–h) the mass-basis soil water content during the period of dormancy in tree transpiration at the places and the depths corresponding to the matric potential measurements.
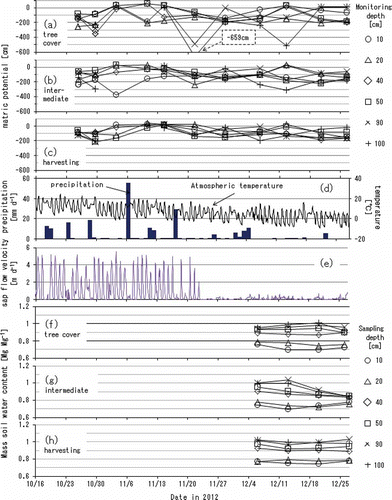
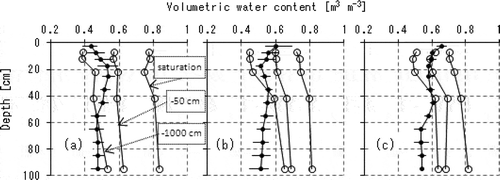
Figure 5 The depth profiles of volume fraction of soil water capacity (open circles) and of measured water content (black circles) for (a) the tree cover, (b) the intermediate and (c) the harvesting areas, respectively. The three curves with open circles on each of the sub-graphs denote the conditions in saturation (the right-most curve on each sub-graph), at field water capacity (the center, with the notation of -50 cm of matric potential) and at the lowest water content of readily available moisture (the left-most, with -1000 cm), respectively. The measured water contents with black circles were derived from the mass soil water contents in and the soil bulk densities in . The plots and the error bars on the measured water contents indicate the time-averaged values and the range between the minima and the maxima among the chances of measurements over the period of dormancy in tree transpiration.
MATERIALS AND METHODS
Study site descriptions
The study site (36°29' 23“N, 139°59'14“E) was set in a meadow of 1.4 ha (350 × 40 m), called “3-1A field”, in the Utsunomiya University Farm in Moka city, Tochigi, Japan. The annual means of temperature and precipitation in this city were 13.0°C and 1276 mm, respectively, from 1981 to 2010. This meadow had been managed without renovation since 1999, and adopted mixed intercropping with orchard grass (Dactylis glomereta L. cv. Natsumidori), perennial ryegrass (Lolium perenne L. cv. Friend), tall fescue (Festuca arundinacea Schreb. cv. Kentucky 31) and white clover (Trifolium repens L. cv. Huia).
shows the annual schedule of grassland practices. Vehicle loads on the study field resulted mainly from harvesting plants and fertilizing the top soil. The forbs and grasses had been harvested early in May and July, and late in October. Additional fertilizers had been applied in March [with nitrogen (N), phosphorus (P) and potassium (K) of 42, 54 and 48 kg ha−1], June (40, 0, 40 kg ha−1) and October (42, 54, 48 kg ha−1). Weeding in July also led to a chance for soil compaction. The loads from herbicide application came from human feet since the solution of 37% asulam diluted by a factor of at least 50 had been applied by hand on the leaves and stems of invasive plants.
Measurements
In order to quantify the effects of vehicle loads on soil physical conditions, we excavated three pits to 120 cm depth at 1, 2 and 4 m away from a larch tree row in October 2012 (). The larch trees had been planted for 30 years along a straight contour line extending 320 m in a north-south direction. And when an agricultural vehicle ran in parallel with the larch tree row, the vehicle wheels on the side of the trees would pass about 2 m away from the trees. Therefore, the area within a 2-m distance of the 320-m length of the tree row had received almost no vehicle loads. As we found when digging the pits, the roots of the larch trees extended to 1 or 2 m from their bodies but not to 4 m. Thus, the three sampling locations at 1, 2 and 4 m away from the larch tree row are called “the tree cover area”, “the intermediate area” and “the harvesting area”, respectively, in the later sections.
The soil profiles in the excavated pits consisted of an Andosol layer of 40 cm in thickness overlying a Kanto loam layer. The soil hardness was measured at every 5 cm in depth with five replicates each using a Yamanaka’s hardness meter (Yamanaka and Matsuo Citation1962; produced by Fujiwara Scientific Company Co. Ltd., Tokyo, Japan). The undisturbed soil cores of 50 cm3 were sampled from 7, 12, 22, 42 and 92 cm in depth with three replicates each, and were used to obtain soil water characteristics, soil bulk density and particle density. The soil particle density was evaluated based on the Japan industrial standards (JIS) A 1202 using Gay-Lussac-type pycnometers. The soil bulk density was measured by the gravimetric method in which the soil core samples of 50 cm3 were weighed before and after drying at 105–110°C for 24 h.
The volume fractions of readily available moisture in the soil layers were evaluated based on the soil water characteristics. The soil water retention curves were measured by the hanging water column method and the pressure plate method (Dane and Hopmans Citation2002). After saturating the undisturbed soil cores for 24 h in a laboratory at 25°C, we imposed suction on the soil water step by step from -5 to -100 cm using the hanging water column, and from -100 to -1000 cm using the pressure plate. At each pressure equilibrium, the soil cores were weighed to identify the volumetric water content. The threshold matric potentials to define the volume fraction of the readily available moisture for plants were set at -50 cm as the field water capacity and -1000 cm as the depletion of soil water content for optimum plant growth.
For identifying the time period of dormancy in the transpiration of the larch trees, we monitored the sap flow in a larch tree which stood just beside the excavated pits () by applying the thermal dissipation method (Granier Citation1987; Lu et al. Citation2002). The thermocouples and the heater to monitor the sap flow were installed into the outer 2.0 cm of the tree trunk radius with 10-cm intervals in vertical direction at breast height.
The precipitation at the study site was recorded every hour using a rain gauge installed 200 m from the study site. The air temperature on the study site was also measured once an hour by a temperature logger (CO-U23-002; Climatec Inc.; Tokyo, Japan) inside a radiation shield (CO-RS1; Climatec Inc.; Tokyo, Japan).
For monitoring the plant availability of the soil water both in the topsoil and in the subsoil, we measured soil matric potentials in the soil layers once every week or two from October to December 2012. The tensiometers were installed at 10, 20, 40, 50, 90 and 100 cm depth at each pit. A tensiometer was made of a ceramic cup 60 mm in length and 18 mm in diameter attached to the tip of a transparent polyvinyl chloride (PVC) tube 18 mm in diameter, into which water was poured. The matric potential at each sampling point was evaluated by manually measuring the air pressure in the top part of the tensiometer tube with a digital manometer (PG-100-102VP; NIDEC COPAL ELECTRONICS CORP.; Tokyo, Japan), assuming that the soil matric potential is equilibrated with the air pressure in the top of the tube.
For estimating the actual quantity of soil water, we obtained the depth profiles of mass soil water content at the distances of the three pits from the larch tree rows (). The sampling depths were every 5 cm in the top 20 cm layer and every 10 cm from 20 to 100 cm in depth. The soil samples were taken using a boring stick, coincidently with the matric potential measurements during the period of dormancy in tree transpiration, and the mass soil water contents were evaluated through weighing the samples before and after drying for 24 h at 105–110°C.
RESULTS AND DISCUSSION
Depth profiles of particle density, bulk density and hardness of the soil
shows the depth profiles of soil particle density, soil bulk density and soil hardness at the three sampling locations. The depth profiles of the soil particle density (a–c) indicated that each profile could be separated into the topsoil, where the particle density was lower than 2.7 Mg m−3, and the subsoil, where the particle density stayed at more than 2.75 Mg m−3 with a boundary at around 40 cm in depth. This boundary depth agreed with the observed depth at which the Andosol and the Kanto loam layers were separated.
The values of both the bulk density and the soil hardness were highly variable, especially in the tree cover area (d and g), suggesting a spatial variation of the degree to which soil matrix had been loosened or compacted by the trees extending their roots. In general, tree standing seemingly causes the soil water holding capacity to enlarge and promotes the development of a macro-pore network, encouraging the infiltration of soil water. Some studies reported that tree standings in a grassland made soil water contents under a tree cover area higher than in the adjacent area (e.g. Joffre and Rambal Citation1988; Puerto and Rico Citation1989), supporting the idea that planting trees for such practical purposes as erosion control or windbreak can be an alternative for introducing precipitated water into deep soil layers in grasslands.
The profiles of soil hardness had peaks at a 15-cm depth of 1.47 and 1.89 MPa in the intermediate area (h) and in the harvesting area (i), respectively. These values were high enough to indicate the development of hardpan as the soil hardness of around 0.98 MPa can be defined in Japan as the threshold value for severely restricting plant root growth regardless of plant species or soil types (Miyoshi Citation1972).
Soil water characteristics
shows comparisons of the soil water retention curves among the three sampling locations. For every sampling depth, the tree cover area showed a higher saturated water content and a larger capacity of soil water equivalent to the range of matric potential above -1000 cm than the other two sampling locations, meaning that the soil layers in the tree cover area can contain a higher amount of readily available moisture.
It was also likely that the more heterogeneous pore structures had developed around the trees since almost every data plot for the tree cover area showed larger variation among the replicates of the measurement than those for the other distances.
The total porosity was 0.69 or more for every sample and it became larger for the deeper layers. However, the deeper layers in the intermediate and the harvesting areas had greater amounts of pores corresponding to the matric potential range of lower than -1000 cm, and, as a result, could contain a lower amount of readily available moisture. For example, the soil cores sampled from 95 cm in depth in the tree cover area had a saturated water content of 0.81 on average while their water contents could be 0.64 even at -1000 cm, meaning that readily available moisture can occupy only a volume fraction of 0.17.
Transition of soil matric potentials
includes the time series data of matric potentials of soil water at the three sampling locations (a–c). d and e show the daily precipitation with the atmospheric temperature (d) and the sap flow velocities in the outermost 2 cm along the radius of the larch tree trunk (e). f–h shows the mass-basis soil water contents during the period of dormancy in tree transpiration at the places and the depths corresponding to the matric potential measurements.
The sap flow rate (e) abruptly ceased on November 22. Thus, that date was likely to be the boundary between the transpiring period and the dormant period of the larch tree. The cumulative precipitation during the measurement period was 146 mm for the transpiring period and 41 mm for the dormant period. Daily mean atmospheric temperature had simply decreased from 17.1 to -1.5°C during the measurement period.
The matric potentials were consistently no less than -1000 cm in the three sampling locations, meaning that the soil water in the study site was in readily available for plants in terms of soil water pressure. The lowest case among the observations occurred in the tree cover area. For instance, on November 21, at the end of the transpiring period, the matric potential decreased to less than -450 cm at both 90-cm and 100-cm depths (a), probably because of the tree water uptake occurring very sporadically in both time and place.
Through the fluctuating behaviors, the soil matric potentials in the tree cover area got close to the saturated condition several times at all sampling depths but the 20-cm depth. These prompt behaviors with peaks and dips in matric potential suggested that the water-filled pores in the tree cover area could easily interconnect in the vertical direction across the entire soil profile when the soil layers were being wetted.
In the harvesting area, by contrast, every sampling depth rarely fell below -200 cm of matric potential during the entire period of measurement (c). The matric potentials observed in the intermediate area showed transitional features between those observed in the tree cover and the harvesting areas. These relatively calm features of the matric potentials were presumably due to the development of hardpan impeding the water exchange between the topsoil and the subsoil. The absence of, or lower intensity of, tree water uptake in the harvesting and the intermediate areas also explains the steady behaviors along the time course of the matric potentials.
Comparison of the matric potentials with the quantity of soil water
shows the depth profiles of volume fractions of soil water capacity compared with the measured soil water content for the three sampling locations. The soil water capacity in each sub-graph was divided by three threshold lines as follows: (i) the saturated water contents derived from the soil particle densities and the soil bulk densities (), (ii) the field water capacity with the matric potential of -50 cm found in and (iii) the lower limit of readily available moisture with the matric potential of -1000 cm found in , respectively. The measured water contents were calculated using the soil bulk densities () and the mass soil water contents (f–h). The plots and the error bars on the measured water contents indicate the time-averaged values and the range between the minima and the maxima among the chances of measurements over the period of dormancy in tree transpiration.
Both the total porosity and the volume fraction of readily available moisture were higher in the tree cover area than in the other two sampling locations (a–c), suggesting the positive effects of tree standings on the increase in total porosity of soil layers. The total porosities in the tree cover, the intermediate and the harvesting areas were calculated as 308, 302 and 293 mm in the top 40-cm depth and 800, 784 and 768 mm in the entire 100-cm depth, respectively. The amounts of readily available moisture that could be stored in the top 40 cm were also larger in the tree cover area (70 mm) than in the intermediate area (61 mm) or the harvesting area (47 mm), and those for the whole 100-cm layer were 154, 103, 91 mm, respectively. In addition, the volume fractions of easily drainable water equivalent to the matric potential of more than -50 cm were larger in the tree cover area. These features suggested that the continuous grassland use had discouraged the development of soil structure while the tree standings had enhanced it.
The measured soil water contents proved that the top 15-cm layer in the harvesting area had retained more water than it can contain in the condition of field water capacity (c), while the top soil in the tree cover area (a) was better drained than in the other two sampling locations. These relatively high water contents in the top layer in the harvesting area can also be explained by the development of the hardpan at 15-cm depth (i).
The matric potential values observed in the three sampling locations (a–c) indicated that the soil water in the subsoil layers below the 40-cm depth, particularly in the intermediate and the harvesting area (b and c), was readily available with the matric potentials of above -1000 cm in the entire period of measurement. The amount of water actually stored there, on the other hand, consistently remained lower than the volumes of soil water that should be expected with a matric potential of -1000 cm.
This apparent discrepancy between the observed matric potentials and the amounts of actually stored water meant that a portion of the subsoil water had been consumed in a severely dried condition, and that it was not recharged as much as expected from the matric potentials observed in this study. This contradiction may be explained if the soil water characteristics of the subsoil layers had strong moisture hysteresis.
About hysteretic behaviors of soil water retention curves, however, so far, only a small number of past studies are available, dealing mainly with coarse-textured soils (e.g. Miller et al. Citation2002; Chen et al. Citation2007; Muraleetharan et al. Citation2009), and few cases have measured volcanic ash soils like those found in the eastern part of the main Japanese island. Thus, further studies are required to observe the cycle of wetting and drying in subsoil layers in grasslands, which will allow us to estimate the long-term sustainability of soil water storage in grasslands.
CONCLUSIONS
We probed the plant availability and the quantity of soil water in a meadow which had not been renovated for 13 years. The depth profiles of the soil matric potential to 100-cm depth and the amounts of actually stored soil water were monitored from autumn to winter when plant transpiration was dormant. Soil water characteristics were used to classify the soil water capacities in terms of the availability of soil water for plants. The measurements were made in both the harvesting area which had experienced agricultural vehicle loads and the tree cover area on which almost no vehicle loads had been imposed.
The depth profiles of soil water capacities showed that the harvesting area could have a smaller capacity for readily available moisture than the tree cover area, while the tree cover area contained a larger amount of macro-pores, implying that the tree standings had maintained or developed the macro-pore structures that could have been lost under the continuous grassland use.
The top 20 cm of soil in the harvesting area had been poorly drained as its field water capacity was often fulfilled while the topsoil in the tree cover area was well drained. In the tree cover area, the matric potentials in the entire soil profile varied more rapidly than those in the harvesting area. This suggested that the pore water was able to connect in the vertical direction more easily in the tree cover area than in the harvesting area, and that the hardpan found in the harvesting area worsened the hydraulic conductivity of soil water through the soil profile.
In the subsoil layers, particularly in the intermediate and the harvesting areas, the actually stored soil water was never more than the possible amount of soil water corresponding to the matric potential of -1000 cm, while the matric potential measured at each sampling location was consistently higher than -1000 cm during the entire period of measurement. This meant that the soil water is readily available for plants in terms of matric potential of the soil water while its actual quantity may have deteriorated through the years of grassland management.
These contradictory features between the matric potentials and the quantity of soil water suggested that the subsoil layers had experienced even drier conditions than the conditions observed during the monitoring period, and that soil moisture hysteresis had restricted the recovery of the subsoil water storage, which can be a likely cause of the long-term degradation of water resources in the subsoil layer. Further studies are necessary to monitor the wetting-drying cycles of subsoil layers for estimating the long-term sustainability of water storage in grasslands.
ACKNOWLEDGMENTS
The authors thank Mr. N. Yamaguchi and Dr. M. Kashiwazaki at Utsunomiya University Farm who provided us with useful information about grassland management and the weather data in the study sites. The authors also thank Ms. H. Akutsu, Ms. C. Tsuji, Ms. C. Tsunekawa and Mr. N. Koike in the Faculty of Agriculture, Utsunomiya University, for their technical support to obtain the data in the field and the laboratory.
References
- Alegre JC, Cassel DK 1996: Dynamics of soil physical properties under alternative systems to slash-and-burn. Agric. Ecosyst. Environ., 58 (1), 39–48. doi:10.1016/0167-8809(95)00654-0
- Bormann H, Klaassen K 2008: Seasonal and land use dependent variability of soil hydraulic and soil hydrological properties of two Northern German soils. Geoderma, 145 (3–4), 295–302. doi:10.1016/j.geoderma.2008.03.017
- Chen LX, Miller GA, Kibbey TCG 2007: Rapid pseudo-static measurement of hysteretic capillary pressure-saturation relationships in unconsolidated porous media. Geotech Test J, 30 (6), 1–10.
- Dane JH, Hopmans JW 2002: Water retention and storage. In Methods of Soil Analysis: Part4, Physical Methods, SSSA Book Series 5, Eds. Dane JH, Topp GC, pp. 680–683 and 688–690. Soil Science Society of America, Inc, Madison, Wisconsin.
- Eissenstat DM 1991: On the relationship between specific root length and the rate of root proliferation: A field study using citrus rootstocks. New Phytol., 118, 63–68. doi:10.1111/j.1469-8137.1991.tb00565.x
- Fujii H, Yoshida M, Shingu Y, Nakamura N 2011: Effects of artificial irrigation simulating daily precipitation of drought year on the growth of Timothy (Phleum pratense L.) ageing plants at northern district of Hokkaido, Japan. J. Jpn. Soc. Grassland Sci., 57 (3), 142–150 (in Japanese with English summary).
- Gallardo M, Turner NC, Ludwig C 1994: Water relations, gas exchange and abscisic acid content of Lupinus cosentiniileaves in response to drying different proportions of the root system. J. Exp. Bot., 45, 909–918. doi:10.1093/jxb/45.7.909
- Garwood EA, Williams TB 1967: Growth, Water use and nutrient uptake from the subsoil by grass swards. J. Agric. Sci., 69 (1), 125–130. doi:10.1017/S002185960001652X
- Granier A 1987: Evaluation of transpiration in a Douglas fir stand by means of sap flow measurements. Tree Physiol., 3, 309–320. doi:10.1093/treephys/3.4.309
- Green SR, Clothier BE 1995: Root water uptake by kiwifruit vines following partial wetting of the root zone. Plant and Soil, 173, 317–328. doi:10.1007/BF00011470
- Grieu P, Lucero DW, Ardiani R, Ehleringer JR 2001: The mean depth of soil water uptake by two temperate grassland species over time subjected to mild soil water deficit and competitive association. Plant and Soil, 230 (2), 197–209. doi:10.1023/A:1010363532118
- Hashimoto T, Takabayashi M, Saito T 1962: The effects of soil moisture on the growth, yields and their seasonal changes of Ladino clover. J. Jpn. Soc. Grassland Sci., 8 (3), 221–225 (in Japanese with English summary).
- Hojito M, Nishimune A 1990: Effects of precipitation on grass growth decline caused by grassland acidification. Jpn. J. Soil Sci. Plant Nutr., 61, 48–53 (in Japanese with English summary).
- Hotta R, Uchiyama S 1964: On productivity of fodder crops in Upland field transposed from paddy Field. J. Jpn. Soc. Grassland Sci., 10, 89–94 (in Japanese with English summary).
- Huang B, Duncan RR, Carrow RN 1997: Drought-resistance mechanisms of seven warm-season turfgrasses under surface soil drying: II. Root Aspects. Crop Sci., 37, 1863–1869. doi:10.2135/cropsci1997.0011183X003700060033x
- Ido T, Abe T, Toyoda H 1978: A survey on conservation of sloping grassland. Bull. Natl. Grassland Res. Inst., 13, 130–136 (in Japanese with English summary).
- Iuchi H 2008: Over-seeding of legumes to the reed canarygras dominant meadow on mineral soil in Tenpoku region of Hokkaido. Bull. Hokkaido Pref. Agric. Exper. Stn., 92, 69–72 (in Japanese).
- Joffre R, Rambal S 1988: Soil water improvement by trees in the rangelands of southern Spain. Acta Oecol. (Oecologia Plantarum), 9, 405–422.
- Klepper B 1991: Crop root system response to irrigation. Irrig. Sci., 12, 105–108. doi:10.1007/BF00192280
- Kramer PJ, Boyer JS 1995: Water relations of plants and soils. Academic Press, San Diego, California, Inc. ISBN0-12-425060–2.
- Lai CT, Katul G 2000: The dynamic role of root-water uptake in coupling potential to actual transpiration. Adv. Water Resour., 23 (4), 427–439. doi:10.1016/S0309-1708(99)00023-8
- Liang YM, Hazlett DL, Lauenroth WK 1989: Biomass dynamics and water use efficiencies of five plant communities in the shortgrass steppe. Oecologia, 80, 148–153.
- Lu P, Woo KC, Liu ZT 2002: Estimation of whole-plant transpiration of bananas using sap flow measurements. J. Exp. Bot., 53, 1771–1779. doi:10.1093/jxb/erf019
- Martı́nez LJ, Zinck JA 2004: Temporal variation of soil compaction and deterioration of soil quality in pasture areas of Colombian Amazonia. Soil and Tillage Research, 75 (1), 3–18. doi:10.1016/j.still.2002.12.001
- Martiniello M. 1999: Effects of irrigation and harvest management on dry-matter yield and seed yield of annual clovers grown in pure stand and in mixtures with graminaceous species in a Mediterranean environment. Grass and Forage Sci., 54, 52–61. doi:10.1046/j.1365-2494.1999.00153.x
- McKenzie BA, Gyamtsho P, Lucas RJ 1990: Productivity and water use of lucerne and two lucerne-grass mixtures in Canterbury. Proc New Zealand Grassland Assoc, 52, 35–39.
- Miki N, Takao K, Nishimune A 1986: The effects of annual and seasonal change in precipitation and soil water retentivity on the grassland productivity in Tenpoku district. Bull. Hokkaido Pref. Agric. Exper. Stn., 54, 21–30 (in Japanese with English summary).
- Miller CJ, Yesiller N, Yaldo K, Merayyan S 2002: Impact of soil type and compaction conditions on soil water characteristic. J. Geotech. Geoenviron. Eng., 128 (9), 733–742. doi:10.1061/(ASCE)1090-0241(2002)128:9(733)
- Miyoshi H 1972: Effective layer for upland field from a view point of suitable soil condition for root development. Jpn. J. Soil Sci. Plant Nut., 43, 92–97 (in Japanese).
- Muraleetharan KK, Liu C, Wei C, Kibbey TCG, Chen L 2009: An elastoplatic framework for coupling hydraulic and mechanical behavior of unsaturated soils. Int. J. Plast., 25, 473–490. doi:10.1016/j.ijplas.2008.04.001
- Puerto A, Rico M 1989: Influence of tree canopy (Quercus rotundifolia Lam.) on content in surface soil water in Mediterranean grasslands. Ekologia (CSSR), 8, 225–238.
- Saeed IAM, El-Nadi AH 1998: Forage sorghum yield and water use efficiency under variable irrigation. Irrig. Sci., 18, 67–71. doi:10.1007/s002710050046
- Schwartz RC, Evett SR, Unger PW 2003: Soil hydraulic properties of cropland compared with reestablished and native grassland. Geoderma, 116 (1–2), 47–60. doi:10.1016/S0016-7061(03)00093-4
- Taniguchi K, Asahida Y, Hirose Y 1979: Grassland productivity assessed with the animal performance under cut forage feeding system. Res. Bull. Univ. Farm Hokkaido Univ., 21, 78–84 (in Japanese with English summary).
- Yamanaka K, Matsuo K 1962: Study about soil hardness –1. Soil hardness and soil water content -. Jpn. J. Soil. Sci. Plant Nutr., 33, 343–347 (in Japanese).
- Zimmermann B, Elsenbeer H, de Moraes JM 2006: The influence of land-use changes on soil hydraulic properties: Implications for runoff generation. For. Ecol. Manage., 222 (1–3), 29–38. doi:10.1016/j.foreco.2005.10.070