
ABSTRACT
This paper is a cross-comparative examination of how tropical forested islands were populated by humans. It first describes the unique ecological conditions of these environments, how they fluctuated during glacial cycles, and the challenges and affordances they provided people. The paper then explores the global archaeological record, classifying modes of colonisation that led insular tropical forests to be populated. These modes include terrestrial colonisation followed by insularisation (Mode A), maritime colonisation followed by major landmass reconfiguration (Mode B), maritime colonisation of uninhabited islands that always remained insular (Mode C), and maritime colonisation of already inhabited islands (Mode D). Finally, the paper discusses how, amongst Homo sapiens, ongoing dynamism between human adaptive behaviours and environmental flux stimulated processes of diversification, specialisation, and connectivity in these crucial ecologies; by contrast, archaic hominins like Homo erectus, Homo floresiensis, and Homo luzonensis may have found changes associated with forest expansion and insularity extremely challenging.
Introduction
Islands and tropical forests are two environments with the capacity to isolate and differentiate living organisms, including hominins. In biogeographic terms, insularity constrains the variety of plants and animals present but encourages the emergence of novel endemic species (Whittaker and Fernández-Palacios Citation2007). On the other hand, tropical forests produce high biomass and biodiversity but low densities of large fauna and carbohydrate-rich plant resources (Corlett and Primack Citation2011). The combination of these environments – island rainforests – should therefore exacerbate processes of isolation and diversification in peculiar ways. Despite this, there have been few attempts to bring these two environments together on a broad theoretical level to understand the implications for the people that interacted with island rainforests in the deep past. There is scope to explore how hominin adaptive flexibility over the longue durée became interlinked with these novel and perhaps challenging places to live, especially through periods of environmental fluctuation.
This paper expressly explores the peopling of rainforested islands at the global scale. It does so by first synthesising biogeographic and environmental studies to identify the distinctive ecological contingencies of these spaces. It then classifies the modes by which tropical forested islands were populated,Footnote1 identifying patterns in behavioural trajectories and geophysical processes. This classification structures a critical review of archaeological data. Here, the review is concerned exclusively with forested islands between the Tropic of Capricorn and the Tropic of Cancer. The focus is primarily on the islands of Southeast Asia, Near Oceania, and Sri Lanka where the highest density of tropical forests exists today, but the discussion at times extends to forested parts of Madagascar, the Caribbean, and Remote Oceania. It therefore omits other important archipelagos like the Mediterranean, and sub-tropical forested islands like Te Ika-a-Māui (New Zealand) and Kyushu (Japan); however, many of the macro-level inferences about adaptation and environmental flux, drawn together below, may extend to those settings too. Although by no means exhaustive, the synthesis draws out some key trends in how these important ecological spaces were populated and reshaped in the deep past.
The ecological contingencies of island rainforests
Island physiographic features have important implications for humans. Island size structures the diversity of environmental zones and directly corresponds to the terrestrial plant and animal species available for subsistence, material culture production, and exchange; large islands tend to support more diverse ecotones and biota than small ones (MacArthur and Wilson Citation1967; Whittaker, Triantis, and Ladle Citation2008). Geological substrates, interlinked with island size and environmental variability, also regulate land systems and soils that are important for cultivation. Isolation and remoteness further influence the terrestrial plant and animal species present because they shape colonisation and extinction processes; remote and isolated islands usually have depauperate biota (Heaney Citation2000; MacArthur and Wilson Citation1963). Moreover, distance from other landmasses influences the accessibility of islands by watercraft, with implications for when they were first populated, and how they were connected with established maritime networks (Leppard et al. Citationin press; Keegan and Diamond Citation1987). Water temperature, currents, and surface winds can further determine the difficulties of navigating around the coast and between islands (Anderson Citation2018; Irwin Citation1991). Warm tropical waters, in particular, are also associated with abundant pelagic, reef, and littoral resources, meaning that although some small and remote islands have depauperate terrestrial biota, they can be rich in marine opportunities (Szabó and Amesbury Citation2011).
Tropical rainforests present a similar range of challenges and affordances for humans. Canopy stratification allows numerous distinct ecotones and microclimates to exist within small horizontal areas, producing high biotic diversity (Allaby Citation2006, 172). The canopy tends to be replete with climbers, epiphytes, and bromeliads. The understory consists of smaller, younger trees, and shrub layers often include saplings and bushes. The forest floor is comparatively depauperate or inhabited by plants that thrive in low light conditions such as mosses, ferns, and some herbs (Erwin Citation2001). Although large-bodied animals are often present in extremely low numbers and edible plant resources are irregularly distributed under the canopy (Montagnini and Jordan Citation2005, 19), tropical forests can present humans with a wide variety of resources as they grade between evergreen rainforests, limestone and heath forests, peat and swamp forests, and mangroves. A host of physiological challenges were associated with these environments, however, including meeting energetic demands, thermoregulation, locomotion, and navigation (Roberts Citation2019; Roberts et al. Citation2016).
Rainforested islands are essentially forest fragments divided by water. One characteristic of forest fragments is an increased forest perimeter, which produces ‘edge effects’ (Laurance and Yensen Citation1991). As shown in , edge effects consist of a variety of biophysical changes unique to marginal areas such as thinner canopy cover and variable microclimates (Laurance Citation1991), which have important implications for human habitability (Gaffney Citation2021b). Around the sea, these effects are often related to increased wind velocity and damage to the forest edge and exposed areas of the canopy. The combination of tall canopies and shallow roots makes the rainforest highly susceptible to adverse winds, especially during tropical monsoons and cyclones (Corlett and Primack Citation2011, 14). This is compounded by the depletion of ground water and increased evapotranspiration near the edge of tropical forests (Bierregaard Jr et al. Citation1992). In the case of humans, however, edge effects often positively impact mobility and settlement, which are expedited by open spaces along the coast, nearby rivers, or in clearances within the forest (Roberts et al. Citation2016).
Figure 1. Edge effects at the margins of rainforest fragments, displaying the distance into the forest fragment each effect is felt (data from Laurence et al. Citation2002).
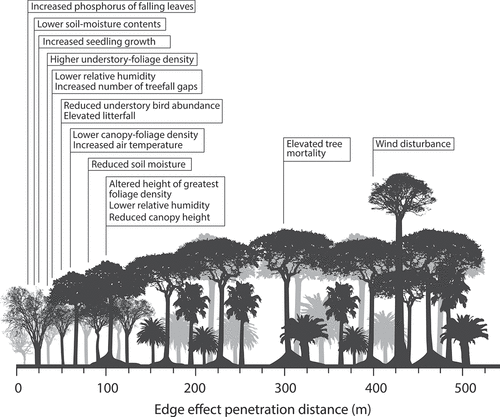
Forest fragments also affect the fauna and flora available to humans, with different species feeding on successional and secondary plants on the margins, and increased competition brought about by limited resource patches (Gibson et al. Citation2013). Fruit and nut bearing trees in the under storey of the forest grow more readily around forest edges (Mountain Citation1991a). In terms of fauna, forest fragments constrain the movement of most forest-adapted species (Malcolm Citation1988, Citation1991) and lead to compressed population sizes (Bierregaard Jr & Lovejoy Citation1989). Despite overall reductions to diversity and abundance on island rainforests, edge effects also encouraged specific animals to congregate at the margins; all of these factors exposed fauna to the impact of humans hunting (Cullen, Bodmer, and Valladares Pádua Citation2000).
Processional forcing and climatic cycling during the Quaternary made island rainforests dynamic, with fluctuations in sea-level, precipitation, and reef development drastically changing their extent and topography throughout archaeological time. As sea levels fluctuated, they connected and separated islands from continental shelves by revealing and then submerging land bridges (Fernández-Palacios et al. Citation2016). During interglacials, sea level rise submerged large tracts of continental shelf including forested regions. However, increases in atmospheric CO2, ambient temperatures, and precipitation simultaneously encouraged the expansion of lowland C3 plants, forming archipelagos of rainforested islands. In Asia, the Palk Strait infilled during the Holocene, cutting Sri Lanka off from India (Banerjee Citation2000). The Sunda shelf, forming an extension to mainland Southeast Asia, has been repeatedly submerged and exposed during marine transgressions. This turned Sunda into the islands of Borneo, Java, and Sumatra at the start of the Holocene, although the Philippines and Wallacea had always remained insular, separated from the mainland by deep-sea trenches (Sathiamurthy and Voris Citation2006). To the east, the islands of New Guinea, Aru, and parts of the Raja Ampat and Massim groups were attached to Australia during low-stands, comprising the continent of Sahul, with the Gulf of Carpentaria at times forming a large paleo-lake (Torgersen et al. Citation1983). Off Sahul’s northeast coast, the Bismarck Archipelago and Solomon Island chain were always separated from the mainland. Throughout human history, Madagascar has remained an isolated continental island, experiencing small fluctuations in size (Ali and Huber Citation2010). Similarly, except for Trinidad and Tobago, the Greater and Lesser Antilles of the Caribbean were never connected to the mainland, but many Pleistocene islands were submerged or reduced in size at the start of the Holocene (Cooke et al. Citation2017).
Patterns in the peopling of rainforested islands
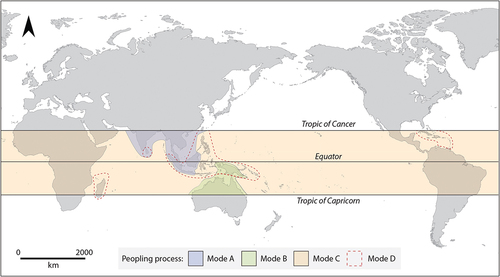
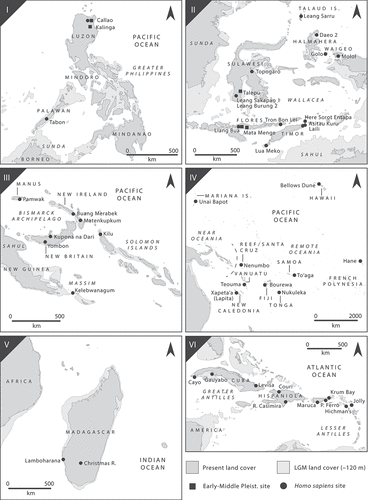
We can identify patterns in the coeval climatic processes and behavioural trajectories that led island rainforests to become populated. Here, I group these into four peopling processes or modes (). Mode A refers to a process of initial terrestrial occupation followed by insularisation, leading pieces of forested land and their human inhabitants to be cut off from the mainland. Mode B refers to large landmasses initially being peopled by water crossings, after which they were insularised. This process is behaviourally distinct from Mode A because it involved seafaring practices emerging prior to insularisation. Mode C refers to water crossings leading uninhabited forested islands to be populated, but despite sea level fluctuations the islands remained insular during the Pleistocene, or the islands were reached after Holocene marine transgression. Mode D refers to water crossings to already-inhabited marine islands, leading to overlapping and distinct cultural groups living on the same island. This classification scheme is now used to critically assess the global archaeological dataset.
Figure 2. The modes by which island rainforests were populated. Mode A: terrestrial colonisation followed by insularisation. Mode B: water crossings to large landmasses followed by large-scale insularisation. Mode C: water crossings to uninhabited marine islands that remained insular. Mode D: colonisations involving the settlement of marine islands already occupied by humans.
Mode A: insularisation
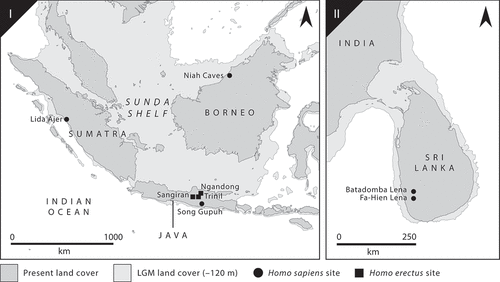
Homo erectus was present around Sunda from about 1.3 million to perhaps c.100,000 years ago (Matsu’ura et al. Citation2020; Rizal et al. Citation2020) (). During this time, the shelf experienced major fluctuations to its shape and size, at times turning what is now Java into a peninsula and, particularly in Marine Isotope Stage (MIS) 5 (130,000–80,000 years ago), an island (van den Bergh et al. Citation1996). Based on what we know about H. erectus mobility and niche selection, it is likely that these hominins moved into Java during successive sea level lowstands at c.1.8 million years ago and again c.1.2 million years ago (Sémah et al. Citation2010). During these times, mosaics of open tropical forests and savannah were dominant, lagoons formed around volcanic domes, and mangroves were present around coastlines (Bettis et al. Citation2009; Sémah and Sémah Citation2012). There is evidence that H. erectus foraged for shellfish (Joordens et al. Citation2009) and reused the shell remains for tools and mark making (Choi and Driwantoro Citation2007; Joordens et al. Citation2015); in these coastal and lacustrine areas H. erectus may have even experimented with rudimentary swimming and rafting (Gaffney Citation2021a). There is yet to be direct evidence for H. erectus living in the dense tropical rainforests that expanded during interstadials (Roberts Citation2019, 88), although forest margins could have plausibly been targeted (Sémah et al. Citation2016). It may not be a coincidence that the latest evidence for H. erectus on Java in MIS-5 corresponds with a period of rapid fluctuation, insularisation, and high precipitation (Cutler et al. Citation2003). Around Sunda, this period was associated with the increased temperature and humidity that prompted the expansion of tropical forests (Van Der Kaars and Dam Citation1997) and faunal turnover leading grassland and open woodland species to be supplanted by rainforest varieties (Storm et al. Citation2005). H. erectus may have been unwilling and unwitting participants in the final stages of these Mode A processes, encountering difficulties in navigation and subsistence maintenance under the canopy, and suffering gradual population bottlenecks having been cut off from mainland Southeast Asia.
Figure 3. Settings for Mode A peopling of rainforested islands (overland movement followed by insularisation), with key archaeological sites showing species presence. I: the Sunda Shelf in Southeast Asia; II: South Asia.
By contrast, the earliest evidence for Homo sapiens on Sunda at Lida Ajer dates to MIS-4, 73,000–63,000 years ago (Westaway et al. Citation2017). This was a period of lowered sea levels and, most parsimoniously, our species entered the region over land. However, following the LGM, the insularisation of formerly continental savannah and the expansion of lowland rainforests turned Sundalanders into rainforest-islanders. On large newly formed islands, humans took advantage of rainforest expansion rather than totally abandoning habitation sites. Sites like the Niah Caves demonstrate that some parts of northern Borneo were continually surrounded by patches of closed canopy rainforest in the Late Pleistocene (Stimpson Citation2012), and people hunted a variety of rainforest fauna throughout the period of insularisation, particularly including large ungulates like wild suids, as well as orangutan, gibbons, monkeys, fruit bats, forest birds, rodents, and sun bears (Arifin Citation2017; Piper and Rabett Citation2009). The encroachment of the sea to the north, however, did lead to the development of mangrove forests that encouraged increased foraging for large soft-shelled turtles (Barker et al. Citation2011; Pritchard, Rabett, and Piper Citation2009), while warming temperatures facilitated hunter-gatherers to frequent the dense rainforested interior of the island as well (Kusmartono, Hindarto, and Herwanto Citation2017). Similarly, when Sri Lanka separated from the subcontinent, sites like Fa-Hein Lena demonstrate that humans continued to hunt in the lowland rainforests from MIS-3 to MIS-1, where they specialised on small and hard-to-catch game like monkeys and squirrels (Roberts et al. Citation2015; Wedage et al. Citation2019). Composite bone tool production, present throughout the Late Pleistocene to Mid Holocene on both Borneo and Sri Lanka, suggest specialised rainforest hunting practices continued unbroken, and actually intensified, despite the insularisation of these large shelf islands (Barton et al. Citation2009; Perera, Roberts, and Petraglia Citation2016; Rabett and Piper Citation2012).
Mode B: overwater movement and landscape reconfiguration
H. sapiens moved from Sunda to Sahul via Wallacea (to which we will return) certainly before 50,000 years ago (O’Connell et al. Citation2018) and very possibly before 65,000 years ago (Clarkson et al. Citation2017) (). In doing so, watercraft were used to enable deliberate movements of people (Balme Citation2013), leading many hundreds of individuals to gradually arrive on the shores of what is now submerged shelf off the northwest coast of Australia and/or on the Bird’s Head of New Guinea (Bird et al. Citation2019; Bradshaw et al. Citation2019; Norman et al. Citation2018). These movements were not unidirectional and people would have returned to their home communities in Wallacea to tell stories of their discoveries and recruit individuals to establish viable founder populations. These factors suggest that the initial phase of frequentation was associated with strong kin and marriage ties between Wallacea and Sahul that operated for several centuries or a millennia before being replaced by networks operating primarily within Sahul.
Figure 4. Northern Sahul, the setting for Mode B peopling of rainforested islands (overwater movement followed by insularisation), with key archaeological sites.
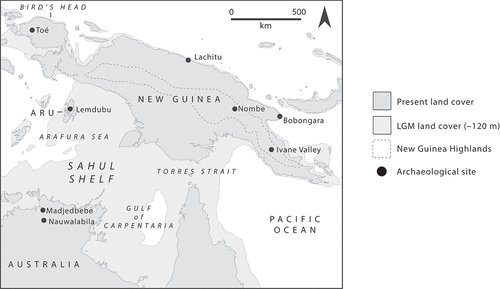
Once on Sahul, populations likely spread quickly and across large distances into a myriad different ecological zones (Bradshaw et al. Citation2021; Crabtree et al. Citation2021). The Madjedbebe humans had already moved hundreds of kilometres inland from the northwest Sahul coast by 65,000 years ago (Clarkson et al. Citation2017), and there is currently little evidence for humans using arid coastlines again until around 30,000 years ago (O’Connor and Veth Citation2008). By contrast, in the Ivane Valley of northeast Sahul, hunter-gatherers were moving between lowland and montane rainforests from sometime before 42,000 years ago (Summerhayes et al. Citation2010), and the lowland and hilly rainforests around coastlines, like at Bobongara and Lachitu, continued to be visited (Groube et al. Citation1986; O’Connor et al. Citation2011a). This initial phase of occupation was associated with complex plant processing in the montane and lowland rainforests of what is now New Guinea (Summerhayes et al. Citation2010), and the open tropical forests and savannah of what is today Australia (Florin et al. Citation2020, Citation2021). Plant foods were probably supplemented with medium to large bodied marsupial hunting, although the evidence is scarce (Field et al. Citation2008; Mountain Citation1991b).
Upon insularisation after the LGM, many areas of Sahul were slowly abandoned. The separation of the Aru Islands from northern Sahul was associated with changes from open grasslands to wetter forest conditions (Hope and Aplin Citation2005), leading to the abandonment of Liang Lemdubu, which had formerly been used as a base camp to hunt a variety of wallaby species (O’Connor et al. Citation2002). Similar trends can be seen around Australia, where the newly formed Barrow Island (Veth et al. Citation2017), High Cliffy Island (O’Connor Citation1994), Kangaroo Island (Lampert Citation1979), and the Furneaux Group (Sim Citation1998), came to be depopulated in the Early and Mid Holocene. By contrast, on the Torres Strait Islands (formerly hilltops surrounded by a large plain) archaeological evidence suggests people arrived in the Early Holocene following insularisation, and that they began to specialise in hunting large sea mammals (David et al. Citation2004; Wright Citation2011). Moreover, rainforested inland areas like the New Guinea Highlands and the Bird’s Head Peninsula continued to be frequented by foragers (Gaffney and Denham Citation2021). In some of these places, lowland–highland seasonal mobility likely continued until the terminal Pleistocene (Gaffney, Ford, and Summerhayes Citation2015, Gaffney Citation2021a). Those groups frequenting southern New Guinea would have been acutely aware of the major changes that were taking place to the shoreline, as the Arafura and Torres plains flooded, creating the island of New Guinea. People visiting the north coast may have been totally unaware that they were becoming islanders, with only small alterations occurring to the steep northern shoreline (until information spread between highland groups).
Mode C: overwater movement to truly insular rainforests
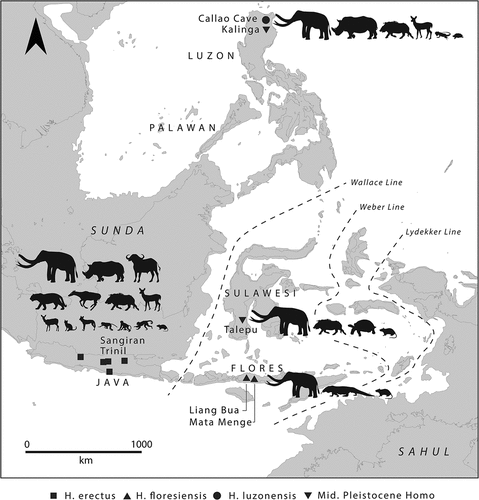
The first hominins to disperse into tropical forested islands proper moved from Sunda to Flores over 1 million years ago (Brumm et al. Citation2010), to Sulawesi sometime between 780,000 and 200,000 years ago (van den Bergh et al. Citation2016a), and to the Philippines before 777,000–631,000 years ago (Ingicco et al. Citation2018). Although these islands were reconfigured in shape and size during the Pleistocene (), the changes were minor compared with flooding of the Sunda and Sahul shelves. Insular and forested conditions had important impacts on hominins throughout climatic fluctuations, producing small-bodied and small-brained Homo floresiensis and Homo luzonensis (Brumm et al. Citation2016; Détroit et al. Citation2019; van den Bergh et al. Citation2016b). It is unclear what the ‘Denisovan’ branch of hominins looked like that may have lived somewhere in western Wallacea (Teixeira et al. Citation2021). What is clear is that substantial morphological variation existed amongst these hominins in the insular tropics, mimicking what we see in non-hominoid primates (Foley Citation1991), and likely reflects limited geneflow between mainland and island populations (Leppard Citation2015).
Figure 5. Settings for Mode C peopling of rainforested islands (overwater movement to island rainforests that remain insular) with key archaeological sites. I: the Philippines; II: Wallacea; III: Near Oceania; IV: Remote Oceania; V: Madagascar; VI: the Caribbean. Note, this is not an exhaustive list, and omits the Indian Ocean, Andaman Islands, and several other minor theatres of migration.
Despite occupying these insular environments for one million years, either continuously or in a recurring series of colonisation and extinction events, there is little evidence that Early–Middle Pleistocene hominins substantially adapted their behaviour to island life. For instance, there is no evidence that marine resources like fish and shellfish were incorporated into the diet (Gaffney Citation2021a; O’Connor et al. Citation2017a; Rabett Citation2018), although it is important to note that none of the sites currently on record were coastal during Early to Middle Pleistocene occupation. It is possible, however, that hominins were exploring closed canopy rainforests on these islands. The hominins hunted megafauna including stegodon, rhinoceroses, cervids, suids, and varanids, which suggests the islands were mosaics of grassland, open woodland, and tropical forests (). Stegodon likely thrived in dense closed canopy tropical forests (Ma et al. Citation2019; Zhang et al. Citation2017), while varanids tend to inhabit more open woodlands, savannah grasslands, and, particularly, dry deciduous rainforest (Imansyah et al. Citation2008). Several cervid remains associated with H. luzonensis display cutmarks, akin to that produced using bamboo knives (Manalo Citation2011), which may further suggest explorations of tropical forest. However, the absence of familiar megafauna likely prevented hominin populations establishing themselves in the smaller and faunally depauperate islands east of the Weber biogeographic line, where animals tend to be small, agile, arboreal, and nocturnal and therefore difficult to capture ().
Figure 6. Faunal complexes associated with key Early–Middle Pleistocene hominin sites in Wallacea and the Philippines (data from Gaffney Citation2021a, Table S1). This includes the Ngandong fauna from Java, associated with H. erectus at Trinil and Sangiran, the Mata Menge fauna from Flores associated with an ancestor to H. floresiensis, and the Talepu fauna from Sulawesi and the Kalinga fauna from Luzon, both associated with an as-yet unidentified hominin. Base map shows present coastline transposed over a – 120 m shoreline, which can be considered an approximate maximum lowstand throughout the Middle Pleistocene.
Several discrete H. sapiens populations dispersed into insular tropical rainforests from MIS-4 to MIS-3, first in Wallacea pushing beyond the Weber Line (O’Connor Citation2010), then in the Philippines (Pawlik and Ronquillo Citation2003), and Near Oceania (Summerhayes and Ford Citation2014). Later populations moved into the Caribbean (Napolitano et al. Citation2019) and possibly Madagascar (Hansford et al. Citation2018) during the Early to Mid Holocene (MIS-1), and Remote Oceania (Rieth and Athens Citation2019) and finally parts of the Indian Ocean (Fuller et al. Citation2011) in the Late Holocene. In these contexts, H. sapiens made longer and more frequent water-crossings than previous hominins had, employing what must have been well-developed seafaring and wayfinding practices to move out of sight of land to reach, for example, Manus (Fredericksen, Spriggs, and Ambrose Citation1993), the Talaud group (Tanudirjo Citation2005), and Grenada (Wilson Citation2007, 45).
Initial Mode C movements followed long-term behavioural trajectories. As shown in , upon arriving to Sulawesi H. sapiens clearly hunted rainforest fauna (Aubert et al. Citation2019; Brumm et al. Citation2018; Simons and Bulbeck Citation2004) that were, in many ways, similar to those encountered in the equatorial forest refugia of Sunda (Piper and Rabett Citation2016). On Madagascar, the earliest tentative sites dated to the Early to Mid Holocene, where megafauna were possibly hunted and butchered, occur in areas today characterised by open forests (spiny thickets and succulent woodlands) (Douglass et al. Citation2019), which would have been similar to the increasingly open conditions of eastern Africa (McWethy et al. Citation2016).
Table 1. Key sites with associated faunal remains reflecting initial Mode C occupation. Primarily terrestrial faunal assemblages are marked in green, primarily marine assemblages are marked blue, and mixed assemblages are marked orange. Note, these lists omit the important terrestrial plant food component of the diet.
However, as H. sapiens moved into smaller island rainforests, the earliest faunal assemblages suggest colonists increasingly employed broad-spectrum foraging practices (Piper and Rabett Citation2014). Owing to the reduced diversity of available game, they intensively targeted a number of rainforest and littoral resources (Chinique de Armas et al. Citation2019). Interregional commonalities in these resources included fruit bats which could be found in large caves, along with small and arboreal forest game such as monkeys, squirrels, possums, cuscus, and rodents. Some groups in Wallacea may have even developed pelagic fishing technology by MIS-3 (O’Connor et al. Citation2011b) and incorporated substantial marine resources into their diets (Roberts et al. Citation2020). Although small forested islands can pose an existential risk to terrestrially adapted groups, those populations with a long-term history of exploiting the littoral and pelagic zones would find small islands ideal, for instance, in the later expansion of Pleistocene groups onto Talaud (Ono et al. Citation2015), Gebe (Bellwood et al. Citation1998), and Kisar (O’Connor et al. Citation2019). It is in these areas, particularly in eastern Wallacean islands, that we find the highest marine ecodiversity on the planet, with an abundance of marine and coastal resources.
Part of what facilitated expansions into increasingly small and biotically depauperate islands was people’s niche constructive tendencies (Fitzpatrick and Giovas Citation2021), which made these ecologies more familiar to incomers. For instance, after several millennia of movements around the Bismarck Archipelago, people began to translocate marsupials from mainland Sahul for hunting (Heinsohn Citation2003). In the small islands of Remote Oceania, human movements to new islands more rapidly led to the import and cultivation of exotic tree species (Huebert and Allen Citation2020), as well as clearances for agricultural crops and the transport of animals for hunting and husbandry (Anderson Citation2009).
Mode D: palimpsests of dispersal
During the Mid to Late Holocene, some technologically Neolithic seafaring groups dispersed into islands that had been occupied for thousands of years by foragers, fishers, and agroforesters, adding to the palimpsest of cultures, languages, and technologies. This occurred as Austronesian speaking groups moved into areas populated by Non-Austronesian speakers in Wallacea and Near Oceania during the fifth and fourth millennia before present (Bellwood Citation2017, 218, 267; Kirch Citation2017, 74; Matsumura et al. Citation2018; Oliveira et al. Citation2022; Skoglund et al. Citation2016), as mainland African populations interacted with Malagasy on Madagascar during the third and second millennia before present (Ekblom et al. Citation2016; Pierron et al. Citation2014), and as pottery making Saladoid populations encountered, and in many places replaced, ‘Archaic’ Casimiroid and Ortoiroid groups in the Caribbean during the third millennium before present (Fernandes et al. Citation2020; Keegan Citation1994; Reid et al. Citation2018). Similar processes may have taken place as various iron-using South Asian populations moved to Sri Lanka in the third millennium before present and gradually intermarried with ancestral Vedda groups (Kulatilake Citation2016; Papiha, Mastana, and Jayasekara Citation1996). In many of these settings, it is clear that dynamic population movements, mixing, and replacements had occurred throughout the Pleistocene and Early Holocene, particularly visible in genetic studies and possibly even involving multiple species in Island Southeast Asia (e.g. Larena et al. Citation2021; Purnomo et al. Citation2021). Novel material culture associated with Neolithicisation, however, make the Mid to Late Holocene dispersal horizons archaeologically salient.
Incoming populations were sometimes accompanied by totally new agricultural systems. For instance, rice farming was introduced from mainland Asia and Taiwan into parts of Island Southeast Asia (Bellwood Citation2011). In other settings, crops already established in the islands were processed in different ways, as in the case of Saladoid populations introducing new types of ceramics and griddles for cooking cassava and breads (Hofman, Ramos, and Jiménez Citation2018). On top of this, new domesticated animals were translocated around islands, as in the case of pigs, dogs, and chickens being introduced into the Pacific from Island Southeast Asia (Summerhayes et al. Citation2019). The gradual domestication of tropical plants and animals, generated both from long-term cultivation practices and wholesale introductions from continental areas, represents a new threshold in how humans interacted with rainforested islands (Roberts et al. Citation2017a). This often came alongside forest clearances in order to make space for gardens, agricultural field systems, and expanding settlements, even further fracturing island rainforest fragments. These changes are marked in palaeoenvironmental records by rapid increases in biomass burning (Haberle, Hope, and van der Kaars Citation2001; Kirleis, Pillar, and Behling Citation2011; Krigbaum Citation2003; Le Blanc Citation2017), and floral and faunal extinctions, which took place, in particular, on smaller oceanic islands (Fall Citation2010; Olson Citation1989; Prebble and Dowe Citation2008; Steadman Citation1995), but also on larger islands like Madagascar (Hixon et al. Citation2021).
What made Mode D peopling processes comparably unique, was that the advent of new migration flows between continental zones to forested islands led communities to mix with, supplant, and even avoid each other; that is, the presence of existing groups and interaction networks actively shaped how incoming groups moved into rainforested islands. In some areas for instance, in the Bismarck Archipelago, incoming Lapita groups were able to settle themselves at arm’s length from established islander groups by occupying marginal, uninhabited islands off the coast of New Ireland and New Britain (Specht Citation2007; Spriggs Citation1997, 88). Lapita seafarers may have even bypassed the Solomon Islands owing to the presence of existing populations (Sheppard Citation2019). These seafarers maintained social and genetic separation over several centuries by reorienting their mobility strategies to frequently cross large stretches of ocean and maintain trade and marriage links with related groups (Lipson et al. Citation2018; Shaw et al. Citation2022; Skoglund et al. Citation2016). As a proxy for this, obsidian from the Bismarck Archipelago was widely distributed to Lapita groups as far east as Fiji (Best Citation1987), and in the opposite direction to contemporary potting communities on Borneo (Chia Citation2003). Keegan and Diamond (Citation1987) characterised these seafaring groups as ‘supertramps’: excluded from larger landmasses by established populations but able to make use of a wide range of resources on several smaller islands. This demonstrates how distinct communities of humans can behaviourally differentiate themselves to carve out different niches in the same ecological space.
Human adaptation and the peopling of island rainforests
The diversity and dynamism engendered by island rainforests had important implications for how hominins moved through, lived in, transformed, and adapted to these ecological spaces. During the Pleistocene and Holocene, humans have come to inhabit highly diverse island rainforest environments such as large islands with dense lowland evergreens, rugged interiors covered in montane rainforest, and small peripheral islands with patchy forest fragments (Keegan and Diamond Citation1987; Roberts and Petraglia Citation2015). In keeping with classical biogeographic models (e.g. MacArthur and Wilson Citation1967), insular rainforests compounded the adaptive challenges produced by isolation, remoteness, and the unevenness and unpredictability of resources, and eventually generated exceptional novelty in human behaviour, language, and physiology. For many foraging groups, these contingencies encouraged a kind of synchronic dynamism, involving frequent movement under the canopy or around the coast, and, in many H. sapiens populations, interisland connectivity during initial stages of the colonisation process and at times of population growth.
However, the ontogenic processes that shaped human behaviour and these wider ecologies were also highly variable. Expanding upon recent paradigm shifts in the general dynamic theory of biogeography (Ávila et al. Citation2019; Borregaard, Matthews, and Whittaker Citation2016; Fernández-Palacios et al. Citation2016; Whittaker, Triantis, and Ladle Citation2008), it is clear that rainforested islands experienced repeated cycles of insularisation and forest expansion and contraction, which presented people with unique and often productive affordances around shifting forest perimeters and shorelines. Diachronically, this produced a continuous dynamism between environmental flux and human behavioural adaptation. In many ways, these developmental pathways played out differently amongst regional hominin populations, and made island rainforests distinct from rainforested continental zones.
Amongst H. sapiens, insularisation encouraged the efflorescence of behavioural adaptive shifts, at times including abandonment, but more commonly, at least on larger rainforested islands, involving persistence and intensification of established rainforest foraging or maritime traditions. In several settings, the movement through a range of forest ecotones, particularly from the coast to rainforested interiors, seems to have been key to maintaining an aggregate of reliable and varied resources amongst mobile hunter gatherers. This is in contrast with H. erectus which became extinct when forests expanded and islands took shape. Perhaps this species was not able to shift regularly between a wide array of foraging strategies or devise ways to take advantage of novel subsistence opportunities when they were encountered.
In a similar way, movements into marine islands led H. sapiens initially to improvise and diversify, but then specialise on maritime resources or small native island fauna and flora, pushing the envelope of what constituted habitable spaces for our species. Initial expansions probably involved frequenting islands by boat for exploration and resource acquisition (speculatively, to acquire fruit bats, birds, small docile mammals, marine animals, and endemic plants) prior to sustained colonisation. That is, there are advantages that uninhabited islands could provide mobile foraging expeditions, and incorporating such spaces into growing archipelagic resource networks was probably a key factor in acclimatising humans to the conditions of small and depauperate forested islands. By occupying these islands regularly, and learning to live on them over many generations, it then scaffolded the possibility for future expansions, allowing later peoples to push into tiny coral atolls, small volcanic islands, temperate and even, on occasion, sub-Antarctic waters. On the other hand, Homo floresiensis, Homo luzonensis, and their ancestors lived on tropical forested islands, and possibly made use of rainforest zones, but did not dramatically reconfigure their subsistence strategies beyond those practiced on Sunda, did not push into small islands (<10,000 km2) lacking megafauna, and became extinct at some time in the Late Pleistocene.
As this paper demonstrates, adaptive flexibility in the tropics emerged in tandem with ecological flux both inside and out of Africa. These developing capacities enabled, and were further encouraged by, movements into novel and challenging environments, including island rainforests. Amongst H. sapiens, innovation and improvisation during initial stages of colonisation were often followed by long periods of specialisation, as humans became increasingly attuned to, and enskilled within, these environments. Specialisation, facilitated by tightly regulated learning systems, is sometimes viewed as ‘stasis’ in the archaeological record, but it can be highly adaptive and went hand in hand with diversification in island rainforests. Lastly, human tendencies to reshape the environment acted as adaptive buffers, bringing these novel environments closer to the familiar, and allowed humans more readily to engage with these new worlds.
Acknowledgments
The ideas in this article were first developed for my PhD and I thank Cyprian Broodbank and Graeme Barker for their supervision. I am grateful to Anna Florin, Tom Leppard, and Cyprian Broodbank for comments on the draft manuscript; I thank also Adam Brumm and one anonymous reviewer for their feedback. The article was written while a visiting fellow in the Archaeology Programme, School of Social Sciences, University of Otago.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Notes
1. Throughout the text, I use the terms peopling, populating, and colonisation in the biogeographic sense to mean the ways that humans established themselves in a new environment.
References
- Ali, J.R., and M. Huber. 2010. “Mammalian Biodiversity on Madagascar Controlled by Ocean Currents.” Nature 463 (7281): 653–656. doi:10.1038/nature08706.
- Allaby, M. 2006. Tropical Forests. New York: Chelsea House Publishers.
- Allen, J., C. Gosden, R. Jones, and J.P. White. 1988. “Pleistocene Dates for the Human Occupation of New Ireland, Northern Melanesia.” Nature 331 (6158): 707–709. doi:10.1038/331707a0.
- Anderson, A. 2009. “The Rat and the Octopus: Initial Human Colonization and the Prehistoric Introduction of Domestic Animals to Remote Oceania.” Biological Invasions 11 (7): 1503–1519. doi:10.1007/s10530-008-9403-2.
- Anderson, A. 2018. “Ecological Contingency Accounts for Earliest Seagoing in the Western Pacific Ocean.” Journal of Island and Coastal Archaeology 13 (2): 220–230. doi:10.1080/15564894.2016.1277286.
- Arifin, K. 2017. “Terminal Pleistocene and Early Holocene Human Occupation in the Rainforests of East Kalimantan.” In New Perspectives in Southeast Asian and Pacific Prehistory, edited by P.J Piper, H. Matsumura, and D. Bulbeck, 97–124. Canberra: ANU Press.
- Aubert, M., R. Lebe, A.A. Oktaviana, M. Tang, B. Burhan, Hamrullah, A. Jusdi 2019. “Earliest Hunting Scene in Prehistoric Art.” Nature 576 (7787): 442–445. doi:10.1038/s41586-019-1806-y.
- Ávila, S.P., C. Melo, B. Berning, S. Nuno, R. Quartau, K.F. Rijsdijk, R.S. Ramalho. 2019. “Towards a ‘Sea-Level Sensitive’ Dynamic Model: Impact of Island Ontogeny and Glacio-Eustasy on Global Patterns of Marine Island Biogeography.” Biological Reviews 94 (3): 1116–1142. doi:10.1111/brv.12492.
- Balme, J. 2013. “Of Boats and String: The Maritime Colonisation of Australia.” Quaternary International 285: 68–75. doi:10.1016/j.quaint.2011.02.029.
- Banerjee, P.K. 2000. “Holocene and Late Pleistocene Relative Sea Level Fluctuations along the East Coast of India.” Marine Geology 167 (3–4): 243–260. doi:10.1016/S0025-3227(00)00028-1.
- Barker, G., L. Lloyd-Smith, H. Barton, F. Cole, Chrishunt, P.J. Piper, R. Rabett, V. Paz, and K. Szabó. 2011. “Foraging-farming Transitions at the Niah Caves, Sarawak, Borneo.” Antiquity 85 (328): 492–509. doi:10.1017/S0003598X00067909.
- Barton, H., P.J. Piper, R. Rabett, and I. Reeds. 2009. “Composite Hunting Technologies from the Terminal Pleistocene and Early Holocene, Niah Cave, Borneo.” Journal of Archaeological Science 36 (8): 1708–1714. doi:10.1016/j.jas.2009.03.027.
- Bellwood, P. 2011. “The Checkered Prehistory of Rice Movement Southwards as a Domesticated Cereal-From the Yangzi to the Equator.” Rice 4 (3–4): 93–103. doi:10.1007/s12284-011-9068-9.
- Bellwood, P. 2017. First Islanders. Hoboken, New Jersey: John Wiley & Sons.
- Bellwood, P., G. Nitihaminoto, G. Irwin, Gunadi, A. Waluyo, and D. Tanudirjo. 1998. “35,000 Years of Prehistory in the Northern Moluccas.” In Bird’s Head Approaches: Irian Jaya Studies, A Programme for Interdisciplinary Research, edited by G.-J. Bartstra, 233–275. Rotterdam: Balkema.
- Best, S. 1987. “Long Distance Obsidian Travel and Possible Implications for the Settlement of Fiji.” Archaeology in Oceania 22 (1): 31–32. doi:10.1002/j.1834-4453.1987.tb00161.x.
- Bettis, E.A., A.K. Milius, S.J. Carpenter, R. Larick, Y. Zaim, Y. Rizal, R.L. Ciochon 2009. “Way Out of Africa: Early Pleistocene Paleoenvironments Inhabited by Homo Erectus in Sangiran, Java.” Journal of Human Evolution 56 (1): 11–24. doi:10.1016/j.jhevol.2008.09.003.
- Bierregaard, R.O., Jr, and T.Ε. Lovejoy. 1989. “Effects of Forest Fragmentation on Amazonian Understory Bird Communities.” Acta Amazonica 19: 215–241. doi:10.1590/1809-43921989191241.
- Bierregaard, R.O., Jr, T.E. Lovejoy, V. Kapos, A.A. Dos Santos, and R.W. Hutchings. 1992. “The Biological Dynamics of Tropical Rainforest Fragments: A Prospective Comparison of Fragments and Continuous Forest.” BioScience 42 (11): 859–866. doi:10.2307/1312085.
- Bird, M.I., S.A. Condie, S. O’Connor, D. O’Grady, C. Reepmeyer, S. Ulm, M. Zega, F. Saltré, and C.J.A. Bradshaw. 2019. “Early Human Settlement of Sahul Was Not an Accident.” Scientific Reports 9 (1): 1–10. doi:10.1038/s41598-019-42946-9.
- Borregaard, M.K., T.J. Matthews, and R.J. Whittaker. 2016. “The General Dynamic Model: Towards a Unified Theory of Island Biogeography?” Global Ecology and Biogeography 25 (7): 805–816. doi:10.1111/geb.12348.
- Bradshaw, C.J.A., K. Norman, S. Ulm, A.N. Williams, C. Clarkson, J. Chadœuf, S.C. Lin 2021. “Stochastic Models Support Rapid Peopling of Late Pleistocene Sahul.” Nature Communications 2021 12:1 12 (1): 1–11.
- Bradshaw, C.J.A., S. Ulm, A.N. Williams, M.I. Bird, R.G. Roberts, Z. Jacobs, F. Laviano 2019. “Minimum Founding Populations for the First Peopling of Sahul.” Nature Ecology and Evolution 3 (7): 1057–1063. doi:10.1038/s41559-019-0902-6.
- Brumm, A., B. Hakim, M. Ramli, M. Aubert, G.D. van den Bergh, B. Li, B. Burhan 2018. “A Reassessment of the Early Archaeological Record at Leang Burung 2, A Late Pleistocene Rock-Shelter Site on the Indonesian Island of Sulawesi.” PLOS ONE 13 (4): e0193025. doi:10.1371/journal.pone.0193025.
- Brumm, A., G.M. Jensen, G.D. van den Bergh, M.J. Morwood, I. Kurniawan, F. Aziz, and M. Storey. 2010. “Hominins on Flores, Indonesia, by One Million Years Ago.” Nature 464 (7289): 748–752. doi:10.1038/nature08844.
- Brumm, A., G.D. van den Bergh, M. Storey, I. Kurniawan, B. V. Alloway, R. Setiawan, E. Setiyabudi 2016. “Age and Context of the Oldest Known Hominin Fossils from Flores.” Nature 534 (7606): 249–253. doi:10.1038/nature17663.
- Bulbeck, D., I. Sumantrai, and P. Hiscock. 2004. “Leang Sakapao 1, a Second Dated Pleistocene Site from South Sulawesi, Indonesia.” In Quaternary Research in Indonesia, edited by S.G. Keates and J.M. Pasveer, 111–128. Ledien: A.A. Balkema.
- Carson, M.T., and H.C. Hung. 2017. Substantive Evidence of Initial Habitation in the Remote Pacific: Archaeological Discoveries at Unai Bapot in Saipan, Mariana Islands. Oxford: Archaeopress.
- Chia, S. 2003. “Obsidian sourcing at Bukit Tengkorak, Sabah, Malaysia.” Sabah Socieity Journal 20: 45–63.
- Chinique de Armas, Y., R. Rodríguez Suárez, W.M. Buhay, and M. Roksandic. 2019. “Subsistence Strategies and Food Consumption Patterns of Archaic Age Populations from Cuba.” In Early Settlers of the Insular Caribbean: Dearchaizing the Archaic, edited by C. Hofman and A.T. Antczak, 107–120. Leiden: Sidestone Press.
- Choi, K., and D. Driwantoro. 2007. “Shell Tool Use by Early Members of Homo Erectus in Sangiran, Central Java, Indonesia: Cut Mark Evidence.” Journal of Archaeological Science 34 (1): 48–58. doi:10.1016/j.jas.2006.03.013.
- Clarkson, C., Z. Jacobs, B. Marwick, R. Fullagar, L. Wallis, M. Smith, R.G. Roberts, et al. 2017. “Human Occupation of Northern Australia by 65,000 Years Ago.” Nature 547 (7663): 306–310. doi:10.1038/nature22968.
- Cooke, S.B., L.M. Dávalos, A.M. Mychajliw, S.T. Turvey, and N.S. Upham. 2017. “Anthropogenic Extinction Dominates Holocene Declines of West Indian Mammals.” Annual Review of Ecology, Evolution, and Systematics 48 (1): 301–327. doi:10.1146/annurev-ecolsys-110316-022754.
- Corlett, R.T., and R.B. Primack. 2011. Tropical Rain Forests: An Ecological and Biogeographical Comparison. 2nd ed. Oxford: Wiley-Blackwell.
- Crabtree, S.A., D.A. White, C.J.A. Bradshaw, F. Saltré, A.N. Williams, R.J. Beaman, M.I. Bird, and S. Ulm. 2021. “Landscape Rules Predict Optimal Superhighways for the First Peopling of Sahul.” Nature Human Behaviour 5 (10): 1303–1313. doi:10.1038/s41562-021-01106-8.
- Cullen, L., R.E. Bodmer, and C. Valladares Pádua. 2000. “Effects of Hunting in Habitat Fragments of the Atlantic Forests, Brazil.” Biological Conservation 95 (1): 49–56. doi:10.1016/S0006-3207(00)00011-2.
- Cutler, K.B., R.L. Edwards, F.W. Taylor, H. Cheng, J. Adkins, C.D. Gallup, P.M. Cutler, G.S. Burr, and A.L. Bloom. 2003. “Rapid Sea-Level Fall and Deep-Ocean Temperature Change since the Last Interglacial Period.” Earth and Planetary Science Letters 206 (3–4): 253–271. doi:10.1016/S0012-821X(02)01107-X.
- David, B., I. McNiven, R. Mitchell, M. Orr, S. Haberle, L. Brady, and J. Crouch. 2004. “Badu 15 and the Papuan-Austronesian Settlement of Torres Strait.” Archaeology in Oceania 39 (2): 65–78. doi:10.1002/j.1834-4453.2004.tb00564.x.
- Détroit, F., A.S. Mijares, J. Corny, G. Daver, C. Zanolli, E. Dizon, E. Robles, R. Grün, and P.J. Piper. 2019. “A New Species of Homo from the Late Pleistocene of the Philippines.” Nature 568 (7751): 181–186. doi:10.1038/s41586-019-1067-9.
- Douglass, K., S. Hixon, H.T. Wright, L.R. Godfrey, B.E. Crowley, B. Manjakahery, T. Rasolondrainy, Z. Crossland, and C. Radimilahy. 2019. “A Critical Review of Radiocarbon Dates Clarifies the Human Settlement of Madagascar.” Quaternary Science Reviews 221: 105878. doi:10.1016/j.quascirev.2019.105878.
- Ekblom, A., P. Lane, C. Radimilahy, J.-A. Rakotoarisoa, P. Sinclair, and M. Virah-Sawmy. 2016. “Migration and Interaction between Madagascar and Eastern Africa, 500 BCE–1000 CE: An Archaeological Perspective.” In Early Exchange between Africa and the Wider Indian Ocean World, edited by G. Campbell, 195–230. London: Palgrave Macmillan.
- Erwin, T.L. 2001. “Forest Canopies, Animal Diversity.” In Encyclopedia of Biodiversity, edited by S. Levin, 19–25. London: Academic Press.
- Fall, P.L. 2010. “Pollen Evidence for Plant Introductions in a Polynesian Tropical Island Ecosystem, Kingdom of Tonga.” In Altered Ecologies: Fire, Climate and Human Influence, edited by S. Haberle, J. Stevenson, and M. Prebble, 253–271. Canberra: ANU Press.
- Fernandes, D.M., K.A. Sirak, H. Ringbauer, J. Sedig, N. Rohland, O. Cheronet, M. Mah 2020. “A Genetic History of the Pre-Contact Caribbean.” Nature 2020 590:7844 590 (7844): 103–110.
- Fernández-Palacios, J.M., K.F. Rijsdijk, S.J. Norder, R. Otto, L. de Nascimento, S. Fernández-Lugo, E. Tjørve, and R.J. Whittaker. 2016. “Towards a Glacial-Sensitive Model of Island Biogeography.” Global Ecology and Biogeography 25 (7): 817–830. doi:10.1111/geb.12320.
- Field, J., M. Fillios, and S. Wroe. 2008. “Chronological Overlap between Humans and Megafauna in Sahul (Pleistocene Australia-New Guinea): A Review of the Evidence.” Earth-Science Reviews 89 (3–4): 97–115. doi:10.1016/j.earscirev.2008.04.006.
- Fitzpatrick, S.M., and C.M. Giovas. 2021. “Tropical Islands of the Anthropocene: Deep Histories of Anthropogenic Terrestrial–Marine Entanglement in the Pacific and Caribbean.” Proceedings of the National Academy of Sciences 118 (40): 2022209118. doi:10.1073/pnas.2022209118.
- Florin, S.A., A.S. Fairbairn, M. Nango, D. Djandjomerr, B. Marwick, R. Fullagar, M. Smith, L.A. Wallis, and C. Clarkson. 2020. “The First Australian Plant Foods at Madjedbebe, 65,000–53,000 Years Ago.” Nature Communications 2020 11:1 11 (1): 1–8.
- Florin, S.A., P. Roberts, B. Marwick, N.R. Patton, J. Shulmeister, C.E. Lovelock, L.A. Barry 2021. “Pandanus Nutshell Generates a Palaeoprecipitation Record for Human Occupation at Madjedbebe, Northern Australia.” Nature Ecology & Evolution 5 (3): 295–303. doi:10.1038/s41559-020-01379-8.
- Foley, R.A. 1991. “How Many Species of Hominid Should There Be?” Journal of Human Evolution 20 (5): 413–427. doi:10.1016/0047-2484(91)90007-I.
- Fox, R.B. 1970. The Tabon Caves: Archaeological Explorations and Excavations on Palawan Island, Philippines. Manilla: National Museum of the Philippines.
- Fredericksen, C., M. Spriggs, and W. Ambrose. 1993. “Pamwak Rockshelter: A Pleistocene Site on Manus Island, Papua New Guinea.” In Sahul in Review: Pleistocene Archaeology in Australia, New Guinea and Island Melanesia, edited by M.A. Smith, M. Spriggs, and B. Fankhauser, 144–152. Canberra: Department of Prehistory, Research School of Pacific Studies, Australian National University.
- Fuller, D.Q., N. Boivin, T. Hoogervorst, and R. Allaby. 2011. “Across the Indian Ocean: The Prehistoric Movement of Plants and Animals.” Antiquity 85 (328): 544–558. doi:10.1017/S0003598X00067934.
- Gaffney, D. 2021a. “Pleistocene Water Crossings and Adaptive Flexibility within the Homo Genus.” Journal of Archaeological Research 29 (2): 255–326.
- Gaffney, D. 2021b. “Human Behavioural Dynamics in Island Rainforests: Evidence from the Raja Ampat Islands, West Papua.” PhD diss., University of Cambridge.
- Gaffney, D., and T. Denham. 2021. “The Archaeology of Social Transformation in the New Guinea Highlands.” In The Oxford Handbook of the Archaeology of Indigenous Australia and New Guinea, edited by I. McNiven and B. David, 1–41. Oxford: Oxford University Press.
- Gaffney, D., A. Ford, and G. Summerhayes. 2015. “Crossing the Pleistocene-Holocene Transition in the New Guinea Highlands: Evidence from the Lithic Assemblage of Kiowa Rockshelter.” Journal of Anthropological Archaeology 39: 223–246. doi:10.1016/j.jaa.2015.04.006.
- Gibson, L., A.J. Lynam, C.J.A. Bradshaw, F. He, D.P. Bickford, D.S. Woodruff, S. Bumrungsri, and W.F. Laurance. 2013. “Near-complete Extinction of Native Small Mammal Fauna 25 Years after Forest Fragmentation.” Science 341 (6153): 1508–1510. doi:10.1126/science.1240495.
- Green, R.C., and M. Cresswell, eds. 1976. Southeast Solomon Islands Cultural History. Auckland: Royal Society of New Zealand.
- Groube, L., J. Chappell, J. Muke, and D. Price. 1986. “A 40,000 year-old Human Occupation Site at Huon Peninsula, Papua New Guinea.” Nature 324 (4): 453–455. doi:10.1038/324453a0.
- Haberle, S.G., G.S. Hope, and S. van der Kaars. 2001. “Biomass Burning in Indonesia and Papua New Guinea: Natural and Human Induced Fire Events in the Fossil Record.” Palaeogeography, Palaeoclimatology, Palaeoecology 171 (3–4): 259–268. doi:10.1016/S0031-0182(01)00248-6.
- Hansford, J., P.C. Wright, A. Rasoamiaramanana, V.R. Pérez, L.R. Godfrey, D. Errickson, T. Thompson, and S.T. Turvey. 2018. “Early Holocene Human Presence in Madagascar Evidenced by Exploitation of Avian Megafauna.” Science Advances 4 (9): 1–7. doi:10.1126/sciadv.aat6925.
- Hawkins, S. 2015. “Human Behavioural Ecology, Anthropogenic Impact and Subsistence Change at the Teouma Lapita Site, Central Vanuatu, 3000-2500 BP.” PhD diss., Australian National University.
- Hawkins, S., S. O’Connor, T.R. Maloney, M. Litster, S. Kealy, J.N. Fenner, K. Aplin 2017. “Oldest Human Occupation of Wallacea at Laili Cave, Timor-Leste, Shows Broad-Spectrum Foraging Responses to Late Pleistocene Environments.” Quaternary Science Reviews 171: 58–72. doi:10.1016/j.quascirev.2017.07.008.
- Heaney, L.R. 2000. “Dynamic Disequilibrium: A Long-Term, Large-Scale Perspective on the Equilibrium Model of Island Biogeography.” Global Ecology and Biogeography 9 (1): 59–74. doi:10.1046/j.1365-2699.2000.00163.x.
- Heinsohn, T. 2003. “Animal Translocation: Long-term Human Influences on the Vertebrate Zoogeography of Australasia (Natural Dispersal versus Ethnophoresy).” Australian Zoologist 32 (3): 351–376. doi:10.7882/AZ.2002.014.
- Hixon, S.W., K.G. Douglass, B.E. Crowley, L.M.A. Rakotozafy, G. Clark, A. Anderson, S. Haberle 2021. “Late Holocene Spread of Pastoralism Coincides with Endemic Megafaunal Extinction on Madagascar.” Proceedings of the Royal Society B 288 (1955): 20211204. doi:10.1098/rspb.2021.1204.
- Hofman, C.L., R.R. Ramos, and J.R.P. Jiménez. 2018. “The Neolithisation of the Northeastern Caribbean: Mobility and Social Interaction.” In The Archaeology of Caribbean and Circum-Caribbean Farmers 6000 BC–AD 1500, edited by B.A. Reid, 71–97. Oxford: Routledge.
- Hope, G., and K. Aplin. 2005. “Environmental Change in the Aru Islands.” In The Archaeology of the Aru Islands, Eastern Indonesia, edited by S. O’Connor, M. Spriggs, and P. Veth, 25–40. Canberra: ANU Press.
- Huebert, J.M., and M.S. Allen. 2020. “Anthropogenic Forests, Arboriculture, and Niche Construction in the Marquesas Islands (Polynesia).” Journal of Anthropological Archaeology 57: 101122. doi:10.1016/j.jaa.2019.101122.
- Imansyah, M.J., T.S. Jessop, C. Ciofi, and Z. Akbar. 2008. “Ontogenetic Differences in the Spatial Ecology of Immature Komodo Dragons.” Journal of Zoology 274 (2): 107–115. doi:10.1111/j.1469-7998.2007.00368.x.
- Ingicco, T., G.D. van den Bergh, C. Jago-On, J.J. Bahain, M.G. Chacón, N. Amano, H. Forestier 2018. “Earliest Known Hominin Activity in the Philippines by 709 Thousand Years Ago.” Nature 557 (7704): 233–237. doi:10.1038/s41586-018-0072-8.
- Irwin, G. 1991. “Pleistocene Voyaging and the Settlement of Greater Australia and its Near Oceanic Neighbours.” In The Report of the Lapita Homeland Project, edited by J. Allen and C. Gosden, 9–19. Canberra: Department of Prehistory, Research School of Pacific Studies. The Australian National University.
- Jones, S.R. 2015. “Eating Identity: An Exploration of Fijian Foodways in the Archaeological Past.” Journal of Indo-Pacific Archaeology 37 (2015): 64–71. doi:10.7152/jipa.v37i0.15000.
- Joordens, J.C.A., F. D’Errico, F.P. Wesselingh, S. Munro, J. De Vos, J. Wallinga, C. Ankjærgaard 2015. “Homo Erectus at Trinil on Java Used Shells for Tool Production and Engraving.” Nature 518 (7538): 228–231. doi:10.1038/nature13962.
- Joordens, J.C.A., F.P. Wesselingh, J. de Vos, H.B. Vonhof, and D. Kroon. 2009. “Relevance of Aquatic Environments for Hominins: A Case Study from Trinil (Java, Indonesia).” Journal of Human Evolution 57 (6): 656–671. doi:10.1016/j.jhevol.2009.06.003.
- Keegan, W.F. 1994. “West Indian Archaeology. 1. Overview and Foragers.” Journal of Archaeological Research 1994 2:3 2 (3): 255–284.
- Keegan, W.F., and J. Diamond. 1987. “Colonization of Islands by Humans: A Biogeographical Perspective.” Advances in Archaeological Method and Theory 10: 49–92.
- Kirch, P. V. 1973. “Prehistoric Subsistence Patterns in the Northern Marquesas Islands, French Polynesia.” Archaeology and Physical Anthropology in Oceania 8 (1): 24–40.
- Kirch, P.V. 2017. On the Road of the Winds: An Archaeological History of the Pacific Islands before European Contact. 2nd ed. Oakland: University of California Press.
- Kirleis, W., V.D. Pillar, and H. Behling. 2011. “Human-environment Interactions in Mountain Rainforests: Archaeobotanical Evidence from Central Sulawesi, Indonesia.” Vegetation History and Archaeobotany 20 (3): 165–179. doi:10.1007/s00334-010-0272-0.
- Kozuch, L., and E.S. Wing. 2010. “Animal Remains from Archaeological Sites on Nevis.” In The Prehistory of Nevis, A Small Island in the Lesser Antilles, edited by S.M. Wilson, 145–186. New Haven: Yale University Press.
- Krigbaum, J. 2003. “Neolithic Subsistence Patterns in Northern Borneo Reconstructed with Stable Carbon Isotopes of Enamel.” Journal of Anthropological Archaeology 22 (3): 292–304. doi:10.1016/S0278-4165(03)00041-2.
- Kulatilake, S. 2016. “The Peopling of Sri Lanka from Prehistoric to Historic Times: Biological and Archaeological Evidence.” In A Companion to South Asia in the Past, edited by G.R. Schug and S.R. Walimbe, 426–436. Hoboken, NJ: John Wiley & Sons.
- Kusmartono, V.P.R., I. Hindarto, and E. Herwanto. 2017. “Late Pleistocene to Recent: Human Activities in the Deep Interior Equatorial Rainforest of Kalimantan, Indonesian Borneo.” Quaternary International 448: 82–94. doi:10.1016/j.quaint.2016.09.025.
- Lampert, R.J. 1979. “The Great Kartan Mystery.” PhD diss., Australian National University.
- Larena, M., F. Sanchez-Quinto, P. Sjödin, J. McKenna, C. Ebeo, R. Reyes, O. Casel 2021. “Multiple Migrations to the Philippines during the Last 50,000 Years.” Proceedings of the National Academy of Sciences of the United States of America 118 (13): 2026132118. doi:10.1073/pnas.2026132118.
- Laurance, W.F. 1991. “Edge Effects in Tropical Forest Fragments: Application of a Model for the Design of Nature Reserves.” Biological Conservation 57 (2): 205–219. doi:10.1016/0006-3207(91)90139-Z.
- Laurance, W.F., and E. Yensen. 1991. “Predicting the Impacts of Edge Effects in Fragmented Habitats.” Biological Conservation 55 (1): 77–92. doi:10.1016/0006-3207(91)90006-U.
- Laurence, W.F., T. Lovejoy, H.L. Vasconcelos, E.M. Bruna, R.K. Didham, P.C. Stouffer, C. Gascon, R.O. Bierredaard, S.G. Laurance, and E. Sampaio. 2002. “Ecosystem Decay of Amazonian Forest Fragments: A 22-year Investigation.” Conservation Biology 16 (3): 605–618. doi:10.1046/j.1523-1739.2002.01025.x.
- Leavesley, M.G. 2004. “Trees to the Sky: Prehistoric Hunting in New Ireland, Papua New Guinea.” PhD diss., Australian National University.
- Le Blanc, A.R. 2017. “Lake Sediment-Based Reconstructions of Late-Holocene Lowland Environments of Dominican Republic and Barbuda, Northern Caribbean.” PhD diss., State University, Blacksburg, Virginia.
- Leppard, T.P. 2015. “Passive Dispersal versus Strategic Dispersal in Island Colonization by Hominins.” Current Anthropology 56 (4): 590–595. doi:10.1086/682325.
- Leppard, T.P., E. Cochrane, D. Gaffney, C. Hofman, J. Laffoon, M. Schmid, and C. Broodbank. inpress. “Global Patterns in Island Colonisation during the Holocene.” Journal of World Prehistory. doi:10.1007/s10963-022-09168-w.
- Lipson, M., P. Skoglund, M. Spriggs, F. Valentin, S. Bedford, R. Shing, H. Buckley 2018. “Population Turnover in Remote Oceania Shortly after Initial Settlement.” Current Biology 28 (7): 1157–1165. doi:10.1016/j.cub.2018.02.051.
- Lundberg, E. 1989. “Preceramic Procurement Patterns at Krum Bay, Virgin Islands.” PhD diss., University of Illinois.
- MacArthur, R.H., and E.O. Wilson. 1963. “An Equilibrium Theory of Insular Zoogeography.” Evolution: International Journal of Organic Evolution 17 (4): 373–387. doi:10.1111/j.1558-5646.1963.tb03295.x.
- MacArthur, R.H., and E.O. Wilson. 1967. The Theory of Island Biogeography. Princeton, NJ: Princeton University Press.
- Mahirta. 2003. “Human Occupation on Rote and Sawu Islands, Nusa Tenggara Timur.” PhD diss., Australian National University.
- Malcolm, J.R. 1988. “Small Mammal Abundances in Isolated and Non-Isolated Primary Forest Reserves near Manaus, Brazil.” Acta Amazonica 18 (3–4): 67–83. doi:10.1590/1809-43921988183083.
- Malcolm, J.R. 1991. “Comparative Abundances of Neotropical Small Mammals by Trap Height.” Journal of Mammalogy 72 (1): 188–192. doi:10.2307/1381995.
- Manalo, K.A. 2011. Preliminary Identification of Cutmark Morphology on Animal Bones: Methods & Applications. MA thesis. University of the Philippines.
- Matsumura, H., K.I. Shinoda, T. Shimanjuntak, A.A. Oktaviana, S. Noerwidi, H.O. Sofian, D. Prastiningtyas 2018. “Cranio-morphometric and aDNA Corroboration of the Austronesian Dispersal Model in Ancient Island Southeast Asia: Support from Gua Harimau, Indonesia.” PLoS ONE 13 (6): e0198689. doi:10.1371/journal.pone.0198689.
- Matsu’ura, S., M. Kondo, T. Danhara, S. Sakata, H. Iwano, T. Hirata, I. Kurniawan 2020. “Age Control of the First Appearance Datum for Javanese Homo Erectus in the Sangiran Area.” Science 367 (6474): 210–214. doi:10.1126/science.aau8556.
- Ma, J., Y. Wang, C. Jin, Y. Hu, and H. Bocherens. 2019. “Ecological Flexibility and Differential Survival of Pleistocene Stegodon orientalis and Elephas maximus in Mainland Southeast Asia Revealed by Stable Isotope (C, O) Analysis.” Quaternary Science Reviews 212: 33–44. doi:10.1016/j.quascirev.2019.03.021.
- McWethy, D.B., F.H. Neumann, F. Steinbruch, C.M. Ryan, and V. Valsecchi. 2016. “Late Quaternary Vegetation Development and Disturbance Dynamics from a Peatland on Mount Gorongosa, Central Mozambique.” Quaternary Science Reviews 137: 221–233. doi:10.1016/j.quascirev.2016.02.004.
- Mijares, A.S., F. Détroit, P. Piper, R. Grün, P. Bellwood, M. Aubert, G. Champion, N. Cuevas, A. De Leon, and E. Dizon. 2010. “New Evidence for a 67,000-year-old Human Presence at Callao Cave, Luzon, Philippines.” Journal of Human Evolution 59 (1): 123–132. doi:10.1016/j.jhevol.2010.04.008.
- Montagnini, F., and C.F. Jordan. 2005. Tropical Forest Ecology: The Basis for Conservation and Management. Berlin: Springer.
- Morwood, M.J., F. Aziz, P. O’Sullivan, Nasruddin, D.R. Hobbs, and A. Raza. 1999. “Archaeological and Palaeontological Research in Central Flores, East Indonesia: Results of Fieldwork 1997-98.” Antiquity 73 (280): 273–286. doi:10.1017/S0003598X00088244.
- Mountain, M.-J. 1991a. “Landscape Use and Environmental Management of Tropical Rainforest by Pre-Agricultural Hunter-Gatherers in Northern Sahulland.” Bulletin of the Indo-Pacific Prehistory Association 11: 54–68. doi:10.7152/bippa.v11i0.11373.
- Mountain, M.-J. 1991b. “Highland New Guinea Hunter-Gatherers: The Evidence of Nombe Rockshelter, Simbu, with Emphasis on the Pleistocene.” PhD diss., Australian National University.
- Nagaoka, L. 1993. “The Faunal Assemlbages from the To’aga Site.” In The To’aga Site, edited by P. V. Kirch and T.L. Hunt, 189–216. Berkeley: Archaeological Research Facility, University of California.
- Napolitano, M.F., R.J. DiNapoli, J.H. Stone, M.J. Levin, N.P. Jew, B.G. Lane, J.T. O’Connor, and S.M. Fitzpatrick. 2019. “Reevaluating Human Colonization of the Caribbean Using Chronometric Hygiene and Bayesian Modeling.” Science Advances 5 (12): 7806–7824. doi:10.1126/sciadv.aar7806.
- Narganes, Y.M. 1991. “Los restos faunísticos del Sitio de Puerto Ferro, Vieques, Puerto Rico.” Proceedings of the Fourteenth Congress of the International Association for Caribbean Archaeology, Barbados Museum and Historical Society, Barbados eds. A. Cummins and P. King. Barbados: Barbados Museum. 94–114.
- Newsom, L.A. 1993. “Native West Indian Plant Use.” PhD diss., University of Florida.
- Newsom, L.A., and E. Wing. 2004. On Land and Sea: Native American Uses of Biological Resources in the West Indies. Tuscaloosa: University of Alabama Press.
- Norman, K., J. Inglis, C. Clarkson, J.T. Faith, J. Shulmeister, and D. Harris. 2018. “An Early Colonisation Pathway into Northwest Australia 70-60,000 Years Ago.” Quaternary Science Reviews 180: 229–239. doi:10.1016/j.quascirev.2017.11.023.
- O’Connell, J.F., J. Allen, M.A.J. Williams, A.N. Williams, C.S.M. Turney, N.A. Spooner, J. Kamminga, G. Brown, and A. Cooper. 2018. “When Did Homo Sapiens First Reach Southeast Asia and Sahul?” Proceedings of the National Academy of Sciences 115 (34): 8482–8490. doi:10.1073/pnas.1808385115.
- O’Connor, S. 1994. “A 6700 BP Date for Island Use in the West Kimberley, Western Australia: New Evidence from High Cliffy Island.” Australian Archaeology 39 (1): 102–107. doi:10.1080/03122417.1994.11681534.
- O’Connor, S. 2010. “Pleistocene Migration and Colonization in the Indo-Pacific Region.” In The Global Origins and Development of Seafaring, edited by A. Anderson, J.H. Barrett, and K. V. Boyle, 41–55. Cambridge: McDonald Institute of Archaeological Research.
- O’Connor, S., K.P. Aplin, M. Spriggs, P. Veth, and L.K. Ayliffe. 2002. “From Savannah to Rainforest: Changing Environments and Human Occupation at Liang Lembudu, Aru Islands, Maluka (Indonesia).” In Bridging Wallace’s Line: The Environmental and Cultural History and Dynamics of the SE-Asian-Australian Region, edited by P. Kershaw, B. David, N. Tapper, D. Penny, and J. Brown. Ämelgasse, Germany: Catena Verlag. 279–306.
- O’Connor, S., A. Barham, K. Aplin, K. Dobney, A. Fairbairn, and M. Richards. 2011a. “The Power of Paradigms: Examining the Evidential Basis for Early to Mid-Holocene Pigs and Pottery in Melanesia.” Journal of Pacific Archaeology 2 (2): 238–240.
- O’Connor, S., A. Barham, M. Spriggs, P. Veth, K. Aplin, and E.S. Pierre. 2010. “Cave Archaeology and Sampling Issues in the Tropics: A Case Study from Lene Hara Cave, a 42,000 Year Old Occupation Site in East Timor,Island Southeast Asia.” Australian Archaeology 71 (1): 29–40. doi:10.1080/03122417.2010.11689382.
- O’Connor, S., J. Louys, S. Kealy, and S.C.S. Carro. 2017a. “Hominin Dispersal and Settlement East of Huxley’s Line the Role of Sea Level Changes, Island Size, and Subsistence Behavior.” Current Anthropology 58: S567–82. doi:10.1086/694252.
- O’Connor, S., S.C. Samper Carro Mahirta, S. Hawkins, S. Kealy, J. Louys, and R. Wood. 2017b. “Fishing in Life and Death: Pleistocene Fish-Hooks from a Burial Context on Alor Island, Indonesia.” Antiquity 91 (360): 1451–1468. doi:10.15184/aqy.2017.186.
- O’Connor, S., M. Mahirta, S. Kealy, C. Boulanger, T. Maloney, S. Hawkins, M C. Langley 2019. “Kisar and the Archaeology of Small Islands in the Wallacean Archipelago.” The Journal of Island and Coastal Archaeology 14 (2): 198–225. doi:10.1080/15564894.2018.1443171.
- O’Connor, S., R. Ono, and C. Clarkson. 2011b. “Pelagic Fishing at 42,000 Years before the Present and the Maritime Skills of Modern Humans.” Science 334 (6059): 1117–1121. doi:10.1126/science.1207703.
- O’Connor, S., G. Robertson, and K.P. Aplin. 2014. “Are Osseous Artefacts a Window to Perishable Material Culture: Implications of an Unusually Complex Bone Tool from the Late Pleistocene of East Timor.” Journal of Human Evolution 67 (1): 108–119. doi:10.1016/j.jhevol.2013.12.002.
- O’Connor, S., and P. Veth. 2008. “Revisiting the Past: Changing Interpretations of Pleistocene Settlement Subsistence and Demography in Northern Australia.” In Archaeology of Oceania: Australia and the Pacific Islands, edited by I. Lilley, 29–47. London: John Wiley & Sons.
- Oliveira, S., K. Nägele, S. Carlhoff, I. Pugach, T. Koesbardiati, A. Hübner, M. Meyer 2022. “Ancient Genomes from the Last Three Millennia Support Multiple Human Dispersals into Wallacea.” Nature Ecology and Evolution 6: 1024–1034.
- Olson, S.L. 1989. “Extinction on Islands: Man as a Catastrophe.” In Conservation for the Twenty-First Century, edited by D. Western and M.C. Pearl, 50–53. New York: Wildlife Conservation International.
- Ono, R., R. Fuentes, A. Pawlik, H.O. Sofian, Sriwigati, N. Aziz, N. Alamsyah, and M. Yoneda. 2020. “Island Migration and Foraging Behaviour by Anatomically Modern Humans during the Late Pleistocene to Holocene in Wallacea: New Evidence from Central Sulawesi, Indonesia.” Quaternary International 554: 90–106. doi:10.1016/j.quaint.2020.03.054.
- Ono, R., N. Nakajima, H. Nishizawa, S. Oda, and S. Soegondho. 2015. “Maritime Migration and Lithic Assemblage on the Talaud Islands in Northern Wallacea during the Late Pleistocene to the Early Holocene.” In Emergence and Diversity of Modern Human Behavior in Paleolithic Asia, edited by Y. Kaifu, M. Izuho, T. Goebel, H. Sato, and A. Ono, 201–213. College Station: Texas A&M University Press.
- Papiha, S.S., S.S. Mastana, and R. Jayasekara. 1996. “Genetic Variation in Sri Lanka.” Human Biology 68 (5): 707–737.
- Pawlik, A.F., and W.P. Ronquillo. 2003. “The Palaeolithic in the Philippines.” Lithic Technology 28 (2): 79–93. doi:10.1080/01977261.2003.11721004.
- Pearson, R.J., P.V. Kirch, and M. Pietrusewsky. 1971. “An Early Prehistoric Site at Bellows Beach, Waimanalo, Oahu, Hawaiian Islands.” Archaeology & Physical Anthropology in Oceania 6 (3): 204–234.
- Perera, N., P. Roberts, and M. Petraglia. 2016. “Bone Technology from Late Pleistocene Caves and Rockshelters of Sri Lanka.” In Osseous Projectile Weaponry, edited by M. Langley, 173–188. Dordrecht: Springer.
- Pierron, D., H. Razafindrazaka, L. Pagani, F.X. Ricaut, T. Antao, M. Capredon, C. Sambo 2014. “Genome-wide Evidence of Austronesian-Bantu Admixture and Cultural Reversion in a Hunter-Gatherer Group of Madagascar.” Proceedings of the National Academy of Sciences of the United States of America 111 (3): 936–941. doi:10.1073/pnas.1321860111.
- Piper, P.J., and R.J. Rabett. 2009. “Hunting in a Tropical Rainforest: Evidence from the Terminal Pleistocene at Lobang Hangus, Niah Caves, Sarawak.” International Journal of Osteoarchaeology 19 (4): 551–565. doi:10.1002/oa.1046.
- Piper, P., and R. Rabett. 2014. “Late Pleistocene Subsistence Strategies in Island Southeast Asia and Their Implications for Understanding the Development of Modern Human Behaviour.” In Southern Asia, Australia and the Search for Human Origins, edited by R. Dennell and M. Porr, 118–134. Cambridge: Cambridge University Press.
- Piper, P., and R. Rabett. 2016. “Vertebrate Fauna from the Niah Caves.” In Archaeological Investigations in the Niah Caves, Sarawak, edited by G. Barker and L. Farr, 401–438. Cambridge: McDonald Institute of Archaeological Research.
- Poulsen, J. 1987. Early Tongapn Prehistory. Canberra: Department of Prehistory, Research School of Pacific Studies, Australian National University.
- Prebble, M., and J.L. Dowe. 2008. “The Late Quaternary Decline and Extinction of Palms on Oceanic Pacific Islands.” Quaternary Science Reviews 27 (27–28): 2546–2567. doi:10.1016/j.quascirev.2008.09.015.
- Pritchard, P.C.H., R.J. Rabett, and P.J. Piper. 2009. “Distinguishing Species of Geoemydid and Trionychid Turtles from Shell Fragments: Evidence from the Pleistocene at Niah Caves, Sarawak.” International Journal of Osteoarchaeology 19 (4): 531–550. doi:10.1002/oa.1038.
- Purnomo, G.A., K.J. Mitchell, S. O’connor, S. Kealy, L. Taufik, S. Schiller, A. Rohrlach 2021. “Mitogenomes Reveal Two Major Influxes of Papuan Ancestry across Wallacea following the Last Glacial Maximum and Austronesian Contact.” Genes 12 (7): 965. doi:10.3390/genes12070965.
- Rabett, R.J. 2018. “The Success of Failed Homo Sapiens Dispersals Out of Africa and into Asia.” Nature Ecology and Evolution 2 (2): 212–219. doi:10.1038/s41559-017-0436-8.
- Rabett, R.J., and P.J. Piper. 2012. “The Emergence of Bone Technologies at the End of the Pleistocene in Southeast Asia: Regional and Evolutionary Implications.” Cambridge Archaeological Journal 22 (1): 37–56. doi:10.1017/S0959774312000030.
- Reid, B.A., P.E. Siegel, N.P. Dunning, C.L. Hofman, S. Rostain, V.D. Thompson, and S.M. Fitzpatrick. 2018. “Caribbean and circum-Caribbean Farmers: An Introduction.” In The Archaeology of Caribbean and Circum-Caribbean Farmers 6000 BC–AD 1500, edited by B.A. Reid, 1–32. Oxford: Routledge.
- Rieth, T.M., and J.S. Athens. 2019. “Late Holocene Human Expansion into near and Remote Oceania: A Bayesian Model of the Chronologies of the Mariana Islands and Bismarck Archipelago.” Journal of Island and Coastal Archaeology 14 (1): 5–16. doi:10.1080/15564894.2017.1331939.
- Rizal, Y., K.E. Westaway, Y. Zaim, G.D. van den Bergh, E.A. Bettis, M.J. Morwood, O.F. Huffman 2020. “Last Appearance of Homo Erectus at Ngandong, Java, 117,000–108,000 Years Ago.” Nature 577 (7790): 381–385. doi:10.1038/s41586-019-1863-2.
- Roberts, P. 2019. Tropical Forests in Prehistory, History, and Modernity. Oxford: Oxford University Press.
- Roberts, P., N. Boivin, J. Lee-Thorp, M. Petraglia, and J. Stock. 2016. “Tropical Forests and the Genus Homo.” Evolutionary Anthropology 25 (6): 306–317. doi:10.1002/evan.21508.
- Roberts, P., C. Hunt, M. Arroyo-Kalin, D. Evans, and N. Boivin. 2017. “The Deep Human Prehistory of Global Tropical Forests and Its Relevance for Modern Conservation.” Nature Plants 3 (8): 17093. doi:10.1038/nplants.2017.93.
- Roberts, P., J. Louys, J. Zech, C. Shipton, S. Kealy, S.S. Carro, S. Hawkins 2020. “Isotopic Evidence for Initial Coastal Colonization and Subsequent Diversification in the Human Occupation of Wallacea.” Nature Communications 11 (1): 1–11. doi:10.1038/s41467-020-15969-4.
- Roberts, P., N. Perera, O. Wedage, S. Deraniyagala, J. Perera, S. Eregama, A. Gledhill, M.D. Petraglia, and J.A. Lee-Thorp. 2015. “Direct Evidence for Human Reliance on Rainforest Resources in Late Pleistocene Sri Lanka.” Science 347 (6227): 1246–1249. doi:10.1126/science.aaa1230.
- Roberts, P., and M. Petraglia. 2015. “Pleistocene Rainforests: Barriers or Attractive Environments for Early Human Foragers?” World Archaeology 47 (5): 718–739. doi:10.1080/00438243.2015.1073119.
- Samper Carro, S.C., S. O’Connor, J. Louys, S. Hawkins, and M. Mahirta. 2016. “Human Maritime Subsistence Strategies in the Lesser Sunda Islands during the Terminal Pleistocene–early Holocene: New Evidence from Alor, Indonesia.” Quaternary International 416: 64–79. doi:10.1016/j.quaint.2015.07.068.
- Sand, C. 2019. “Archaeological Report on Localitites WKO013A and WKO013B at the Site of Lapita (Koné, New Caledonia).” The Journal of the Polynesian Society 107 (1): 7–33.
- Sathiamurthy, E., and H.K. Voris. 2006. “Maps of Holocene Sea Level Transgression and Submerged Lakes on the Sunda Shelf.” The Natural History Journal of Chulalongkorn University 2: 1–43. Supplement.
- Sémah, A.M., and F. Sémah. 2012. “The Rain Forest in Java through the Quaternary and Its Relationships with Humans (Adaptation, Exploitation and Impact on the Forest).” Quaternary International 249: 120–128. doi:10.1016/j.quaint.2011.06.013.
- Sémah, A.M., F. Sémah, T. Djubiantono, and B. Brasseur. 2010. “Landscapes and Hominids’ Environments: Changes between the Lower and the Early Middle Pleistocene in Java (Indonesia.” Quaternary International 223–224: 451–454. doi:10.1016/j.quaint.2009.07.017.
- Sémah, A.M., F. Sémah, A.M. Moigne, T. Ingicco, A. Purnomo, T. Simanjuntak, and H. Widianto. 2016. “The Palaeoenvironmental Context of the Palaeolithic of Java: A Brief Review.” Quaternary International 416: 38–45. doi:10.1016/j.quaint.2015.11.026.
- Shaw, B., S. Hawkins, L. Becerra-Valdivida, C.S.M. Turney, S. Coxe, V. Kewebu, J. Haro, et al. 2022. “Frontier Interaction with Papuan Populations Set Stage for Pacific Colonisation.” Nature Ecology and Evolution. 6 (6): 802–812.
- Sheppard, P. 2019. “Early Lapita Colonisation of Remote Oceania: An Update on the Leapfrog Hypothesis.” In Debating Lapita: Distribution, Chronology, Society and Subsistence, edited by S. Bedford and M. Spriggs, 158–161. Canberra: ANU Press.
- Sim, R. 1998. “The Archaeology of Isolation? Prehistorical Occupation in the Furneux Group of Islands, Bass Strait, Tasmania.” PhD diss., Autralian National University.
- Simons, A., and D. Bulbeck. 2004. “Late Quaternary Faunal Successions in South Sulawesi, Indonesia.” In Quaternary Research in Indonesia, edited by S.G. Keates and J.M. Pasveer, 167–189. Leiden: A.A. Balkema.
- Skoglund, P., C. Posth, K. Sirak, M. Spriggs, F. Valentin, S. Bedford, G.R. Clark 2016. “Genomic Insights into the Peopling of the Southwest Pacific.” Nature 538 (7626): 510–513. doi:10.1038/nature19844.
- Specht, J. 2007. “Small Islands in the Big Picture: The Formative Period of Lapita in the Bismarck Archipelago.” In Oceanic Explorations: Lapita and Western Pacific Settlement, edited by S. Bedford, C. Sand, and S.P. Connaughton, 51–70. Canberra: ANU Press.
- Spriggs, M. 1997. The Island Melanesians. Cambridge, MA: Blackwell.
- Steadman, D. 1995. “Prehistoric Extinctions of Pacific Island Birds: Biodiversity Meets Zooarchaeology.” Science 267 (5201): 1123–1131. doi:10.1126/science.267.5201.1123.
- Stimpson, C.M. 2012. “Local Scale, Proxy Evidence for the Presence of Closed Canopy Forest in North-western Borneo in the Late Pleistocene: Bones of Strategy I Bats from the Archaeological Record of the Great Cave of Niah, Sarawak.” Palaeogeography, Palaeoclimatology, Palaeoecology 331–332: 136–149. doi:10.1016/j.palaeo.2012.03.007.
- Storm, P., F. Aziz, J. de Vos, D. Kosasih, S. Baskoro, Ngaliman, and L.W. van den Hoek Ostende. 2005. “Late Pleistocene Homo Sapiens in a Tropical Rainforest Fauna in East Java.” Journal of Human Evolution 49 (4): 536–545. doi:10.1016/j.jhevol.2005.06.003.
- Summerhayes, G.R., and A. Ford. 2014. “Late Pleistocene Colonisation and Adaptation in New Guinea Implications for Modelling Modern Human Behaviour.” In Southern Asia, Australia and the Search for Human Origins, edited by R. Dennell and M. Porr, 213–227. Cambridge: Cambridge University Press.
- Summerhayes, G.R., M. Leavesley, A. Fairbairn, H. Mandui, J. Field, A. Ford, and R. Fullagar. 2010. “Human Adaptation and Plant Use in Highland New Guinea 49,000 to 44,000 Years Ago.” Science 330 (6000): 78–81. doi:10.1126/science.1193130.
- Summerhayes, G.R., K. Szabó, A. Fairbairn, M. Horrocks, S. Mcpherson, and A. Crowther. 2019. “Early Lapita Subsistence: The Evidence from Kamgot, Anir Islands, New Ireland Province, Papua New Guinea.” In Debating Lapita, edited by S. Bedford and M. Spriggs, 379–402. Canberra: ANU Press.
- Sutikna, T., M.W. Tocheri, J.T. Faith, Jatmiko, R. Due Awe, H.J.M. Meijer, E. Wahyu Saptomo, and R.G. Roberts. 2018. “The Spatio-Temporal Distribution of Archaeological and Faunal Finds at Liang Bua (Flores, Indonesia) in Light of the Revised Chronology for Homo Floresiensis.” Journal of Human Evolution 124: 52–74. doi:10.1016/j.jhevol.2018.07.001.
- Szabó, K., and J.R. Amesbury. 2011. “Molluscs in a World of Islands: The Use of Shellfish as a Food Resource in the Tropical Island Asia-Pacific Region.” Quaternary International 239 (1–2): 8–18. doi:10.1016/j.quaint.2011.02.033.
- Tanudirjo, D.A. 2001. “Islands in Between: Prehistory of the Northeastern Indonesian Archipelago.” PhD diss., Australian National University.
- Tanudirjo, D.A. 2005. “Long-continuous or Short-Occasional Occupation? The Human Use of Leang Sarru Rockshelter in the Talaud Islands, Norteastern Indonesia.” Bulletin of the Indo-Pacific Prehistory Association 25 (1): 15–19.
- Teixeira, J.C., G.S. Jacobs, C. Stringer, J. Tuke, G. Hudjashov, G.A. Purnomo, H. Sudoyo 2021. “Widespread Denisovan Ancestry in Island Southeast Asia but No Evidence of Substantial Super-Archaic Hominin Admixture.” Nature Ecology & Evolution 2021 5:5 5 (5): 616–624. doi:10.1038/s41559-021-01408-0.
- Torgersen, T., M.F. Hutchinson, D.E. Searle, and H.A. Nix. 1983. “General Bathymetry of the Gulf of Carpentaria and the Quaternary Physiography of Lake Carpentaria.” Palaeogeography, Palaeoclimatology, Palaeoecology 41 (3–4): 207–225. doi:10.1016/0031-0182(83)90088-3.
- van den Bergh, G.D., J. de Vos, P.Y. Sondaar, and F. Aziz. 1996. “Pleistocene Zoogeographic Evolution of Java (Indonesia) and Glacio-Eustatic Sea Level Fluctuations: A Background for the Presence of Homo.” Bulletin of the Indo-Pacific Prehistory Association 14: 7–21. doi:10.7152/bippa.v14i0.11583.
- van den Bergh, G.D., Y. Kaifu, I. Kurniawan, R.T. Kono, A. Brumm, E. Setiyabudi, F. Aziz, and M.J. Morwood. 2016a. “Homo floresiensis-like Fossils from the Early Middle Pleistocene of Flores.” Nature 534 (7606): 245–248. doi:10.1038/nature17999.
- van den Bergh, G.D., B. Li, A. Brumm, R. Grün, D. Yurnaldi, M.W. Moore, I. Kurniawan 2016b. “Earliest Hominin Occupation of Sulawesi, Indonesia.” Nature 529 (7585): 208–211. doi:10.1038/nature16448.
- Van Der Kaars, S., and R. Dam. 1997. “Vegetation and Climate Change in West-Java, Indonesia during the Last 135,000 Years.” Quaternary International 37: 67–71. doi:10.1016/1040-6182(96)00002-X.
- Veth, P., I. Ward, T. Manne, S. Ulm, K. Ditchfield, J. Dortch, F. Hook 2017. “Early Human Occupation of a Maritime Desert, Barrow Island, North-West Australia.” Quaternary Science Reviews 168: 19–29. doi:10.1016/j.quascirev.2017.05.002.
- Wedage, O., N. Amano, M.C. Langley, K. Douka, J. Blinkhorn, A. Crowther, S. Deraniyagala 2019. “Specialized Rainforest Hunting by Homo Sapiens ~45,000 Years Ago.” Nature Communications 10 (1): 1–8. doi:10.1038/s41467-019-08623-1.
- Westaway, K.E., J. Louys, R.D. Awe, M.J. Morwood, G.J. Price, J.X. Zhao, M. Aubert 2017. “An Early Modern Human Presence in Sumatra 73,000–63,000 Years Ago.” Nature 548 (7667): 322–325. doi:10.1038/nature23452.
- Whittaker, R.J., and J.M. Fernández-Palacios. 2007. Island Biogeography. Oxford: Oxford University Press.
- Whittaker, R.J., K.A. Triantis, and R.J. Ladle. 2008. “A General Dynamic Theory of Oceanic Island Biogeography.” Journal of Biogeography 35 (6): 977–994. doi:10.1111/j.1365-2699.2008.01892.x.
- Wilson, S.M. 2007. Archaeology of the Caribbean. Cambridge: Cambridge University Press.
- Wright, D. 2011. “Mid Holocene Maritime Economy in the Western Torres Strait.” Archaeology in Oceania 46 (1): 23–27. doi:10.1002/j.1834-4453.2011.tb00096.x.
- Zhang, H., Y. Wang, C.M. Janis, R.H. Goodall, and M.A. Purnell. 2017. “An Examination of Feeding Ecology in Pleistocene Proboscideans from Southern China (Sinomastodon, Stegodon, Elephas), by Means of Dental Microwear Texture Analysis.” Quaternary International 445: 60–70. doi:10.1016/j.quaint.2016.07.011.