
SUMMARY
Dietary fibre has traditionally been considered an energy-diluting component of laying hen diets. With low energetic values, and sometimes negative impacts on digesta viscosity and gut function, formulations have often aimed to minimise crude fibre contents. Increasing fibre contents can mean that the level of fat required to meet nutritional standards must be increased to account for the decrease in energy, with consequential impacts on feed handling. By contrast, recent research has shown that some dietary fibres can have beneficial effects on laying performance, driven by changes in digestibility, gut structure and function, as well as shifts in the gut microbiota. However, there is often a lack of clarity as to the quantity and type of fibre required to yield production benefits. Broadly, soluble fibres – while largely detrimental to nutrient digestibility – can act as prebiotics, favouring beneficial intestinal microbiomes, while insoluble fibres stimulate intestinal development and may have some preventative effects on feather pecking and cannibalism.
Introduction
Eggs represent one of the most abundant and affordable sources of animal protein and are generally accepted by most cultures and religions (Bain, Nys, and Dunn Citation2016). The poultry industry has selectively bred laying hens to be capable of laying over 300 eggs a year. However, modern production practices pose numerous challenges for optimal bird health and productive output. Furthermore, the 2006 EU ban on non-therapeutic antibiotics adds to the challenge of maintaining flock health and welfare. The inclusion of additional dietary fibre is a strategy that has the potential to support multiple aspects of laying production. Historically, the inclusion of dietary fibre in poultry diets was considered to have a diluting effect on nutrient content, thought to cause poor nutrient utilisation (Jha et al. Citation2019). However, in recent years, there has been a change in attitude towards the industry recognising the value of certain types and quantities of fibre in poultry diets, and the use of dietary fibre has been revisited due to its potential benefits on gut health, gastrointestinal development, performance and bird welfare. Therefore, this review aims to summarise modern thinking on the use of dietary fibre in laying hens.
What is fibre?
Physiologically, dietary fibre can be defined as: the consumable component of plants or analogous carbohydrates (polysaccharides, oligosaccharides, lignin, and associated plant substances) that are resistant to digestion and absorption in the small intestine with complete or partial fermentation in the large intestine (AACC Citation2001). There are a number of types of fibre, as shown in , all of which can possess different functional properties – exerting positive, negative or neutral effects on the host’s digestive function (Jha et al. Citation2019). Approximate soluble and insoluble fibre contents of common feed ingredients used in reviewed studies is shown in .
Table 1. Types of fibre (adapted from The British Nutrition Foundation, 2020)
Table 2. Approximate insoluble and soluble fibre content of common feed materials
Fibre composition and chemistry have been thoroughly reviewed elsewhere (Yangilar Citation2013; Fuller et al. Citation2016), and this is not the scope of this paper. Briefly, components of fibre can be either soluble (SF) or insoluble (IF). Soluble fibres can be readily fermented by gut microbes in the hind gut; however, they have also been shown to increase the viscosity of digesta (Cameron-Smith, Collier, and O’dea Citation1994; Choct Citation1997). Increased viscosity reduces passage rate due to increased satiety and ultimately reduces overall feed intake (Jha et al. Citation2019). Reduction of viscosity caused by dietary soluble fibre is the traditional reason for application of xylanases to poultry diets (Choct et al. Citation2004; Kiarie, Romero, and Ravindran Citation2014). Insoluble fibre is poorly fermented and passes through the tract mainly undigested. This increases the passage rate and faecal bulking and can result in increased feed intake by the host (Jha and Berrocoso Citation2015).
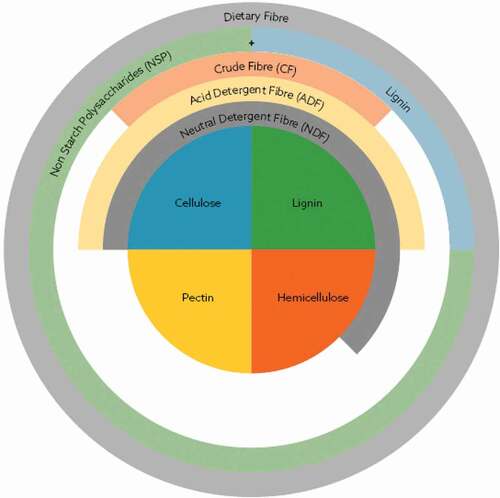
Fibre as a nutrient is difficult to define and there are many terms used in animal nutrition to describe and measure fibre components. Crude fibre quantification came into existence following the development of proximate analysis in Germany in the 19th century. This method separates carbohydrates into two sections of either more digestible (nitrogen free extract) or less digestible (crude fibre) (Henneberg and Stohmann Citation1859, Citation1860). Crude fibre is defined as the organic remnants of feed following immersion in hot sulphuric acid and sodium hydroxide and is composed of some insoluble non-starch polysaccharides (NSP), cellulose and some hemi-cellulose (). Whilst this method has been in use for many years, it has innate flaws, such as not accounting for any of the soluble NSP content (Choct Citation2015). Due to this flaw, the neutral detergent (NDF) and acid detergent fibre methods (ADF) were developed (Van Soest Citation1963). This method is generally agreed to well quantify the levels of NSP in cereal-based raw materials; however, there is a lack of consensus for oilseeds and legumes, due to the levels of soluble NSP, such as arabinose, galacturonans, and arabinogalactans, that the NDF analysis fails to account for (Choct, Citation2015). Despite the values of this method, from a chemistry perspective, it fails to determine any specific fibre components.
Figure 1. Fibre composition. Adapted from Choct, Citation2015
In recent years, there has been a significant increase in research output surrounding fibre in poultry diets, and the postulation that dietary fibre may have beneficial effects. Both insoluble and soluble fibres have been shown to support various parameters of poultry production and improvements in performance, egg production, nutrient digestibility, behaviour and welfare have all been cited suggesting that fibre may be a viable, broad spectrum tool for successful production (Mateos, Lazaro, and Gracia Citation2002; Incharoen and Maneechote Citation2013). An overview of the general effects is shown in , and a summary of the literature and reported effects is found in .
Table 3. The effects of dietary soluble and insoluble fibre in the gastrointestinal tracts of laying hens
Table 4. Summary of published literature assessing the effect of dietary fibre on the performance, gut health and feather pecking in laying hens
Effects of dietary fibre on laying hen performance and egg quality
The performance effects of fibre inclusion have been varied and not always consistent. This is likely due to the varying quantities and types of fibre investigated, making comparisons difficult.
During the pullet stage (8–18 weeks) the use of insoluble fibre has been shown to be beneficial for GIT development and subsequent performance (Abdallah and Beshara Citation2015; Panaite et al. Citation2016). Panaite et al. (Citation2016) studied the effect of pelleted alfalfa at 0%, 4%, 8% and 19.44% on Lohmann Brown hybrid pullets from 9 to 16 weeks of age. Results showed that pullets supplemented with 8% alfalfa had significantly increased liveweights compared to the control at the end of the study. Furthermore, the 8% treatment group achieved liveweights that were 6.73% higher than the Lohmann Brown management guidelines. Abdallah and Beshara (Citation2015) found similar results. Supplementing pullets with sunflower meal (7% and 14%), olive cake (5% and 10%) and a combination of sunflower and olive cake (7% sunflower and 14% olive cake) compared to a control (3.5% CF) from 11 to 19 weeks, followed by feeding all birds a control diet from week 20 to 40. The results showed that increasing CF by up to 6% during the pullet phase resulted in significantly improved liveweight and FCR compared to the control. CF digestibility was also significantly increased for all fibre treatments compared to the control. The study found that increasing the CF significantly increased the gizzard and caeca weights compared to the control, suggesting that improved gizzard development and function may be a possible mechanism for improved growth. This study went on to look at the subsequent egg production performance and found that increasing the level of fibre in the diet significantly increased egg numbers, laying rate % and egg mass compared to the control birds, potentially suggesting ongoing benefits of fibre in the growing phase diets.
Guzmán et al. (Citation2015) investigated the effect of supplementing laying pullets with sunflower hulls, cereal straw or sugar beet pulp at either 0%, 2% or 4% from hatching to 5 weeks of age, and demonstrated significant improvements in ADG, FI and energy efficiency. Furthermore, FCR was significantly improved when fibre was supplemented at 2% rather than 0% or 4%, suggesting that there may be a maximum inclusion rate to gain benefits from supplemental fibre. Yokhana, Parkinson, and Frankel (Citation2016) found that a commercial insoluble fibre, fed at 1% to 8-week-old pullets, increased body weight gain compared to the control. Sozcu and Ipek (Citation2020) investigated the effect of lignocellulose at 0, 0.5, 1 and 2 kg/t feed on performance, and showed that 0.5 and 1 kg/t of lignocellulose significantly increased bodyweight, feed intake and feed efficiency compared to the control at all sampling points between 18 and 38 weeks of age. Interestingly, supplementation at 2 kg/t had no further beneficial effect; this is similar to the findings of Guzmán et al. (Citation2015), though the optimal dose differs between studies.
However, other studies have found either no effect or detrimental effects on hen bodyweight when supplementing dietary fibre. Panaite et al. (Citation2015) trialled numerous sources of fibre, including DDGS, rice bran wheat bran and sunflower meal, comparing the inclusion of these fibre sources at both a low (3.67% starter, 3.86% grower and 5.39% finisher) and high (4% starter, 5% grower, 7.24% finisher) levels. Inclusion of these fibres had no significant impact, indicating that there were no detrimental effects of feeding up to 7% of dietary fibre. Similarly, Walugembe, Rothschild, and Persia (Citation2014) found no significant differences in the feed intake or average daily gain of layer chicks supplemented with a 60 g/kg (6%) mix of DDGS and wheat bran compared to a control. Zdunczyk et al. (Citation2019) saw no effect of increasing fibre levels when substituting lupins for soybeans in isocalorific diets, and Röhe et al. (Citation2019) report that a 10% addition of lignocellulose resulted in significantly decreased bodyweight in dual-purpose birds compared to a standard commercial diet. However, this study also revealed a significant improvement in the number of eggs laid, which was also correlated with significantly improved feed conversion efficiency.
Increased egg production following fibre supplementation was also seen with lignocellulose by Sozcu and Ipek (Citation2020), where 0.5 and 1 kg/t of lignocellulose (IF, NDF 60–90%) resulted in significantly increased egg production; 2 kg/t was unable to change performance versus the unsupplemented control. Röhe et al. (Citation2020) also reported some egg production benefits when supplementing dual-purpose birds (Lohmann Dual) with 10% lignocellulose (IF, NDF 60–90%) and showed that the addition of lignocellulose resulted in significantly increased egg production. Likewise, shell breaking strength and thickness were numerically increased in the treatment group. However, the eggs produced by the control were significantly larger and heavier than the treated group. The authors suggest that the results seen may be related to bird weight, as the treatment group weighed significantly less than the control and it has been demonstrated that egg weight is directly related to bird weight. It is of note that the experimental birds were dual purpose, and therefore ingested energy was divided between muscle/fat deposition and egg production. Modern laying strains have been heavily selected over long periods of time to achieve maximum laying potential; it is plausible that the use of a dual-purpose bird complicates the laying response to fibre.
By contrast, De Arruda et al. (Citation2018) reported lower egg weights with increasing levels of insoluble fibre, while Kocer et al. (Citation2021) reported mixed findings when supplementing laying diets (from 21 weeks of age) with 3%, 4%, or 5% fibre (high fibre sunflower meal). In a 24-week study, the authors report that body weight, egg production rate, and shell mass were all linearly significantly increased by increasing fibre inclusion. Furthermore, the shell-less egg rate and cracked/broken egg rate were numerically reduced linearly with increased fibre inclusion. A typical layer requires 2.2 g of calcium for each egg laid – more than 60% of this is supplied in the diet, but the remainder is obtained via mobilisation of calcium from the medullary bone (Bouvarel Citation2011). Insufficient calcium absorption from the diet can contribute to the onset of osteoporosis. Therefore, provision and bioavailability of calcium are vital for both shell strength and bird welfare. Van der Aar et al. (Citation1983) reported that the inclusion of 8% lignin (IF, NDF 60–90%) resulted in reduced mineral absorption, consequently reducing cation-binding protein at the brush border of the small intestine. However, there is also evidence that the reduced intestinal pH often observed with increased dietary fibre, can result in increased mineral digestibility, possibly due to increased mineral solubility (Engberg et al. Citation2004; Yokhana, Parkinson, and Frankel Citation2016; Kim et al. Citation2019). The increase in shell mass described above by Kocer et al. (2020), as well as the increase in shell breaking strength described by Röhe et al. (2018), seem to refute the earlier findings of Van der Aar et al. (Citation1983); however, as the fibre sources and inclusion rates were different between studies, it is hard to compare fairly.
Sozcu and Ipek (Citation2020) also looked at egg quality parameters and found that once again the supplementation of 1 kg/t of lignocellulose resulted in significantly increased eggshell breaking strength, eggshell thickness, increased yolk index percentage and increased yolk ratio percentage compared to the other treatments. As with the previous results of this study, the 0.5 kg/t treatment demonstrated significant improvements compared to the control, but to a lesser degree. However, the 2 kg/t treatment in general only delivered statistically similar results to the 0.5 kg/t group and a linear dose response was not observed. Hassan, Morsy, and Hasan (Citation2013) investigated egg yolk cholesterol and production performance of layers fed 3%, 4%, 5% or 6% crude fibre in the form of alfalfa meal. This study found that layers fed 3% fibre laid significantly more eggs than the higher inclusion rates; however, the inclusion of 6% fibre resulted in significantly increased shell, albumin, and yolk weights compared to the other inclusion rates. Likewise, the cholesterol per yolk was significantly decreased linearly in line with increased fibre inclusion. This may indicate that including increased levels of dietary fibre may help to reduce the cholesterol in eggs, consequently improving the nutritional profile of eggs for the consumer.
There is a paucity of information on the effect of fibre on flock uniformity. Incharoen and Maneechote (Citation2013) looked at the effect of rice hulls at 0%, 3% and 6% as a form of insoluble fibre on flock uniformity of laying hens. A 28-week study found that rice hulls significantly improved flock uniformity. Conversely, Asensio et al. (Citation2020) found no significant effect of increasing fibre (Wheat bran) from 3% to 8.8% and reducing AMEn by 15% on flock uniformity. Whilst no detrimental effects were found, clearly more information is required to better understand this parameter.
Effects of dietary fibre on nutrient digestibility
Whilst it has been reported that nutrient digestibility is inversely proportional to the quantity of dietary fibre included in the ration (Jha et al. Citation2019), a number of studies have demonstrated positive effects when changing from diets lacking in fibre to formulations with significant fibre content. In some cases, the addition of fibre can result in improved AME (Hetland and Svihus Citation2001; Gonzales-Alvarado et al. Citation2007; Biggs and Parsons Citation2009). Likewise, Biggs and Parsons (Citation2009) report that feeding broilers 10–20% whole wheat resulted in improved AA digestibility. A number of studies have reported that the inclusion of 3–10% oat hulls results in increased digestibility of faecal starch, protein, and fat, as well as ileal starch (Rogel, Annison, and Bryden Citation1987; Hetland and Svihus Citation2001; Jimenez-Moreno et al. Citation2009). Conversely, Singh et al. (Citation2014) investigated the effect of replacing ground maize with 150, 300, 450 or 600 g/kg of coarse maize (10 mm sieve). The only effect observed between treatments was a significant increase in feed intake, and no differences in energy utilisation were seen.
The reasons for the improved utilisation of nutrients seen with the addition of insoluble fibre are likely to be multifactorial. It has been demonstrated that the inclusion of insoluble fibre decreases the particle size of digesta entering the small intestine. Hetland, Svihus, and Olaisen (Citation2010) investigated the effect of feeding whole grains (wheat, oats and barley) to broilers. The study measured duodenal particle size by laser diffraction and found that the moderate inclusion of whole cereals (300 g/kg) reduced the particle size in the duodenum significantly compared to the ground cereal control. Likewise, the high inclusion of whole grains at 440 g/kg reduced the particle size still further. This ultimately resulted in a significant increase in starch digestibility for the inclusion of both whole wheat and barley. The authors suggested a hypothesis for this is that the gizzard grinds diet components to a critical particle size before releasing them into the duodenum. It has also been suggested that some constituents of the diet may be encapsulated within large particles and, therefore, harder to degrade sufficiently for absorption by the bird. This theory is supported by the work of Péron et al. (Citation2007), who used scanning electron microscopy to elucidate the digestive process. This study was able to show starch granules trapped within larger particles in the small intestine. In contrast, another study by Péron et al. (Citation2005) demonstrated that fine grinding of a hard wheat variety that had generally shown low starch digestibility, resulted in increased starch digestibility. This may suggest that the process is more complicated than simply providing structural fibre to increase gizzard efficiency and that rather the provision of a ground diet with a degree of structural components may provide more benefit to digestibility.
Adibmoradi, Navidshad, and Jahromi (Citation2016) reported that the inclusion of rice hulls (IF87.3%, SF 2.7%, NDF ~60%) at both 0.75% and 1.5% significantly increased crude protein digestibility; improved gizzard function and pepsin secretion as a result of increased HCL production may be the mechanism for this improvement. This is in agreement with the results of Gonzales-Alvarado et al. (Citation2007), and Amerah, Ravindran, and Lentle (Citation2009). Similar results have been reported in pigs (Kirchgessner et al. Citation1994), where improvements were assigned to changes in the microbiome favouring N utilisation in the hind gut and thus shifting from urine N excretion to bacterial protein, lowering N volatilisation in the manure and improving environmental outcomes in both pigs and laying hens (Canh et al. Citation1998; Roberts et al. Citation2007). It is noted that much of the literature reviewed here is from broilers due to the paucity of layer data. Therefore, some discrepancies in the effect of adding fibre to layer diets may exist due to the variable composition and form of broiler diets.
It has been widely demonstrated that fibre inclusion can reduce the pH in the GIT. Jimenez-Moreno et al. (Citation2009) looked at the effect of cereal type on gizzard pH and nutrient utilisation in broilers. In this study, we compared the inclusion of 3% oat (IF83.3%, SF1.7%, NDF ~75%) or soy hulls (IF49.3%, SF13.3%, NDF 67%) to a control and showed that the inclusion of either oat or soy hulls improved the total tract apparent retention of nutrients and reduced the pH in the gizzard. The authors suggest that the reduced pH in the gizzard may be responsible for further degrading nutrients and to make them more available for subsequent absorption. Engberg et al. (Citation2004) found similar results and suggested the same mechanism of increased nutrient degradation in the gizzard. The reduced pH in the gizzard is likely to have further benefits for nutrient digestibility. Pepsin activity is stimulated by low pH (Yokhana, Parkinson, and Frankel Citation2016). Therefore, if fibre inclusion is reducing intestinal pH, it is plausible that this may be the mechanism for increased pepsin activity observed by Yokhana, Parkinson, and Frankel (Citation2016) in layers fed 1% insoluble fibre. Likewise, reduced gizzard pH has also been demonstrated to increase calcium digestibility (Kim et al. Citation2019). The reduction in pH improves the solubility of limestone in the diet, rendering increased calcium available for absorption. This mechanism may be beneficial for layer production, particularly in later lay when shell strength starts to diminish.
Svihus (Citation2011) suggests that there is a link between the proventriculus and gizzard, whereby the functionality of the gizzard can control the preceding secretion of HCl and pepsinogen in the proventriculus. The author theorises that when the gizzard is stimulated by the presence of fibre, the proventriculus can respond by increasing gastric secretions. This may therefore aid the further degradation of nutrients, increasing the availability of absorption.
Fibre has also been shown to influence digesta retention time, and therefore digestibility (Hetland, Svihus, and Olaisen Citation2002; Tejeda and Kim Citation2021). Sacranie et al. (Citation2012) demonstrated that the supplementation of 15% oat (IF 83.3%, SF 1.7%, NDF ~75%) and barley hulls (IF 20.3%, SF 9.8%, NDF 60–75%) resulted in a larger particle of dietary fibre leaving the gizzard. This reduced passage rate in the upper digestive tract and increased retro-peristalsis, therefore increasing exposure to HCl and digestive enzymes, and consequently increasing the digestibility of starch. However, Hetland et al. (Citation2002) suggests that the accumulation of insoluble fibre in the gizzard may trigger temporary satiety and could therefore reduce feed intake and subsequent growth and/or performance.
Effects of dietary fibre on gut development and function
The beneficial effects of dietary fibre inclusion (particularly insoluble fibre) on the development and function of the GIT are well reported in poultry (Hetland, Svihus, and Krogdahl Citation2003; Hetland, Svihus, and Choct Citation2005; Sacranie et al. Citation2012). It is likely that this stems from the physical/mechanical demands of the consumption of fibre. Sacranie et al. (Citation2012) reported that the typical gizzard content is higher in fibre than the contents of other parts of the intestinal tract, demonstrating that the fibre component of the diet is physically harder to grind. In the absence of teeth, poultry relies on the gizzard to grind digesta (Svihus Citation2011). The gizzard is comprised of myelinated smooth muscle and sits immediately after the proventriculus (glandular stomach). The body of the gizzard is composed of two thick, laterally opposed muscles and two smaller anterior and posterior muscles (Akester Citation1986). Together, these allow rotary and crushing movements upon contraction. Within the gizzard, the surface is coated in a thick layer of glycol-proteins that are hardened by the low pH environment that increase grinding ability (De Lima and Sasso Citation1985). The adequate function of the gizzard is essential as feed particles can only leave the gizzard when a critical size is reached (Clemens, Stevens, and Southworth Citation1975; Moore Citation1999). Likewise, feed must be adequately ground to optimise nutrient uptake, as unground feed particles will prevent enzyme access and subsequent degradation. However, increased particle size is only part of the answer. Whilst indeed, increasing particle size can increase gizzard size and function, it is the insolubility of the fibre that is the most active factor. Increasing pellet-size is likely to increase feed intake; however, if the pellet is mainly soluble, much of this structure will be lost, along with the potential beneficial effect during digestion. The entire gastrointestinal tract relies heavily on retro peristalsis and reflux. Due to the small size of the proventriculus (glandular stomach), feed retention tends to be low causing much of the gastric secretion activity to take place within the gizzard. The reflux effect allows insufficiently degraded feed to be refluxed back to the proventriculus for additional gastric secretions to be applied (Duke Citation1986). It is well established that feed particle size and structure have a significant effect on the development of both the proventriculus and the gizzard. Svihus (Citation2011) states that gizzard size can increase by up to 100% its original size with the addition of insoluble fibre and structural diet components, such as whole grains and wood shavings. Interestingly, the degree of weight change in the gizzard content is often far greater than that of size change, with the content weight often increasing two-fold (Svihus Citation2011). It is generally considered that increased gizzard size allows for additional feed intake and throughput, and therefore may be beneficial for growth performance.
The work of Yokhana, Parkinson, and Frankel (Citation2016) demonstrates this effect: when supplementing 8-week-old layers with 1% insoluble fibre (Commercial source), birds fed insoluble fibre had significantly increased organ weights for liver, gizzard, and small intestine compared to the control. GIT enzyme activities were also analysed, and again revealed that the inclusion of insoluble fibre resulted in significantly increased pepsin, trypsin, and chymotrypsin activities compared to the control. Similarly, Röhe et al. (Citation2020) saw increased gizzard weights following supplementation of 10% lignocellulose and Sacranie et al. (Citation2012) showed that the addition of 15% oat and barley hulls either coarsely or finely ground significantly increased gizzard weight and significantly reduced gizzard pH. Whilst both fine and coarse hull inclusions significantly increased gizzard weight compared to the control, the effect was not as pronounced for the fine ground treatment. This could suggest that the increased particle size is partly responsible for the benefits derived. This study revealed no significant differences in gut motility between treatments but did confirm complete reflux with the inert marker applied to the cloaca being detected in all parts of the GIT.
The lack of fibre provision may also be detrimental to bird health and downstream productivity. As already explained, it has been widely demonstrated that the provision of fibre improves the development and function of the proventriculus and gizzard. However, if fibre is not provided to serve this function, the proventriculus can be poorly developed affecting functionality in terms of gastric secretions (Gonzales-Alvarado et al. Citation2008; Jimenez-Moreno et al. Citation2009; Mateos et al. Citation2012) resulting in widening of the gastric isthmus and subsequent rupture of the proventriculus during processing, resulting in increased condemnations (birds used for meat products, stock and soups) and financial loss (Dormitorio, Giambrone, and Hoerr Citation2007; Svihus Citation2011; Mateos et al. Citation2012).
In addition to the upper GIT, fibre may also influence the architecture of the small intestine. Sozcu and Ipek (Citation2020) investigated the effect of lignocellulose (at 0, 0.5, 1 and 2 kg/t) on the histomorphological traits of layers in peak lay and showed that birds supplemented with 2 kg/t of lignocellulose had significant longer jejunal villi compared to other treatments. This result is interesting as the 1 kg/t treatment performed best in terms of performance, and 2 kg/t tended to cause negative or negligible results in comparison. Similarly, Röhe et al. (Citation2020) saw that the addition of 10% lignocellulose had a larger villus absorptive area, longer villi and shorter crypts. Zdunczyk et al. (Citation2019) saw that increased crude fibre from lupin inclusion resulted in increases in both duodenal and jejunal mucosa thickness, crypt depth and villus height. Conversely, Sadeghi, Toghyani, and Gheisari (Citation2015) investigated the effect of 3% sugar beet pulp rice hulls or a 3% combination of both compared to a control in a broiler trial. This trial showed that the inclusion of sugar beet pulp significantly reduced duodenal and ileal villi heights compared to the control at 21 days of age. This may suggest that the type, inclusion rate and form of the fibre used may influence its effects on gut development and function. Interestingly, the inclusion of sugar beet pulp (Sadeghi, Toghyani, and Gheisari Citation2015) also significantly increased the antibody titre against Newcastle Disease.
Effects of dietary fibre on the intestinal microbiome
The microbiome can be affected by innumerable factors, including diet, breed, environment and genetics (Shang et al. Citation2018). However, diet is considered one of the most potent regulating factors (Mahmood and Guo Citation2020). Dietary fibres can contain varying quantities of non-starch polysaccharides (NSPs) which are complex carbohydrates derived from the structural components of plant cell walls (Mahmood and Guo Citation2020). NSPs have been widely shown to have detrimental effects on poultry performance, mainly associated with the increased viscosity of digesta as they absorb water and form viscous gels (Bedford Citation2018). Consequently, the use of NSP degrading exogenous enzymes is common in the industry. Degradation of NSPs is thought to depolymerise polysaccharide components into oligosaccharides (SF) which are considered prebiotics due to their ability to enhance beneficial bacteria in the GIT (Ding et al. Citation2018; Li et al. Citation2017). This prebiotic shift following supplementation with either oligosaccharides or NSP-degrading enzymes is well documented, and is shown to work through ‘cross-feeding’ mechanisms that favour mutualistic microbial populations and ultimately upregulate the production of butyrate and other SCFAs in the gut (De Maesschalck et al. Citation2015; Rivière et al. Citation2015).
Nutrients that remain undigested upon reaching the hind gut (distal small intestine and large intestine) can be utilised as a vital nutrient source for commensal bacteria. The commensal bacteria play a role in immune defence by stimulating the development of the intestinal mucus layer, epithelial monolayer and intestinal immune cells. Furthermore, the provision of fermentable substrate for the microbiota results in the production of microbial metabolites. Metabolites produced can have a variety of beneficial functions, such as antimicrobial and anti-inflammatory properties and the production of short-chain fatty acids (SCFAs), such as butyric and propionic acids (Shang et al. Citation2018). Butyrate is the primary energy source for enterocytes, allowing cells to be renewed and therefore helping to prevent intestinal permeability (leaky gut) (Jha et al. Citation2019). Likewise, butyrate encourages growth of beneficial bacteria, contributes towards optimal immune function and also plays a role in brain function (Belkaid and Hand Citation2014; Liu et al. Citation2018).
The huge variety of chemical structure of dietary fibre is likely to have equally variable effects on the resident bacteria: while Wanzenböck et al. (Citation2020) saw no significant effects of native or fermented wheat bran on the microbiome of laying hens, Ding et al. (Citation2018) studied the effects of dietary xylo-oligosaccharides (SF) (XOS, at 0, 0.01, 0.02, 0.03, 0.04 and 0.05%) on the gut microbiota of laying hens and saw a significant increase in the numbers of Bifidobacteria in the caeca (linear increase in line with inclusion rate). Furthermore, XOS inclusion significantly increased the digesta content of butyrate and tended to increase acetic acid in the caeca (again linear increase with inclusion rate). This may suggest that provision of only a very small insoluble fibre component may be beneficial.
Conversely, the provision of excess undigested feed can contribute to dysbiosis and the proliferation of pathogenic bacteria (Shang et al. Citation2018; Mahmood and Guo Citation2020). Dysbiosis has been reported to reduce intestinal barrier function (leaky gut), reduce nutrient digestibility and increased likelihood of systemic translocation of luminal bacteria (Shang et al. Citation2018). This may suggest that there is an optimal inclusion level of fibre to derive maximum benefits – as seen previously when assessing the effects of fibre on performance – and that the provision of excess fibre may be detrimental to the bird. Furthermore, the increased viscosity, which has been well described in the literature as a result of NSPs in the diet, has also been shown to have a detrimental effect on the proliferation of potentially pathogenic bacteria in the hind gut. Johnson and Gee (Citation1981) state that the increased viscosity of digesta causes the thickness of the unstirred water layer that covers the mucosa cells, to increase. In turn, this limits contact between nutrients and gastrointestinal secretions (pancreatic enzymes and bile acids), resulting in increased quantities of undigested protein and starch reaching the hindgut and acting as a substrate for proliferation of potentially pathogenic bacteria (Bedford and Cowieson Citation2012). This effect has been described by Shakouri et al. (Citation2006), who found that 3% supplementation of methylated citrus pectin resulted in increased colonisation of Enterobacteriacaeca spp. in the caeca of broilers. Baurhoo et al. (Citation2007) found similar results demonstrating that supplementing purified lignin at 2.5% resulted in inhibition of beneficial bacteria, such as Lactobacilli and Bifidobacteria in the caeca. It is of note that neither of these studies used an NSP degrading enzyme, so it is unclear whether this may mitigate the problem.
Effects of dietary fibre on excreta consistency and litter quality
Due to the evolutionary adaptation of birds to excrete both solid and liquid waste in one combined movement, maintaining litter quality can be a difficult task. Poultry excreta can contain approximately 80% moisture (Henuk and Dingle Citation2003). In broiler production, this can become problematic, leading to wet litter and numerous problems, including pododermatitis, breast blisters/scald, hock burn, and poor air quality (Dunlop et al. Citation2016; Kheravii et al. Citation2018)
However, in laying hens, excreta consistently is more problematic in terms of dirty eggshells and contaminated plumage. Dirty eggs are problematic for the laying industry due to downgrades at processing that can be costly to producers. The effect of fibre on excreta consistency seems to depend largely on the type and quantity of fibre used. A number of studies have reported that provision of dietary soluble NSPs can significantly increase the incidence of water and sticky droppings, due to the increased viscosity of content (Choct and Annison Citation1992; Jimenez-Moreno et al. Citation2013). However, other studies – particularly those looking at insoluble fibre – have shown that the provision of dietary fibre can significantly improve excreta consistency, consequently improving litter quality. Rezaei, Karimi Torshizi, and Rouzbehan (Citation2011) investigated the effect of feeding 0%, 0.3%, 0.4% and 0.5% of micronised insoluble fibre (commercial source) on litter moisture in broilers; there was a linear reduction in litter moisture with the increased inclusion of micronised insoluble fibre. The improvements observed with insoluble fibre may be due either to an effect on the mucosal layer of the intestine, allowing increased absorption of nutrients and water from the digesta, or that the micronised insoluble fibre itself, as the material has a high water-holding capacity (up to 4–8 times its weight). Similarly, Farran, Pietsch, and Chabrillat (Citation2013) found that litter moisture could be reduced by 10% with the addition of either 0.8% wheat bran or lignocellulose, while Kheravii et al. (Citation2017) also found significantly reduced litter moisture with the supplementation of 1% and 2% lignocellulose. Kheravii et al. (Citation2017) suggest two potential mechanisms for this: firstly the reduced litter moisture may be due to the increased water-holding capacity of the feed material, consequently reducing the moisture in the excreta, or alternatively, the increased production of SCFAs as a result of increased fermentation of fibre fractions in the hind gut may be increasing intestinal enterocyte proliferation, consequently increasing the surface area available for water resorption. However, as both lignin and cellulose are insoluble, the degree of fermentation may be questionable. Whilst this literature is mainly on broilers, the principle of improved excreta quality is likely to have beneficial effects on the occurrence of dirty shells. Higher moisture levels in the excreta can increase the propensity of plumage and nests becoming contaminated, thus increasing dirty shells. Soluble fibres such as β-glucan and xylan that have been shown to increase the viscosity of digesta, can actually have a negative impact on excreta consistency. Increased viscosity has been reported to increase thirst, consequently causing birds to increase their water intake, and moisture content of the excreta (Choct Citation1997Kalmendal and Wall Citation2012). However, the provision of insoluble fibres such as lignin and cellulose has been shown to increase the structure of manure (Kalmendal and Wall Citation2012; Johansson Citation2014).
Effects of dietary fibre on feather pecking and cannibalism
Feather pecking and cannibalism are abnormal behaviours of poultry that can cause significant welfare, health and productivity problems for the industry (Hartini et al. Citation2002; Van Krimpen et al. Citation2009). Nutritional approaches to preventing these issues have been extensively reviewed elsewhere (Rodenburg et al. Citation2013; Mens, Van Krimpen, and Kwakkel Citation2020) As far back as 1940, the use of dietary fibre was cited as an effective strategy for preventing cannibalistic behaviour (Bearse, Miller, and McClary Citation1940). However, there is also a significant weight of modern research to confirm this (Hartini et al. Citation2002; Van Krimpen et al. Citation2009). Feather pecking is generally considered a redirected foraging behaviour, antagonised by the lack of need to forage for food due to commercial feed being available ad libitum (Blokhuis Citation1986; Huber-Eicher and Wechsler Citation1997; Steenfeldt, Kjaer, and Engberg Citation2007; Albiker and Zweifel Citation2017). The provision of increased fibre (generally insoluble fibre) as a source of roughage has been widely shown to reduce feather pecking and/or cannibalistic behaviours (Bearse, Miller, and McClary Citation1940; Scott, Holm, and Reynolds Citation1954; Seemann Citation1982; Wahlström, Tauson, and Elwinger Citation1998;; Steenfeldt, Kjaer, and Engberg Citation2007; Albiker and Zweifel Citation2017). This effect is generally attributed to increased time spent eating, thus reducing redirected behaviours (Hartini et al. Citation2002; van Krimpen et al. Citation2008; Walser Citation1997). Furthermore, the act of actually reducing the metabolisable energy, by diluting the diet with fibre has been demonstrated to reduce stereotypic behaviours by encouraging birds to spend extra time eating in order to intake sufficient nutrients (Hartini et al. Citation2002; van Krimpen et al. 2008).
When considering the effect of various fibrous raw materials, it has been reported that the inclusion of oats seems to have more beneficial effects on cannibalistic behaviours than wheat (Bearse, Miller, and McClary Citation1940; Scott, Holm, and Reynolds Citation1954; Seemann Citation1982; Wahlström, Tauson, and Elwinger Citation1998). This may be due to the increased level of crude fibre present, as Abrahamsson et al. (Citation1996) found that diets high in barley were more effective than diets high in wheat. However, this effect has also been reported to vary with strain in birds, with some studies finding no effect (Karlson Citation1996).
Bearse, Miller, and McClary (Citation1940) investigated the effect of low (3% crude fibre) and high (11–12%) fibre diets on the respective damage to the integument and found that the birds receiving the lower fibre diets suffered a higher incidence of integument injury. The study went on to add oat hull water extract, oat hull ash or sodium silicate to layer diets to try and determine how the oat hulls were helping to prevent injury. However, this part of the study revealed no reduction in integument injuries, suggesting that the benefit is derived from fractions other than those investigated. The work of Qaisrani, Van Krimpen, and Kwakkel (Citation2013) reports that the use of nutrient diluted diets (7.5% diluted sunflower seed extract (SF) and oat hulls (IF, NDF, NDF 75%) 15% diluted, sunflower seed extract and 15% diluted with oat hulls) resulted in a significant reduction in feather damage in line with the increased dilution percentage and that oat hulls were more effective in preventing feather damage than sunflower seed extract. Van Krimpen et al. (Citation2009) are in agreement, finding that the use of diluted by 15% resulted in birds compensating for this by directly increasing feed intake. Consequently, a significant and sustained reduction in feather pecking behaviour was observed, although not completely mitigated.
In contrast, Van Krimpen et al. (Citation2009), when supplementing ISA brown layers with either high or normal NSP levels (13.3% or 19.5%) combined with normal and low-energy diets (2540 or 2825 kcal ME/kg) found no significant differences in the feather scores or culling rates due to cannibalism between treatments. The results of these two studies would seem to indicate that the type of fibre used is vital for improvements to be observed. In general, it appears that insoluble fibre is more effective in this parameter.
The form in which the feed is provided may also be important. Bearse et al. (Citation1940) observed that high fibre diets were more effective in reducing cannibalistic behaviours when fed as a mash rather than pelleted. It seems plausible that this is due to the increased time required to ingest mash as opposed to pellets.
The effect of fibre on layer mortality is generally associated with feather pecking and cannibalistic behaviours (Hartini et al. Citation2002; Wahlström, Tauson, and Elwinger Citation1998; Hartini and Choct Citation2011; Walugembe, Rothschild, and Persia Citation2014). Hartini et al. (Citation2002) reported reduced rates of mortality when feeding high levels of soluble fibre (Barley); however, mortality was reduced still further when fibre was used in conjunction with beak trimming. Wahlström, Tauson, and Elwinger (Citation1998) report that increasing the quantity of crude fibre in laying diets from 44 to 64 g/kg resulted in a 31% reduction in mortality. This reduction was also accompanied by a significant reduction in traumatic skin wounds and so is assumed to be related more to the effect of fibre on feather pecking behaviours, rather than a direct effect on mortality. Hartini and Choct (Citation2011) found no significant difference in mortality between fibre sources in an experiment comparing wheat, guar gum and lucerne diets containing identical crude fibre percentages.
The effect of adding additional fibre to diet formulations
Typically, the addition of fibre to poultry diets has been thought to have detrimental effects on performance and the utilisation of nutrients, due to the low metabolisable energy content of ingredients with high fibre content (Lee et al. Citation2003). A traditional peak-lay (26–50 weeks) diet contains approximately 2–3% crude fibre. Additional fibre in the diet generally results in an energy deficit; this is often compensated by the inclusion of additional oil. Unfortunately, mash diets containing more than 4% oil are highly likely to have handling problems due to the high moisture content. An example of formulations containing increasing quantities of fibre is shown in .
Table 5. Effect of increasing the level of dietary fibre on example feed formulations for laying hens in peak lay
It is of note that laying hens are generally regarded to have good capability to increase feed intake when diets are diluted; however, considerations have to be made for the potential extra costs of feed processing and transportation of diluted diets. To avoid the use of high quantities of soya oil, the increased use of oil seeds such as sunflower and rapeseed can be used. Whilst this solution still results in increased crude fat inclusion, the handleability of the diet is improved. Diets containing high levels of oil are thought to be typically well tolerated by layers (Kleyn Citation2013); however, interactions between fats and dietary fibre have been reported. Galbraith and Miller (Citation1973) showed that lipids can reduce the microbial degradation of fibre by forming a layer on the surface of the fibre matrix. When the oil level is increased (particularly with soya oil) the linoleic acid level is also likely to increase; increased linoleic acid has been associated with increased egg weight (Grobas, Mateos, and Mendez Citation1999a), and at maximum levels of 3.45% are unlikely to be problematic to production.
Increasing the fibre content of diets may also be associated with an increase in NSPs (), which has been shown to increase the viscosity of digesta (Choct Citation1997). Increased digesta viscosity has been associated with reduced digestibility of various nutrients, including lipids (Smits Citation1996; Choct Citation1997). This indicates the need for an NSP degrading enzyme to be added to the diet to counteract the antinutritional factors associated with higher levels of NSPs. Adding NSP degrading enzymes may also have the benefit of reducing the amount of oil required in high fibre diets. The addition of a typical NSP degrading enzyme to wheat or barley can increase the AME by 3–6% (Lázaro et al. Citation2003; Mirzaie et al. Citation2012), thus reducing the energy deficit from adding fibre.
Summary
Despite the historical aversion to the inclusion of dietary fibre in poultry diets, there is increasing evidence that the use of some forms and quantities of fibre may be beneficial to the host. Primary benefits include enhanced nutrient digestibility, improved gut health and development, reduced cannibalistic behaviours, and increased hind gut fermentation of undigested substrate. However, the type, form and quantity of dietary fibre used appears to be a crucial factor, with excess amounts potentially counteracting any beneficial effects.
The main factor to determine is what effects are desired and, therefore, what type of fibre may induce this effect. In the literature reviewed, soluble fibre may have significant effects on the microbiome, resulting in increased SCFA production, potentially producing an energy sparing and antimicrobial effects. The inclusion of insoluble forms of fibre appears to have beneficial effects on excreta consistency, egg hygiene, litter quality, increased GIT passage rate and reduced feather pecking and cannibalism. Both soluble and insoluble forms of fibre seem to play a role in enhancing digestibility of nutrients. There is extensive evidence that the inclusion of insoluble fibre can help improve gizzard development and function, often resulting in reduced GIT pH, allowing increased degradation of nutrients, thereby making additional nutrients available for absorption. These changes suggest that the use of increased quantities of both insoluble and soluble fibre, may have the potential to support increased persistency of lay, where issues such as poor intestinal health, reduced eggshell strength and long-term welfare issue resulting from feather pecking and cannibalism combine to reduce the length of laying cycles.
There is a paucity of literature looking at the effect of fibre in free-range systems, so it remains unclear whether similar effects could be expected in that setting. Likewise, few studies looking at uniformity were found. As such, further research in these areas would be valuable to obtain a holistic view of the subject.
There does seem to be a clear maximum inclusion rate, with many studies reporting detrimental effects when fibre inclusion reaches levels over ~10%, with most benefits seeming to be found with inclusion rates of around 5–7%. With the continuing cost increases of raw materials and trend towards reduced meat prices, it is unsurprising that identifying opportunities to reduce production costs is an industry priority. The use of a limited quantity of fibre has been demonstrated to be beneficial to both bird health and production performance. Typically, sources of fibre are inexpensive and therefore the potential is there to reduce production costs. Likewise, the benefits of fibre inclusion appear to support the maintenance of a healthy microbiome. As such, there is additional potential for the use of increased fibre inclusion to support efficient and profitable production.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Notes on contributors
Alexandra Desbruslais
Alexandra Desbruslais got her PhD at Nottingham Trent University, where she focused on poultry nutrition and oxidative stress. Today, Alexandra's research interests are broad, including sustainable animal production, oxidative reactions in feed and production animals and fundamental monogastric nutrition.
Alexandra Wealleans
Alexandra Wealleans gained her PhD from the University of Reading, where she focused on the nutrition of pigs in alternative husbandry systems. Since then, her research focus has been primarily on improving nutrient digestion and absorption of monogastrics, and the ways in which this interacts with their health and performance.
David Gonzalez-Sanchez
David Gonzalez Sanchez received his degree in Agronomic engineering and MSc in poultry nutrition from the department of Animal Production of Polytechnic University of Madrid. He worked as poultry and livestock feed formulator, feed ingredient purchaser and nutritionist for a multinational feed company and later on being involved in technical support positions for several companies in the feed additive industry. His main research activities and interests have been oriented to the role of fiber in poultry diets, carbohydrase enzymes and feed formulation.
Mauro di Benedetto
Mauro Di Benedetto received his degree in Veterinary Medicine from the department of Animal Nutrition of Bologna University. He worked as veterinarian and nutritionist in livestock production primarily giving technical assistance in the animal nutrition and animal health area. In the last years his main research and application interest is dedicated to the link between nutrition and health and its reflection on animal welfare, both in poultry and swine.
References
- AACC. 2001. “Cereal Foods World.” 46, 112–126.
- Abdallah, A. G., and M. M. Beshara. 2015. “Effect of Different Levels and Sources of Dietary Fibre on Productive and Economic Performance in Local Laying Hens during Growing Period and Subsequent Laying Performance.” Egyptian Poultry Science Journal 35: 367–398.
- Abrahamsson, P., R. Tauson, K. Elwinger. 1996. “Effects on Production, Health and Egg Quality of Varying Proportions of Wheat and Barley in Diets for Two Hybrids of Laying Hens Kept in Different Housing Systems.” Acta Agriculturae Scandinavica, Section A - Animal Science 46(3): 173–182.
- Adibmoradi, M., B. Navidshad, and M. F. Jahromi. 2016. “The Effect of Moderate Levels of Finely Ground Insoluble Fibre on Small Intestine Morphology, Nutrient Digestibility and Performance of Broiler Chickens.” Italian Journal of Animal Science 15: 310–317. doi:https://doi.org/10.1080/1828051X.2016.1147335.
- Akester, A. R. 1986. “Structure of the Glandular Layer and Koilin Membrane in the Gizzard of the Adult Domestic Fowl (Gallus Gallus Domesticus).” Journal of Anatomy 147: 1–25. PMC1261543.
- Albiker, D., and R. Zweifel. 2017. “Performance, Behaviour and Plumage of Brown Layers with Intact Beaks”. 21st European Symposium on Poultry Nutrition, Spain
- Amerah, A. M., V. Ravindran, and R. G. Lentle. 2009. “Influence of Insoluble Fibre and Whole Wheat Inclusion on the Performance, Digestive Tract Development and Ileal Microbiota Profile of Broiler Chickens.” British Poultry Science 50 (4): 366–375. doi:https://doi.org/10.1080/00071660902865901.
- Asensio, X., N. Abdelli, J. Piedrafita, M. D. Soler, and A. C. Barroeta. 2020. “Effect of Fibrous Diet and Vitamin C Inclusion on Uniformity, Carcass Traits, Skeletal Strength, and Behaviour of Broiler Breeder Pullets.” Poultry Science 99: 2633–2644. doi:https://doi.org/10.1016/j.psj.2020.01.015.
- Bailoni, L., S. Schiavon, G. Pagnin, F. Tagliapietra, and M. Bonsembiante. 2005. “Quanti-Qualitative Evaluation of Pectins in the Dietary Fibre of 24 Foods.” Italian Journal of Animal Science 4: 49–58. doi:https://doi.org/10.4081/ijas.2005.49.
- Bain, M. M., Y. Nys, and I. C. Dunn. 2016. “Increasing Persistency in Lay and Stabilising Egg Quality in Longer Laying Cycles, What are the Challenges?” British Poultry Science 57 (3): 330–338. doi:https://doi.org/10.1080/00071668.2016.1161727.
- Baurhoo, B., L. Phillip, C. A. Ruiz-Feria. 2007. “Effects of Purified Lignin and Mannan Oligosaccharides on Intestinal Integrity and Microbial Populations in the Ceca and Litter of Broiler Chickens.” Poultry Science 86(6): 1070–1078. doi:https://doi.org/10.1093/ps/86.6.1070
- Bearse, G. E., V. L. Miller, and C. F. McClary. 1940. “The Cannibalism Preventing Properties of the Fiber Fraction of Oat Hulls.” Poultry Science 18: 210–215. doi:https://doi.org/10.3382/ps.0190210.
- Bedford, M., and A. Cowieson. 2012. “Exogenous Enzymes and Their Effects on Intestinal Microbiology.” Animal Feed Science and Technology 173: 76–85. doi:https://doi.org/10.1016/j.anifeedsci.2011.12.018.
- Bedford, M. R. 2018. “The Evolution and Application of Enzymes in the Animal Feed Industry: The Role of Data Interpretation.” British Poultry Science 59 (5): 347–365. doi:https://doi.org/10.1080/00071668.2018.1484074.
- Belkaid, Y., and T. Hand. 2014. “Role of the Microbiota in Immunity and Inflammation.” Cell 157 (1): 121–141. doi:https://doi.org/10.1016/j.cell.2014.03.011.
- Biggs, P., and C. M. Parsons. 2009. “The Effects of Whole Grains on Nutrient Digestibilities, Growth Performance, and Cecal Short-chain Fatty Acid Concentrations in Young Chicks Fed Ground Corn-soybean Meal Diets.” Poultry Science 88: 1893–1905. doi:https://doi.org/10.3382/ps.2008-00437.
- Blokhuis, H. J. 1986. “Feather-pecking in Poultry: Its Relation with Ground-Pecking.” Applied Animal Behaviour Science 16: 63–67. doi:https://doi.org/10.1016/0168-1591(86)90040-7.
- Bouvarel, I. 2011. “Hen Nutrition for Sustained Egg Quality.” In Improving the Safety and Quality of Eggs and Egg Products, Vol 1: Egg Chemistry, Production and Consumption, edited by Y. Nys, M. Bain, and F. Vanimmerseel. Cambridge: Woodhead Publishing.
- Cameron-Smith, D., G. R. Collier, and K. O’dea. 1994. “Effect of Soluble Dietary Fibre on the Viscosity of Gastrointestinal Contents and Acute Glycaemia Response.” British Journal of Nutrition 71 (4): 563–571. doi:https://doi.org/10.1079/BJN19940163.
- Canh, T. T., A. L. Sutton, A. J. A. Aarnink, M. W. A. Verstegen, J. W. Schrama, and G. C. M. Bakker. 1998. “Dietary Carbohydrates Alter the Fecal Composition and pH and the Ammonia Emission from Slurry of Growing Pigs.” Journal of Animal Science 76: 1887–1895. doi:https://doi.org/10.2527/1998.7671887x.
- Chaplin, M. F. 2003. “Fibre and Water Binding.” Proceedings of the Nutrition Society 62: 223–227. doi:https://doi.org/10.2527/1998.7671887x.
- Choct, M. 1997. “Feed Non-starch Polysaccharides: Chemical Structures and Nutritional Significance.” Feed Milling International, 1997 (6): 13–26.
- Choct, M. 2015. “Feed Non-starch Polysaccharides for Monogastric Animals: Classification and Function.” Animal Production Science55(11): 1360–1366.
- Choct, M., A. Kocher, D. L. E. Waters, D. Pettersson, and G. L. Ross. 2004. “A Comparison of Three Xylanases on the Nutritive Value of Two Wheats for Broiler Chickens.” British Journal of Nutrition 92 (1): 53–61. doi:https://doi.org/10.1079/BJN20041166.
- Choct, M., and G. Annison. 1992. “The Inhibition of Nutrient Digestion by Wheat Pentosans.” British Journal of Nutrition 67: 123–132. doi:https://doi.org/10.1079/BJN19920014.
- Clemens, E. T., C. E. Stevens, and M. Southworth. 1975. “Sites of Organic Acid Production and Pattern of Digesta Movement in the Gastrointestinal Tract of Geese.” Journal of Nutrition 105: 1341–1350. doi:https://doi.org/10.1093/jn/105.10.1341.
- De Arruda, M. B. R., M. E. Borges, M. V. A. Santos, S. L. Santos Faustino, M. Jesus Teodoro, G. Cassio Souza, C. G. Silva Salles, and S. A. Barbosa. 2018. “Fiber in Laying Poultry Diets and Its Effects on Egg Quality.” 55ª Reunião Anual da Sociedade Brasileira de Zootecnia, 28º Congresso Brasileiro de Zootecnia, 27 a 30 de agosto de 2018. Goiânia, Brasil: Sociedade Brasileira de Zootecnia-SBZ, Associação Brasileira dos Zootecnistas. http://www.adaltech.com.br/anais/zootecnia2018/listaresumos.htm
- De Lima, M. A., and W. S. Sasso. 1985. “Histochemical Detection of Glycoproteins in the Gastric Epithelia of Columba Livia”.” Acto Histochemica 76 (2): 145–150. doi:https://doi.org/10.1016/S0065-1281(85)80051-9.
- De Maesschalck, C., V. Eeckhaut, L. Maertens, L. De Lange, C. Marchal, C. Nezer, S. De Baere, et al. 2015. “Effects of Xylo-Oligosaccharides on Broiler Chicken Performance and Microbiota.” Applied and Environmental Microbiology 81 (17): 5880–5888. doi:https://doi.org/10.1128/AEM.01616-15.
- Ding, M., D. D. Li, S. P. Bai, J. P. Wang, Q. F. Zeng, Z. W. Su, and K. Y. Zhang. 2018. “Effect of Dietary Xylooligosaccharides on Intestinal Characteristics, Gut Microbiota, Cecal Short-Chain Fatty Acids, and Plasma Immune Parameters of Laying Hens.” Poultry Science 97 (3): 874–881. doi:https://doi.org/10.3382/ps/pex372.
- Dormitorio, T. V., J. J. Giambrone, and F. J. Hoerr. 2007. “Transmissible Proventriculitis in Broilers.” Avian Pathology 36: 87–91. doi:https://doi.org/10.1080/03079450601142588.
- Duke, G. E. 1986. “Alimentary Canal: Secretion and Digestion, Special Digestive Functions, and Absorption.” In Avian Physiology, edited by P. D. Sturkie, 291. New York: Springer-Verlag. doi:https://doi.org/10.1007/978-1-4612-4862-0_12.
- Dunlop, M. W., A. F. Moss, P. J. Groves, S. J. Wilkinson, R. M. Stuetz, and P. H. Selle. 2016. “The Multidimensional Causal Factors of Wet Litter in Chicken-Meat Production.” Science of the Total Environment 562: 766–776. doi:https://doi.org/10.1016/j.scitotenv.2016.03.147.
- Engberg, R. M., M. S. Hedemann, S. Steenfeldt, and B. B. Jensen. 2004. “Influence of Whole Wheat and Xylanase on Broiler Performance and Microbial Composition and Activity in the Digestive Tract.” Poultry Science 83: 925–938. doi:https://doi.org/10.1093/ps/83.6.925.
- Farran, M., M. Pietsch, and T. Chabrillat 2013. “Effect of Lignocellulose on the Litter Quality and the Ready to Cook Carcass Yield of Male Broilers.” Actes des 10èmes Journées de la Recherche Avicole et Palmipèdes à Foie Gras du 26 au 28 mars, 2013, La Rochelle, France, 917–921.
- Fischer, M. M., A. M. Kessler, L. R. De Sa, F. O. Vasconcello, S. P. Roberti Filho, M. Nogueira, M. C. C. Oliverira, and A. C. Carciofi. 2012. “Fiber Fermentability Effects on Energy and Macribility, Fecal Traits, Postprandial Metabolite Responses, and Colon Histology of Overweight Cats.” Journal of Animal Science 90: 2233–2245. doi:https://doi.org/10.2527/jas.2011-4334.
- Fuller, S., E. Beck, H. Salman, and L. Tapsell. 2016. “New Horizons for the Study of Dietary Fiber and Health: A Review.” Plants Foods for Human Nutrition 71 (1): 1–12. doi:https://doi.org/10.1007/s11130-016-0529-6.
- Galbraith, H., and T. B. Miller. 1973. “Effect of Long Chain Fatty Acids on Bacterial Respiration and Amino Acid Uptake.” Journal of Applied Bacteriology 36: 659–675. doi:https://doi.org/10.1111/j.1365-2672.1973.tb04151.x.
- Gonzales-Alvarado, J. M., E. Jimenez-Moreno, D. G. Valencia, R. Lazaro, and G. G. Mateos. 2008. “Effects of Fiber Source and Heat Processing of the Cereal on the Development and pH of the Gastrointestinal Tract of Broilers Fed Diets Based on Corn or Rice.” Poultry Science 87: 1779–1795. doi:https://doi.org/10.3382/ps.2008-00070.
- Gonzales-Alvarado, J. M., E. Jimenez-Moreno, R. Lazaro, and G. G. Mateos. 2007. “Effects of Type of Cereal, Heat Processing of the Cereal, and Inclusion of Fiber in the Diet on Productive Performance and Digestive Traits of Broilers.” Poultry Science 86: 1705–1715. doi:https://doi.org/10.1093/ps/86.8.1705.
- Gonzalez-Alvarado, J. M., E. Jimenez-Moreno, D. Gonzalez-Sanchez, R. Lazaro, G. G. Mateos. 2010. “Effect of Inclusion of Oat Hulls and Sugar Beet Pulp in the Diet on Productive Performance and Digestive Traits of Broilers From 1 to 42 days of Age.” Animal Feed Science and Technology 62 (1-2): 37–46 anifeedsci.2010.08.010
- Grobas, S., G. G. Mateos, and J. Mendez. 1999a. “Influence of Dietary Linoleic Acid on Production and Weight of Eggs and Egg Components in Young Brown Hens.” Journal of Applied Poultry Research 8: 177–184. doi:https://doi.org/10.1093/japr/8.2.177.
- Guzmán, P., B. Saldana, H. A. Mandalawi, A. Perez-Bonilla, R. Lazaro, and G. G. Mateos. 2015. “Productive Performance of Brown-Egg Laying Pullets from Hatching to 5 Weeks of Age as Affected by Fiber Inclusion, Feed Form, and Energy Concentration of the Diet.” Poultry Science 94 (2): 249–261. doi:https://doi.org/10.3382/ps/peu072.
- Hartini, S., G. Hinch, A. Kocher, and J. V. Nolan. 2002. “Effects of Light Intensity during Rearing and Beak Trimming and Dietary Fiber Sources on Mortality, Egg Production, and Performance of ISA Brown Laying Hens.” Journal of Applied Poultry Research 11: 104–110. doi:https://doi.org/10.1093/japr/11.1.104.
- Hartini, S., and M. Choct. 2011. “Effect of Mash Dietary Fibre on Performance and Cannibalism in Laying Hens.” Journal of the Indonesian Tropical Animal Agriculture 36 (1): 50–54. doi:https://doi.org/10.14710/jitaa.36.1.50-54.
- Hassan, M. M., A. S. Morsy, and A. M. Hasan. 2013. “Egg Yolk Cholesterol and Productive Performance of Laying Hens Influenced by Dietary Crude Fibre Levels under Drinking Natural Saline Water.” Journal of Animal and Poultry Production 4 (3): 161–176. doi:https://doi.org/10.21608/jappmu.2013.71020.
- Henneberg, W., and F. T. Stohmann. 1859. “Uber Das Erhaltungsfutter Volljahrigen Rindviehs.” Journal Der Landwirtschaft 3: 485–551.
- Henneberg, W., and F. T. Stohmann. 1860. Beiträge Zur Begründung Einer Rationellen Fütterung Der Wiederkäuer I & II. Braunschweig: C.A. Schwetschke und sohn.
- Henuk, Y. L., and J. G. Dingle. 2003. “Poultry Manure: Source of Fertilizer, Fuel and Feed.” World’s Poultry Science Journal 59: 350–360. doi:https://doi.org/10.1079/WPS20030022.
- Hetland, H., and B. Svihus. 2001. “Effect of Oat Hulls on Performance, Gut Capacity and Feed Passage Time in Broiler Chickens.” British Poultry Science 42: 354–361. doi:https://doi.org/10.1080/00071660120055331.
- Hetland, H., B. Svihus, and A. Krogdahl. 2003. “Effects of Oat Hulls and Wood Shavings on Digestion in Broilers and Layers Fed Diets Based on Whole or Ground Wheat.” British Poultry Science 44 (2): 275–282. doi:https://doi.org/10.1080/0007166031000124595.
- Hetland, H., B. Svihus, and M. Choct. 2005. “Role of Insoluble Fiber on Gizzard Activity in Layers.” Journal of Applied Poultry Research 14: 38–46. doi:https://doi.org/10.1093/japr/14.1.38.
- Hetland, H., B. Svihus, and V. Olaisen. 2002. “Effect of Feeding Whole Cereals on Performance, Starch Digestibility and Duodenal Particle Size Distribution in Broiler Chickens.” British Poultry Science 43: 416–423. doi:https://doi.org/10.1080/00071660120103693.
- Hetland, H., B. Svihus, and V. Olaisen. 2010. “Effect of Feeding Whole Cereals on Performance, Starch Digestibility and Duodenal Particle Size Distribution in Broiler Chickens.” British Poultry Science 43 (3): 416–423. doi:https://doi.org/10.1080/00071660120103693.
- Huber-Eicher, B., and B. Wechsler. 1997. “Feather Pecking in Domestic Chicks: Its Relation to Dustbathing and Foraging.” Animal Behaviour 54: 757–768. doi:https://doi.org/10.1006/anbe.1996.0506.
- Incharoen, T., and P. Maneechote. 2013. “The Effects of Dietary Whole Rice Hulls as Insoluble Fiber on the Flock Uniformity of Pullets and on Egg Performance and Intestinal Mucosa of Laying Hens.” American Journal of Agricultural and Biological Sciences 8 (4): 323–329. doi:https://doi.org/10.3844/ajabssp.2013.323.329.
- Jaworski, N. W., H. N. Laerke, K. E. Bach Knudsen, and H. H. Stein. 2015. “Carbohydrate Composition and in Vitro Digestibility of Dry Matter and Non-starch Polysaccharides in Corn, Sorghum Wheat and Coproducts from These Grains.” Journal of Animal Science 93 (3): 1103–1113. doi:https://doi.org/10.2527/jas.2014-8147.
- Jha, R., and J. D. Berrocoso. 2015. “Dietary Fiber Utilization and Its Effects on Physiological Functions and Gut Health of Swine.” Animals 9: 1441–1452. doi:https://doi.org/10.1017/S1751731115000919.
- Jha, R., J. M. Fouhse, U. P. Tiwari, L. Li, and B. P. Willing. 2019. “Dietary Fiber and Intestinal Health of Monogastric Animals.” Frontiers in Veterinary Science 6: 48. doi:https://doi.org/10.3389/fvets.2019.00048.
- Jimenez-Moreno, E., A. De Coca-Sinova, J. Garcia, and G. G. Mateos. 2013. “Oat Hulls and Sugar Beet Pulp in Diets for Broilers 1. Effects on Growth Performance and Nutrient Digestibility.” Animal Feed Science and Technology 182: 33–43. doi:https://doi.org/10.1016/j.anifeedsci.2013.03.011.
- Jimenez-Moreno, E., J. M. Gonzalez-Alvarado, R. Lazaro, and G. G. Mateos. 2009. “Effects of Type of Cereal, Heat Processing of the Cereal, and Fiber Inclusion in the Diet on Gizzard pH and Nutrient Utilization in Broilers at Different Ages.” Poultry Science 88: 1925–1933. doi:https://doi.org/10.3382/ps.2009-00193.
- Johansson, F. 2014. Effect of Insoluble Fibre Enrichment on Egg Quality in Laying Hens. Swedish University of Agricultural Sciences. https://stud.epsilon.slu.se/6492/7/johansson_f_140310.pdf.
- Johnson, I., and J. M. Gee. 1981. “Effect of Gel-Forming Gums on the Intestinal Unstirred Layer and Sugar Transport in Vitro.” Gut 22: 398–403. doi:https://doi.org/10.1136/gut.22.5.398.
- Kalmendal, R., W. Bessei. 2012. “The Preference for High-fiber Feed in Laying Hens Divergently Selected on Feather Pecking.” Poultry Science 91(8): 1785–1789.
- Kalmendal, R., and H. Wall. 2012. “Effects of a High Oil and Fibre Diet and Supplementary Roughage on Performance, Injurious Pecking and Foraging Activities in Two Layer Hybrids.” British Poultry Science 53 (2): 153–161. doi:https://doi.org/10.1080/00071668.2012.682146.
- Karlson, A. 1996. Effekt Av Olika Spannmålsslag På Fjäderplockning Och Kannibalism Hos Värphöns I Olika Inhysninssystem. Examensarbete 74. Uppsala: Swedish University of Agric. Sci.
- Kheravii, S., N. K. Morgan, R. A. Swick, M. Choct, and S. B. Wu. 2018. “Roles of Dietary Fibre and Ingredient Particle Size in Broiler Nutrition.” World’s Poultry Science Journal 74 (2): 301–316. doi:https://doi.org/10.1017/S0043933918000259.
- Kheravii, S., R. A. Swick, M. Choct, and S. B. Wu. 2017. “Coarse Particle Inclusion and Lignocellulose-Rich Fiber Addition in Feed Benefit Performance and Health of Broiler Chickens.” Poultry Science 96: 3272–3281. doi:https://doi.org/10.3382/ps/pex123.
- Kiarie, E., L. F. Romero, and V. Ravindran. 2014. “Growth Performance, Nutrient Utilization, and Digesta Characteristics in Broiler Chickens Fed Corn or Wheat Diets without or with Supplemental Xylanase.” Poultry Science 5 (1): 1186–1196. doi:https://doi.org/10.3382/ps.2013-03715.
- Kim, S. W., W. Li, R. Angel, and P. W. Plumstead. 2019. “Modification of a Limestone Solubility Method and Potential to Correlate with Limestone Calcium Digestibility.” Poultry Science 98 (12): 6837–6848. doi:https://doi.org/10.3382/ps/pez423.
- Kirchgessner, M., M. Kreuzer, A. Machmuller, and D. A. Roth-Maier. 1994. “Evidence for a High Efficiency of Bacterial Protein Synthesis in the Digestive Tract of Adult Sows Fed Supplements of Fibrous Feedstuffs.” Animal Feed Science and Technology 46: 293–306. doi:https://doi.org/10.1016/0377-8401(94)90147-3.
- Kleyn, R. 2013. Chicken Nutrition: A Guide for Nutritionists and Poultry Professionals. United Kingdom: Context Publishing.
- Kocer, B., M. Bozkurt, G. Ege, A. E. Tuzun. 2021. “Effects of Sunflower Meal Supplementation in the Diet on Productive Performance, Egg Quality and Gastrointestinal Tract Traits of Laying Hens.” British Poultry Science 62(1): 101–109.
- Lázaro, R., M. Garcia, M. J. Aranibar, and G. G. Mateos. 2003. “Effect of Enzyme Addition to Wheat-, Barley-and Rye-based Diets on Nutrient Digestibility and Performance of Laying Hens.” British Poultry Science 44 (2): 256–265. doi:https://doi.org/10.1080/0007166031000085616.
- Lee, K. W., H. J. Everts, M. Kappert, M. Frehner, R. Losa, A. C. Beyen. 2003. “Effects of Dietary Essential Oil Components on Growth Performance, Digestive Enzymes and Lipid Metabolism in Female Broiler Chickens.” British Poultry Science 44 (3):450–457. doi:https://doi.org/10.1080/0007166031000085508
- Li, D. D., X. M. Ding, K. Y. Zhang, J. P. Wang, Z. W. Zeng, and L. Kang. 2017. “Effects of Dietary Xylooligosaccharides on the Performance, Egg Quality, Nutrient Digestibility and Plasma Parameters of Laying Hens.” Animal Feed Science and Technology 225: 20–26. doi:https://doi.org/10.1016/j.anifeedsci.2016.12.010.
- Liu, H., J. Wang, T. He, S. Becker, G. Zhang, D. Li, and X. Ma. 2018. “Butyrate: A Double-edged Sword for Health?” Advances in Nutrition 9 (1): 21–29. doi:https://doi.org/10.1093/advances/nmx009.
- Mahmood, T., and Y. Guo. 2020. “Dietary Fiber and Chicken Microbiome Interaction: Where Will It Lead To?” Animal Nutrition 6 (1): 1–8. doi:https://doi.org/10.1016/j.aninu.2019.11.004.
- Mateos, G. G., E. Jimenez-Moreno, M. P. Serrano, and R. P. Lazaro. 2012. “Poultry Response to High Levels of Dietary Fiber Sources Varying in Physical and Chemical Characteristics.” Journal of Applied Poultry Research 21 (1): 156–174. doi:https://doi.org/10.3382/japr.2011-00477.
- Mateos, G. G., R. Lazaro, and M. I. Gracia. 2002. “The Feasibility of Using Nutritional Modifications to Replace Drugs in Poultry Feeds.” Journal of Applied Poultry Research 11: 437–452. doi:https://doi.org/10.1093/japr/11.4.437.
- Mens, A. J. W., M. M. Van Krimpen, and R. P. Kwakkel. 2020. “Nutritional Approaches to Reduce or Prevent Feather Pecking in Laying Hens: Any Potential to Intervene during Rearing?” World’s Poultry Science Journal 76 (3): 591–610. doi:https://doi.org/10.1080/00439339.2020.1772024.
- Mirzaie, S., M. Zaghari, S. Aminzadeh, M. Shivazad, and G. G. Mateos. 2012. “Effects of Wheat Inclusion and Xylanase Supplementation of the Diet on Productive Performance, Nutrient Retention, and Endogenous Intestinal Enzyme Activity of Laying Hens.” Poultry Science 91 (2): 413–425. doi:https://doi.org/10.3382/ps.2011-01686.
- Moore, S. J. 1999. “Food Breakdown in an Avian Herbivore; Who Needs Teeth?” Australian Journal of Zoology 47: 625–632. doi:https://doi.org/10.1071/ZO99051.
- Nabizadeh, A. 2012. “The Effect of Inulin on Broiler Chicken Intestinal Microflora, Gut Morphology, and Performance.” Journal of Animal Feed Science 2: 725–734. doi:https://doi.org/10.22358/jafs/66144/2012.
- NRC. 2001. Nutrient Requirements of Beef Cattle. Seventh revised ed. Washington DC, USA: National Academies Press.
- Panaite, C., D. Dragatoiu, R. Marti, T. Panaite. 2015. “Use of High-fibre Feedstuffs in Pullet Diets Starting With the Starter Stage.” Atchiva Zootechnica 18(1): 53–63.
- Panaite, C. V., R. D. Criste, D. Dragotoiu, T. D. Panaite, and M. Olteanu 2016. “Effect of Crude Fibre Concentration in Pullet Diets (9-16 Weeks) on Their Subsequent Performance.” The International Conference of the University of Agronomic Sciences and Veterinary Medicine of Bucharest Agriculture for Life, Life for Agriculture, Bucharest, Romania.
- Péron, A., B. Svihus, I. Gabriel, S. Berot, D. Tanguy, and B. Bouchet. 2007. “Effects of Two Wheat Cultivars on Physico-Chemical Properties of Wheat Flours and Digesta from Two Broiler Chicken Lines (D+ and D-) Differing in Digestion Capacity.” British Poultry Science 48: 370–380. doi:https://doi.org/10.1080/00071660701341963.
- Péron, A., D. Bastianelli, F. X. Oury, J. Gomez, and B. Carre. 2005. “Effects of Food Deprivation and Particle Size of Ground Wheat on Digestibility of Food Components in Broilers Fed on a Pelleted Diet.” British Poultry Science 46: 223–230. doi:https://doi.org/10.1080/00071660500066142.
- Podolske, J. 2013. “Rice Hulls Fibre: Food Application, Physiological Benefits, and Safety.” Cereal Foods World 2013 (58): 127–131. doi:https://doi.org/10.1094/CFW-58-3-0127.
- Qaisrani, S. N., M. M. Van Krimpen, and R. P. Kwakkel. 2013. “Effects of Dietary Dilution Source and Dilution Level on Feather Damage, Performance, Behaviour, and Litter Condition in Pullets.” Poultry Science 92 (3): 591–602. doi:https://doi.org/10.3382/ps.2012-02378.
- Rezaei, M., M. A. Karimi Torshizi, H. Wall, and E. Ivarsson. 2018. “Body Growth, Intestinal Morphology and Microflora of Quail on Diets Supplemented with Micronized Wheat Fibre.” British Poultry Science 59: 422–429. doi:https://doi.org/10.1080/00071668.2018.1460461.
- Rezaei, M., M. A. Karimi Torshizi, and Y. Rouzbehan. 2011. “The Influence of Micronized Insoluble Fibre on Performance and Litter Moisture.” Poultry Science 90: 2008–2012. doi:https://doi.org/10.3382/ps.2011-01352.
- Rivière, A., M. Gagnon, S. Weckx, D. Roy, and L. De Vuyst. 2015. “Mutual Cross-Feeding Interactions between Bifidobacterium Longum Subsp. Longum NCC2705 and Eubacterium Rectale ATCC 33656 Explain the Bifidogenic and Butyrogenic Effects of Arabinoxylan Oligosaccharides.” Applied and Environmental Microbiology 81 (22): 7767–7781. doi:https://doi.org/10.1128/AEM.02089-15.
- Roberts, S. A., H. Xin, B. J. Kerr, J. R. Russell, and K. Bregendahl. 2007. “Effects of Dietary Fiber and Reduced Crude Protein on Ammonia Emission from Laying-Hen Manure.” Poultry Science 86 (8): 1625–1632. doi:https://doi.org/10.1093/ps/86.8.1625.
- Rodenburg, T. B., M. M. Van Krimpen, I. C. De Jong, E. N. De Hass, M. S. Kops, B. J. Riedstra, R. E. Nordquist, J. P. Wagenaar, M. Bestman, and C. J. Nicol. 2013. “The Prevention and Control of Feather Pecking in Laying Hens: Identifying the Underlying Principles.” World's Poultry Science Journal 69(2): 361–374. doi:https://doi.org/10.1017/S0043933913000354
- Rogel, A. M., E. F. Annison, and W. L. Bryden. 1987. “The Digestion of Wheat Starch in Broiler Chickens.” Australian Journal of Agricultural Research 38: 639–649. doi:https://doi.org/10.1071/AR9870639.
- Röhe, I., J. Urban, A. Dijkslag, J. Te Paske, and J. Zentek. 2019. “Impact of an Energy and Nutrient Reduced Diet Containing 10% Lignocellulose on Animal Performance, Body Composition and Egg Quality of Dual-Purpose Laying Hens.” Archives of Animal Nutrition 73 (1): 1–17. doi:https://doi.org/10.1080/1745039X.2018.1551950.
- Röhe, I., W. Vahjen, F. Metger, and J. Zentek. 2020. “Effect of a “Diluted” Diet Containing 10% Lignocellulose on the Gastrointestinal Tract, Intestinal Microbiota, and Excreta Characteristics of Dual-Purpose Laying Hens.” Poultry Science 99 (1): 310–319. doi:https://doi.org/10.3382/ps/pez492.
- Sacranie, A., B. Svihus, V. Denstadli, B. Moen, P. A. Iji, and M. Choct. 2012. “The Effect of Insoluble Fiber and Intermittent Feeding on Gizzard Development, Gut Motility, and Performance of Broiler Chickens.” Poultry Science 91 (3): 693–700. doi:https://doi.org/10.3382/ps.2011-01790.
- Sadeghi, M., M. Toghyani, and A. Gheisari. 2015. “Effect of Various Fiber Types and Choice Feeding of Fiber on Performance, Gut Development, Humoral Immunity, and Fiber Preference in Broiler Chicks.” Poultry Science 94 (11): 2734–2743. doi:https://doi.org/10.3382/ps/pev292.
- Scott, M. L., E. R. Holm, and R. E. Reynolds. 1954. “Studies on Pheasant Nutrition. 3. Effects of Antibiotics, Arsenicals and Thyroactive Compounds upon Growth and Feathering.” Poultry Science 33: 1261–1265. doi:https://doi.org/10.3382/ps.0331261.
- Seemann, M. 1982. Die Reaktion Von Legehybriden Auf Wahlfütterung. Stuttgart: Universität Hohenheim, Institut für Tierhaltung und Tierzüchtung, Fachgebiet Kleintierzucht.
- Shakouri, M. D., H. Kermanshahi, M. Mohsenzadeh. 2006. “Effect of Different Non Starch Polysaccharides in Semi Purified Diets on Performance and Intestinal Microflora of Young Broiler Chickens.” International Journal of Poultry Science 5(6): 557–561.
- Shang, Y., S. Kumar, H. Thippareddi, W. K. Kim. 2018. “Effect of Dietary Fructooligosaccharide (FOS) Supplementation on Ileal Microbiota in Broiler Chickens.” Poultry Science 97(10): 3622–3634. doi:https://doi.org/10.3382/ps/pey131
- Shang, Y., S. Kumar, B. Oakley, and W. K. Kim. 2018. “Chicken Gut Microbiota: Importance and Detection Technology.” Frontiers in Veterinary Science 5: 254. doi:https://doi.org/10.3389/fvets.2018.00254.
- Silva, V. K., V. Morita, and I. C. Bolei. 2013. “Effect of Pectin Extracted from Citrus Pulp on Digesta Characteristics and Nutrient Digestibility in Broiler Chickens.” Revista Brasileira De Zootechnia 42: 575–582. doi:https://doi.org/10.1590/S1516-35982013000800007.
- Singh, Y., V. Ravindran, T. J. Wester, A. L. Molan, G. Ravindran. 2014. “Influence of Feeding Coarse Corn on Performance, Nutrient Utilization, Digestive Tract Measurements, Carcass Characteristics, and Cecal Microflora Counts of Broilers.” Poultry Science 93(3): 607–616.
- Smits, C. H. M. 1996. “Viscosity of Dietary Fibre in Relation to Lipid Digestibility in Broiler Chickens”. PhD Thesis, Wageningen University, Netherlands.
- Sozcu, A., and A. Ipek. 2020. “The Effects of Lignocellulose Supplementation on Laying Performance, Egg Quality Parameters, Aerobic Bacterial Load of Eggshell, Serum Biochemical Parameters, and Jejunal Histomorphological Traits of Laying Hens.” Poultry Science 99 (6): 3179–3187. doi:https://doi.org/10.1016/j.psj.2020.01.024.
- Steenfeldt, S., J. B. Kjaer, and R. M. Engberg. 2007. “Effect of Feeding Silages or Carrots as Supplements to Laying Hens on Production Performance, Nutrient Digestibility, Gut Structure, Gut Microflora and Feather Pecking Behaviour.” British Poultry Science 48: 454–468. doi:https://doi.org/10.1080/00071660701473857.
- Svihus, B. 2011. “Limitations to Wheat Starch Digestion in Growing Broiler Chickens: A Brief Review.” Animal Production Science, 51(7): 583–589. doi:https://doi.org/10.1071/AN10271
- Svihus, B. 2011. “The Gizzard: Function, Influence of Diet Structure and Effects on Nutrient Availability.” World’s Poultry Science Journal 67 (1): 207–223. doi:https://doi.org/10.1017/S0043933911000249.
- Swanson, K. S., C. M. Grieshop, G. M. Clapper, R. G. Shields, T. Belay, N. R. Merchen, and G. C. Fahey. 2001. “Fruit and Vegetable Fibre Fermentation by Gut Microflora from Canines.” Journal of Animal Science 79 (4): 919–926. doi:https://doi.org/10.2527/2001.794919x.
- Tejeda, O. J., W. K. Kim. 2021. “Role of Dietary Fiber in Poultry Nutrition.” Animals 11(2): 461.doi:https://doi.org/10.3390/ani11020461
- Van der Aar, P. J., G. C. Fahey Jr, S. C. Ricke, S. E. Allen, and L. L. Berger. 1983. “Effects of Dietary Fibers on Mineral Status of Chicks.” Journal of Nutrition 113: 653–661. doi:https://doi.org/10.1093/jn/113.3.653.
- Van Krimpen, M. M., R. P. Kwakkel, V. der Peet-Schwering, L. A. Den Hartog, M. W. A. Verstegen. 2008. “Low Dietary Energy Concentration, High Nonstarch Polysaccharide Concentration, and Course Particle Sizes of Nonstarch Polysaccharides Affect the Behavior of Feather-Pecking-Prone Laying Hens.” Poultry Science 87(3): 485–496. doi:https://doi.org/10.3382/ps.2007–00279
- Van Krimpen, M. M., R. P. Kwakkel, C. M. C. van der Peet-Schwering, L. A. den Hartog, and M. W. A. Verstegen. 2009. “Effects of Nutrient Dilution and Nonstarch Polysaccharide Concentration in Rearing and Laying Diets on Eating Behavior and Feather Damage of Rearing and Laying Hens.” Poultry Science 88: 759–773. doi:https://doi.org/10.3382/ps.2008-00194.
- Van Soest, P. J. 1963. “Use of Detergents in the Analysis of Fibrous Feeds. 1. Preparation of Fibre Residues of Low Nitrogen Content.” Journal of Association of Official Analytical Chemists 46: 825–829. doi:https://doi.org/10.1093/jaoac/73.4.487.
- Wahlström, A., R. Tauson, and K. Elwinger. 1998. “Effects on Plumage Condition, Health and Mortality of Dietary Oats/Wheat Ratios to Three Hybrids of Laying Hens Different Housing System.” Acta Agriculturae Scandinavica 48: 250–259. doi:https://doi.org/10.1080/09064709809362428.
- Walser, P.T. 1997. “Einfluss unterschiedlicher Futterzusammensetzung und -aufarbeitung auf das Auftreten von Federpicken, das Nahrungsaufnahmeverhalten, die Leistung und den Gesamtstoffwechsel bei verschiedenen Legehennenhybriden.” Doctoral Thesis, ETH Zurich. doi:https://doi.org/10.3929/ethz-a–001889734
- Walugembe, M., M. Rothschild, and M. Persia. 2014. “Effects of High Fiber Ingredients on the Performance, Metabolizable Energy and Fiber Digestibility of Broiler and Layer Chicks.” Animal Feed Science and Technology 188: 46–52. doi:https://doi.org/10.1016/j.anifeedsci.2013.09.012.
- Wanzenböck, E., U. Zitz, C. Steinbauer, W. Kneifel, K. J. Domig, and K. Schedle. 2020. “A Diet Containing Native or Fermented Wheat Bran Does Not Interfere with Natural Microbiota of Laying Hens.” Animal 14: 1147–1155. doi:https://doi.org/10.1017/S1751731119003343.
- Yangilar, F. 2013. “The Application of Dietary Fibre in Food Industry: Structural Features, Effects on Health and Definition, Obtaining and Analysis of Dietary Fibre: A Review.” Journal of Food and Nutrition Research 1: 13–23. http://article.foodnutritionresearch.com/pdf/jfnr-1-3-1.pdf.
- Yokhana, S., G. Parkinson, and T. L. Frankel. 2016. “Effect of Insoluble Fiber Supplementation Applied at Different Ages on Digestive Organ Weight and Digestive Enzymes of Layer-Strain Poultry.” Poultry Science 95: 550–559. doi:https://doi.org/10.3382/ps/pev336.
- Zdunczyk, Z., D. Mikulski, J. Jankowski, B. Przybylska-Gornowicz, and J. Juskiewicz. 2019. “Gastrointestinal Response of Laying Hens to Graded Dietary Inclusion Levels of Yellow Lupine Seeds.” Animal Feed Science and Technology 255: 114214. doi:https://doi.org/10.1016/j.anifeedsci.2019.114214.