Abstract
To understand the relationship between ploidy levels and fruit characters in kiwifruit, the ploidy of 30 important Chinese cultivars is determined by flow cytometry and eight fruit characters are described following the UPOV descriptors. The provenance of the cultivars is noted. The type of fruit hairs, fruit flesh colour and fruit weight are found to be related to the ploidy. Cultivars at different ploidies vary in fruit hairs, ranging from downy to hispid. Diploid and tetraploid cultivars have mostly yellow or golden yellow fruit flesh, whereas hexaploid cultivars have mostly green fruit flesh. Partial correlation analysis indicates a positive correlation between ploidy and fruit weight. However, fruit shape, fruit skin colour, soluble solids content, vitamin C content and total acid content vary and are not significantly correlated to ploidy. Cultivars are predominantly selected from the Xuefeng mountains, Mufu mountains, Qinling mountains and Daba mountains, centres of morphological polymorphism and ploidy variation of Actinidia chinensis var. chinensis and A. chinensis var. deliciosa.
Introduction
Polyploidy is common in plants and has classically been recognized as one of the most important evolutionary events because of polyploid genome ‘buffering’ increased allelic diversity, and increased or ‘fixed’ heterozygosity (Masterson Citation1994; Udall and Wendel Citation2006; Leitch & Leitch Citation2008; Soltis et al. Citation2009). With increased allelic diversity and heterozygosity, and with the opportunity for novel phenotypic variation to arise from duplicated genes acquiring new functions, polyploids have long been considered an important source of diversity for crop improvement (Stebbins Citation1950), even in modern plant breeding programmes. Nevertheless, the complexity of the polyploidy process involving changes of the organization and function of the genome at both genetic and epigenetic levels is still a major challenge if polyploids are to be used in crop improvement (Soltis et al. Citation2004). Successful selection of polyploid cultivars is dependent on understanding and unravelling the complex genetic basis underlying the phenotype.
Kiwifruit (Actinidia) are native to China and are extraordinarily diverse in both genetic and morphological variation (Li et al. Citation2007). For instance, Actinidia plants occur in nature as diploids, tetraploids, hexaploids and even octoploids in a reticulate structure (Xiong Citation1992; Yan et al. Citation1994; Ferguson et al. Citation1997; Huang & Ferguson Citation2007). Most kiwifruit cultivars are selections of Actinidia chinensis var. chinensis and A. chinensis var. deliciosa (Li et al. Citation2007, Citation2009) domesticated and bred in China and other countries during the past century (Xiao et al. Citation1999; Cui et al. 2002; Huang & Ferguson Citation2003). Kiwifruit breeding, however, is difficult because of the need to choose male parents whose contribution to fruit characters is not known, and determine the ploidy levels of the parents (e.g., A. kolomikta, A. polygama, A. arguta, Start et al. Citation2007) that can restrict crossing. The long generation time is another constraint. Therefore, the direct selection of the superior genotypes from the wild is very rewarding. This does raise another problem: what are the important fruit characters by which genotypes of varying ploidy should be selected from the variation existing in the wild? Since polyploidy is as a major force for plant evolution and diversification, we consider it important for germplasm improvement to examine the correlation between the fruit characters of Actinidia plants selected and their ploidy levels.
A. chinensis var. chinensis and A. chinensis var. deliciosa naturally occur across much of China from east to west, with considerable overlaps (Liu et al. Citation2006; Li et al. Citation2007; Zhang et al. Citation2007). Recent studies suggest that the most variable populations, in terms of ploidy and morphology, of the species are centred in mid-western China, presenting a large topographical and eco-geographical gradient (unpublished data). Cultivars selected from wild A. chinensis var. chinensis and A. chinensis var. deliciosa plants also came originally from a wide geographical range from Fujian Province to Sichuan Province (Cui et al. Citation2002). However, little is known about which geographical regions are most promising for cultivar selection and improvement. As a fruit tree with a known short history for domestication (about 100 years from 1904), the main kiwifruit cultivars domesticated in China were mostly collected during the nationwide investigation of germplasm resources of Actindia genus in 1977–78 (Cui Citation1993). To take advantage of the diversity in wild Actinidia populations, breeders need to know the potentially most useful areas from which good germplasm could be collected (Huang & Ferguson Citation2007). Thus, a comparison of the overall geographical distribution of A. chinensis var. chinensis and A. chinensis var. deliciosa and the main localities from which the most commercially important cultivars were collected would be useful for future germplasm collections and breeding programmes.
In the present study, we used flow cytometry to determine the ploidy levels of the most important kiwifruit cultivars in China. Some fruit characters of these cultivars were described following the International Union for the Protection of New Varieties of Plants descriptors (UPOV), and the geographical localities from which they were initially collected were noted. To improve our current breeding programmes, these data were examined to determine first, whether there is a correlation between the main fruit characters and ploidy, and second, where the major kiwifruit cultivars were selected and where future collecting should focus.
Materials and methods
Plant materials, morphological and geographical origin analyses
In total, 30 important female kiwifruit cultivars, accounting for about 80% of Chinese kiwifruit production, were studied. Plants were grown in the national Actinidia germplasm repository of the Wuhan Institute of Botany (WIB). Four qualitative traits—the nature of the fruit hairs, fruit shape, fruit skin colour and flesh colour—were described using the UPOV descriptors, and four quantitative traits—fruit weight (FW), vitamin C content (Vc), soluble solids content (SSC) and total acid content (TA)—were measured according to the methods of Cheng et al. (Citation2004). Most of the quantitative traits were collected in 3 consecutive years, and the average values of each trait for these three years were listed in . In addition to observations on fruit characters collected from the WIB, we also referred to the literature for descriptions of the fruit characters and for information on provenance.
Table 1 Ploidy levels, fruit characters and provenances of the 30 most important kiwifruit (Actinidia) cultivars in China
Ploidy analysis
Ploidy was determined by flow cytometry (FCM). Hardwood cuttings of the 30 cultivars were collected during the winter in 2007. They were placed in a warm room to induce budburst and the newly expanding leaves were harvested. To release individual nuclei for FCM measurements, the leaves were chopped in 0.5 ml of nuclear extraction buffer (solution A of High Resolution Kit, Partec, Germany) and incubated for 2 min, which was then filtered through a nylon sieve with a mesh diameter of 30 µm (CellTricsTM, Partec, Germany), followed by the addition of 2 ml 6-diamidino-2-phenylindole (DAPI, solution B of the kit) held at 4°C. Two minutes later, the samples were measured to estimate their ploidy levels on Ploidy Analyser-II (Partec, Munster, Germany) automatically. Generally, only samples producing two histogram peaks (sample and standard) with a low coefficient of variation (<6%) were retained. The clones of previously cytologically studied diploid (2n =58) cultivars of A. chinensis ‘Hongyang’ were used as an internal reference standard for the relative DNA content measurements.
Statistical analyses
According to the ploidy levels examined, we separated the cultivars into three groups: the diploid cultivar group, the tetraploid cultivar group and the hexaploid cultivar group. One-way analysis of variance (ANOVA) was used to test for differences among different ploidy cultivars in fruit weight, vitamin C content, soluble solid content and total acid content. When ANOVA gave a significant result (P<0.05), Tukey tests were used a posteriori to identify the source of the significance (Sokal & Rohlf Citation1981). Pearson coefficients were calculated to assess correlation between ploidy levels and fruit characters. In all cases, data were arcsine transformed to meet the assumptions of normality and homogeneity of variances (Sokal & Rohlf Citation1981). All analysis of data was conducted using SPSS version 13.0 (Zar Citation1999).
Results
Ploidy levels and fruit characteristics
The main fruit characters, the provenances and the ploidy levels of the 30 most important kiwifruit cultivars in China are summarized in . Three cytotypes were observed, including six diploids, 13 tetraploids and 11 hexaploids. The predominant fruit shapes were oblong, ovoid, ellipsoid and spheroid. The fruit skins were mostly light brown, medium brown and reddish green. Within a ploidy group, the fruit hairs were mostly similar, but between ploidy groups ranged from downy (diploid) to tomentose (diploid or tetraploid) and to hispid (hexaploid). The flesh colour also changed from yellow (diploid and tetraploid) to green (hexaploid), except for hexaploid ‘Chuhong’ and diploid ‘Hongyang’ which had red pigments in the inner pericarp. Of the four qualitative characters, the nature of the fruit hairs and flesh colour altered according to the different ploidy of the cultivars.
For the four quantitative characters (FW, Vc, SSC and TA), the ANOVA revealed that there were no significant differences except for fruit weight (F = 5.045, P<0.05) (). The Tukey HSD test revealed that the fruit weight of the hexaploid cultivars is greater than that of diploid and tetraploid cultivars, whereas there were no significant differences between diploid and tetraploid cultivars (). Moreover, the Pearson's correlated analysis (Pearson's correlation coefficient, PCC = 0.490, n=30, P<0.05) indicated that there was a significant positive correlation between the ploidy and fruit weight. Apart from fruit weight, there were no significant correlations between other quantitative characters (Vc, SSC and TA) and ploidy levels (P >0.05).
Table 2 ANOVA and Tukey analyses of fruit weight (FW), vitamin C content (Vc), soluble solid content (SSC) and total acid content (TA) of the 30 most important kiwifruit (Actinidia) cultivars in China
The geographical origin
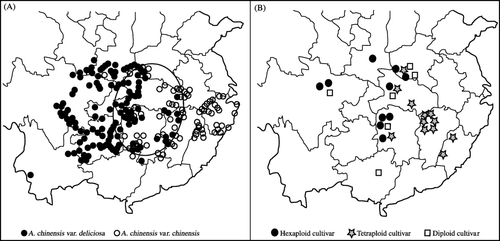
The cultivars were selected mainly from Jiangxi Province (six cultivars), Hunan Province (six cultivars), Henan Province (five cultivars), Hubei Province (four cultivars) and Sichuan Province (three cultivars). There is an obvious trend for hexaploid cultivars to be selected from western provinces, while tetraploid and diploid cultivars came from eastern and central provinces of China. The comparison between the original localities of the 30 dominant cultivars and the natural distribution of A. chinensis var. chinensis and A. chinensis var. deliciosa () indicated that the original areas of the cultivars overlapped in sympatric areas of A. chinensis var. chinensis and A. chinensis var. deliciosa. For instance, the Xuefeng mountains, which is the sympatric area of A. chinensis var. chinensis and A. chinensis var. deliciosa, are the original locality of ‘Miliang-1’, ‘Chuhong’, ‘Cuiyu’ and ‘Fengyue’, and the Mufu mountains are the original locality of ‘Jintao’, ‘Jinzao’, ‘Jinfeng’, ‘Kuimi’ and ‘Lushanxiang’.
Fig. 1 A) Natural distribution of Actinidia chinensis var. chinensis (open circles) and A. chinensis var. deliciosa (solid circles) in China (according to Li et al. Citation2007). Each point denotes a collection site of a herbarium specimen and the area in which the two species overlap is circled by a solid line. B) Localities where the main kiwifruit cultivars in China were selected.
Discussion
The nature of the fruit hairs and fruit flesh colour, but not fruit shape or skin colour, is correlated with ploidy. In general, the fruit shape and skin colour varies considerably within a cultivar ploidy group. Random selection is thus the principal method for improvement of fruit shape and skin colour in China. Although the hairs protect the developing fruit from mechanical damage or some insect pests, the long coarse hairs of hexaploid cultivars are generally considered a fault or disadvantage to consumers that should be selected against (Huang & Ferguson Citation2007). The tendency of the diploid and tetraploid cultivars to have downy or tomentose hairs, and hexaploid cultivars to have bristly or hispid hairs indicated that the fruit hairs differentiate between different ploidy groups. The hexaploid cultivars, such as ‘Qinmei’, are necessary to improve their long coarse hairs. The hexaploid cultivars, ‘Chuhong’, which belong to A. chinensis var. deliciosa, have downy hairs, making the fruit more appealing. This cultivar could be used in breeding programmes to get fruit with softer hairs.
Differences in fruit flesh colour are more obvious and appeal more to the consumer than any other differences between the fruit of the species A. chinensis var. chinensis and A. chinensis var. deliciosa except perhaps for skin hairiness. Flesh colour results from different combinations of chlorophylls, carotenoids and anthocyanins, which may change as the fruit mature and ripen (McGhie & Ainge Citation2002; Minchin et al. Citation2003). The fruit flesh of all A. chinensis var. deliciosa cultivars studied so far is a brilliant, translucent emerald green colour, and the only occasional variation is in intensity of colour. Different genotypes of A. chinensis var. chinensis have fruit ranging in colour from lime green to yellow, the final shade depending on the maturity of the fruit. Of the 30 cultivars studied, hexaploid cultivars have uniform green flesh colour, except for ‘Chuhong’ (Wang et al. Citation2004), and most of the diploid and tetraploid cultivars have a yellow or golden yellow flesh colour except for ‘Cuiyu’ which has green flesh and ‘Hongyang’ with red in the inner pericarp. Ploidy is generally correlated with the fruit flesh colour.
Based on the Pearson's correlated analysis, of the four quantitative fruit characters (FW, Vc, SSC and TA), ploidy is correlated only with fruit weight. Fruit weight is considered to be one of the most important initial criteria in determining which cultivar has commercial potential. Our results confirm the positive correlation between ploidy level and fruit size (weight); however, significant differences in average fruit weight were not found between diploid and tetraploid cultivars, but only between hexaploid cultivars and diploid or tetraploid cultivars. The possibility that tetraploids may have larger fruit than diploids may not have been disproved because fewer diploid cultivars were available for statistical analyses.
Kiwifruit are well known for their nutritional value and good flavour. The outstanding nutritional quality of kiwifruit is their remarkably high vitamin C content, and the good flavour is largely determined by the relative concentrations of acids and sugars present (Huang & Ferguson Citation2007). The range of vitamin C content varies considerably between and within species (Yin & Li Citation1982; Huang et al. Citation2004). We found great variation in the vitamin C content, SSC and TA in the 30 cultivars studied, but there was no consistent pattern between the ploidy cultivar groups. Cheng et al. (Citation2004) suggested that vitamin C content, SSC and TA have high heritabilities in A. chinensis var. chinensis. For the genetic correlations between these characters, selection applied to one character can affect other characters, e.g., increased soluble sugar content should lead to a simultaneous increase in ascorbate content (Cheng et al. Citation2004). Hence, selection for different concentrations of vitamin C, SSC or TS in new cultivars should be based on the concentrations in their parents (Cheng et al. Citation2004).
The relationship between ploidy and fruit flesh colours, fruit hairs and fruit weight has potentially worked in the breeding and commerce of kiwifruit. Nowadays, the kiwifruit market can be segmented according to fruit flesh colour (Zambujo Citation2003). On the basis of fruit flesh colour, nearly 90% of kiwifruit produced anywhere in the world (both A. chinensis var. chinensis and A. chinensis var. deliciosa) have green flesh, and the remainder have yellow flesh (all A. chinensis var. chinensis) (Huang & Ferguson Citation2007). Hexaploid cultivars (A. chinensis var. deliciosa) with big fruit weight and green flesh colour are still the predominant cultivars in the kiwifruit industry. However, this pattern is changing with the commercialization of tetraploid cultivars such as ‘Jintao’ and ‘Jinyan’ which have fruit with tetraploid characters, such as skins covered in soft, fine hairs, striking yellow-gold flesh and a sweeter, more aromatic flavour. This change may be accelerated by a series of new red-fleshed cultivars. The diploid ‘Hongyang’, which has yellow-fleshed fruit with brilliant red around the central core, is cultivated widely in China and is being exported to Japan (Huang & Ferguson Citation2003). These new diploid and tetraploid cultivars provide different types of kiwifruit for the market and provide opportunities for new types with the improved characteristics. The abundant natural resource, ploidy variation and unique fruit characteristic within A. chinensis var. chinensis is greater than within A. chinensis var. deliciosa and this variety may eventually turn out to be more important commercially than A. chinensis var. deliciosa.
As a crop, kiwifruit has been subjected to little selection pressure and is still very similar to plants in the wild (Ferguson Citation2007). The current kiwifruit cultivars in China are mostly selections directly from the wild and are not the products of planned breeding programmes. The current wide range of kiwifruit cultivars available in China represents the superior genotypes of natural resources of Actinidia. Our results have revealed that these cultivars are mostly, directly or indirectly, from the areas of Xuefeng mountains, Mufu mountains, Qingling mountains and Daba mountains. These areas are the transit zone of the second step to the third step of China's topography, and also the region where both A. chinensis var. chinensis and A. chinensis var. deliciosa are found. Our recent extensive and systemic investigation of ploidy and morphological characters for the natural A. chinensis var. chinensis and A. chinensis var. deliciosa populations revealed that there is considerable ploidy and morphological variation in these areas (unpublished data). The diverse habitats in these mountain areas could allow different ploidy races to co-exist and provide chances for hybridization. Frequent hybridization and gene introgression among different ploidy races could result in the reticulate evolutionary structure and many phenotypically transitional or special forms in these areas. The abundant morphological polymorphism and special forms from these areas could be observed both in wild individuals (unpublished data) and cultivars, such as red fruit flesh diploid ‘Hongyang’ and the hairless and red fruit flesh hexaploid ‘Chuhong’. Thus, the Xuefeng mountains, Mufu mountains, Qinling mountains and Daba mountains, which are the centres of morphological and ploidy variation of A. chinensis var. chinensis and A. chinensis var. deliciosa, are the pivotal regions for exploitation and utilization of wild germplasm for cultivar development and species conservation.
Acknowledgements
We sincerely thank A. Ross Ferguson and Canhong Cheng for their critical comments on a previous version of this manuscript. This study was funded by a key initiative grant of the Chinese Academy of Sciences (KSCX2-YW-N-061) and NSF of China grant (30771479). We likewise acknowledge Key Laboratory of Plant Germplasm Enhancement and Speciality Agriculture of WIB-CAS, Research Center for Kiwifruit of WIB-CAS and CAS Key Laboratory of Plant Conservation and Sustainable Utilization of SCBG.
Additional information
Notes on contributors
Caihong Zhong
The first two authors have contributed equally to this workReferences
- Cheng , CH , Seal , AG , Boldingh , HL , Marsh , KB , MacRae , EA , Murphy , AJ and Ferguson , AR . 2004 . Inheritance of taste characters and fruit size and number in a diploid Actinidia chinensis (kiwifruit) population . Euphytica , 138 : 185 – 195 .
- Cui , ZX . 1993 . Actinidia in China , 2 – 12 . China : Shandong Scientific Press .
- Cui , ZX , Huang , HW and Xiao , XG . 2002 . Actinidia in China , Beijing, , China : Agricultural Science and Technology Press .
- Ferguson , AR , O'Brien , IEW and Yan , GJ . 1997 . Ploidy in Actinidia . Acta Horticulturae , 444 : 67 – 71 .
- Ferguson , AR . 2007 . The need for characterisation and evaluation of germplasm: kiwifruit as an example . Euphytica , 154 : 371 – 382 .
- Huang , HW and Ferguson , AR . 2003 . Kiwifruit (Actinidia chinesis and A. deliciosa) plantings and production in China, 2002 . New Zealand Journal of Crop and Horticultural Science , 31 : 197 – 202 .
- Huang , HW and Ferguson , AR . 2007 . Genetic resources of kiwifruit: domestication and breeding . Horticultural Reviews , 33 : 1 – 121 .
- Huang , HW , Wang , Y , Zhang , ZH , Jiang , ZW and Wang , SM . 2004 . Actinidia germplasm resources and kiwifruit industry in China . HortScience , 39 : 1165 – 1172 .
- Leitch , AR and Leitch , IJ . 2008 . Genomic plasticity and the diversity of polyploid plants . Science , 320 : 481 – 483 .
- Li , XW , Li , JQ and Djendoel , SD . 2007 . New synonyms in Actinidiaceae from China . Acta Phytotaxonomica Sinica , 45 ( 5 ) : 633 – 660 .
- Li , XW , Li , JQ and Djendoel , SD . 2009 . Advances in the study of the systematics of Actinidia Lindley . Frontiers of Biology in China , 10 : 55 – 61 .
- Liu , YL , Li , ZZ , Zhang , PF , Jiang , ZW and Huang , HW . 2006 . Spatial genetic structure in natural populations of two closely related Actinidia species (Actinidiaceae) as revealed by SSR analysis . Chinese Biodiversity , 14 : 421 – 434 .
- Masterson , J . 1994 . Stomatal size in fossil plants: evidence for polyploidy in majority of angiosperms . Science , 264 : 421 – 424 .
- McGhie , TK and Ainge , GD . 2002 . Colour in fruit of the genus Actinidia: carotenoid and chlorophyll compositions . Journal of Agricultural and Food Chemistry , 50 : 117 – 121 .
- Minchin , PEH , De Silva , N , Snelgar , WP , Richardson , AC and Thorp , TG . 2003 . Modelling of colour development in the fruit of Actinidia chinensis ‘Hort16A’ . New Zealand Journal of Crop and Horticultural Science , 31 : 41 – 53 .
- Sokal , RS and Rohlf , FJ . 1981 . Biometry: the principles and practice of statistics in biological research , 2nd edn , New York, , USA : Freeman .
- Soltis , DE , Soltis , PS and Tate , JA . 2004 . Advances in the study of polyploidy since Plant speciation . New Phytologist , 161 : 173 – 191 .
- Soltis , DE , Albert , VA , Leebens-Mack , J , Bell , CD , Paterson , AH , Zheng , C , Sankoff , D , DePamphilis , CW , Wall , PK and Soltis , PS . 2009 . Polyploidy and angiosperm diversification . American Journal of Botany , 96 : 336 – 348 .
- Start , MA , Luby , J , Filler , D , Riera-Lizarazu , O and Guthrie , R . 2007 . Ploidy levels of cold-hardy Actinidia accessions in the United States determined by flow cytometry . Acta Horticulturae , 753 : 161 – 168 .
- Stebbins , GL . 1950 . Variation and evolution in plants , New York : Columbia University Press .
- Udall , JA and Wendel , JF . 2006 . Polyploidy and crop improvement . Crop Science , 46 : 3 – 14 .
- Wang , ZY , Zhong , CH and Bu , FW . 2004 . A new red-fleshed kiwifruit cultivar ‘Chuhong’ . Agriculture Science & Technology (China) , 5 ( 4 ) : 23 – 24 .
- Xiao , XG . 1999 . Progress of Actinidia selection and breeding in China . Acta Horticulturae , 498 : 25 – 35 .
- Xiong , ZT . 1992 . Chromosome studies on cultivated Chinese gooseberry . Guihaia , 12 : 79 – 82 .
- Yan , GJ , Ferguson , AR and McNeilage , MA . 1994 . Ploidy races in Actinidia chinensis . Euphytica , 78 : 175 – 183 .
- Yin , WF and Li , X . 1982 . A study of the variation in the vitamin C content of the fruits of Actinidia chinensis Planch . Acta Horticulturae Sinica , 9 ( 2 ) : 31 – 35 .
- Zambujo , CV . 2003 . Du nouveau … hors de France . Arboriculture Fruitière , 568 : 32 – 33 .
- Zar , JH . 1999 . Biostatistical analysis , 4th edn , Englewood Cliffs, NJ : Prentice-Hall .
- Zhang , T , Li , ZZ , Liu , YL , Jiang , ZW and Huang , HW . 2007 . Genetic diversity, gene introgression and homoplasy in sympatric populations of the genus Actinidia as revealed by chloroplast microsatellite markers . Chinese Biodiversity , 15 : 1 – 22 .