
ABSTRACT
Understanding the differences in fruit quality within apricot tree canopies is necessary before developing strategies to reduce variation. Fruit set, fresh weight (FW) and soluble solids and dry matter concentrations (SSC, DMC) were lower, by up to 30%, 28 g, 2.4% and 2.5%, respectively, in fruit from 1-year-old long shoots and terminal spurs or from short shoots and spurs on 4-year-old wood, than on those from 2- and 3-year-old wood, for the six genotypes examined. Genotype, year and wood age affected mealiness, gel development, rubberiness and/or juiciness in fruit after cold storage. Effects of position along the shoot, fruit number and fruiting node leaf area on fruit quality were examined for one cultivar. FW, SSC and DMC were higher in fruit on distal zones of 2-year-old wood, but were lower in fruit from distal zones of 1-year-old wood. Greater fruit number per shoot before thinning decreased FW on 2-year-old wood. On 1-year-old wood, there was an interaction between fruiting node leaf area, position and fruit FW. These results will assist in devising management techniques, such as differential thinning and pruning, to reduce fruit quality variation.
Introduction
Fruit quality within an apricot tree (Prunus armeniaca L.) varies considerably. The benefit of understanding the sources of variation in fruit quality within a tree is substantial, if the variation can then be reduced to provide more consistent and higher quality. Approximately 25% of consumers purchasing apricots delayed repurchasing by more than 2 weeks or did not repurchase them because of the negative eating experience, according to a New Zealand shopping survey (Summerfruit New Zealand Citation2011). Related to this, consumer liking of apricots increased as soluble solids concentration (SSC) increased (Azodanlou et al. Citation2003). These factors emphasise the importance of ensuring fruit achieves optimum quality by the time of harvest.
In a range of fruit types, variation in fruit quality within trees has been studied in some detail. Position in the canopy (Luchsinger et al. Citation2002), light exposure (Robinson et al. Citation1983; George et al. Citation1996), shoot type (Volz et al. Citation1994), wood age (Corelli Grappadelli & Coston Citation1991), leaf area and competition between fruit, both on a whole-tree basis (Crisosto et al. Citation1997) and within single shoots (Corelli Grappadelli & Coston Citation1991), have all been examined. In apricot, the effect of crop load on a whole tree basis has been shown to affect fruit weight (FW) and SSC (Stanley et al. Citation2014).
The source of variation in fruit quality within whole trees has been identified previously. Peach and nectarine fruit from upper canopy positions had greater FW and higher SSC, and fruit were well exposed to light (Luchsinger et al. Citation2002). In apple, light exposure accounted for a large proportion of the variation in fruit size and quality, whereas spur age accounted for a much smaller proportion of the variation (Robinson et al. Citation1983).
Fruit quality was found to vary along a branch or shoot, while fruit number and leaf area also affected fruit quality. Apricot fruit have been reported to be larger on spurs than long shoots, although no data were presented (Lichou et al. Citation1999). Higher fruit density on a shoot reduced fruit size and sometimes SSC for peach (Corelli Grappadelli & Coston Citation1991; Marini & Sowers Citation1994). The location of fruit along the shoot was also an important factor in peach, although results from different trials were sometimes contradictory (Corelli Grappadelli & Coston Citation1991; Marini & Sowers Citation1994). The reason may be related to the presence of axillary shoots on the fruiting shoot (Marini & Sowers Citation1994). Fruit at the distal region of the shoot competed with the terminal shoot for photosynthate, resulting in fruit that had lower FW and SSC than basal fruit (Corelli Grappadelli & Coston Citation1991; Marini & Sowers Citation1994). However, when lateral shoots were present, the source–sink relationship changed so that distal fruit were more competitive and subsequently had greater FW than basal fruit (Marini & Sowers Citation1994). The number of fruits per shoot and overall tree crop density affected the relationship between FW and fruit position (Marini & Sowers Citation1994). In addition, spur leaf area and leaf to fruit ratio were shown to affect fruit quality (Corelli Grappadelli et al. Citation1996).
While the interrelationship between fruit quality and the arrangement and presence of fruit and shoots along a branch may be complex, one key factor was the basis for all of them: the ability of each sink to compete for photoassimilate. The total sink demand of the whole crop and the relative strength of an individual fruit sink compared with other sinks, such as adjacent fruit and growing shoot apices, are factors that enable resources to be accumulated by any particular fruit (Genard et al. Citation1998). Assimilate supply is also important, the main sources of carbohydrate being the leaves. However, stored carbohydrate may play an important role in the early season, particularly since flowering occurs before leaf emergence in apricot. A reduction in stored carbohydrate reserves, from early defoliation in the previous autumn, reduced fruit set in apple, indicating that there was competitive demand for the carbohydrate supply stored as reserves in the roots and other storage organs during early spring (Tustin et al. Citation1997).
Light is necessary for enabling the supply of photoassimilate and this has been shown to directly affect the quality of fruit in different locations within a tree. Apricot fruit had higher SSC in positions most exposed to light, but no data were provided to support this statement (Lichou et al. Citation1999). The effect of high light exposure on apricot fruit SSC has been confirmed from other trials (Dichio et al. Citation1999; Stanley et al. Citation2014). Lower and inner canopy positions had the lowest light interception, where apricot fruit weight and fruit SSC were also lowest (Stanley et al. Citation2014).
Therefore, the aim of these studies was to identify the key factors involved in variation in apricot fruit quality along a shoot or branch. The first study compared the variation of fruit set and fruit quality within branch units, compared among apricot genotypes. A second study examined the competition for resources within single shoots of one cultivar and how the location along the shoot affected the quality of that fruit at harvest. The hypothesis was that there would be patterns in fruit quality along an apricot branch that could be explained by wood age and resource competition, with genotype affecting the extent of such effects.
Materials and methods
Wood type classifications
For the purposes of these trials, wood was classified into the following wood types:
One-year-old spurs: terminal spurs that had very reduced internodes and were approximately 2–6 cm in length, and followed by 2-year-old wood of at least 15 cm in length (A).
One-year-old shoots: long shoots that had at least 15 cm of 1-year-old wood. These were strong shoots with diameters at the base of the 1-year-old wood typically greater than 9 mm (B). Sylleptic shoots were included as 1-year-old shoots as long as the total length of the longest shoot was more than 15 cm.
Spurs on 2-year-old wood: spurs of 1-year-old shoots that had very complex, short internodes, usually less than 5 cm in length, which were attached to 2-year-old wood (C).
Short shoots on 2-year-old wood: short shoots of 1-year-old wood less than 15 cm that were attached to 2-year-old wood. These were weak shoots with diameters at the base of the shoots of approximately 4–5 mm and they typically did not have sylleptic shoots (D).
Spurs on 3- or 4-year-old wood: spurs of 1-year-old shoots that had very complex, short internodes and in some cases had very short sections of 2- and 3-year-old shoots, which arose from 3- and 4-year-old wood, respectively (E). The total spur complex was usually less than 5 cm in length, but older spur complexes could sometimes be longer.
Short shoots on 3- or 4-year-old wood: short shoots of less than 15 cm that arose from 3- and 4-year-old wood, respectively. These were weak shoots with diameter at the base of the shoot of approximately 4–5 mm (F). Many of these may have been latent or adventitious buds that had only grown out in the previous year, but all were completely 1-year-old wood shoots or, if they included wood that was 2 or 3 years old, they were very short sections.
Trial 1. Variation of fruit quality within a branch among apricot genotypes
Trial layout
Five trees each of six genotypes from the Plant & Food Research breeding programme, thought to have differing fruit-bearing habits, were tagged on 5 September 2011. The trees were not randomised as they were established for breeding evaluations, therefore, the trees for any one genotype were adjacent to each other in the same row. The genotypes were: ‘Clutha Summer’, a recently released cultivar previously named D14/1; CM1/4; H3/3; StB6/6; StB7/1; and StB14/15. All trees were located at the Clyde Research Orchard, Earnscleugh, Central Otago, New Zealand, and were 5 years old in 2011 except CM1/4 trees, which were 6 years old. At this location, the chilling requirement for apricots is easily reached well before flowering every year. More than 1200 and 1100 Richardson chill units were accumulated by the end of July in 2011 and 2012, respectively. Two branches from each tree were tagged and the age of wood was marked. Branches that had minimal pruning were selected so that wood age was easy to trace along the branch. The location of flowers were categorised into two wood types for each wood age: shoots or spurs. This resulted in eight possible combinations (four ages of wood × two wood types).
Figure 1. Illustrations identifying: A, 1-year-old terminal spur; B, 1-year-old long shoot; C, spur on 2-year-old wood; D, short shoot on 2-year-old wood; E, spur on 3-year-old wood; F, short shoot on 3-year-old wood.

The fruit on the trees, including the tagged branches, were thinned according to commercial thinning practices (fruit spaced at least 6–8 cm apart) on 14 and 15 November 2011, while ensuring that some fruit from each wood age × wood type combination were retained. In the following year, the same numbers of branches were tagged for four of the genotypes: ‘Clutha Summer’, StB6/6, StB7/1 and StB14/15. Fruit on the trees were thinned in a similar manner on 30 October 2012. The results from the 2 years are referred to by the year in which the fruit were harvested i.e. 2012 and 2013, respectively.
Measurements
The numbers of flowers were counted for each combination on each tagged branch between 5 and 9 September 2011 and between 12 and 14 September 2012. The numbers of fruit that had set were counted for each combination on each tagged branch on 27 and 28 October 2011 and on 23 October 2012. The proportional fruit set was then calculated for each branch before thinning. For each genotype, all fruit on tagged shoots were inspected on two or three dates in January 2012 and January 2013. On each date, those that were at commercial maturity, based on skin background colour, were harvested for assessment. Fruit were weighed individually. Flesh firmness was measured on opposite sides of each fruit along the equatorial zone using a 7.9 mm diameter probe attached to a GUSS FTA penetrometer (GUSS Manufacturing Ltd) after a small circle of skin had been removed. Penetration speed, trigger force and penetration distance were set to 10 mm s−1, 0.5 Newtons (N) and 8 mm, respectively. Data were calculated as the mean of measurements from a fruit sample and expressed in Newtons. Soluble solids concentration of expressed juice was measured using a digital refractometer (Atago). Dry matter concentration (DMC) was determined by taking the fresh and dry weight of two vertical slices from opposite sides of the fruit (including skin and flesh but excluding the stone).
In addition, five fruit of commercial maturity were harvested from each tree for each of five combinations: 1-year-old shoots, shoots and spurs on 2-year-old wood, and shoots and spurs on 3-year-old wood. These fruit were stored in air at 0 °C for 3 weeks and then at 20 °C for 4 days before postharvest assessment. Each fruit was assessed for gel formation and rubberiness. The gel score was based on the proportion of flesh that had a glassy appearance: from 0 (no glassy flesh) to 3 (>50% glassy flesh). Rubberiness, or leatheriness, was considered to be a rubbery texture of the intact fruit. Leatheriness in peaches has been described as a dry texture when fruit have a flesh firmness of >30 N (Ju et al. Citation2000). Rubberiness was scored on a scale from 0 (not rubbery) to 3 (very rubbery). A score of 3 was given when the penetrometer did not penetrate the flesh on both sides of the fruit, but ‘bounced’ off the flesh surface. Each fruit was tasted by at least two of three trained assessors for mealiness and juiciness. Mealiness was considered to be a woolly or lumpy texture and fruit were scored on a scale from 0 (no mealiness) through to 3 (high mealiness). Juiciness was considered to take into account the amount of free fluid released from the sample during chewing and fruit were scored for juiciness on a scale from 0 (no juice) through to 3 (very juicy).
The total number of fruit assessed at harvest was 1222 and 754 and after cold storage was 500 and 372 for 2012 and 2013, respectively.
Trial 2. Effect of competition for resources and location along shoots on fruit quality
Trial layout
Ten shoots of similar length of 2-year-old wood (mean length 112.2 mm, standard error of mean [SEM] 3.4 mm; basal shoot diameter 14.4 mm, SEM 0.4 mm) and 10 shoots of similar length of 1-year-old wood (mean length 69.2 mm, SEM 3.4 mm; basal shoot diameter 7.6 mm, SEM 0.2 mm) were tagged at flowering on each of 10, 17-year-old ‘CluthaGold’ apricot trees at the Clyde Research Orchard, Earnscleugh, Central Otago, New Zealand. Spurs but not short shoots were present on the 2-year-old wood and no sylleptic shoots were present on any shoots. The 1-year-old shoots had no 2-year-old wood section along the same shoot, i.e. they had grown out as laterals from older wood the previous season. The number of fruit on the 2-year-old wood and the 1-year-old wood were counted on 25 and 26 October 2012, respectively.
Fruit on six of the shoots of 2-year-old wood per tree were thinned to form a range of thinning patterns, leaving either three or six fruit per shoot (i.e. two different crop loads), on 30 October 2012. The thinning patterns were: three or six fruit were distributed in the basal half of the 2-year-old wood, three or six fruit were distributed evenly along the whole 2-year-old wood section of the shoot, or three or six fruit were distributed in the distal half of the 2-year-old wood section of the shoot. Very few fruit set on the 1-year-old wood sections along the shoots with our 2-year-old wood treatments, and all remaining fruit on the 1-year-old wood were removed. Each treatment combination (thinning pattern × no. fruit) occurred on each of 10 trees once. Fruit on six shoots of the 1-year-old wood per tree were thinned to form a similar range of thinning patterns, but leaving only two fruit per shoot, on 30 October 2012. The thinning patterns of fruit distribution were the same as for the 2-year-old wood shoots i.e. two fruit were left in the basal half of the 1-year-old wood, one fruit was left in the basal half and one fruit left in the distal half of the 1-year-old wood, or two fruit were left in the distal half of the 1-year-old wood. Each thinning pattern occurred twice on each of the same 10 trees as for the 2-year-old wood shoots. The remaining fruit on the trees were thinned according to commercial thinning practices, spacing fruit out to be approximately 5–8 cm apart.
Measurements
The distance from the base of each shoot to each fruit and the total length of 1-year-old wood and/or 2-year-old wood per shoot were measured. All fruit on tagged shoots were assessed on 25 January, 29 January, 31 January and 7 February 2013 and those that were at commercial maturity were harvested for assessment. All fruit were harvested on one of these four dates. The total number of fruit assessed was 304. Fruit were weighed and SSC and DMC were determined, as described for Trial 1.
Once fruit were harvested, the leaf number and leaf area of each shoot arising from a fruiting node was estimated. The leaf numbers and total node numbers were recorded for each shoot and the leaves kept for leaf area measurement. The leaves for each shoot were scanned on a photocopier along with a ruler for scale and saved as a pdf file. The leaf areas were calculated using the software ImageJ (Reinking Citation2007).
Statistical analysis
A combined set of the two years of data was used for analysis of variance (ANOVA) models for Trial 1. Data were aggregated for each wood type × wood age combination on each branch of every tree for each year. As the data were unbalanced because there were fewer genotypes and only three ages of wood in 2013, ANOVA was performed first to assess the main effects. The interactions between factors were then assessed using ANOVA on genotype × wood age × wood type × year for main effects and two-way interactions. Estimated denominator degrees of freedom for Trial 1 were high (300–400 for harvest data and approximately 150 for storage data). For all analyses, any three- and four-way interactions assessed were not significant and were therefore left out.
In Trial 2, ANOVA was performed using the full data set (individual fruit), to consider the main effects and two-way interactions between position of the fruit along the shoot × the fruiting spur leaf area for 1-year-old wood and 2-year-old wood separately. A contour plot of FW was drawn using the contour function within R 2.15.0 (R Development Core Team Citation2012) based on the ANOVA results for the interaction between position of the fruit along the shoot × the fruiting spur leaf area on 1-year-old wood. Data were then aggregated for each wood age by obtaining the means of each trait for fruit from each shoot and ANOVA performed for main effects and two-way interactions. FWs were grouped into three categories containing equal numbers of samples based on their number of fruit pre-thinning, and the mean FW was calculated for each grouping.
All linear mixed models were fitted using the asreml package (Butler et al. Citation2009) within R 2.15.0 (R Development Core Team Citation2012). Many of the ANOVA results are presented as dot plot grids for easy trend comparisons and for space-saving reasons. Dot plot grids are horizontal bar graphs in which the bar is separated from the standard error by a small gap for easy recognition of the error range.
Results
Trial 1
Effect of bud type and wood age on fruit set
Genotype significantly affected fruit set depending on wood type, wood age and year (). Fruit set on 1-year-old wood was lower than on the other three wood ages for CM1/4 and lower than 2- and/or 3-year-old wood for ‘Clutha Summer’, StB14/15 and StB6/6, but was similar for different wood ages in most other cases (). This interaction was influenced by year, but the three-way interaction was not significant. Although the interaction between genotype and wood age was significant (P = 0.05), the high degrees of freedom in the analysis suggests that the interaction was not important (data not presented). Fruit set was lower on 1-year-old long shoots than on 1-year-old terminal spurs (). There were no differences in fruit set among shoots or spurs on 2-year-old, 3-year-old wood or 4-year-old wood. Fruit set was much lower in 2012 than in 2013 for ‘Clutha Summer’, StB14/15 and StB7/1 in almost all cases (), possibly caused by five frosts during flowering (first 10 days in September) in the 2012 season whereas there were no frosts during flowering in the 2013 season.
Figure 2. Interaction between genotype and wood age for fruit set of six apricot genotypes during two fruiting seasons (solid line: 2012; dotted line: 2013) in three-way ANOVA. Error bars represent ± standard error of the mean.
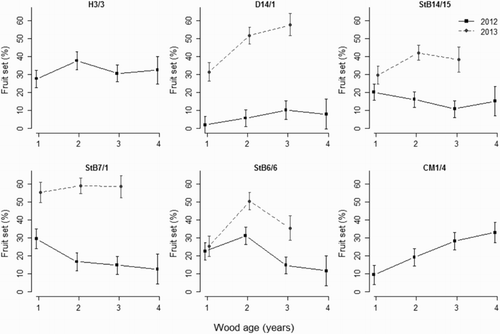
Figure 3. Effect of interaction between wood age and wood type (solid lines: shoots; dotted lines: spurs) on fruit set of six apricot genotypes during two fruiting seasons (squares: 2012; circles: 2013) in three-way ANOVA analysis. Error bars represent ± standard error of the mean.
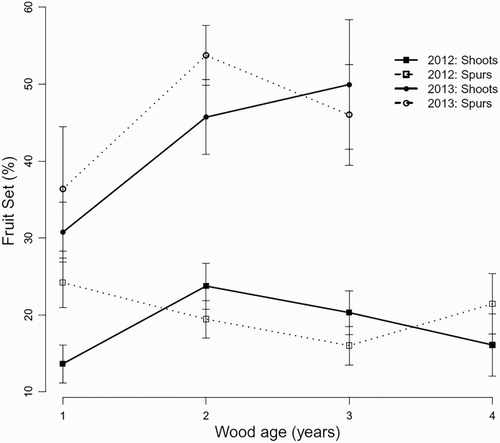
Table 1. ANOVA P-values for main effects and two-way interactions between genotype × wood age × wood type × year for fruit set, fresh weight, soluble solids concentration (SSC) and dry matter concentration (DMC) of apricot fruit for six genotypes at commercial harvest. ANOVA P-values for main effects and two-way interactions between genotype × wood age × wood type × year for mealiness score, rubbery score and flesh firmness of apricot fruit after cold storage at 0 °C for 3 weeks and shelf-life at 20 °C for 4 days of apricot fruit for four genotypes.
Effect of bud type and wood age on fruit quality at harvest
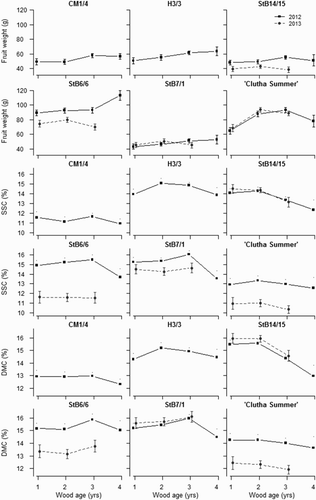
Genotype affected FW depending on wood age and year (). FW was lower from 1-year-old wood of ‘Clutha Summer’ than from older wood, whereas FW was highest from 4-year-old wood for StB6/6 and there was a trend for increasing FW on older wood for CM1/4, H3/3 and StB7/1 (). Fruit from spurs were heavier (65.6 g ± 1.3) than from shoots (61.7 g ± 1.3) (). Year and genotype affected FW, SSC and DMC, and wood age also affected SSC and DMC for some genotypes (; ). All fruit were harvested within the commercial maturity range; however, those of ‘Clutha Summer’, StB6/6 and StB7/1 were harvested at a slightly more mature stage in 2012 than in 2013 (flesh firmness was 10 to 15 N lower in 2012) and fruit SSC was higher in 2012.
Figure 4. Effect of interaction between genotype, wood age and year on flesh firmness, soluble solids concentration (SSC) and dry matter concentration (DMC) of apricots of six genotypes at harvest during two fruiting seasons (solid line: 2012; dotted line: 2013) in three-way ANOVA. Error bars represent ± standard error of the mean.
Effect of bud type and wood age on fruit quality after cold storage
Mealiness score, rubbery score, gel score and juiciness scores for fruit assessed after cold storage were all affected by genotype and year (). Some genotypes had much greater mealiness scores, rubbery scores or gel scores than other genotypes, but often the severe symptoms were observed in one year only (). The year in which the symptoms of rubberiness were greater, differed for StB14/15 and StB6/6 (). Rubberiness was significantly less for fruit on 2-year-old wood in 2012 than in 2013 or on 3-year-old wood in either year (). Gel development was greater on 2-year-old shoots than on 3-year-old shoots or spurs, whereas 3-year-old shoots and spurs were similar in gel development to 1-year-old shoots and 2-year-old spurs ().
Figure 5. Effect of genotype × year on mealiness score, rubbery score, gel score and juiciness score of fruit of four apricot genotypes after 3 weeks at 0 °C and 4 days at 20 °C in two-way ANOVA. A higher score indicates greater mealiness, rubberiness, gel formation and juiciness. Error bars represent ± standard error of the mean.
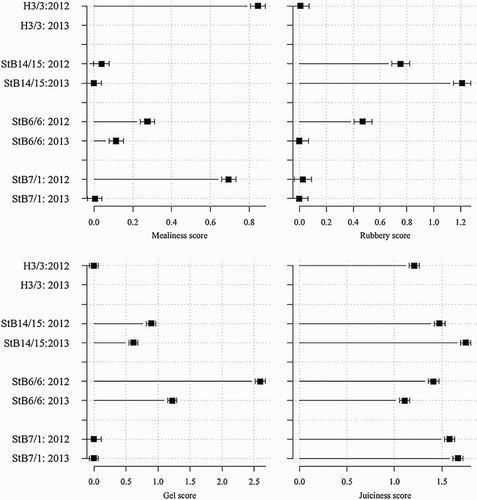
Table 2. Effects of wood age × year on rubbery score ± standard error of the mean, and wood age × shoot type on gel score ± standard error of the mean, of fruit of four apricot genotypes after 3 weeks at 0 °C and 4 days at 20 °C in two-way ANOVA. A higher score indicates greater rubberiness and gel formation.
Trial 2
Effect of position along shoot and wood age on fruit quality
Fruit from mid and distal regions of 1-year-old wood had lower FW than those from basal regions and from all positions on 2-year-old wood (P < 0.001) (). Fruit from the basal region of 1-year-old shoots had higher FW when the leaf area from that node was large e.g. >40 mm2 (). In contrast, fruit from the distal region of 1-year-old shoots had lower FW when the leaf area from that node was greater. Leaf area of fruiting nodes, and its interaction with position along the shoot, had no effect on fruit weight of fruit on 2-year-old shoots (A). Leaf area from fruiting nodes was greater on 2-year-old wood than on 1-year-old wood (P < 0.001), but position along the shoot had no effect on leaf area of the fruiting node (data not shown).
Figure 6. Effect of position along the shoot, wood age and the interaction between position along the shoot × wood age on fruit fresh weight, soluble solids concentration and dry matter concentration of ‘CluthaGold’ apricots in two-way ANOVA. Error bars represent ± standard error of the mean (1 YOW: 1-year-old wood; 2 YOW: 2-year-old wood).
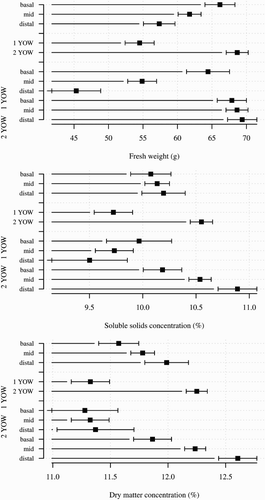
Figure 7. Contour plot showing the fitted trend surface for fruit fresh weight (represented by the contour lines in grams) based on the relationship between leaf area at the fruiting node immediately after fruit harvest (mm2) and the position of the fruit along the shoot (distance from shoot base as a percentage of length) on 1-year-old wood of ‘CluthaGold’ apricots. Raw data points are shown in grey.
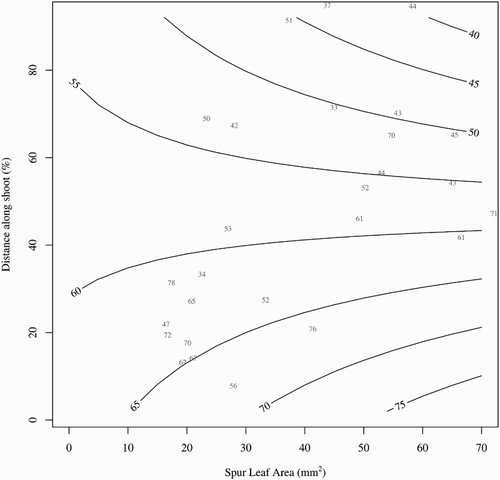
Table 3. A, ANOVA P-values for main effects and two-way interactions between position along the shoot × leaf area at the fruiting node, for fruit fresh weight for ‘CluthaGold’ apricot fruit for each shoot on either 1-year-old wood (1 YOW) or 2-year-old wood (2 YOW). B, ANOVA P-values for main effects and two-way interactions between the number of fruit before fruit thinning × thinning pattern on fresh weight of ‘CluthaGold’ apricot fruit using data aggregated for each shoot on either 1 YOW or 2 YOW.
Fruit from 1-year-old wood had lower SSC and DMC than those from 2-year-old wood, except basal fruit from both 1-year-old and 2-year-old regions had similar SSC (). SSC and DMC were higher in fruit located more distally along 2-year-old wood. Leaf area at the fruiting node had no effect on fruit SSC or DMC (data not presented).
Effect of early season competition on fruit quality
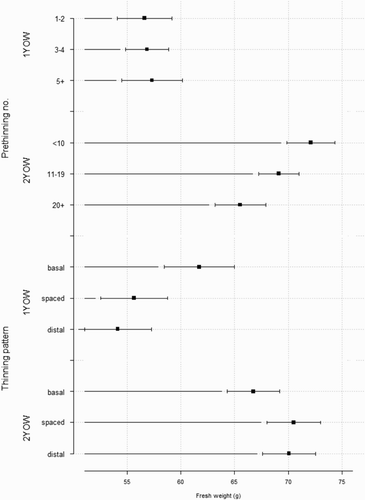
Fewer fruit on 2-year-old wood before thinning resulted in higher FW but this response was not evident on 1-year-old wood (B, ). Fruit on 1-year-old shoots that were thinned to the basal region had greater FW than fruit from shoots thinned to the distal region, or than fruit from shoots thinned uniformly along the shoot length (). There was no effect of thinning pattern on FW of fruit on 2-year-old wood. Number of fruit pre-thinning and thinning patterns had no significant effects on SSC or DMC (data not presented).
Figure 8. Effect of the number of fruit pre-thinning and thinning pattern on fruit fresh weight of ‘CluthaGold’ apricots in two-way ANOVA using data aggregated for each shoot on either 1-year-old wood (1 YOW) or 2-year-old wood (2 YOW). Data were grouped into three categories containing equal numbers of samples based on their number of fruit pre-thinning, and the mean FW was calculated for each grouping. Thinning patterns were: basal half only, evenly spaced along shoot or distal half only. Fruit were thinned on 30 October to two fruit for 1 YOW and to three or six fruit for 2 YOW. Error bars represent ± standard error of the mean.
Discussion
Effect of genotype
Genotype affected fruit set, confirming previous observations on apricot (Alburquerque et al. Citation2004; Cirillo et al. Citation2010). In contrast to earlier results, fruit set in New Zealand was affected by season as much as it was by genotype, possibly because of the variable and damaging temperatures that can be experienced during flowering. Selections in breeding programmes that consistently have low fruit set may not be suitable for commercial production (Alburquerque et al. Citation2004).
Fruit quality was significantly affected by the genotype, not only FW, SSC and DMC at harvest, but also attributes after cold storage, including mealiness, gel formation, rubberiness and juiciness. This emphasises the importance of selecting cultivars that produce fruit with inherently high-quality traits. To identify a genotype that has all fruit quality traits within an acceptable range is challenging to achieve, and some traits may be more important than others. Consumers found sweetness, aroma and texture (firmness and lack of mealiness) to be of greatest importance (Gatti et al. Citation2009). Therefore, the best guide that a new cultivar has an acceptable combination of fruit quality traits is probably through consumer preference studies (Gatti et al. Citation2009). However, even once a cultivar has been selected and planted, there are significant variations in fruit quality within a tree, as these results and those of others have shown (Lichou et al. Citation1999; Stanley et al. Citation2014).
Effect of wood age and wood type on fruit set
Fruit set was higher on spurs on 2-year-old wood and 3-year-old wood than on shoots on 1-year-old wood, but there were no differences in fruit set between spurs and short shoots on 2-year-old wood or older. A similar result was found for the apricot cultivar ‘Ninfa’, although fruit set was higher on spurs than on either 1-year-old shoots or short shoots on older wood (Cirillo et al. Citation2010). One possible cause of these differences could be the starch content in the ovaries and ovules of flowers, which was found to be higher in short shoots than on 1-year-old shoots (Julian et al. Citation2010a). Lower fruit set on long shoots compared with short shoots in the cultivar ‘Moniqui’ was related to a higher incidence of underdeveloped pistils (Julian et al. Citation2010b). Other possible causes to reduced fruit set on younger wood include late flower initiation, which reduced the effective pollination period and flower dry weight in apple (Robbie & Atkinson Citation1994), and flower receptacle weight at full bloom (Volz et al. Citation1994; Julian et al. Citation2010b). Higher flower weight on spurs may be related to earlier flower initiation prior to dormancy (Sakayarote et al. Citation2005) and a lower chilling requirement for spurs than for long shoots (Austin et al. Citation1992), enabling earlier flower development in spring. Lower fruit set on 1-year-old long shoots may also have been related to greater competition from shoot apices about to emerge.
Effect of wood age and wood type on fruit quality
In general, fruit on shoots and spurs arising from 2-year-old wood and the base of 1-year-old long shoots had the highest FW, SSC and DMC.
Poorer fruit quality on older wood may be a result of being located nearer the centre of the tree. Fruit FW, SSC and DMC have been shown to be less in lower regions of the tree in stonefruit, where light transmission was lower (Luchsinger et al. Citation2002; Stanley et al. Citation2014). Artificial shading during fruit growth stage I early in the season, reduced FW in peach; however, it was shading in growth stage III close to harvest that reduced SSC (George et al. Citation1996).
Fruit FW, SSC and DMC of some genotypes were not significantly different or were higher in fruit from 2-year-old wood than those from 1-year-old wood. The 1-year-old shoots in our trials were closer to the outside of the canopy and more likely to be in the higher light environment. However, even in a higher light environment, FW, SSC and DMC of fruit from 1-year-old shoots, for ‘CluthaGold’ and many of the genotypes in Trial 1, were lower than from 2-year-old fruit, evidence that light was not the only factor involved.
A possible explanation for the poorer fruit quality nearer the apex of the shoot could be the close proximity of the fruit to the growing shoot apex, which may compete for resources at peak growth periods. The shoot apex has been shown to be the strongest growth sink for peach and Japanese apricot (Prunus mume Sieb. et Zucc.) in the early season (Corelli Grappadelli et al. Citation1996; Tsuchida et al. Citation2011), when cell division is also occurring in most fruits (Jackson & Coombe Citation1966; Yamaguchi et al. Citation2002). This period of fruit growth in the early season is also a critical contributor to determining final fruit size in apple (Stanley et al. Citation2000), and fruit size at pit hardening correlated with final fruit size in apricot (Jackson & Coombe Citation1966). This theory matches the results in this trial and is further supported by results for early maturing peach cultivars, in which the most rapid periods of fruit growth and shoot growth coincided, resulting in lower fruit FW (Dejong et al. Citation1987).
Leaf area and the development of axillary shoots may also be important. Initial size of leafy shoots was found to be one of the important factors that affected peach fruit size (Genard et al. Citation1998). However, in cherry, in which fruit matures even more rapidly than apricot and peach, carbon labelling showed that fruit close to the carbon source were very strong sinks compared with shoot growth, but higher leaf to fruit ratio reduced the proportion of 13C that proximal fruit received (Ayala & Lang Citation2008). Studies of labelled carbon distribution from leaves in different positions along the shoot at different times during fruit growth would be required to provide a greater understanding of these dynamics.
Effect of wood age and wood type on chilling injury
Differences in chilling injury were observed in cold-stored fruit, among genotypes, between years, on different aged wood, and between spurs and short shoots in some cases. The location of the different aged wood in the canopy may be affecting these differences in postharvest chilling injury, as previously noted in peach (Crisosto et al. Citation1997). Nutrition of the fruit may be involved, as found with canopy location affecting calcium content in apples, which led to differences in bitter pit development during cold storage (Volz et al. Citation1993). Canopy position also affects fruit temperature and this has been associated with chilling injury in avocado (Woolf et al. Citation1999). The relationship between pre-harvest factors and chilling injury in cold-stored apricots needs further evaluation, in particular focusing on exposure to high temperatures prior to harvest and nutrient status of fruit.
Effect of crop load on the shoot
A smaller number of initial fruit on a shoot resulted in greater fruit FW, therefore, if flower numbers are high, a lower fruit set may be a desirable trait. However, if fruit set was too low, yields could be poor (Alburquerque et al. Citation2004). In general, higher fruit set can be adjusted at thinning time, to achieve a desirable fruit density. In Trial 1, while fruit set in the first season was low, the resulting final fruit density was similar to that in the second season, when the fruit set was higher. Until the fruit were thinned, however, the crop load per shoot would have been greater in the second year than in the first year. Fruit FW was greater in two of the four genotypes, indicating that early season crop load had an effect on FW. The lack of an effect on FW in the other two genotypes suggests that some genotypes have an ability to compensate better than others, possibly because of different growth habits (Muleo et al. Citation2006).
The negative effect of higher initial fruit numbers per shoot on FW has implications for those cultivars that have high fruit set and earlier fruit maturity. While high fruit set may be seen as a positive attribute for apricots as a safeguard against low yields after late spring frosts, it had a negative effect on fruit weight in this trial. Greater competition in the early season could be reduced to some extent by earlier thinning (Rettke & Dahlenburg Citation1999).
Practical implications
These results have implications for orchard management to improve fruit quality and uniformity within apricot trees. Possible techniques that could utilise knowledge of the variation include training and pruning to favour fruit on 2- and 3-year-old wood. Methods that reduce shading in lower canopy positions, for example utilising reflective cloth (Stanley et al. Citationin press) or removing larger limbs, would increase FW and SSC in inner canopy fruit. Other possibilities include differential thinning to reduce fruit numbers on distal regions of 1-year-old wood and basal regions on older wood. This could be combined with successful techniques that involved thinning to the same overall crop load, but reducing the proportion of fruit from lower canopy, shady locations and retaining more fruit in upper-canopy, high-light locations. This method resulted in an increase in the proportion of fruit that had higher FW and SSC (Stanley et al. Citationin press). Earlier thinning would reduce competition among fruit sooner. The longer-term goal would be to redesign training and pruning systems to favour certain wood types for each cultivar.
Conclusions
Genotype affected fruit set and fruit quality, both at harvest and during cold storage. Apricot fruit located on 2-year-old wood or 3-year-old wood had better fruit quality, in terms of greater FW, SSC and DMC, and fruit set was higher, than on 1-year-old long shoots or older wood, for most genotypes. For ‘CluthaGold’, fruit had higher FW, SSC and DMC when located on the distal end of 2-year-old wood or the basal end of 1-year-old wood. Greater fruit numbers prior to commercial thinning reduced final FW on fruit arising off 2-year-old wood. Possible underlying factors affecting fruit set and fruit quality are likely to be associated with competition for carbon resources and light interception. Further research is needed to separate the effects of shading and carbohydrate competition on apricot fruit quality. In addition, our results indicated there may be some effect of pre-harvest factors on chilling injury during cold storage.
Acknowledgements
Thanks to Ross Marshall and Claire Scofield, who assisted with data collection and Peter Alspach, who advised on statistical analyses. This paper has been adapted from a chapter in a thesis by the author entitled ‘Control of pre-harvest and postharvest factors to improve fruit quality of apricot’, which was submitted and conferred at Griffith University, Australia, in 2015.
Disclosure statement
No potential conflict of interest was reported by the author.
Additional information
Funding
References
- Alburquerque N, Burgos L, Egea J. 2004. Influence of flower bud density, flower bud drop and fruit set on apricot productivity. Sci Horti. 102:397–406. doi: 10.1016/j.scienta.2004.05.003
- Austin PT, Atkins TA, Plummer JA, Noiton DA, Hewett EW. 1992. Influence of shoot-type on time of apricot flowering. Acta Horti. 313:325–330. doi: 10.17660/ActaHortic.1992.313.39
- Ayala M, Lang GA. 2008. 13C-photoassimilate partitioning in sweet cherry on dwarfing rootstocks during fruit development. Acta Horti. 795:625–632. doi: 10.17660/ActaHortic.2008.795.97
- Azodanlou R, Darbellay C, Luisier JL, Villettaz JC, Amado R. 2003. Development of a model for quality assessment of tomatoes and apricots. Lebensmittel-Wissenschaft Und-Technologie-Food Sci Technol. 36:223–233. doi: 10.1016/S0023-6438(02)00204-9
- Butler DG, Cullis BR, Gilmour AR, Gogel BJ. 2009. ASReml-R reference manual. Brisbane, QLD: Department of Primary Industries and Fisheries; [cited 2016 May 23]. Available from: http://www.vsni.co.uk/downloads/asreml/release3/asreml-R.pdf
- Cirillo C, Basile B, Hernandez G, Pannico A, Giaccone M, Forlani M. 2010. Influence of fruiting shoot on flowering pattern and fruit quality of Vesuvian apricot cultivars. Acta Horti. 862:557–564. doi: 10.17660/ActaHortic.2010.862.88
- Corelli Grappadelli L, Coston DC. 1991. Thinning pattern and light environment in peach-tree canopies influence fruit-quality. HortScience. 26:1464–1466.
- Corelli Grappadelli L, Ravaglia G, Asirelli A. 1996. Shoot type and light exposure influence carbon partitioning in peach cv Elegant Lady. J Hortic Sci. 71:533–543. doi: 10.1080/14620316.1996.11515433
- Crisosto CH, Johnson RS, DeJong T, Day KR. 1997. Orchard factors affecting postharvest stone fruit quality. HortScience. 32:820–823.
- Dejong TM, Doyle JF, Day KR. 1987. Seasonal patterns of reproductive and vegetative sink activity in early and late maturing peach (Prunus persica) cultivars. Physiol Plantarum. 71:83–88. doi: 10.1111/j.1399-3054.1987.tb04621.x
- Dichio B, Celano G, Montanaro G, Nuzzo V, Xiloyannis C, Giandomenico I. 1999. Leaf area evolution, light interception, yield and quality of fruits in apricot trees (cultivar Tirynthos) trained to transverse Y and vase. Acta Hortic. 488:527–532. doi: 10.17660/ActaHortic.1999.488.85
- Gatti E, Defilippi BG, Predieri S, Infante R. 2009. Apricot (Prunus armeniaca L.) quality and breeding perspectives. J Food Agric Environ. 7:573–580.
- Genard M, Lescourret F, Ben Mimoun M, Besset J, Bussi C. 1998. A simulation model of growth at the shoot-bearing fruit level. II. Test and effect of source and sink factors in the case of peach. Eur J Agron. 9:189–202. doi: 10.1016/S1161-0301(98)00036-7
- George AP, Hieke S, Rasmussen T, Ludders P. 1996. Early shading reduces fruit yield and late shading reduces quality in low-chill peach (Prunus persica (L) Batsch) in subtropical Australia. J Hortic Sci. 71:561–571. doi: 10.1080/14620316.1996.11515436
- Jackson DI, Coombe BG. 1966. The growth of apricot fruit. I. Morphological changes during development and the effects of various tree factors. Aust J Agr Res. 17:465–477. doi: 10.1071/AR9660465
- Ju ZG, Duan JS, Ju ZQ. 2000. Leatheriness and mealiness of peaches in relation to fruit maturity and storage temperature. J Hortic Sci Biotech. 75:86–91. doi: 10.1080/14620316.2000.11511205
- Julian C, Rodrigo J, Herrero M. 2010a. Apricot flower bud development and fruit set in different types of shoots in ‘Moniqui’ variety. Acta Hortic. 862:337–342. doi: 10.17660/ActaHortic.2010.862.52
- Julian C, Herrero M, Rodrigo J. 2010b. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca L.). Trees. 24:833–841. doi: 10.1007/s00468-010-0453-6
- Lichou J, Jay M, Combe J. 1999. Variation of apricot quality - influence of fruit location within the tree. Acta Hortic. 488:539–542. doi: 10.17660/ActaHortic.1999.488.87
- Luchsinger L, Ortin P, Reginato G, Infante R. 2002. Influence of canopy fruit position on the maturity and quality of ‘Angelus’ peaches. Acta Hortic. 592:515–521. doi: 10.17660/ActaHortic.2002.592.69
- Marini RP, Sowers DL. 1994. Peach fruit weight is influenced by crop density and fruiting shoot length but not position on the shoot. J A Soc Hortic Sci. 119:180–184.
- Muleo R, Remorini D, Iacona C, Guerriero R, Massai R. 2006. Cultivar-specific adaptation of growth and photosynthetic activity in developing apricot dense canopies. Acta Hortic. 717:55–58. doi: 10.17660/ActaHortic.2006.717.7
- R Development Core Team. 2012. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Available from: http://www.R-project.org/
- Reinking L. 2007. Examples of image analysis using ImageJ. Millersville, PA: Millersville University; [cited 2015 May 23]. Available from: http://rsb.info.nih.gov/ij/docs/pdfs/examples.pdf
- Rettke MA, Dahlenburg AP. 1999. Effect of timing of hand thinning on productivity of Moorpark apricots destined for drying. Aust J Exp Agr. 39:885–889. doi: 10.1071/EA99018
- Robbie FA, Atkinson CJ. 1994. Wood and tree age as factors influencing the ability of apple flowers to set fruit. J Hortic Sci. 69:609–623. doi: 10.1080/14620316.1994.11516493
- Robinson TL, Seeley EJ, Barritt BH. 1983. Effect of light environment and spur age on delicious apple fruit size and quality. J Am Soc Hortic Sci. 108:855–861.
- Sakayarote K, Krisanapook K, Pichakum A, Intharasri W. 2005. Time of floral initiation and flowering of Japanese apricots at the Royal Agricultural Station Angkhang. ACIAR Tech Rep Ser. 61: 10–112.
- Stanley J, Marshall R, Scofield C, Alspach P. in press. The effects of reflective cloth and adjusted fruit distribution on apricot fruit quality. Acta Hortic.
- Stanley J, Marshall R, Tustin S, Woolf A. 2014. Preharvest factors affect apricot fruit quality. Acta Hortic. 1058:269–276. doi: 10.17660/ActaHortic.2014.1058.31
- Stanley CJ, Tustin DS, Lupton GB, McArtney S, Cashmore WM, De Silva HN. 2000. Towards understanding the role of temperature in apple fruit growth responses in three geographical regions within New Zealand. J Hortic Sci Biotechnol. 75:413–422. doi: 10.1080/14620316.2000.11511261
- Summerfruit New Zealand. 2011. Summerfruit New Zealand shopper behaviour study 2011. Nielsen Homescan survey. Wellington: Summerfruit New Zealand; [cited 2016 May 23]. Available from: http://www.summerfruitnz.co.nz/users/Image/Files/PDFs/2011%20Final%20Summerfruit%20Report%20Shopper%20Behaviour%207%20pages%20.pdf
- Tsuchida Y, Yakushiji H, Negoro K, Hishiike M. 2011. Characteristics of assimilated carbohydrate competition between fruit and shoot, and effect of fruit load on assimilted carbohydrate translocation and tree growth of Japanese apricot (Prunus mume Sieb. et Zucc.) ‘Nanko’. Hortic Res (Japan). 10:551–558. doi: 10.2503/hrj.10.551
- Tustin DS, Stanley CJ, Adams HM. 1997. Physiological and phenological responses of apple trees to artificial reduction of the growth period from harvest to leaf fall. Acta Hortic. 451:383–404. doi: 10.17660/ActaHortic.1997.451.45
- Volz RK, Ferguson IB, Bowen JH, Watkins CB. 1993. Crop load effects on fruit mineral-nutrition, maturity, fruiting and tree growth of cox orange pippin apple. J Hortic Sci. 68:127–137. doi: 10.1080/00221589.1993.11516336
- Volz RK, Ferguson IB, Hewett EW, Woolley DJ. 1994. Wood age and leaf-area influence fruit size and mineral-composition of apple fruit. J Hortic Sci. 69:385–395. doi: 10.1080/14620316.1994.11516468
- Woolf AB, Bowen JH, Ferguson IB. 1999. Preharvest exposure to the sun influences postharvest responses of ‘Hass’ avocado fruit. Postharvest Biol Tec. 15:143–153. doi: 10.1016/S0925-5214(98)00077-5
- Yamaguchi M, Haji T, Miyake M, Yaegaki H. 2002. Varietal differences in cell division and enlargement periods during peach (Prunus persica Batsch) fruit development. J Jpn Soc Hortic Sci. 71:155–163. doi: 10.2503/jjshs.71.155