
ABSTRACT
Pear fruit are subject to sun damage from high fruit surface temperature (FST). Shade netting has been shown to decrease FST and sunburn damage of apples but also decreased fruit colour. The aim of this study was to investigate the effects of netting on FST, fruit sunburn and fruit colour of the red-blushed pear cultivar ‘ANP-0131’. FST was measured using fine-wire thermocouples inserted into 54 fruit in a netted block of trees and into 56 fruit in adjoining rows of non-netted trees. Netting reduced the maximum FST by 10%. FST under netting did not exceed the reported sunburn damage threshold of 47°C despite air temperature reaching 38.6°C. The maximum FST in the non-netted site regularly exceeded the sunburn damage threshold and reached a maximum of 50.3°C. Assessments of sunburn damage showed a clear decrease in sunburn (amount and severity) for fruit from the netted trees; however, netting negatively affected colour intensity and coverage.
Introduction
Pear breeding programmes throughout the world have released new fresh market cultivars that are premium quality and aim to increase world consumption of pears. Most of the selections are either red-blushed (eg ‘Celina’, ‘Gem’ and ‘ANP-0131’) or full-red peel cultivars (eg PremP009) that are more popular with consumers (Human Citation2013). Australian growers, predominantly in the Goulburn Valley, are switching to these new cultivars and removing traditional plantings of ‘Williams’ Bon Chretien’ and ‘Packham’s Triumph’. Moving away from high yields of green peel cultivars brings new challenges, in particular, the need for good colour to meet market expectations without any bleaching, browning or necrosis from high fruit surface temperature (FST).
Exposure of fruit to direct solar radiation (DSR) is the primary cause of elevated FST and resultant sun damage. Sun damage of apples has been extensively studied (see review by Racsko and Schrader Citation2012). A more recent study in red-blushed pear established threshold FSTs of 47.1°C and 50°C for sunburn browning and necrosis, respectively (McClymont et al. Citation2016). In addition, high FST has been reported to suppress anthocyanin synthesis in apple (Ubi Citation2004) and degrade anthocyanins in pear (Steyn et al. Citation2004). On the other hand, solar radiation is essential for anthocyanin synthesis in many fruits including pear (Dussi et al. Citation1995; Zoratti et al. Citation2014). Unlike other fruits, the highest anthocyanin concentrations in full-red and red-blushed Pyrus communis cultivars are generally observed early in the season (Steyn et al. Citation2005). Later in the season, high FST can cause red colour to fade and play a critical role in fruit quality at harvest.
Many of the new red-blush cultivars are being grown using dwarfing rootstocks, tree densities (>2000 trees/ha) and tree training systems (eg Open Tatura trellis, van den Ende Citation2011) to increase early bearing and hence profitability. These management systems combined with increased fruitfulness result in greater exposure of fruit to DSR. Initial colour development will more than likely be enhanced compared to traditional systems but radiation exposure later in the season will increase the risk of colour fading and sun damage.
Management strategies devised to minimise sun damage, such as evaporative cooling systems, shading by netting and spray-on protectants, are all effective to some extent in decreasing FST and sunburn damage of apples (Evans et al. Citation1995; Glenn et al. Citation2002; Dussi et al. Citation2005; Gindaba and Wand Citation2005; Iglesias et al. Citation2005; Iglesias and Alegre Citation2006; Schrader Citation2011; Darbyshire et al. Citation2015). However, negative impacts of netting on apple colour development and fruit quality have been reported (Gindaba and Wand Citation2005; Iglesias and Alegre Citation2006). Wand et al. (Citation2005) described the use of evaporative cooling to improve colour development and reduce sunburn of red-blush pears and concluded that evaporative cooling could be effective in reducing sunburn of pears. Colavita et al. (Citation2011) reported that kaolin sprays were effective in decreasing FST of ‘Packham’s Triumph’ pears and decreasing sunburn of ‘Packham’s Triumph’ and ‘Beurré d’Anjou’. To date, the effect of netting on the incidence of sun damage in pears has not been reported.
The purpose of this study was to investigate the effectiveness of netting in decreasing FST and sunburn damage. A secondary aim was to evaluate possible negative impacts of netting on colour development. FSTs were continuously monitored on trees under netting and trees that were not netted, planted at a moderate density and grown on Open Tatura trellis. At harvest, sunburn incidence was determined, and blush intensity and coverage were measured.
Materials and methods
FSTs of a new red-blushed pear (P. communis) ‘ANP-0131’(marketed as Deliza®) were monitored at two neighbouring sites (‘netted’ and ‘non-netted’) at the Agriculture Victoria research farm, Tatura, Australia (36.44°S, 145.27°E; 115 m APSL). The soil type is a Red Sodosol (Isbell Citation2002), a duplex red-brown earth consisting of a 0.2 m deep sandy loam topsoil overlying a medium clay known locally as Shepparton fine sandy loam (Skene and Poutsma Citation1962). Top soil was mounded in the tree line to form a bed and allow surface drainage. The region has a temperate climate with warm to hot summers. The long-term mean maximum temperature in January is 29.3°C.
At each site, three 72 m rows of BP1 rootstock were planted in winter 2009 and budded with ‘ANP-0131’ scion in spring 2009 at 0.75 and 4.0 m tree and row spacing, respectively, and grown on Open Tatura trellis in north–south-oriented rows. Trees were drip irrigated to meet crop water requirements. Pest and disease protection methods were similar to commercial orchards with regular monitoring to determine spray requirements. The inter-rows were regularly slashed to maintain a grass sward and the tree rows were sprayed with herbicide as per commercial practice. The ‘netted’ site was within a 1 ha block enclosed by white HDPE monofilament knitted netting (12 mm Q Net, NetPro, Stanthorpe, Queensland, Australia). The netting extended to the ground on all sides of the block.
At each site, 13–15 fruit were selected for monitoring FST from each of four trees. Two of the trees at each site were grown on the eastern arm of the trellis and two were grown on the western arm. Monitored fruit were selected from the upper, mid and lower canopy, and were exposed to DSR for at least part of the day. FST was measured using fine-wire thermocouples (32 g copper-constantan) (Type T, Tranzflo NZ Ltd, Palmerston North, New Zealand). Thermocouples were installed between 23 December 2014 and 2 January 2015, and fruit were continuously monitored until 23 February 2015. A single thermocouple was inserted just under the peel on the sun-exposed side of each fruit. The thermocouples were positioned directly beneath the epidermis and forced 5–10 mm along the interface of the epidermis and the cortex of the fruit to place it away from the entry wound. FSTs were measured at 10 s intervals. One-minute averages of fruit temperature were then recorded using a datalogger (CR1000, Campbell Scientific, Logan, UT, USA). Fruit were inspected once or twice weekly to ensure sensors were in place. Sensors that became dislodged were re-inserted in the fruit and data collected while the sensors were dislodged were removed from the analysis. Peel and flesh of the fruit showed no visual signs of deterioration over the study period.
A visual grading scale was developed for assessment of sunburn browning and sunburn necrosis, as per McClymont et al. (Citation2016) where no sunburn = 0; bleaching of colour or very slight sunburn browning = 1; slight browning = 2; moderate browning = 3; severe browning = 4; slight sunburn necrosis = 5; moderate sunburn necrosis = 6; severe sunburn necrosis = 7. Monitored fruit were assessed for sunburn damage on 13 February 2015. The rest of the fruit on the eight trees were harvested on 12 February 2015 and assessed for sunburn damage. Differences in the occurrence of sunburn browning (scores 1–4) and sunburn necrosis (scores 5–7) between non-netted and netted sites were analysed using a two-sample two-sided binomial proportion test (Genstat 14.1, VSN International Limited, Oxford, UK).
A sample of 10 fruit were collected at harvest from each of 26 trees in the non-netted site (n = 257) and 32 trees in the netted site (n = 305) to determine fruit fresh weight (g), firmness (kg), soluble solids concentration (SSC, °Brix) and blush colour intensity (0–5 scale) and coverage (%). Fruit with severe pest damage were excluded from the analysis. Fruit firmness was measured on the blush cheek and opposing cheek with a penetrometer using an 8 mm probe. SSC was measured on expressed juice from two opposing cheeks using a digital hand-held refractometer (Model PR-1; Atago Co. Ltd, Tokyo, Japan). Colour intensity was assessed by scaling according to a colour chart developed specifically for the cultivar ‘ANP-0131’. Differences in fruit quality between non-netted and netted sites were analysed using a two-sample two-sided t-test (Genstat 14.1, VSN International Limited, Oxford, UK).
The microclimate was monitored at the netted and non-netted sites at 1 min intervals. Ten-minute averages were recorded using a datalogger (DataTaker DT80M, Thermo Fisher Scientific Inc., Yokohama, Japan). Air temperature (AT, °C) and relative humidity (%; HMP155, Vaisala Oyj, Vantaa, Finland) were measured at 1.5 m height with the sensor mounted in a cylindrical white aluminium screen. Accuracy of the HMP155 air temperature sensor was ± 0.2–0.3°C. Wind speed (m/s; PA2 Wittich and Visser, Rijswijk, Netherlands) was measured in the middle of the canopy at 2 m height. Diffuse and global radiation (W/m2; SPN1, Delta T Devices, Cambridge, UK) were monitored above the canopy at 3 m height. Accuracy of the SPN1 radiation sensor was ± 10 W/m2.
Direct solar radiation (DSR, W/m2) was calculated as DSR = global radiation − diffuse radiation. Daily total global radiation (MJ/m2) and daily mean, maximum and minimum AT, relative humidity and wind speed were calculated from 10 min data. Differences between weather variables at the non-netted and netted site were examined using 10 min and daily data collected over the period of fruit temperature monitoring. Differences in daily weather parameters between non-netted and netted sites were analysed using a one-sample two-sided t-test for the null hypothesis that the mean of the difference between each site (non-netted − netted) is equal to zero (Genstat 14.1, VSN International Limited, Oxford, UK). Relationships between the FST at non-netted and netted sites and weather variable were determined by linear regression in Genstat 14.1 (VSN International Limited, Oxford, UK).
Results
Effect of netting on weather variables
Daily maximum AT, mean wind speed and daily total global radiation were reduced by 1.3%, 75% and 15%, respectively, in the netted compared to the non-netted site (). Relative humidity tended to be higher under netting.
Table 1. Summary of daily weather variables measured at the non-netted and netted sites during the period of fruit surface temperature measurements (23 December 2014 to 23 February 2015).
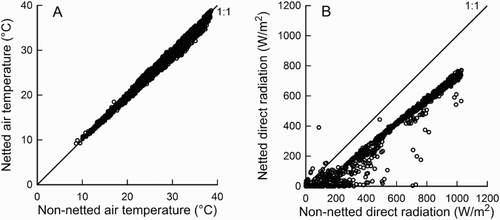
The relationship between 10 min observations of air temperature during the day (between 0600 and 2000 h) in the netted compared to the non-netted sites for the period of the study significantly differed from 1:1 (A). Differences in air temperature between the sites were minimal at low temperatures (9.2°C under netting compared to 9°C) and increased at higher temperatures (37.1°C under netting compared to 38°C). In contrast, 10 min observations of DSR in the netted site for clear sky days were substantially less than in the non-netted site (B), with mean DSR between 1200 and 1230 h of 695 W/m2 under netting and 972 W/m2 in the non-netted site. Hence, netting had little appreciable effect on air temperature but decreased daytime maximum DSR on clear sky days by 29%.
Figure 1. The relationship between netted and non-netted weather variables during the daytime (0600–2000 h) for the period 23 December 2014 to 23 February 2015: A, air temperature, ATNET = 0.96 (± 0.001) ATNONET + 0.57 (± 0.030); P < .001, R2 = 0.993, n = 5292, and B, direct solar radiation for clear sky days, DSRNET = 0.76 (± 0.005) DSRNONET − 48.4 (± 3.05); P < .001, R2 = 0.955, n = 1175. Data points are 10 min averages.
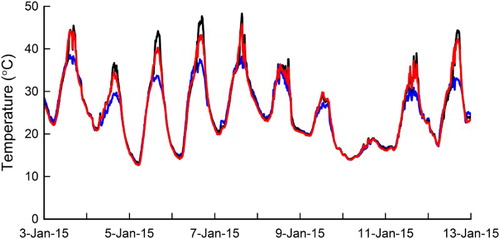
Fruit surface temperatures
Daytime FST was consistently higher than AT in both the non-netted and netted sites apart from during cloudy days (eg 10 January 2015) (). Night time FST was similar to AT. At the non-netted site, daytime FST was consistently higher than at the netted site. Differences between non-netted and netted FST ranged from 0.2°C on a cloudy day (eg total global radiation = 22.0 MJ/m2, maximum air temperature = 30.3°C on 20 January 2015) to 10.6°C on a sunny day (eg total global radiation = 33.4 MJ/m2, maximum air temperature = 30.1°C on 31 December 2015). One particular fruit at the non-netted site reached an FST of 50.3°C, whereas at the netted site, FST of all the monitored fruit did not exceed 46.4°C.
Figure 2. Fruit surface temperature of two monitored fruit within the non-netted (—) and netted sites (![]()
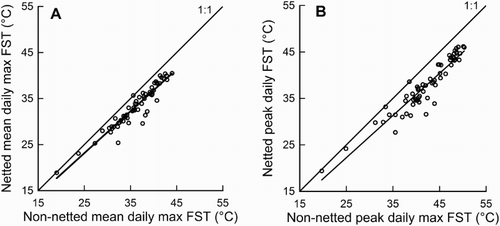
Daily maximum FST was significantly less in the netted site ().
Figure 3. The relationship between daily maximum fruit surface temperature (FST) in the non-netted and netted sites for the A, mean, MXFSTNET = 0.92 (± 0.005) MXFSTNONET; P < .001, R2 = 0.90, n = 63, and B, hottest, PKFSTNET = 0.90 (± 0.006) PKFSTNONET; P < .001, R2 = 0.89, n = 63, of all monitored fruit from 23 December 2014 to 23 February 2015.
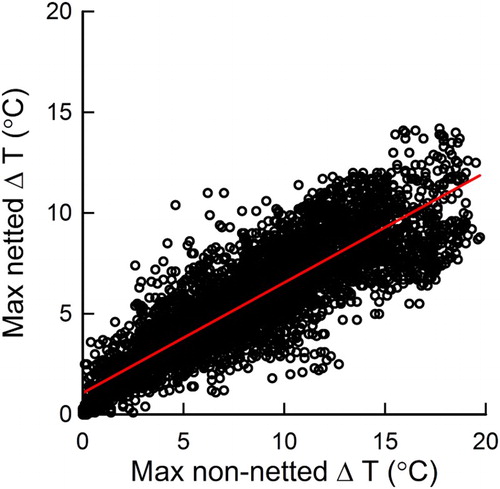
For all the monitored fruit over the measurement period, the greatest difference between FST and AT (ΔT) was 19.7°C and 14.2°C at the non-netted and netted sites, respectively (). The mean ΔT during the daytime was 7.6°C and 5.2°C at the non-netted and netted sites, respectively.
Figure 4. The relationship between fruit surface temperature minus air temperature (ΔT) at non-netted and netted sites for the hottest fruit between 0600 and 2000 h from 23 December 2014 to 23 February 2015, ΔTNET = 0.55 (± 0.004) ΔTNONET + 1.06 (± 0.036); P < .001, R2 = 0.78, n = 5249. Data points are 10 min averages.
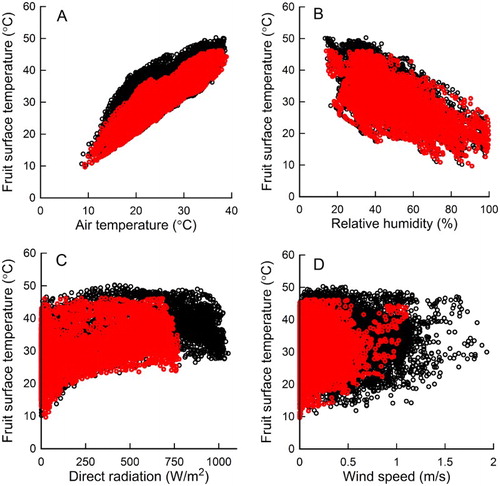
The relationships between FST and weather variables indicated that AT was the best single predictor of maximum FST between 0600 and 2000 h, followed by relative humidity and DSR (). Wind speed was a poor predictor of FST. The minimum ATs when FST exceeded 46°C were 30.2°C and 35.6°C for the non-netted and netted sites, respectively. The minimum AT when FST exceeded 47°C, the threshold for sunburn browning, was 30.8°C for the non-netted site. Despite air temperatures up to 38.9°C in the netted site, FST did not reach 47°C whereas FST in the non-netted sites reached 50.3°C, just exceeding the threshold for sunburn necrosis.
Figure 5. The relationship between fruit surface temperature with weather variables: A, air temperature, B, relative humidity, C, direct radiation, and D, wind speed, at non-netted (○) and netted (![]()
Sunburn damage
Netting substantially decreased the occurrence and severity of sunburn observed at harvest. shows the proportion of all fruit from four trees at each site (non-netted and netted) that were observed to suffer sunburn browning or sunburn necrosis at harvest and the mean sunburn score assigned to sunburnt fruit. Sunburnt fruit most commonly received a score of ‘1’ (bleaching of colour or very slight sunburn browning), 43% and 75% of sunburnt fruit from the non-netted and netted sites, respectively, fell in this category.
Table 2. Proportion of fruit affected by sunburn browning and sunburn necrosis and the mean sunburn score assigned to sunburnt fruit at harvest in non-netted (n = 340) and netted (n = 441) sites.
Fruit quality
Blush coverage and intensity were greater at the non-netted site (). At the non-netted site, 2% of fruit had no red coverage and 85% had 20–90% red coverage. By contrast, under netting, 21% of fruit had no red coverage and 61% had 20–80% red coverage. SSC and firmness also tended to be greater at the non-netted site. There was no difference in fruit fresh weight between sites.
Table 3. Fruit fresh weight, soluble solids concentration (SSC), firmness and colour intensity and coverage of fruit grown at non-netted and netted sites at harvest (12 February 2015).
Discussion
Netting lowered the risk of sunburn damage by decreasing radiation exposure and consequently decreasing FST. The netting used in this study reduced daily maximum DSR on clear sky days by 29%. Daily maximum FST was reduced by approximately 8% with the hottest fruit reduced by 10%.
Daytime FST was consistently greater than AT. Under netting, the difference between FST and AT did not exceed 14.2°C whereas without netting the maximum difference between FST and AT was 19.7°C. For example, on hot days when AT was 35°C, FST under netting ranged from approximately 36°C to 45°C with no risk of sun damage whereas without netting, FST ranged from 36°C to 50°C with a greater risk of sun damage. The decrease in FST of pears under netting observed in this study was similar to differences in FST between netted and non-netted apple trees reported by Iglesias and Alegre (Citation2006) where netted fruit were 2.4–4.1°C cooler than non-netted fruit when air temperature was 32–34°C, and Gindaba and Wand (Citation2005) where netted fruit were 5.4–9.7°C cooler than non-netted fruit when the daily maximum air temperature was 34–37°C. Similarly, McCaskill et al. (Citation2016) found a 4°C reduction in the maximum FST using grey netting that reduced direct radiation by 26%. Continuous monitoring of FST by thermocouples enabled AT to be compared simultaneously with temperature of ∼120 fruit, unlike other studies using digital thermometers or infrared sensors where sample sizes were limited. Additionally, continuous monitoring allows examination of diurnal patterns and enhances understanding of effects of fruit positioning and tree training on fruit exposure and, consequently, FST (McClymont et al. Citation2016).
Sunburn damage was reduced from 29% in the non-netted site to 10% in the netted site. Damage corresponding to a sunburn score of ‘1’ is unlikely to be recognised by consumers or cause fruit to be downgraded during packing. The high proportion of sunburnt fruit in this category means that just 2% of fruit under netting would be expected to be downgraded while approximately 16% of fruit from the non-netted site would be downgraded or culled. FST thresholds for sunburn browning and necrosis (47.1°C and 50°C, respectively; McClymont et al. Citation2016) were observed in the non-netted site where the maximum recorded FST was 50.3°C. In contrast, FST thresholds for sunburn were not reached in the netted site where the maximum FST was 46.2°C. Not all fruit were monitored so it is likely that FST on highly exposed fruit in the netted site exceeded FST damage thresholds as 10% of the fruit suffered sunburn browning although the severity of browning was lower than in the non-netted site. Temperature thresholds for sunburn damage are likely to vary between pear cultivars and the effectiveness of netting in reducing sunburn damage will relate to these thresholds. Thresholds for other pear cultivars are unknown; however, Schrader et al. (Citation2001) report temperature thresholds for apple cultivars ranging from 46°C to 49°C.
A minimum 20% red coverage is required to market ‘ANP-0131’ under the registered name ‘Deliza®’. Under netting, 39% of fruit did not meet this requirement, whereas at the non-netted site 15% did not meet this requirement. There are currently no market requirements for blush intensity. Visually, a blush of intensity score of ‘4’ is most desirable, and fruit from the non-netted site were more likely to receive this score than those from the netted site (24% occurrence compared to 14%, data not shown). Fruit fresh weight was not affected but SSC and firmness tended to be lower under netting. Negative impacts of netting on red colour and SSC of apples have also been reported for ‘Fuji’ (Dussi et al. Citation2005), ‘Cripps Pink’ and ‘Royal Gala’ (Gindaba and Wand Citation2005) and ‘Mondial Gala’ (Iglesias and Alegre Citation2006).
Middleton and McWaters (Citation2002) reported effects of netting on weather conditions similar to those observed in this study. They pointed out that the human perception of cooler air temperatures under netting was due to reduced radiant heat rather than differences in AT. The large reductions in wind speed under netting in this study (up to 90% compared to up to 50% reported by Middleton and McWaters Citation2002) were probably due to the netting extending to the ground, fully enclosing the netted study site. Such a reduction in wind speed reduces the capacity to remove heat from fruit (McCaskill et al. Citation2016). Minimising the effects of netting on wind is desirable in terms of reducing the risk from sunburn so it is recommended that orchards are not fully enclosed unless netting is used for pest exclusion.
AT was the best single predictor of maximum FST regardless of whether trees were netted or not. However, the reduction in FST under netting was attributed to the reduction in DSR. Fruit heats up above AT from DSR striking the fruit. Netting intercepts and scatters a portion of DSR thereby reducing ΔT. The effects of netting on DSR depend on the netting weave density and fibre diameter; the greater the weave density and/or fibre diameter, the lower the DSR and hence FST.
Netting is increasingly being used to protect high value, perennial crops as it provides multi-purpose protection (from hail, pests and sunburn). Lower colour of red-blushed pears is a drawback of netting. However, some studies have shown that filtering particular wavelengths of light increased anthocyanin content in red-blushed pears in comparison to natural sunlight (Dussi Citation1993; Dussi et al. Citation1995). Use of coloured netting that transmits higher proportions of particular wavelengths (Bastías et al. Citation2012) may be necessary to ensure sufficient colour development of red-blush pears. Greater knowledge regarding anthocyanin synthesis and degradation in new pear cultivars, such as described by Dussi (Citation1993), Steyn et al. (Citation2005), Huang et al. (Citation2009) and Qian et al. (Citation2013, Citation2014), and development of techniques to improve colour development in red-blushed pears are needed to ensure that netting remains a management option for pear growers.
Conclusion
Netting decreased pear FST and more than halved the occurrence and severity of sunburn damage. However, netting decreased red colour intensity and coverage, and tended to decrease SSC and fruit firmness. The impact of netting on colour development of pears requires further investigation particularly the wavelengths of light that determine red colour so that netting fibres could be designed to reduce DSR but not impact on the wavelengths of light that contribute anthocyanin synthesis. Alternatively, red colour is known to develop early in the season in pears and there is an opportunity to use retractable netting that is only drawn out after colour is determined. Other netting structures like a half gable in N-S rows that are designed to maximise fruit exposure to radiation in the morning and minimise afternoon exposure should be considered.
Acknowledgements
We would like to thank Dr Subhash Chandra and Dr Des Whitfield for statistical advice.
Disclosure statement
No potential conflict of interest was reported by the authors.
ORCID
Rebecca Darbyshire http://orcid.org/0000-0003-4712-8514
Additional information
Funding
References
- Bastías RM, Manfrini L, Grappadelli LC. 2012. Exploring the potential use of photo-selective nets for fruit growth regulation in apple. Chilean Journal of Agricultural Research. 72:224–231. doi: 10.4067/S0718-58392012000200010
- Colavita GM, Blackhall V, Valdez S. 2011. Effect of kaolin particle films on the temperature and solar injury of pear fruits. Acta Horticulturae. 909:609–615. doi: 10.17660/ActaHortic.2011.909.73
- Darbyshire R, McClymont L, Goodwin I. 2015. Sun damage risk of Royal Gala apple in fruit-growing districts in Australia. New Zealand Journal of Crop and Horticultural Science. 43(3):222–232. doi: 10.1080/01140671.2015.1034731
- Dussi MC. 1993. Fruit color development in red pears [MSc thesis]. Oregon State University.
- Dussi MC, Giardina G, Sosa D, González Junyent R, Zecca A, Reeb P. 2005. Shade nets effect on canopy light distribution and quality of fruit and spur leaf on apple cv. Fuji. Spanish Journal of Agricultural Research. 3:253–260. doi: 10.5424/sjar/2005032-144
- Dussi MC, Sugar D, Wrolstad RE. 1995. Characterizing and quantifying anthocyanins in red pears and the effect of light quality on fruit color. Journal of the American Society for Horticultural Science. 120:785–789.
- Evans RG, Kroeger MW, Mahan MO. 1995. Evaporative cooling of apples by overtree sprinkling. Applied Engineering in Agriculture. 11:93–99. doi: 10.13031/2013.25721
- Gindaba J, Wand SJE. 2005. Comparative effects of evaporative cooling, kaolin particle film, and shade net on sunburn and fruit quality in apples. HortScience. 40:592–596.
- Glenn DM, Prado E, Erez A, McFerson J, Puterka GJ. 2002. A reflective, processed-kaolin particle film affects fruit temperature, radiation reflection, and solar injury in apple. Journal of the American Society for Horticultural Science. 127:188–193.
- Huang C, Yu B, Teng Y, Su J, Shu Q, Cheng Z, Zeng L. 2009. Effects of fruit bagging on coloring and related physiology, and qualities of red Chinese sand pears during fruit maturation. Scientia Horticulturae. 121:149–158. doi: 10.1016/j.scienta.2009.01.031
- Human JP. 2013. Breeding blush pears (Pyrus communis L.) in South Africa. Acta Horticulturae. 976:383–388. doi: 10.17660/ActaHortic.2013.976.52
- Iglesias I, Alegre S. 2006. The effect of anti-hail nets on fruit protection, radiation, temperature, quality and profitability of ‘Mondial Gala’ apples. Journal of Applied Horticulture. 8:91–100.
- Iglesias I, Salvia J, Torguet L, Montserrat R. 2005. The evaporative cooling effects of overtree microsprinkler irrigation on ‘Mondial Gala’ apples. Scientia Horticulturae. 103:267–287. doi: 10.1016/j.scienta.2004.06.010
- Isbell RF. 2002. The Australian soil classification. Melbourne: CSIRO Publishing.
- McCaskill MR, McClymont L, Goodwin I, Green S, Partington DL. 2016. How hail netting reduces apple fruit surface temperature: a microclimate and modelling study. Agricultural and Forest Meteorology. 226–227:148–160. doi: 10.1016/j.agrformet.2016.05.017
- McClymont L, Goodwin I, Turpin S, Darbyshire R. 2016. Fruit surface temperature of red-blushed pear: threshold for sunburn damage. New Zealand Journal of Crop and Horticultural Science. 44(4):262–273. doi: 10.1080/01140671.2016.1216867
- Middleton S, McWaters A. 2002. Hail netting of apple orchards – Australian experience. The Compact Fruit Tree. 35(2):51–55.
- Qian M, Yu B, Li X, Sun Y, Zhang D, Teng Y. 2014. Isolation and expression analysis of anthocyanin biosynthesis genes from the red Chinese sand pear, Pyrus pyrifolia Nakai cv. Mantianhong, in response to methyl jasmonate treatment and UV-B/VIS conditions. Plant Molecular Biology Reporter. 32:428–437. doi: 10.1007/s11105-013-0652-6
- Qian M, Zhang D, Yue X, Wang S, Li X, Teng Y. 2013. Analysis of different pigmentation patterns in ‘Mantianhong’ (Pyrus pyrifolia Nakai) and ‘Cascade’ (Pyrus communis L.) under bagging treatment and postharvest UV-B/visible irradiation conditions. Scientia Horticulturae. 151:75–82. doi: 10.1016/j.scienta.2012.12.020
- Racsko J, Schrader LE. 2012. Sunburn of apple fruit: historical background, recent advances and future perspectives. Critical Reviews in Plant Sciences. 31:455–504. doi: 10.1080/07352689.2012.696453
- Schrader LE. 2011. Scientific basis of a unique formulation for reducing sunburn of fruits. HortScience. 46:6–11.
- Schrader LE, Zhang J, Duplaga W. 2001. Two types of sunburn in apple caused by high fruit surface (peel) temperature. Plant Health Prog. 10:1–5. doi:10.1094/PHP-2001-1004-01-RS.
- Skene JKM, Poutsma TJ. 1962. Soils and land use in part of the Goulburn Valley, Victoria comprising the Rodney, Tongala-Stanhope, North Shepparton and South Shepparton Irrigation Areas. Victoria, Australia: Department of Agriculture. Technical Bulletin No. 14.
- Steyn WJ, Holcroft DM, Wand SJE, Jacobs G. 2004. Anthocyanin degradation in detached pome fruit with reference to preharvest red color loss and pigmentation patterns of blushed and fully red pears. Journal of the American Society for Horticultural Science. 129:13–19.
- Steyn WJ, Wand SJE, Holcroft DM, Jacobs G. 2005. Red colour development and loss in pears. Acta Horticulturae. 671:70–85.
- Ubi BE. 2004. External stimulation of anthocyanin biosynthesis in apple fruit. Journal of Food, Agriculture and Environment. 2:65–70.
- van den Ende B. 2011 Sept 1. Grow perfect Forelle pears on Tatura. Good fruit Grower. http://www.goodfruit.com/grow-perfect-forelle-pears-on-tatura/.
- VSN International. 2011. GenStat for Windows 14th edition. VSN International, Hemel Hempstead, UK. Web page: GenStat.co.uk.
- Wand SJE, Steyn WJ, Holcroft DM, Mdluli MJ, van den Dool K, Jacobs G. 2005. Use of evaporative cooling to improve ‘Rosemarie’ and ‘Forelle’ pear fruit blush colour and quality. Acta Horticulturae. 671:103–111. doi: 10.17660/ActaHortic.2005.671.12
- Zoratti L, Karppinen K, Luengo Escobar A, Häggman H, Jaakola L. 2014. Light-controlled flavonoid biosynthesis in fruits. Frontiers in Plant Science. 5: 534. doi: 10.3389/fpls.2014.00534