Abstract
Studies were undertaken to determine the effects and relationships of compost type, aeration, and brewing time on compost tea properties, phytotoxicity, and efficacy in inhibiting growth of Pythium ultimum in vitro. The effect of compost tea on dry matter of tomato and sweet pepper seedlings cultivated on compost-amended substrate was also investigated. Compost type, aeration, and brewing time significantly affected the microbial and chemical properties of compost teas made from banana leaf (BLC) and lawn clipping (LCC) composts. Both aerated and nonaerated compost teas made from BL and LC composts significantly inhibited mycelial growth of P. ultimum. However, growth inhibition (GI) levels were significantly higher in compost teas produced from LCC. Aerating compost tea (ACT) did not consistently result in higher GI levels across compost types. Neither did increasing brewing time beyond 18 h for ACT and 56 h for nonaerated compost tea (NCT). Across all ACTs, yeast population was positively related to GI whereas bacterial population had a positive relationship with GI across all NCTs. Evidence suggests that multiple groups of microorganisms contributed to GI through antibiosis and competition for resources. All compost teas stimulated seed germination of tomato and root growth in sweet pepper. However, NCTs brewed for 56 h using LCC or BLC, and ACT produced from BLC brewed for 18 h, significantly reduced seed germination of sweet pepper. Application of NCT brewed for 168 h made from BLC significantly increased total dry matter of tomato seedlings by 122% compared with the control.
Introduction
There is a general lack of conclusive scientific evidence on the effect of compost type, aeration, and brewing time on the properties and efficacy of compost teas used as biocontrol agents and liquid fertilizers. Studies to date have focused primarily on the disease suppressive properties of compost tea, with little attention paid to the phytotoxicity and nutrient supply of compost teas (Hibar et al. Citation2006; Haggag and Saber Citation2007).
Moreover, few studies have been done to characterize the brewing process and to relate compost tea production methods to factors affecting the efficacy of compost tea as biocontrol agents and nutrient amendments. In fact, most of the evidence in support of the efficacy of compost tea as biocontrol agents and nutrient amendments has been in the form of anecdotal information provided by professional landscapers and gardeners and from commercial sensitive data held by private companies (Litterick and Wood Citation2009). Furthermore, most research on compost teas has been conducted under temperate environmental conditions using compost made from biomass resources such as grape marc and apple that differ from the agro-waste resources readily available for use in tropical regions (St. Martin and Brathwaite Citation2012). As such, there is a paucity of information on compost and compost teas made from the biomass of plants such as banana, an economic crop for many Asian-Pacific, African, Latin American, and Caribbean countries (Sharrock and Frison Citation1998), which produces a substantial amount of waste materials, including leaves, stems, and fruit during its production cycle. Other readily available local compostable materials, such as lawn clippings from large hotel golf courses and other institutions, yard waste from homes, and agro-waste from farms, are often discarded to the landfills, burnt or left piled along roadsides (Acurio et al. Citation1998).
With serious efforts being made by most developing and developed countries to reduce organic waste sent to landfills, composting and production of compost teas seem to be sustainable options that can be used by farmers, householders, and institutions to produce useful agricultural products. Compost teas may be particularly useful to the growing number of organic and ecological farmers who are faced with the challenge of the lack of approved disease management and plant fertility tools for use in commercial horticulture (Scheuerell and Mahaffee Citation2002; National Organic Standards Board 2004).
Aerated and nonaerated compost teas produced with and without nutrient additives have been shown to consistently suppress damping-off caused by Pythium spp. (Tränkner Citation1992; Scheuerell and Mahaffee Citation2004), a disease which has been reported to cause serious losses of young seedlings of several horticultural and vegetable crops cultivated in both greenhouse and field production systems (Howard et al. Citation1994). Although there has been some successful use, the understanding of, and research into, compost tea is limited (Scheuerell and Mahaffee Citation2002). As such, the efficacy of aerated and nonaerated compost teas with respect to compost source and age, optimum brewing time, benefits of aeration, addition of nutrients, phytotoxicity, cost, energy input, and mechanism of disease suppression continue to be investigated and debated (Ingham and Alms Citation2003; Litterick and Wood Citation2009). For example, Ingham and Alms (Citation2003) claim that aerated compost teas (ACTs) are more effective than nonaerated compost teas (NCTs) because they tend to have higher microbial populations and diversity. Conversely, the majority of scientific literature supports greater suppression of phytopathogens by NCTs (Ketterer and Schwager Citation1992; Tränkner Citation1992; Cronin et al. Citation1996). Interestingly, in the few studies done to directly compare the effect of ACTs and NCTs made from the same compost source, aeration has shown to have no effect on disease control or pathogen suppression (Al-Dahmani et al. Citation2003; Scheuerell and Mahaffee Citation2004; Scheuerell and Mahaffee Citation2006).
Despite contrasting assertions made by Ingham and Alms (Citation2003), correlations of total microbial or bacterial populations with increased disease suppression have been more frequently reported with NCTs (Ketterer et al. Citation1992; Ketterer and Schwager Citation1992) than with ACTs. There is little evidence to substantiate other assertions made, including that optimum fermentation time for peak suppressive properties of ACTs occurs between 18 and 36 h and NCTs are often phytotoxic (Ingham and Alms Citation2003).
For the most part, plant response to the addition of compost teas/extracts has varied from positive effects such as root growth stimulation (Hibar et al. Citation2006), increased yield, and fruit quality (Haggag and Saber Citation2007) to growth retardation, reduction in crop yield (Hargreaves et al. Citation2009), and phytotoxic effects (Tam and Tiquia Citation1994). The objective of this study was to evaluate the effects and relationships of compost type, aeration, and brewing time on compost tea properties, phytotoxicity, and efficacy in inhibiting the growth of Pythium ultimum in vitro. The effect of compost tea on the growth of tomato and sweet pepper seedlings cultivated on compost-amended substrate was also investigated.
Materials and methods
Compost production and characterization
A modified design to the rotary barrel composter developed by Minnich and Hunt (Citation1979) was used to compost and cure banana leaf, including petioles, and lawn clippings. Modifications consisted of the use of a blue 0.25 m3 closed top, food grade polyethylene barrel (0.9 × 0.55 m × 3 mm) rather than a metal drum and the inclusion of a mixing apparatus. The barrel was supported on a 1 m steel X–frame stand with a 19 mm thick steel rod, which ran longitudinally through the centre of the barrel and the mixing apparatus, constructed from 0.22 m inverted L-shaped steel rods welded perpendicularly, at 15 cm apart, onto the longitudinal steel rod inside of the barrel. Holes (16 mm diam.) were made 15 cm within and between rows through the walls of the barrel for aeration and to allow drainage of excess water. All steel rods inside the composter were painted with anticorrosive paints, Epilux 5 base and converter, in accordance with the product specification instruction document.
Lawn clippings and banana leaf were collected from two unweeded lawns and a banana field, respectively, located on a 2 ha estate in Talparo, Trinidad, 10° 30′ 0″ N, 61° 16′ 0″ W, where synthetic pesticides had not been applied for over 10 years. Dry, brown leaves with little or no photosynthetic capacity or latex were removed from banana plants, chopped using a machete, placed on a plastic covering, mixed and subdivided in the field at the time of sampling. Sawdust (particle size 0.03 − 0.6 mm) made from local teak (Tectona grandis) was purchased from a local sawmill. Cow manure stock piled for more than 3 months was collected from the University Field Station, Valsayn, Trinidad, where a semi-intensive forage-based system is used to rear cattle for milk production.
Composite samples of the respective feedstock materials were carried to the laboratory and prepared in accordance with the TMECC method 02.02-F (Thompson Citation2002). Moisture content (105°C for 24 h), total organic carbon (TOC) content by loss on ignition, 550°C for 5 h (Nelson and Sommers Citation1996), total Kjeldahl nitrogen (TKN) (Bremner Citation1996) and bulk density (Blake and Hartge Citation1986) of each feedstock material were determined to develop compost formulations with ideal heap characteristics as described by Thompson (Citation2002). Compost formulations on an air-dried weight basis were banana leaf compost (BLC): 12 kg banana leaf+18 kg sawdust+18 kg cow manure+1 kg gypsum+1 l Nitro-Plus 9TM and lawn clipping compost (LCC): 20 kg lawn clipping (80% Axonopus compressus)+14 kg sawdust+11 kg cow manure+1 kg gypsum+1 l Nitro-Plus 9TM.
Organic waste materials were collected and composted in June 2009. All feedstock materials were air dried for at least three days prior to mixing and shredded to a particle size of < 2 cm width and ≤ 9 cm length. Nitro-Plus 9TM was added to the banana leaf and lawn clippings to achieve the desired C/N ratio (30:1). Compost formulations were mixed for 25 min in a portable cement mixer before being placed in the composters, which were housed in a gable roof, open-sided protective structure (50 × 20 × 6 m) located at The University of the West Indies, St. Augustine. Formulations were loaded into the barrels up to 75% of total capacity, and passively dechlorinated water was added to the compost to adjust moisture level within the desirable range of 50–60%.
For the first two weeks, the composters were manually turned daily for 1 min each in a clockwise then an anticlockwise direction. Thereafter, the barrels were rotated every third day for the same duration. Compost maturity index was determined using the Solvita® CO2 respiration (Group A) and NH3 volatility tests (Group B) on composting day 20 when there was little or no difference between ambient and compost temperature and C/N ratio < 25:1 (California Compost Quality Council Citation2001).
Compost was allowed to cure for 70 more days before it was used to make compost teas. At the end of the curing period, compost maturity index was determined, and homogenized composite samples of compost (4 l per barrel) were analysed for moisture content, TOC, and TKN, using methods previously described. Electrical conductivity (EC) was determined using the TMECC method 04.10-B (Thompson Citation2002); pH using the TMECC method 04.11-A, 1:5 slurry, water extract (Thompson Citation2002); phosphorus by the method of Kuo (Citation1996); and potassium, according to Helmke and Sparks (Citation1996). Methods described by Scheuerell and Mahaffee (Citation2004) were used to enumerate total culturable bacteria, fungi, and yeasts.
Compost tea production
Ninety-day-old banana leaf composts were used to make compost teas. Nineteen litre aerated compost tea brewers were used to make aerated compost teas: 17 l of tap water (25°C ± 3°C) was placed in a 19 l bucket and aerated for 30 min with an air pump without the bucket lid to reduce chlorine present in the water, and a 1000 g compost sample (approximately 50% moisture w/w) was placed in a 100 μm mesh filter bag and immersed in the dechlorinated water. The bucket was closed with the lid and the mixture was brewed over a 36 h period with a constant supply of air from an air pump. The same procedure was used to make aerated lawn clipping compost teas.
Nonaerated BLC and LCC teas were made using the same procedure, but air was not actively pumped into the mixture and brewing was done over a 168 h period. Compost teas were produced in triplicate for each compost type for each brewing method used. No nutrient amendments were added to compost teas before, during, or after the brewing process.
Microbial and chemical characterization of compost teas
Compost tea samples (250 ml) were collected at 18, 27, and 36 h for ACTs, and 56, 112, and 168 h for NCTs. Samples were immediately processed after collection. Electrical conductivity, pH, phosphorus, potassium, total culturable bacteria, fungi, and yeasts were determined using methods described for the analysis of compost.
Ammonia, nitrate, and nitrite were estimated in accordance with Bremner (Citation1996), calcium and magnesium using methods described by Suarez (Citation1996), and copper and zinc in accordance with Reed and Martens (Citation1996). Dissolved oxygen (DO) was measured at predefined time intervals with respect to the brewing method (aerated or nonaerated) used.
Effect of compost teas on the growth of P. ultimum
Suppressive effect of aerated and nonaerated banana leaf and lawn clipping compost teas was investigated using a central disk antagonistic assay. Compost tea treatments included aerated banana leaf or lawn clipping compost teas brewed for 18, 27, and 36 h, and nonaerated banana leaf or lawn clipping compost teas brewed for 56, 112, and 168 h. Two aliquots of 0.1 ml of compost teas were placed in two 5 mm pits 1 cm away from the edge of the Petri dish and equidistant from a central disk of P. ultimum. Autoclaved (123°C, 20 min) composts teas were used to determine whether mycelial inhibition of P. ultimum was related to microbial populations or activity.
P. ultimum cultured on potato dextrose agar without the addition of water or any of the compost teas was used as the control. All Petri dishes were incubated at 25°C for five days. Inhibition was discerned by limited growth or the complete absence of mycelium in the inhibition zone between the pathogen and the colony of the antagonist microbes. Percentage mycelial growth inhibition (GI) was recorded on day 13, when the P. ultimum in control Petri dishes reached its maximum growth. Percentage mycelial GI was calculated in accordance with Skidmore and Dickinson (Citation1976): (Kr − r1) × 100/Kr, where Kr represents the radius of the control pathogens growth, and r1 the radius of the pathogen's growth toward the antagonist. A complete randomized design was used with the experimental unit (Petri dish) being replicated five times and the experiment replicated twice.
Phytotoxic evaluation of compost teas
In vitro germination and root elongation bioassay with tomato cv. Calypso and sweet pepper cv. California Wonder TMECC method 05.05-B (Thompson Citation2002) was used to assess the phytotoxicity of compost teas. Germination index, relative germination, and root elongation rates were used to infer the presence of phytotoxic compounds in compost teas.
Compost teas as liquid fertilizers
The compost tea with the highest nutrient content, root or seed germination stimulatory effects, and an absence of phytotoxic compounds, was used in an in vivo seedling growth study. Three-week-old tomato cv. Calypso and sweet pepper cv. California Wonder seedlings grown in PRO-MIX (‘BX’/Mycorise®) (PM) were transplanted into Styrofoam containers (12.7 × 10.2 × 8.9 cm) filled with PM blended with banana leaf or lawn clipping at ratios of 1:1 and 1:4 v/v, respectively. Applications of Miracle-Gro® (Water Soluble All Purpose Plant Food) (MG) and compost tea were made at transplanting as well as 7 and 14 days after transplanting. Fifteen millimetres of MG or compost tea was applied to each container with three seedlings. Miracle-Gro® applied was mixed at the recommended rate of 4 ml l− 1. Substrates to which only potable water was applied were used as controls.
Studies were done using a complete randomized design with each Styrofoam container as an experimental unit. Experimental units were replicated five times for the respective crop and the experiment was replicated twice. All studies were done under a conventional span-roof, naturally ventilated glasshouse (8.5 × 3.7 × 2.7 m) located at the Department of Food Production, The University of the West Indies, St. Augustine. Mean day and night temperatures were 31°C and 24°C, respectively, at a corresponding relative humidity of 61 and 95%. Total dry matter per seedling was determined 21 days after transplanting.
Statistical analysis
A univariate analysis of variance method of SPSS (ver. 17.0, 2008, SPSS Inc., Armonk, NY) for Windows was used to determine the effect of treatments on response variables, and treatment means were compared using Tukey's test (p ≤ 0.05). Simple effects analyses were used to investigate significant interactions among independent variables using the EMMEANS subcommand (Green and Salkind Citation2004). Data from the in vitro phytotoxic and GI studies expressed as percentages were submitted to arcsine transformation before analysis. Multiple and simple linear regression were used to determine the most important microbial population affecting mycelial GI of the pathogen or the most important chemical factors retarding root elongation, seed germination rate, and germination index. Homogeneity of error variances of replicated experiments were tested using the F-ratio test or Bartlett's test (Gomez and Gomez Citation1984). No significant difference occurred among the repeated experiments; therefore, data from trials were pooled for analysis.
Results and discussion
Compost maturity, chemical and microbial properties
Both BLC and LCC recorded 7 for the Solvita CO2 evolution test and 5 for the NH3 + test, which indicates that they were cured and ammonia content was very low ( < 100 ppm), below the level toxic to seedlings (Woods End Research Laboratories Citation2002). Total nitrogen content was significantly higher in LCC than in BLC, with no significant differences in sodium and nitrate+nitrite concentrations between compost types (Table ). Phosphorus and potassium concentrations as well pH and EC were significantly higher in BLC than in LCC (Table ).
Table 1 Selected chemical characteristics of compost and aerated and nonaerated banana leaf and lawn clipping compost teas.
The predominant microorganisms in banana leaf and lawn clipping composts were bacteria with respective populations of 4.5 × 108 and 1.3 × 108 CFU g dw− 1 (Table ). Fungal populations were less than the square root of the corresponding bacterial populations, with values of 5.2 × 103 CFU g dw− 1 in BLC and 4.6 × 103 CFU g dw− 1 in LCC (Table ).
Table 2 Microbial characteristics of compost, and autoclaved ( + Aut) and nonautoclaved ( − Aut), aerated and nonaerated, banana leaf and lawn clipping compost teas.
Microbial properties of compost teas
The main effects of compost type, aeration, and brewing time on microbial and chemical parameters of compost teas were significant (p < 0.01). DO concentration in the aerated compost teas was>5 mg l− 1 during the brewing process (Table ). According to Davis (Citation1975), this is the minimum aerobic level needed to support a diverse population of aquatic organisms. Contrastingly, DO concentration of nonaerated compost teas was < 4.1 mg l− 1 and generally decreased as brewing time progressed. Although there is no standard DO concentration level used to distinguish ACT from NCT in the literature (Johnson Citation1999), it is evident that the level in ACT was more suitable for the proliferation of aerobic microorganisms.
Data showed that DO concentrations fluctuated during the ACT brewing process, with peak values occurring at 27 h, which corresponded with the lowest bacterial and yeast populations (Table ). The subsequent increase of bacterial and yeast populations at ACT 36 h (Table ) and decrease in the NH4 + − N and oxygen concentration in ACTs made from LCC from 27 to 36 h (Table ) suggest that NH4 + − N and oxygen were used by microbes for the synthesis of ribonucleic acid, enzymes, and other molecules, as well as for maturation and multiplication.
A similar pattern was observed with bacterial populations of NCT at 112 h. However, there was no decrease in NH4 + − N content between NCT-112 h and NCT-168 h (Table ). According to McCarty (Citation1964), the rate and quantity of nitrogen consumption by anaerobic microorganisms is generally slower and lower than that of aerobic microorganisms. This suggests that the rate of NH4 + − N consumption by microorganisms or loss in the NCTs was slower than the rate at which NH4 + − N dissolved into compost teas from compost.
Data showed that total microbial populations were generally higher in ACTs than in NCTs (Table ). According to McCarty (Citation1964), under more anaerobic conditions, less energy is available to microorganisms, therefore their growth rate is slow, and only a small portion of energy produced from the degradation of waste is used to produce new cells. Bacterial and yeast populations were generally higher in ACTs, with fungi being higher in NCTs. Peak yeast populations were recorded at 112 h for both banana leaf and lawn clipping NCTs. The population of fungi also fluctuated in ACT and NCT, with the highest populations at ACT 27 h for both banana leaf and lawn clipping and at 168 h for NCTs (Table ). Jack and Thies (Citation2006) reported similar DO concentration and bacterial population patterns when thermogenic composts were used to make compost teas over a 36 h period without the addition of molasses. They also reported that compost type was a significant factor affecting microbial and chemical characteristics of compost teas.
Chemical properties of compost teas
Despite the increase dissolution of nutrients expected in ACT due to the agitation caused by aeration, the NH4 + − N and NO3 − N+NO2 − − N content was generally higher in NCTs than in ACTs (Table ). Several authors attributed the higher nitrogen content in end products of anaerobic process to lower nitrogen loss through nitrification-denitrification reactions and ammonia volatilization compared with aerobic processes (Shih Citation1987; Wilkie Citation2005; Topper et al. Citation2006). Increases in the surface area between the liquid, which results from increase aeration, means more oxygen is available for microbial reactions to convert ammonium-N to nitrate or nitrite, which can subsequently be converted to and released as N2O or N2. Zhao and Chen (Citation2003) found that at a temperature of 20°C, ammonia volatilization increased with increasing rates of aeration. This may explain the differences in NH4 + − N and NO3 − N+NO2 − − N content between ACTs and NCTs, which were brewed at similar temperatures. The effect of aeration on P and K concentration was not as discernible as it was on NH4 + − N and NO3 − N+NO2 − − N content. This may be due to the lack of volatile-loss pathways for P or K, which resulted in these nutrients being highly conserved.
With the exception of banana aerated compost teas, the concentrations of Ca generally increased with brewing time (Table ). Calcium concentration across compost teas varied between 151.0 and 553.2 mg l− 1 and was generally higher in ACTs and teas made from lawn clipping compost. As with Ca, concentrations of Mg were also higher in ACTs with the highest concentration, 240.1 mg l− 1, recorded in compost teas made from banana leaf compost. Contrastingly, sodium concentrations were considerably lower than that of Mg and fluctuated across brewing time within the respective brewing method. Sodium concentrations across compost teas varied between 2.95 and 28.55 mg l− 1 and were below the 0–50 mg l− 1 target range recommended for the water to be applied to most vegetable crops grown in containers (Whipker Citation1999).
Copper concentration for the respective brewing method and compost type (banana leaf or lawn clipping) generally remained constant throughout the brewing process (Table ). Conversely, Zn concentrations fluctuated across brewing time within the respective brewing method. Concentrations of Cu and Zn were below the respective target ranges of < 0.2 and < .50 mg l− 1, respectively, recommended for water to be used to irrigate vegetable crops grown in containers (Whipker Citation1999).
Solubility of nutrients in the composts as well as the difference rates of consumption and liberation of nutrients by microorganisms present in the compost teas appear to be the main factors affecting the concentration of nutrients across brewing time. Increases in the concentration of nutrients across brewing time may also be the result of a concentration effect (Bernal et al. Citation1996), which occurs when water is lost during the brewing process through evaporation. This may be particularly evident in the production of aerated compost teas, where water molecules are continuously agitated by air supplied by an air pump and evaporation was greater.
Electrical conductivity values for all compost teas were above the standard recommended higher limit (Ayers and Westcot Citation1985), which has been reported to cause salinization of growth substrates and negatively affect crop growth and lower yields (Katembe et al. Citation1998; Munns and Tester Citation2008). There was no consistent pattern in the variation of EC over brewing time within compost type. However, data showed that significant reductions in NH4 + − N usually corresponded to an increase in EC values. This suggests that EC was affected by the release of soluble salts such as ammonium (Huang et al. Citation2004). As with pH, EC was generally higher in compost teas made from BLC.
The pH values of ACTs made from LCC were < 5.3, the standard recommended lower limit for irrigation water to be used in vegetable container production systems (Bailey and Bilderback Citation1997). Continuous application of ACTs with such low pH values as liquid fertilizers may acidify substrates resulting in calcium and/or magnesium deficiency or manganese toxicity in vegetable crops (Russell Citation1988). Table shows that pH generally remained constant across brewing time within aeration method.
Effect of compost type, aeration, brewing time, and autoclaving on the efficacy of compost tea to inhibit the mycelial growth of P. ultimum
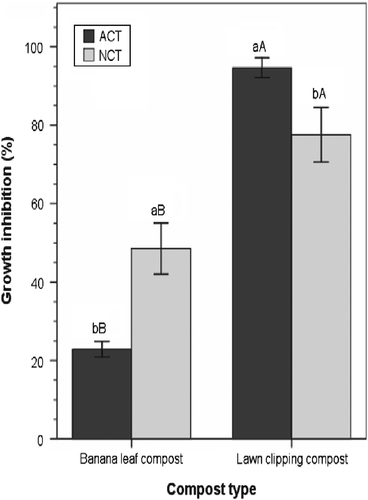
Compost type had a significant main effect on the efficacy of compost teas with higher GI levels achieved with teas produced from LCC (86%) than from BLC (36%). Although the main effect of aeration was not significant, there was a significant compost type × aeration interaction effect (p < 0.05). Figure shows that aerating compost tea did not consistently result in higher GI levels. Despite having lower microbial populations, NCTs made from BLC were more efficacious at inhibiting the mycelial growth of the pathogen than ACTs made from the same compost source (Figure ). These results contradict the claims of Ingham and Alms (Citation2003) that ACTs are generally more effective than NCTs because they tend to have higher microbial populations and diversity. Moreover, the results seem to imply that the presence and activity of specific groups of microorganisms within each compost type rather than total microbial population may be contributing to the suppressive effect. Therefore, an examination of the population metrics of these specific microorganisms rather than total microbial populations may prove to be more reliable in rationalizing the efficacy between aerated and nonaerated compost teas.
Figure 1 Effect of compost type and aeration on the efficacy of compost tea in inhibiting the mycelial growth of P. ultimum. Means with different lowercase letters are significantly different within compost type; those with different uppercase letters are significantly different across compost type within the same aeration method (aerated or nonaerated) at p ≤ 0.05, according to Tukey's test. Error bars indicate one standard error.
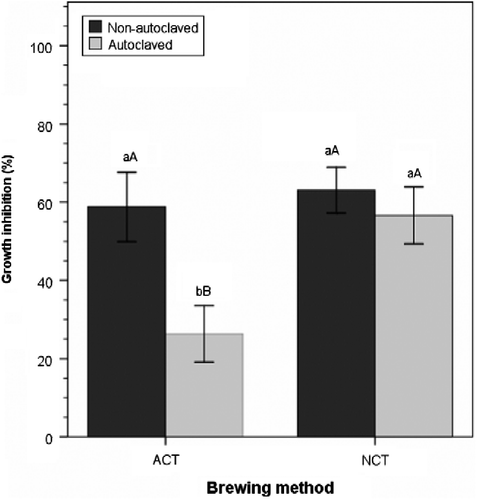
The presence and prevalence of microorganisms were affected by the autoclaving process, which had a significant main effect on ACTs within each compost type (p < 0.01) (Table ). Across compost type, higher levels of inhibition were achieved with nonautoclaved ACTs rather than with autoclaved ACTs (Figure ). Unlike Tränkner and Liesenfeld (Citation1990), autoclaving reduced microbial populations but did not sterilize compost teas. Most of the microorganisms that survived the autoclaving process were from Bacillus, Actinomycetes, and Trichoderma or yeasts species. These heat-tolerant microorganisms were the predominant species both in nonautoclaved and autoclaved compost tea samples and were observed to be present when the mycelial growth of P. ultimum was inhibited. Contrary to the findings of Tränkner and Liesenfeld (Citation1990), heat-treating or autoclaving did not negate all inhibition of pathogen growth in vitro. In fact, autoclaving had no significant effect on the suppressivity of NCT within each compost type (p>0.05) (Table ). Cronin et al. (Citation1996) reported similar results, where autoclaved extracts from spent mushroom substrate compost were less effective than untreated extracts against Venturia inaequalis, but they retained most of their efficacy. According to Cronin et al. (Citation1996), the results seem to indicate the likely role of heat-stable chemical compounds produced by specific microorganisms in the compost teas in suppressing mycelial growth of the pathogen.
Table 3 Probability of significance for sources of variation for mycelial growth inhibition of P. ultimum.
Figure 2 Effect of aeration and autoclaving on mycelial growth inhibition of P. ultimum. Means with different lowercase letters are significantly different at the same aeration method; those with different uppercase letters are significantly different across aeration methods within the same autoclave treatment (autoclaved or nonautoclaved) at p ≤ 0.05, according to Tukey's test. Error bars indicate one standard error.
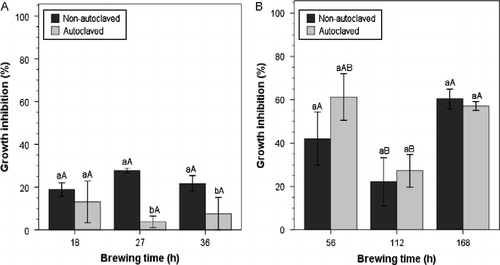
The general trend described by Ketterer (Citation1990) of the suppressive properties of ACTs and NCTs increasing with brewing time to a maximum and then declining was not evident in this research work. There was no significant increase or decrease in the efficacy of ACTs within compost type as brewing time progressed (p>0.05) (Table ). In contrast, the main effect of brewing time within compost type for NCTs was significant (p < 0.01) (Table ), with GI levels for nonautoclaved NCTs declining with brewing time to the lowest level at 112 h, followed by a subsequent increase at 168 h (Figures and ). The results contradict the assertions by Ingham and Alms (Citation2003) that optimum fermentation time for peak suppressive properties, that is ACTs with greater microbial populations and diversity, is usually between 18 and 36 h. In fact, total microbial populations of ACTs across compost types were generally lowest at the brewing time of 27 h (Table ) and the resulting GI levels were not significantly different from 18 or 36 h treatments (Figure and ). Contrary to assertions made by other investigators (Cantisano Citation1998; Ingham and Alms Citation2003), shorter brewing cycles of 18 h for ACT and 56 h for NCTs can be used to produce compost teas of a relatively high level of GI efficacy of P. ultimum.
Figure 3 Effect of (A) aerated and (B) nonaerated lawn clipping compost teas on mycelial growth inhibition of P. ultimum. Means with different lowercase letters are significantly different at the same brewing time; those with different uppercase letters are significantly different across the brewing time within the same autoclave treatment (autoclaved or nonautoclaved) at p ≤ 0.05, according to Tukey's test. Error bars indicate one standard error.
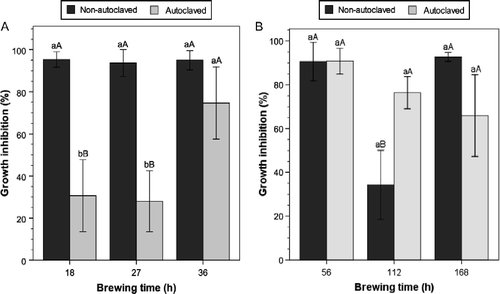
Figure 4 Effect of (A) aerated and (B) nonaerated banana leaf compost teas on mycelial growth inhibition of P. ultimum. Means with different lowercase letters are significantly different at the same brewing time; those with different uppercase letters are significantly different across the brewing time within the same autoclave treatment (autoclaved or nonautoclaved) at p ≤ 0.05, according to Tukey's test. Error bars indicate one standard error.
Analyses showed brewing time × autoclaving interaction effect in nonaerated lawn clipping compost teas was significant (p < 0.05) (Table ). Figure shows that with the exception of NCT-112 h, mycelial GI between autoclaved and nonautoclaved treatments across brewing time was not significantly different. Optimal brewing time for the highest inhibitive properties of compost tea seems to be mainly dependent on the properties of the compost as influenced by feedstock and the compost tea production method used (Litterick and Wood Citation2009).
Relationship between microbiological properties of compost teas and mycelial growth inhibition of P. ultimum
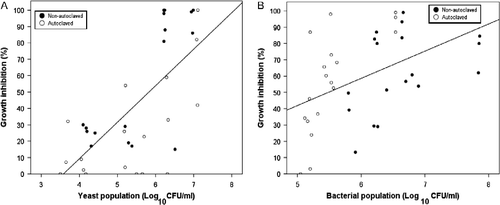
Multiple linear regression analyses of pooled data for all ACTs, including nonautoclaved and autoclaved treatments, showed there was a significant positive relationship between mycelial GI and yeast and fungal populations (R2 = 0.57, p < 0.001). Together, yeast (β = 0.61, p < 0.01) and fungal populations (β = 0.37, p < 0.01) explained 54% of the variation of mycelial GI. Though mycelial GI occurred over a range of yeast populations, 103.5 to 107.5 CFU ml− 1; data seem to suggest that 106 CFU ml− 1 is the minimum threshold to achieve GI levels of 50% or greater (Figure .). Positive significant relationships between yeast population and mycelial GI was also observed within pooled datasets of only nonautoclaved ACTs (R2 = 0.56, p < 0.001) or autoclaved ACTs (R2 = 0.38, p < 0.01), where yeast populations explained 53 and 38% of the variation, respectively. These results seem to indicate that across compost types, yeasts had the greatest influence on the inhibitive effect of the ACTs.
Figure 5 Relationship of (A) yeast population of all aerated compost teas (ACTs) and (B) bacterial population all nonaerated compost teas (NCTs) on mycelial growth inhibition of P. ultimum.
An examination of the linear regression for ACT made from lawn clipping compost indicated that there was a significant positive relationship between mycelial GI and total microbial population (R2 = 0.63, p < 0.001), with total microbial population (β = 0.79, p < 0.001) explaining 60% of the variation (Figure ). Though considerable mycelial GI was achieved with autoclaved ACTs, which did not contain fungi, data showed that the presence of fungi and yeasts in nonautoclaved ACTs consistently resulted in higher GI levels (Figure ). Total microbial populations of ≥ 1014 CFU ml− 1 tended consistently to result in mycelial GI of ≥ 80% (Figure ). Interestingly, there was a significant positive relationship between fungal population and mycelial GI (R2 = 0.43, p < 0.01) in ACT made from banana leaf compost with fungal population (β = 0.66, p < 0.01), explaining 40% of the variation (Figure ).
Figure 6 Relationship of (A) total microbial population of aerated compost tea (ACTs) made from lawn clipping compost (LCC) and (B) fungal population of ACT made of banana leaf (BLC) on mycelial growth inhibition of P. ultimum. Total microbial population is calculated as the sum of the populations of bacteria, fungi, and yeast (each log10 CFU/ml of compost tea).
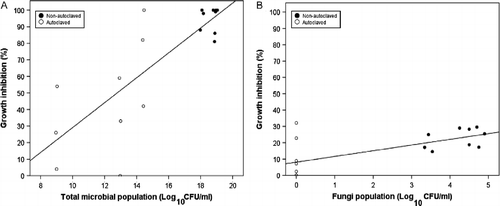
The results for ACTs seem to indicate that multiple microbial groups contributed to the mycelial GI of the pathogen. This may suggest that main mechanisms for inhibition were antibiosis and competition for nutrients. The positive relationship between total microbial population and efficacy of ACTs, as posited by Ingham and Alms (Citation2003), was only evident in ACTs made from LCC. The efficacy of ACT was compost type specific, with populations of particular groups of microorganisms, mainly fungi and yeasts, being more predictive of mycelial GI. Though not as well studied as bacteria as biocontrol agents, yeasts and fungi have been reported to suppress phytopathogens and plant diseases (Punja Citation1997; El-Tarabily Citation2004; El-Mehalawy et al. Citation2007) using similar mechanisms as described by Hoitink and Fahy (Citation1986).
As with Scheuerell and Mahaffee (Citation2004), no significant relationship was found between suppressive efficacy of ACTs and bacterial populations when ACTs were pooled. The results, therefore, contradict the claim by Ingham and Alms (Citation2003), that increasing the population of total bacteria generally increases the level of efficacy of ACTs against diseases and phytopathogens. The lack of a significant relationship between GI and the bacterial population may be explained by the absence of population metrics of unculturable bacteria types, which may have a role in the GI of the pathogen. Scheuerell and Mahaffee (Citation2002) posited that the variability in the populations of specific species of bacteria with the ability to consistently suppress a pathogen may also account for the lack of a significant relationship between bacterial population and suppressivity of compost teas.
Multiple linear regression analysis for pooled datasets of all NCTs showed that both bacterial and fungal populations had a significant relationship with mycelial GI of the pathogen (R2 = 0.35, p < 0.01). Bacterial population was positively related with mycelial GI and together with fungal populations explained 32% of the variation. Contrastingly, the fungal population (β = − 0.63, p < 0.01) was inversely related to mycelial GI, which suggests that its presence and activity neutralized or suppressed the inhibitive effect of indigenous bacteria. Figure shows a weak but positive relationship of the bacterial population of NCTs and mycelial GI of the pathogen.
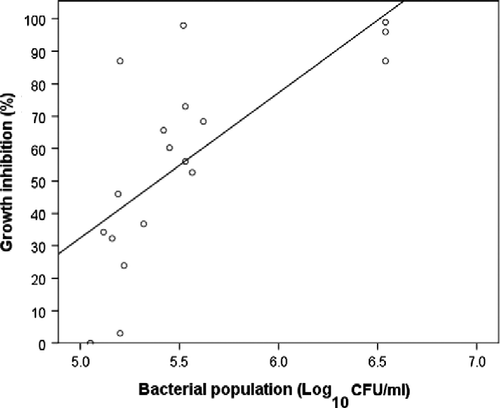
When data for autoclaved NCTs were removed from the pooled datasets, bacterial, fungal, yeast, or total microbial populations showed no significant relationships with mycelial GI (p>0.05). Analyses of the pooled datasets of all autoclaved NCTs showed there was a significant relationship between bacterial population and mycelial GI (R2 = 0.52, p < 0.01) (Figure ). Bacterial populations (β = 0.72, p < 0.01) explained 49% of the variation in mycelial GI.
Figure 7 Relationship of bacterial population of autoclaved nonaerated compost teas (NCTs) on mycelial growth inhibition of P. ultimum.
Multiple linear regression of NCTs made from lawn clipping or banana leaf compost showed that there was no significant relationship between total microbial population and mycelial GI. Neither was there a significant relationship between mycelial GI and bacterial, fungal, or yeast populations. The general lack of significant relationships between mycelial GI and total microbial population or specific subpopulations of NCTs seems to suggest that metrics of specific chemical compounds may better explain the variation of mycelial GI.
Phytotoxicity of compost teas
There were no significant differences in main and interaction effects of compost type (lawn clipping or banana leaf compost), aeration (aerated or nonaerated), and brewing time on the germination index, relative root length, and germination rate of tomato seedlings. The overall mean germination index, relative root length, and germination rate of tomato were 80, 89, and 138%, respectively, which indicates that compost teas were not phytotoxic (Zucconi et al. Citation1985). On the contrary, it was observed that compost teas stimulated the seed germination of tomato (data not shown). Similar results have been reported with green bean (Tam and Tiquia Citation1994) and tomato (Dukare et al. Citation2011) used in phytotoxic and growth bioassays. Several authors have attributed the seed germination and root stimulatory effect to a small fraction of lower molecular mass ( < 3500 Da) components of humic substances present in compost (Nardi et al. Citation2002; Chen et al. Citation2004).
The regression model for root length was significant and showed that Mg was the only predictor that negatively affected root length (β = − 0.48, p < 0.10). The other two predictors of the model, Zn (β = 0.56, p < 0.05) and EC (β = 0.72, p < 0.05), were positive and, together with Mg, explained 23% of the variance (R2 = 0.32, p < 0.05). No negative predictors were identified in the regression model for seed germination rate. Electrical conductivity, the only predictor in the model that was positively related to seed germination rate (β = 0.4, p < 0.05), explained 13% of the variance in seed germination (R2 = 0.16, p < 0.05). This suggests that the threshold tolerance level of electrical conductivity as it relates to imbibition was not exceeded.
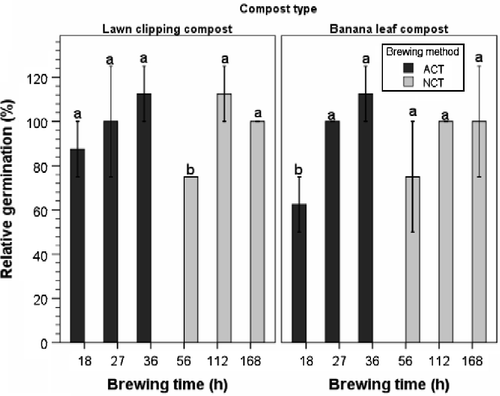
The main and interaction effects of compost type and aeration on the germination index, relative root length, and germination percentage of sweet pepper seedlings were not significant. Relative root length across treatments was 114.1%, which indicates that compost teas had a stimulatory effect on root development of sweet pepper seedlings. Although the germination index was high (110%), results showed that banana leaf and lawn clipping NCTs, brewed at 56 h, and banana ACT, brewed at 18 h, inhibited seed germination to a level below 80% (Figure ), which is indicative of the presence of phytotoxic compounds (Zucconi et al. Citation1985). These results exposed the potential of germination index values to mask the effect of toxic compounds that may affect germination in instances where root elongation is stimulated by substances in compost teas or extracts.
Figure 8 Effect of aerated (ACT) and nonaerated (NCT) lawn clipping and banana leaf compost teas on relative germination of sweet pepper. Means with different lowercase letters are significantly different across the brewing time within the brewing method and compost type at p ≤ 0.05, according to Tukey's test. Error bars indicate one standard error.
Contrary to the findings of Tam and Tiquia (Citation1994), seed germination was more sensitive than root elongation to the presence of phytotoxic compounds in compost teas. Regression models for both germination index and root elongation of sweet pepper were not significant (p>0.05). However, the regression model for the seed germination rate of sweet pepper indicated that two predictors, Cu and Ca, explained 27% of the variance (R2 = 0.33, p < 0.05). Concentrations of Cu was the most significant factor inhibiting seed germination (β = − 0.88, p < 0.001), whereas Ca was positively related to seed germination (β = 0.80, p < 0.001). Low concentrations of Cu and other trace metals have been reported to inhibit seed germination (Wong and Bradshaw Citation1982). According to Ahsan et al. (Citation2007), the exposure of seeds to excess copper not only affects starvation in water uptake by seeds but also results in failure of the reserve mobilization processes, the result of which is inhibition of seed germination.
Compost teas as liquid fertilizers
Tomato seedlings were more responsive to liquid fertilizer treatments than were sweet pepper seedlings. Results of the tomato trials showed that the main effect of substrate and substrate × liquid fertilizers interaction were not significant (p>0.05). However, the application of Miracle-Gro® or banana leaf NCT at 168 h (BNCT 168 h) resulted in a significant (p < 0.001) mean total dry matter increase of 122%, compared with the nonfertilized water control treatment (Figure ).
Figure 9 Effect of Miracle Gro® and nonaerated banana leaf compost tea brewed for 168 h (BNCT 168 h) on total dry matter of six-week-old tomato and sweet pepper seedlings. Means with different lowercase letters within crop type are significantly different at p ≤ 0.05, according to Tukey's test. Error bars indicate one standard error.
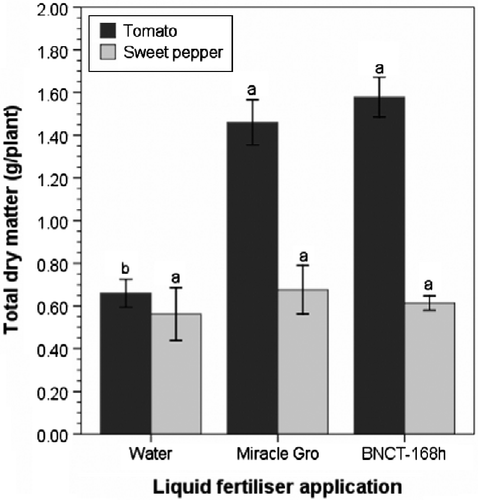
Analysis showed that substrates, nutrient amendments (Miracle Gro® and BNCT 168 h), and substrate × nutrient amendment interaction had no effect on the total dry matter of sweet pepper seedlings (Figure ). These results have serious implications for the time of application of liquid fertilizers and suggest that application of liquid fertilizers one to two weeks after transplanting sweet pepper does not enhance seedling performance.
Conclusions
As the interest in the use of compost tea in commercial horticulture increases, so does the demand for conclusive scientific evidence on factors that can be manipulated to optimize the effectiveness of compost teas. Based on data presented here, it is clear that many of the assertions pertaining to optimum brewing time, phytotoxicity, and the effect of aeration and compost type on the potential use and effectiveness of aerated and nonaerated compost teas need to be revisited.
Shorter brewing cycles of 18 h for ACTs and 56 h for NCTs can be used to produce compost teas with optimal P. ultimum in vitro inhibitive properties using lawn clipping and banana leaf composts. Data showed that ACTs were not consistently more effective than NCTs; rather, the effect of aeration on suppressivity of compost teas was compost type specific.
While further work is needed to determine specific factors responsible for the suppressive effect of compost teas, it is apparent that the presence of particular microorganisms indigenous to compost teas is a prerequisite for inhibition (Koné et al. Citation2010). It is also apparent that microorganisms responsible for the suppressive effect may differ across compost tea type. Further studies on the quantitative relationship between compost tea microbial population and suppressivity (Scheuerell and Mahaffee Citation2006), as well as the mechanism of inhibition, will assist in the development of guidelines for producing suppressive drench formulations for P. ultimum (Scheuerell and Mahaffee Citation2004). Exclusive experimentation on the seed germination and root growth stimulatory properties of compost teas as well as its potential use as a liquid fertilizer on a production scale should result in the production of healthier vegetable seedlings.
Acknowledgements
The authors acknowledge the financial support of the Office for Graduate Studies and Research, Campus Research and Publication Fund, The University of the West Indies, St. Augustine. The authors acknowledge the helpful input and criticism from Drs. G.D. Eudoxie and G. Legall, Mrs. R. Brizan-St. Martin, the editor, and anonymous reviewers.
Related Research Data
References
- Acurio , G , Rossin , A , Teixeira , F and Zepeda , F . 1998 . Diagnosis of municipal solid waste management in Latin America and the Caribbean , Washington (DC) : Pan American Health Organization (PAHO) .
- Ahsan , N , Lee , DG , Lee , SH , Kang , KY , Lee , JJ , Kim , PJ , Yoon , HS , Kim , JS and Lee , BH . 2007 . Excess copper induced physiological and proteomic changes in germinating rice seeds . Chemosphere , 67 : 1182 – 1193 .
- Al-Dahmani , JH , Abbasi , PA , Miller , SA and Hoitink , HAJ . 2003 . Suppression of bacterial spot of tomato with foliar sprays of compost extracts under greenhouse and field conditions . Plant Dis , 87 : 913 – 919 .
- Ayers , RS and Westcot , DW . 1985 . Water quality for agriculture , Rome (Italy) : Food and Agriculture Organization of the United Nations .
- Bailey , DA and Bilderback , T . 1997 . Alkalinity control for irrigation water used in nurseries and greenhouses. Horticulture information leaflets , Raleigh (NC) : North Carolina State University .
- Bernal , MP , Navarro , AF , Roig , A , Cegarra , J and García , D . 1996 . Carbon and nitrogen transformation during composting of sweet sorghum bagasse . Biol. Fert. Soils , 22 : 141 – 148 .
- Blake , GR and Hartge , KH . 1986 . “ Bulk density-core method ” . In Methods of soil analysis. Part 1. Physical and mineralogical methods , Edited by: Klute , EA . 364 – 367 . Madison (WI) : Soil Science Society of America .
- Bremner , JM . 1996 . “ Nitrogen-total ” . In Methods of soil analysis. Part 3. Chemical methods , Edited by: Sparks , DL , Page , AL , Helmke , PA , Loeppert , RH , Soltanpour , PN , Tabatabai , MA , Johnston , CT and Sumner , ME . 1085 – 1121 . Madison (WI) : Soil Science Society of America .
- California Compost Quality Council. 2001. California compost quality council compost maturity index [Internet]. California Compost Quality Council; [cited 2012 May 06]. Available from: http://www.calrecycle.ca.gov/organics/Products/Quality/CompMaturity.pdf
- Cantisano , A . 1998 . Compost teas , Colfax (CA) : Organic Agricultural Advisors Letter .
- Chen , Y , De Nobili , M and Aviad , T . 2004 . “ Stimulatory effects of humic substances on plant growth ” . In Soil organic matter in sustainable agriculture , Edited by: Magdoff , F and Ray , RW . 103 – 129 . Boca Raton (FL) : CRC Press .
- Cronin , MJ , Yohalem , DS , Harris , RF and Andrews , JH . 1996 . Putative mechanism and dynamics of inhibition of the apple scab pathogen Venturia inaequalis by compost extracts . Soil Biol. Biochem , 28 : 1241 – 1249 .
- Davis , JC . 1975 . Waterborne dissolved oxygen requirements and criteria with particular emphasis on the Canadian environment , Ottawa (ON) : National Research Council Canada .
- Dukare , AS , Prasanna , R , Chandra Dubey , S , Nain , L , Chaudhary , V , Singh , R and Saxena , AK . 2011 . Evaluating novel microbe amended composts as biocontrol agents in tomato . Crop Protect , 30 : 436 – 442 .
- El-Mehalawy , AA , Hassanin , SM , Hassanin , NM and Zaki , SA . 2007 . Induction of resistance and biocontrol of Rhizoctonia in cotton against damping-off disease by rhizosphere yeasts and fungi . New Egyptian J. Microbiol , 17 : 148 – 168 .
- El-Tarabily , KA . 2004 . Suppression of Rhizoctonia solani diseases of sugar beet by antagonistic and plant growth-promoting yeasts . J. Appl. Microbiol , 96 : 69 – 75 .
- Gomez , KA and Gomez , AA . 1984 . Statistical procedures for agricultural research , 2nd ed , New York (NY) : John Wiley and Sons .
- Green , SB and Salkind , NJ . 2004 . Using SPSS for Windows and Macintosh: Analyzing and understanding data , 4th ed , Upper Saddle River (NJ) : Prentice Hall .
- Haggag , WM and Saber , MSM . 2007 . Suppression of early blight on tomato and purple blight on onion by foliar sprays of aerated and non-aerated compost teas . Int. J. Food Agric. Environ , 5 : 302 – 309 .
- Hargreaves , JC , Adl , MS and Warman , PR . 2009 . The effects of municipal solid waste compost and compost tea on mineral element uptake and fruit quality of strawberries . Compost Sci. Utiliz , 17 : 85 – 94 .
- Helmke , PA and Sparks , DL . 1996 . “ Lithium, sodium, potassium, rubidium, and cesium ” . In Methods of soil analysis. Part 3. Chemical methods , Edited by: Sparks , DL , Page , AL , Helmke , PA , Loeppert , RH , Soltanpour , PN , Tabatabai , MA , Johnston , CT and Sumner , ME . 551 – 574 . Madison (WI) : Soil Science Society of America .
- Hibar , K , Daami-Remadi , M , Jabnoun-Khiareddine , H , Znaïdi , IEA and El Mahjoub , M . 2006 . Effet des extraits de compost sur la croissance mycélienne et l’ agressivité du Fusarium oxysporum f. sp. radicis-lycopersici [Effects of compost extracts on the mycelial growth and agressiveness of Fusarium oxysporum f. sp. radicis-lycopersici] . Biotechnol Agron Soc Environ , 10 : 101 – 108 .
- Hoitink , HAJ and Fahy , PC . 1986 . Basis for the control of soilborne plant pathogens with composts . Annu. Rev. Phytopathol , 24 : 93 – 114 .
- Howard , RJ , Garland , JA and Seaman , WL . 1994 . Diseases and pests of vegetable crops in Canada , Ottawa (ON) : The Canadian Phytopathological Society .
- Huang , GF , Wong , JWC , Wu , QT and Nagar , BB . 2004 . Effect of C/N on composting of pig manure with sawdust . Waste Manage , 24 : 805 – 813 .
- Ingham , ER and Alms , M . 2003 . The compost tea brewing manual , 4th ed , Corvallis (OR) : Soil Foodweb .
- Jack , ALH and Thies , JE . 2006 . Potential for E. coli survival and replication in aerated and non-aerated compost teas: An analysis of proposed National Organic Program legislation , Santa Cruz (CA) : Organic Farming Research Foundation .
- Johnson , EA . 1999 . “ Anaerobic fermentations ” . In Manual of industrial microbiology and biotechnology , Edited by: Demain , AL and Davies , JE . 139 – 150 . Washington (DC) : ASM Press .
- Katembe , WJ , Ungar , IA and Mitchell , JP . 1998 . Effect of salinity on germination and seedling growth of two Atriplex species (Chenopodiaceae) . Ann. Bot. , 82 : 167 – 175 .
- Ketterer , N . 1990 . Untersuchungen zur wirkung von kompostextracten auf den blattbefall ser kartoffel und tomate durch Phytophthora infestans sowie auf den befall der weinrebe durch Plasmopara viticola, Pseudopeziza tracheiphila und Uncinula necator [Studies of the effect of compost extracts on potato and tomato leaf infections through Phytophthora infestans as well as the incidences of grape disease] [dissertation] , Bonn (Germany) : University of Bonn .
- Ketterer , N , Fisher , B and Weltzien , HC . 1992 . Biological control of Botrytis cinerea on grapevine by compost extracts and their microorganisms in pure culture. 10. International Botrytis Symposium: Recent Advances in Botrytis Research , Heraklion (Crete) : Wageningen Pudoc .
- Ketterer , N and Schwager , L . 1992 . Effect of compost extracts on the disease incidence and the phyllosphere flora of bush bean and tomato leaves . Med van de Fac Land Univ Gent , 57 : 411 – 421 .
- Koné , SB , Dionne , A , Tweddell , RJ , Antoun , H and Avis , TJ . 2010 . Suppressive effect of non-aerated compost teas on foliar fungal pathogens of tomato . Biol. Control , 52 : 167 – 173 .
- Kuo , S . 1996 . “ Phosphorus determination ” . In Methods of soil analysis. Part 3. Chemical methods , Edited by: Sparks , DL , Page , AL , Helmke , PA , Loeppert , RH , Soltanpour , PN , Tabatabai , MA , Johnston , CT and Sumner , ME . 899 – 920 . Madison (WI) : Soil Science Society of America .
- Litterick , A and Wood , M . 2009 . “ The use of composts and compost extracts in plant disease control ” . In Disease control in crops: Biological and environmentally friendly approaches , Edited by: Walters , D . 93 – 121 . Oxford (UK) : Wiley-Blackwell .
- McCarty , PL . 1964 . Anaerobic waste treatment fundamentals . Pub Works , 95 : 107 – 112 .
- Minnich , J and Hunt , M . 1979 . The Rodale guide to composting , Emmaus (PA) : Rodale Press .
- Munns , R and Tester , M . 2008 . Mechanisms of salinity tolerance . Annu Rev Plant Biol , 59 : 651 – 681 .
- Nardi , S , Pizzeghello , D , Muscolo , A and Vianello , A . 2002 . Physiological effects of humic substances on higher plants . Soil Biol. Biochem , 34 : 1527 – 1536 .
- National Organic Standards Board . 2004 . Compost tea task force report , Washington (DC) : National Organic Standards Board .
- Nelson , DW and Sommers , LE . 1996 . “ Total carbon, organic carbon, and organic matter ” . In Methods of soil analysis. Part 3. Chemical methods , Edited by: Sparks , DL , Page , AL , Helmke , PA , Loeppert , RH , Soltanpour , PN , Tabatabai , MA , Johnston , CT and Sumner , ME . 961 – 1010 . Madison (WI) : Soil Science Society of America .
- Punja , ZK . 1997 . Comparative efficacy of bacteria, fungi, and yeasts as biological control agents for diseases of vegetable crops . Can J Plant Pathol , 19 : 315 – 323 .
- Reed , ST and Martens , DC . 1996 . “ Copper and zinc ” . In Methods of soil analysis. Part 3. Chemical methods , Edited by: Sparks , DL , Page , AL , Helmke , PA , Loeppert , RH , Soltanpour , PN , Tabatabai , MA , Johnston , CT and Sumner , ME . 703 – 722 . Madison (WI) : Soil Science Society of America .
- Russell , SJE . 1988 . Soil conditions and plants growth , Harlow (UK) : Longman Science & Technology .
- Scheuerell , SJ and Mahaffee , WF . 2002 . Compost tea: Principles and prospects for plant disease control . Compost Sci. Utiliz , 10 : 313 – 338 .
- Scheuerell , SJ and Mahaffee , WF . 2004 . Compost tea as a container medium drench for suppressing seedling damping-off caused by Pythium ultimum . Phytopathol , 94 : 1156 – 1163 .
- Scheuerell , SJ and Mahaffee , WF . 2006 . Variability associated with suppression of gray mold (Botrytis cinerea) on Geranium by foliar applications of nonaerated and aerated compost teas . Plant Disease , 90 : 1201 – 1208 .
- Sharrock , S and Frison , E . 1998 . Musa production around the world-trends, varieties and regional importance. International Network for the Improvement of Banana and Plantain annual report , Wageningen (The Netherlands) : The Technical Centre for Agricultural and Rural Cooperation (CTA) .
- Shih , JCH . 1987 . Ecological benefits of anaerobic digestion . Poultry Sci , 66 : 946 – 950 .
- Skidmore , AM and Dickinson , CH . 1976 . Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi . Trans. British Mycol. Soc , 66 : 57 – 64 .
- St. Martin , CCG and Brathwaite , RAI . 2012 . Compost and compost tea: Principles and prospects as substrates and soil-borne disease management strategies in soil-less vegetable production . Biol. Agric. Hort , 28 : 1 – 33 .
- Suarez , DL . 1996 . “ Beryllium, magnesium, calcium, strontium, and barium ” . In Methods of soil analysis. Part 3. Chemical methods , Edited by: Sparks , DL , Page , AL , Helmke , PA , Loeppert , RH , Soltanpour , PN , Tabatabai , MA , Johnston , CT and Sumner , ME . 575 – 601 . Madison (WI) : Soil Science Society of America .
- Tam , NFY and Tiquia , S . 1994 . Assessing toxicity of spent pig litter using a seed germination technique . Resour. Conserv. Recycl , 11 : 261 – 274 .
- Thompson , WH . 2002 . Test Methods for the Examination of Composting and Compost (TMECC) , Washington (DC) : United States Composting Council and U.S. Department of Agriculture, U.S. Government Printing Office .
- Topper , PA , Graves , RE and Richard , T . 2006 . The fate of nutrients and pathogens during anaerobic digestion of dairy manure , Lehman (PA) : Penn State University. College of Agricultural Science, Cooperative Extension Bulletin G 71 .
- Tränkner , A . 1992 . “ Use of agricultural and municipal organic wastes to develop suppressiveness to plant pathogens ” . In Biological control of plant diseases , Edited by: Tjamos , ES , Papavizas , GC and Cook , RJ . 35 – 42 . New York (NY) : Plenum Press .
- Tränkner , A and Liesenfeld , R . 1990 . Unterdrueckung von Pythium ultimum, einer keimlingskrankheit der Erbse (Pisum sativum) durich Saatgutbehandlung mit Kompostextrakten [Suppression of Pythium ultimum, a seedling disease of pea (Pisum sativum) seed treatment with compost extract] . Mitt. Biol. Bundesanst Land Forstwirtsch , 266 : 64
- Whipker , B . 1999 . Irrigation water quality for container-grown plants , Ames (IA) : Iowa University Extension .
- Wilkie , AC . 2005 . “ Anaerobic digestion: Biology and benefits ” . In Dairy manure management: Treatment, handling, and community relations. NRAES-176 , 63 – 72 . Ithaca (NY) : Natural Resource, Agriculture, and Engineering Service, Cornell University .
- Wong , MH and Bradshaw , AD . 1982 . A comparison of the toxicity of heavy metals, using root elongation of rye grass, Lolium perenne . New Phytol , 91 : 255 – 261 .
- Woods End Research Laboratories . 2002 . Guide to Solvita® testing for compost maturity index , Mt Vernon (ME) : Woods End Research Laboratories .
- Zhao , B and Chen , S . 2003 . Ammonia volatilization from dairy manure under anaerobic and aerated conditions at different temperature , St. Joseph (MI) : ASABE . ASABE Paper No. 421 034148
- Zucconi , F , Monaco , A , Forte , M and de Bertoldi , M . 1985 . “ Phytotoxins during the stabilization of organic matter ” . In Composting of agricultural and other wastes , Edited by: Gasser , JKR . 73 – 86 . London (UK) : Elsevier Applied Science Publishers .