
ABSTRACT
Repeated sprint ability (RSA) is more closely related to match performance outcomes than single-sprint performance, but the kinetic determinants in youth athletes remain poorly understood. Therefore, the aim of the study was to explore the kinetic determinants of RSA in youth athletes. Twenty trained adolescents (15 girls; 14.4 ± 1.0 years) completed five 15 m repetitions interspersed with 5-s rest. Velocity was measured during each trial using a radar gun at >46 Hz, following which the force–velocity–power (F-v-P) profile was fitted to a velocity–time curve and instantaneous power and force variables calculated. The mechanical efficiency of force application (DRF) was the primary predictor of both single and repeated sprint performance in adolescents. Secondly, hierarchical analyses revealed the percentage reduction in peak velocity, DRF, and allometrically scaled peak force explained 91.5% of the variance in 15 m sprint time from sprints 1–5. Finally, declines in allometrically scaled peak power were more closely related to declines in peak force than reductions in velocity. In conclusion, given DRF was the primary predictor of both single and repeated sprint performance training programmes targeting RSA need to include technique, and skill acquisition, components.
KEYWORDS:
1. Introduction
Repeated sprint ability (RSA), defined as the ability to repeatedly reproduce consistent maximal efforts (Girard et al., Citation2011), is fundamental to athletic performance in team sports and routinely assessed in long-term athlete development (LTAD) programmes and talent identification batteries (Girard et al., Citation2011; Mendez-Villanueva et al., Citation2010; Moran et al., Citation2016; Papaiakovou et al., Citation2009). Indeed, adult athletes are reported to perform more than 50 individual sprints during an international football match (Pons et al., Citation2021), with many being in close succession and almost always in game critical scenarios (i.e., scoring or preventing a goal; Spencer, Bishop, et al., Citation2004). Unsurprisingly, therefore, RSA has been more closely related to performance outcomes than individual sprint ability in adult team sports (Spencer, Bishop, et al., Citation2004; Spencer, Lawrence, et al. Citation2004). Youth team sports are characterised by similar movement profiles (Buchheit et al., Citation2010; McLellan & Lovell, Citation2013), but the determinants of RSA are likely to be dependent on both chronological and biological age, although this requires further investigation. Indeed, determining the mechanisms of RSA in youth athletes would be pertinent for coaches, talent identification, and the development of more comprehensive LTAD models. However, obtaining a reliable measure of RSA is difficult given the high intra- and inter-day variability (17–51% in some cases) and the number of different RSA protocols which are routinely used confounding direct comparisons between studies (Altmann et al., Citation2019).
The development of single-sprint performance during childhood and adolescence is thought to be a non-linear process, with accelerated periods of development around the time of peak height velocity (PHV; Mendez-Villanueva et al., Citation2010; Mujika et al., Citation2009; Philippaerts et al., Citation2006; Spencer et al., Citation2011). However, changes in RSA and its determinants during maturation are less well understood (Mujika et al., Citation2009; Rossi et al., Citation2017). The determinants of RSA have been postulated to be a combination of physiological (i.e., VO2max; Girard et al., Citation2011; Meckel et al., Citation2013; Rumpf et al., Citation2013) and biomechanical factors (Girard et al., Citation2011; Morin et al., Citation2006, Citation2011; Rossi et al., Citation2017; Rumpf et al., Citation2013; Rumpf, Cronin, Oliver, et al., Citation2015; Samozino et al., Citation2016). From a biomechanical perspective, sprint performance is suggested to be directly proportional to peak horizontal force (Fpeak) and peak power (Ppeak) during the initial acceleration (Morin et al., Citation2011; Rabita et al., Citation2015).
An athlete’s capacity to produce Fpeak whilst running is well described by the force–velocity relationship (Morin et al., Citation2006; Rossi et al., Citation2017; Samozino, Citation2018) which characterises the theoretical limits of the entire neuromuscular system and the theoretical exponents of Fpeak, Ppeak and peak velocity (Vpeak; Samozino et al., Citation2016). These mechanical variables (Fpeak, Ppeak, and Vpeak) appear to be the primary determinants of single-sprint performance, irrespective of age or maturity (r2: 0.98–0.99; Rumpf, Cronin, Oliver, et al., Citation2015). However, the relative influence of Fpeak, Ppeak and Vpeak on RSA is less clear and the underpinning mechanisms are thought to be maturity dependant (Meyers et al., Citation2015, Citation2017a; Papaiakovou et al., Citation2009; Philippaerts et al., Citation2006). For instance, vertical stiffness, a reflection of the ability to tolerate and overcome gravitational forces, was reported to be the greatest predictor of sprint performance in pubertal adolescents but not their pre- or post-pubertal counterparts (Rumpf, Cronin, Mohamad, et al., Citation2015; Rumpf, Cronin, Oliver, et al., Citation2015). Whilst the generalisability of these results is limited by the derivation of the biomechanical parameters from only the four fastest consecutive steps on a non-motorised treadmill, they raise interesting questions regarding the interaction between maturity and kinetic determinants of performance.
The mechanisms underpinning Fpeak and Ppeak are multifaceted and represent a complex interaction between neural, anthropometric, and morphological factors, alongside individual muscle properties (Morin et al., Citation2006, Citation2011; Rabita et al., Citation2015). Interestingly, the technical ability to apply force (DRF) has recently been postulated to be more influential than absolute Fpeak (Morin et al., Citation2011; Rossi et al., Citation2017), highlighting that technique and skill proficiency may also be of importance. Whilst the influence of DRF has been reported in world-class (Rabita et al., Citation2015) and masters athletes (Slawinski et al., Citation2017), little is known about its role in sprint performance in youth athletes. It was recently reported that single-sprint DRF improved with age in a sample of 68 children and adolescents (Rossi et al., Citation2017). However, whether DRF is equally important for multiple-sprint performance remains largely unknown in youth athletes and thus further research is required to delineate the potential effects of age and maturation and their interaction with training on the determinants of sprint performance. Therefore, the aim of this study was to assess RSA using radar technology and biomechanical modelling in trained children and adolescents to determine the underlying kinetics during repeated over-ground sprints.
2. Methods
Twenty adolescents (n = 15 girls; ) involved in international age-group field-hockey tournaments and an LTAD programme participated in the study. A pre-screening medical questionnaire and informed parent/guardian consent were completed online using a custom-built consent form (Survey Monkey, Dublin, Ireland). Written participant assent was obtained on the day of testing. Ethics approval was obtained from the institutional ethics committee (approval number: SWA_2019_18) and the study conformed to the Declaration of Helsinki.
Table 1. Participant descriptives.
2.1 Experimental procedures
Standing and sitting stature were measured to the nearest 0.1 cm using a Seca 213 portable stadiometer (Seca 213, Seca, Chino, CA, USA), with body mass measured to the nearest 0.1 kg using a set of electronic scales (Seca 803, Seca, Chino, CA, USA). Maturity offset was subsequently calculated using the equations of Mirwald et al. (Citation2002).
The purpose of the repeated sprint protocol was to mimic gameplay scenarios where near maximal sprints are completed in quick succession with changes in direction. Velocity was measured throughout all sprints using a STALKER ATS II radar gun (STALKER, Plano, Texas, USA), mounted on a tripod positioned 10 m behind the start line, in accord with manufacturer recommendations. The STALKER ATS II has a recording frequency of 46.875 Hz, allowing for near-instantaneous power and force variables to be modelled. Rossi et al. (Citation2017) used radar technology coupled with macroscopic biomechanical modelling to estimate force and power variables in field-based settings. This radar technology has the ability to measure both inbound and outbound velocities (Simperingham et al., Citation2016, Citation2017), thereby enabling the quantification of between sprint differences. Indeed, such high-resolution quantification of the kinetics underpinning repeated sprints could not only further our understanding of, but also facilitate targeted interventions to improve, RSA during childhood and adolescence (Girard et al., Citation2011; Samozino et al., Citation2016; Simperingham et al., Citation2017). Moreover, radar technology has demonstrated high intraday reliability in paediatric populations (Runacres et al., Citation2019).
Prior to undertaking the repeated-sprint protocol, participants undertook a standardised five-minute low-intensity warm-up, culminating in two 15 m sprints that simultaneously served as familiarisation with the sprint protocol. Participants were then given at least 2 min of active rest before completing the repeated-sprint protocol. The 20 m sprinting lane was marked by a pair of coloured cones and prior to undertaking the RSA protocol participants had the opportunity to ask any questions. For the repeated-sprint protocol, participants completed five 20 m sprints, turning 180° every 20 m. To minimise the potential confounding effects of differences in turning speed between participants and sprints, participants were required to stop for up to 5 s before accelerating into the next maximal sprint. A repeated measures ANOVA revealed that there was no significant difference in the times in between sprints. Sprint times and kinetic variables were subsequently derived from the initial 15 m of each sprint to minimise the effects of deceleration. Participants started from a two-point standing start to reduce vertical displacement during the early phase of the sprint and were instructed to start using auditory cues (i.e., “3 … 2 … 1 … GO!”). All sprints were completed on outdoor AstroTurf pitches, with the mean air temperature and wind speed being 15.8 ± 0.8ºC and 1.6 ± 0.8 m ⋅ s−1, respectively. Verbal encouragement was given to each participant for the duration of the RSA protocol to ensure that all participants completed the RSA protocol to the best of their ability.
2.2 Data analysis and statistics
Radar technology has the ability to measure both inbound and outbound velocities (Simperingham et al., Citation2016, Citation2017), thereby enabling the quantification of between sprint differences. Indeed, such high-resolution quantification of the kinetics underpinning repeated sprints could not only further our understanding of but also facilitate targeted interventions to improve, RSA during childhood and adolescence (Girard et al., Citation2011; Samozino et al., Citation2016; Simperingham et al., Citation2017). To assess RSA, the data was first segmented into five sections to represent each repeated sprint and, subsequently, the first reading of each new sprint was assigned time 0, and the first 0.3 s deleted in line with previous recommendations (Samozino, Citation2018). Using the biomechanical model of Samozino et al. (Citation2016), the following parameters were derived for each sprint: time to peak power (t_Ppeak), absolute, relative and allometrically scaled peak power (Ppeak), mean power (Pmean), relative mean power (relative Pmean), peak and mean velocity (Vpeak and Vmean, respectively), 15 m sprint time (15mT), absolute, relative, and allometrically scaled peak force (Fpeak), fatigue rate (FR) and DRF. Furthermore, to aid comparisons with previous literature, the fatigue index (FI) was calculated using the formula reported elsewhere (Mujika et al., Citation2009).
All statistical analyses were conducted in SPSS (Version 26.0, IBM, Armonk, NY, USA), with values presented as mean ± SD. Sex differences in anthropometric variables were assessed using an independent samples t-test, revealing no sex differences in any anthropometric or repeated sprint parameter, so data were pooled for subsequent analyses. Allometric scaling, a common technique to account for body size differences in children (Welsman & Armstrong, Citation2019), was performed on Ppeak and Fpeak, with a b exponent of 1.54 and 1.87 for Ppeak and Fpeak, respectively. Following confirmation that all variables were normally distributed parametric tests were used throughout. A repeated measures ANOVA was used to compare performance parameters between sprints, with Bonferroni corrections applied where necessary. Cohen's d effect sizes were also calculated to determine trivial (≤0.20), moderate (≥0.21–≤ 0.60), large (≥0.61–≤ 0.80), and very large (≥0.81) effects (Cohen, Citation1988).
Hierarchical linear regressions were used to ascertain the biomechanical determinants of each 15 m sprint repetition. Allometrically scaled Ppeak was initially added to the model due to the emerging associations with single-sprint performance in children and adolescents (Rumpf, Cronin, Mohamad, et al., Citation2015; Rumpf, Cronin, Oliver, et al., Citation2015). Subsequently, predictor variables were entered into the models to determine the independent association with each repetition’s 15mT, with inclusion into the model accepted if a significant increase in explained variance was observed at the 0.05 confidence interval. Collinearity checks were conducted using the variance inflation factor (VIF), with a VIF <1, between 1 and 5, and greater than 5 indicating low, moderate and high collinearity, respectively (Daoud, Citation2017). If high multi-collinearity was found between variables, the variable explaining the greatest proportion of variance was included in the model (Daoud, Citation2017). The accuracy and suitability of the model was assessed using the normality of residual values. The correlation coefficients were classified according to Hopkins Citation(2006) as trivial (0.0–0.1), small (0.1–0.3), moderate (0.3–0.5), large (0.5–0.7), very large (0.7–0.9), or nearly perfect (0.9–1.0). A significant correlation was evident between the percentage decline in 15mT and allometrically scaled Ppeak (R2 = 0.76; very large, p < 0.01) but Ppeak was not entered into the regression model due to its high collinearity with Vmax and Fpeak. Declines in Fpeak were more strongly related to declines in Ppeak during the repeated sprints (R2 = 0.89; very large, p < 0.01) than changes in Vmax (R2 = 0.42; moderate, p < 0.05).
To assess the overall RSA, the mean percentage decline from the first to last sprint was calculated for each mechanical variable, similar to Chaouachi et al. (Citation2010), with a subsequent hierarchical stepwise linear regression used to ascertain which variables predicted the decline in performance according to the 15mT. Pearson’s correlations were performed to establish the relationship between the relative declines in kinetic variables over the repeated sprints.
3. Results
The repeated measures ANOVA demonstrated a main effect for sprint number, with post-hoc analyses demonstrating that, as shown in , this was primarily attributable to differences between the first two sprints and all subsequent sprints. More specifically, there was a main effect for Ppeak regardless of how it was expressed, with post-hoc tests revealing Ppeak during sprint 1 was significantly higher than sprint 4 (d = 0.54; ). Pmean, relative Pmean, and DRF were significantly higher during sprint 1 compared to all other sprints (Pmean: F(4,76) = 23.7, p < 0.01, relative Pmean: F(4,76) = 24.7, p < 0.01, DRF: F(4,76) = 2.6, p < 0.01). There was an overall effect of sprint number on Vpeak (F(4,76) = 29.4, p < 0.01), Vmean (F(4,76) = 17.4, p < 0.01) and 15mT (F(4,76) = 17.1, p < 0.01), with post-hoc analyses revealing that Vpeak, Vmean and 15mT decreased from sprints 1 to 4, before increasing slightly in sprint 5 (). However, as shown in , there were no significant differences in Fpeak, irrespective of whether it was expressed in absolute, relative, or allometrically scaled units, over the five sprint repetitions (all p > 0.05). Additionally, t_Ppeak and FR did not significantly change with each sprint repetition. The mean FI over the five sprint repetitions was 11.0 ± 3.9%.
Figure 1. Post-processed representative traces from the five sprint-repetitions for a typical participant showing a) velocity-time profile; b) power-velocity profile; and c) the force velocity profile.
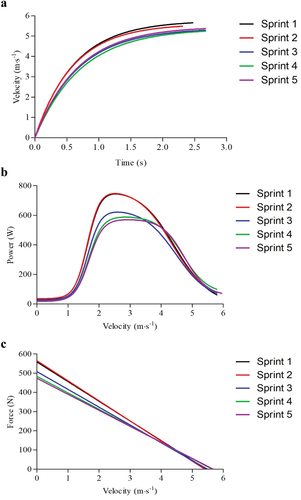
Table 2. Sprint variables from each of the 5 × 15 m sprint repetitions.
3.1 Biomechanical determinants of repeated sprints
In model 1, in which only allometrically scaled Ppeak was entered, 74–82% of the variance in the 15mT from sprints 1 to 4 was explained, with the explained variance lower for sprint 5 only (62%; ). DRF was found to be a significant additional predictor of sprint performance for sprints 1, 2, 3 and 5, increasing the explained variance to between 84% and 92%. Mean allometrically scaled Ppeak across the five sprints explained a large proportion of the variance in the mean 15mT (81%), with the inclusion of mean DRF significantly increasing the explained variance to 90%. The hierarchical linear regression revealed that percentage declines in Vmax was the strongest predictor of 15mT (37.9%), followed by DRF (34.7%) and allometrically scaled Fpeak (18.9%). Subsequently, the percentage decline in 15mT was described by:
Table 3. Biomechanical determinants of 15 m sprint time for each repetition.
Percentage decline in 15mT = −0.903 + (0.629 * Vmax) + (0.143 * DRF) + (0.415 * allometrically scaled Fpeak)
4. Discussion
This study was the first to utilise radar technology in combination with macroscopic biomechanical modelling to gain greater insights into the mechanical properties underpinning repeated-sprint ability in adolescents. The main findings of the current study indicate that declines in allometrically scaled Ppeak with repeated sprints are more closely related to declines in allometrically scaled Fpeak than peak velocity in circa-PHV adolescents. Moreover, the hierarchical analysis revealed that, irrespective of sprint number, the kinetic determinants of single sprints were also key to RSA performance. Finally, DRF was the primary predictor of both single- and multiple-sprint kinetics. The results of this study provide important insights for the design and implementation of training interventions targeting improvements in RSA in youth athletes.
Data from non-motorised treadmills in boys (Rumpf, Cronin, Oliver, et al., Citation2015) and force platforms in girls (Nagahara et al., Citation2019) indicate that Ppeak and Fpeak are key determinants of single-sprint performance during youth. In contrast, the present study suggests that DRF, an indication of mechanical efficiency, is a greater predictor of single-sprint performance than Fpeak. This is in agreement with data in trained youth (Rossi et al., Citation2017) and adult (Morin et al., Citation2011) sprinters, which was attributed to Fpeak being the sum of both horizontal and vertical forces, with the latter not significantly influencing performance (Morin et al., Citation2006, Citation2011). However, DRF represents the linear decline in the ratio of forces (horizontal: vertical) with increasing velocity and may therefore be more performance-orientated (Samozino et al., Citation2016; Slawinski et al., Citation2017). Of note, DRF has also been reported to significantly decline in repeated cycling sprints, whereby participant’s force-efficiency decreased as pedalling frequency increased (Sanderson, Citation1991). Whilst it is not possible to exert zero vertical force (and subsequently have a ratio of forces of 100%) as this would preclude running motion (Morin et al., Citation2006, Citation2011), future research should seek to establish optimal values that could improve both single and repeated sprints.
Allometrically scaled Ppeak and peak velocity were significantly higher in sprint 1 compared to all other sprints (d = 0.54–1.16), with the decline in allometrically scaled Ppeak reaching significance from sprint 4. In contrast, there were no significant differences between any sprints for Fpeak, regardless of how it was expressed. Therefore, it was surprising that declines in Fpeak were the primary cause of the reductions in Ppeak over the course of the five sprints. Nevertheless, previous research in children has reported that relative Fpeak has a very strong relationship with step length and flight length (the distance travelled by the COM from toe-off to touchdown; Lloyd et al., Citation2016). Thus, it may be possible that even small changes in relative Fpeak may negatively impact upon these crucial kinematic variables, thereby lowering velocity (Lloyd et al., Citation2016; Meyers et al., Citation2017b). Of note, allometrically scaled Fpeak and Ppeak explained significantly more variance in both individual sprint 15mT and the overall decline in 15mT across the five sprint repetitions than absolute or relative values. This may be due to the near-linear relationship observed between Ppeak, Fpeak and body mass, irrespective of sex (Doré et al., Citation2005, Citation2008), with ratio-scaling consequently penalising heavier, more mature individuals (Nevill, Bate, & Holder, Citation2005; Welsman & Armstrong, Citation2019).
Allometric scaling, unlike conventional ratio scaling, allows the specific exponent of body mass to be calculated, and is currently the most robust statistical method to account for body mass differences in aerobic fitness (Welsman & Armstrong, Citation2019). Allometric scaling was utilised within this study to provide reference values for future studies exploring the kinetic determinants of RSA in circa-PHV adolescents. Additionally, by employing allometric scaling, comparisons of kinetic RSA determinants across maturity groups are facilitated, which is especially relevant as it is likely that the determinants may be maturity dependent. Indeed, circa-PHV adolescents are often characterised by “adolescent awkwardness” caused by rapid growth of limbs in relation to the trunk inhibiting motor control (Quatman-Yates et al., Citation2012). Consequently, the potential reductions in motor control may explain the importance of DRF for RSA in this study. However, future research is warranted to establish the kinetic determinants of RSA throughout maturity.
In the present study, similar mechanical determinants of single and repeated sprints were evident, therefore suggesting that single- and repeated-sprint ability may be governed by the same mechanical properties. From a physiological perspective, declines in Ppeak during repeated cycling sprints have primarily been attributed to neuromuscular fatigue, arising from increases in blood lactate concentrations and associated reductions in intramuscular pH (Ratel et al., Citation2002; Ratel, Duche, et al., Citation2006). Such reductions inhibit the ability to recruit type II higher-order muscle fibres, with changes in motor-unit recruitment patterns in the quadriceps, measured using an electromyogram, explaining 97% of the total work during repeated cycling sprints, interspersed with 30 s rest in untrained adults (Girard et al., Citation2011). Moreover, a reduction in pH may lead to reductions in motor co-ordination (Doré et al., Citation2005; Ratel, Williams, et al., Citation2006), which may also explain DRF as a predictor of fatigue during repeated sprints. However, although similar fatiguing pathways seem likely (Ratel, Duche, et al., Citation2006), whether the same physiological mechanisms are also responsible for the declines in running performances remains to be established.
The trained hockey players in the present study showed a higher decrement in performance, as measured using FI (11.9%), to that reported in highly trained footballers (4.1–5.5%; Girard & Farooq, Citation2012; Mujika et al., Citation2009), likely due to differences in the repeated-sprint protocol as both groups were highly trained completing ≥7 hours ∙ week−1. Specifically, most repeated-sprint studies incorporate 10–30 s of rest between repetitions (Meckel et al., Citation2009; Mendez-Villanueva et al., Citation2010; Mujika et al., Citation2009; Temfemo et al., Citation2011), whereas this study utilised a near-continuous protocol. Whilst it could be argued that incorporating rest between sprints is more indicative of team-sport scenarios (Mendez-Villanueva et al., Citation2010), the inclusion of rest periods facilitates aerobic recovery. Indeed, previous paediatric research found a significant correlation between aerobic capacity and fatigue resistance during repeated-sprint protocols (Mendez-Villanueva et al., Citation2007). Moreover, Dupont et al. (Citation2005) reported that the magnitude of change in sprint time was negatively correlated with the speed of the pulmonary VO2 kinetics in adults (r2 = 0.80, p < 0.01), with faster pulmonary VO2 kinetics postulated to spare intramuscular phosphocreatine for the later sprints and thereby increase RSA performance (Dupont et al., Citation2005). Therefore, future research should seek to establish physiological determinants of repeated-sprint performance in children and adolescents to further explain the declines in mechanical variables during sprint running.
It may be pertinent to note that the last sprint (sprint 5) was faster than sprints 2, 3 and 4, perhaps highlighting that future research should incorporate an increased number of sprint repetitions to induce greater fatigue, as well as the potential role of pacing resulting in sub-maximal sprints (Impellizzeri et al., Citation2008; Ratel, Williams, et al., Citation2006). In the absence of criteria to determine the maximal effort, it could be postulated that an element of pacing is involved within all repeated-sprint protocols, irrespective of recovery period. The present study utilised specific strategies to increase and maintain motivation during repeated sprints, including a longer finish line distance (Mendez-Villanueva et al., Citation2010) and verbal encouragement (Mujika et al., Citation2009; Philippaerts et al., Citation2006). However, no studies have examined differences between trials with and without motivational techniques, so it is unclear whether these techniques mitigate the role of pacing.
Although there are numerous strengths to this study, certain limitations must be noted. Data collection was curtailed by COVID-19 resulting in a relatively small sample size, thereby limiting generalisability. Given the small sample size, the results of this study should be interpreted as a preliminary investigation into the potential underpinning mechanisms of RSA in highly trained youth. Furthermore, all participants were classified as circa peak height velocity and so the results of this study should be interpreted as the kinetic determinants of RSA in this maturity group only. Whilst the macroscopic biomechanical model used provides an overview of the kinetics underpinning sprint performance, muscle-specific inferences cannot be made. Consequently, the specific muscles responsible for the observed power reduction cannot be established, limiting intervention specificity. Nevertheless, radar technology coupled with biomechanical modelling offers a foundation for quantifying potential sex, training and maturational differences in RSA.
4.1 Practical recommendations
The results of this study indicate that DRF was the most important factor in determining RSA, suggesting that future interventions and training programmes seeking to improve RSA in youth should incorporate a focus on technique and skill acquisition components. This is important as many LTAD models focus almost exclusively on physiological conditioning with little consideration for the wider determinants of talent development. Furthermore, coaches and practitioners working with youth team sport athletes should endeavour to incorporate strength, or power, training into their training regimes as Ppeak was found to be a key determinant of 15mT.
5. Conclusions
In conclusion, DRF was the primary determinant of both single- and repeated-sprint performance in youth and therefore future training interventions aiming to improve RSA in youth should include some technique, or skill acquisition, components. Future studies should seek to establish the RSA development in untrained children and adolescents, so the trainability of RSA, and any sex, or maturity, differences can be established.
Highlights
Mechanical efficiency (DRF) was the primary determinant of single- and repeated-sprint performance in youth.
Declines in peak power with repeated sprints were more closely related to reductions in peak force than velocity.
Peak velocity, DRF, and allometrically scaled peak force explained 91.5% of the variance in RSA.
Ethics approval
Ethics approval was granted by the A-STEM Ethics Committee (Approval Number: SWA_2019_18), and the study was conducted in accordance with the Declaration of Helsinki.
Acknowledgments
The authors would like to thank Dr Rachel Hughes and Tim Evans for their help in the initial design of this study. The authors would also like to thank all the athletes, coaches, and parents without whom this research would not have been possible.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data is available from the corresponding author (Professor Melitta McNarry; [email protected]) upon reasonable request.
Additional information
Funding
References
- Altmann, S., Ringhof, S., Neumann, R., Woll, A., Rumpf, M. C., & Mirkov, D. (2019). Validity and reliability of speed tests used in soccer: A systematic review. PLos One, 14(8), e0220982. https://doi.org/10.1371/journal.pone.0220982
- Buchheit, M. V., Mendez-Villanueva, A., Simpson, B. M., & Bourdon, P. C. (2010). Repeated-sprint sequences during youth soccer matches. International Journal of Sports Medicine, 31(10), 709–716. https://doi.org/10.1055/s-0030-1261897
- Chaouachi, A., Manzi, V., Wong, D. P., Chaalali, A., Laurencelle, L., Chamari, K., & Castagna, C. (2010). Intermittent endurance and repeated sprint ability in soccer players. The Journal of Strength & Conditioning Research, 24(10), 2663–2669. https://doi.org/10.1519/JSC.0b013e3181e347f4
- Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). Erlbaum.
- Daoud, J. (2017). Multicollinearity and regression analysis. Journal of Physics Conference Series, 949, 012009. https://doi.org/10.1088/1742-6596/949/1/012009
- Doré, E., Bedu, M., & Van Praagh, E. (2008). Squat Jump performance during growth in both sexes: Comparison with cycling power. Research Quarterly for Exercise & Sport, 79(4), 517–524. https://doi.org/10.1080/02701367.2008.10599518
- Doré, E., Martin, R., Ratel, S., Duché, P., Bedu, M., & Van Praagh, E. (2005). Gender differences in peak muscle performance during growth. International Journal of Sports Medicine, 26(4), 274–280. https://doi.org/10.1055/s-2004-821001
- Dupont, G., Millet, G., Cuinhouya, C., & Berthoin, S. (2005). Relationship between oxygen uptake kinetics and performance in repeated running sprints. European Journal of Applied Physiology, 95(1), 27–34. https://doi.org/10.1007/s00421-005-1382-8
- Girard, O., & Farooq, A. (2012). Effects of Ramadan fasting on repeated sprint ability in young children. Science & Sports, 27(4), 237–240. https://doi.org/10.1016/j.scispo.2011.09.006
- Girard, O., Mendez-Villanueva, A., & Bishop, D. (2011). Repeated-sprint ability - part i factors contributing to fatigue. Sports Medicine, 41(8), 673–694. https://doi.org/10.2165/11590550-000000000-00000
- Hopkins, W.G. (2006). Estimating sample size for magnitude-based inferences. Sportscience, 10, 63–70.
- Impellizzeri, F., Rampinini, E., Castagna, C., Bishop, D., Ferrari-Bravo, D., Tibaudi, A., & Wisloff, U. (2008). Validity of a repeated-sprint test for football. International Journal of Sports Medicine, 29(11), 889–905. https://doi.org/10.1055/s-2008-1038491
- Lloyd, R., Meyers, R., Hughes, M., Cronin, J., & Oliver, J. (2016). The relationship between vertical and horizontal stiffness and maximal running speed in 11 - 15 year old boys. The Journal of Strength & Conditioning Research, 30, S99.
- McLellan, C. P., & Lovell, D. I. (2013). Performance analysis of professional, semiprofessional, and junior elite rugby league match-play using global positioning systems. The Journal of Strength & Conditioning Research, 27(12), 3266–3274. https://doi.org/10.1519/JSC.0b013e31828f1d74
- Meckel, Y., Bishop, D., Rabinovich, M., Kaufman, L., Nemet, D., & Eliakim, A. (2013). Repeated sprint ability in elite water polo players and swimmers and its relationship to aerobic and anaerobic performance. Journal of Sports Science & Medicine, 12(4), 738–743.
- Meckel, Y., Machnal, O., & Eliakim, A. (2009). Relationship among repeated sprint tests, aerobic fitness, and anaerobic fitness in elite adolescent soccer players. The Journal of Strength & Conditioning Research, 23(1), 163–169. https://doi.org/10.1519/JSC.0b013e31818b9651
- Mendez-Villanueva, A., Buchheit, M., Kuitunen, S., Douglas, A., Peltola, E., & Bourdon, P. (2010). Age-related differences in acceleration, maximal running speed, and repeated sprint-performance in young soccer players. Journal of Sports Science, 29(5), 477–484. https://doi.org/10.1080/02640414.2010.536248
- Mendez-Villanueva, A., Hamer, P., & Bishop, D. (2007). Fatigue responses during repeated sprints matched for initial mechanical output. Medicine & Science in Sport and Exercise, 39(12), 2219–2225. https://doi.org/10.1249/mss.0b013e31815669dc
- Meyers, R., Oliver, J., Hughes, M., Cronin, J. B., & Lloyd, R. (2015). Maximal sprint speed in boys of increasing maturity. Pediatric Exercise Science, 27(1), 85–94. https://doi.org/10.1123/pes.2013-0096
- Meyers, R., Oliver, J., Hughes, M., Lloyd, R., & Cronin, J. (2017a). Influence of age, maturity, and body size on the spatiotemporal determinants of maximal sprint speed in boys. The Journal of Strength & Conditioning Research, 31(4), 1009–1016. https://doi.org/10.1519/JSC.0000000000001310
- Meyers, R., Oliver, J., Hughes, M., Lloyd, R., & Cronin, J. (2017b). New insights into the development of maximal sprint speed in male youth. Journal of Strength and Conditioning, 39(2), 2–10. https://doi.org/10.1519/SSC.0000000000000290
- Mirwald, R. L., Baxter-Jones, A. D. G., Bailey, D. A., & Beunen, G. P. (2002). An assessment of maturity from anthropometric measurements. Medicine and Science in Sport & Exercise Science, 34(4), 689–694. https://doi.org/10.1249/00005768-200204000-00020
- Moran, J., Sandercock, G., Rumpf, M. C., & Parry, D. A. (2016). Variation in responses to sprint training in male youth athletes: A meta-analysis. International Journal of Sports Medicine, 38(1), 1–11. https://doi.org/10.1055/s-0042-111439
- Morin, J. B., Edouard, P., & Samozino, P. (2011). Technical ability of force application as a determinant factor of sprint performance. Medicine and Science in Sport & Exercise, 43(9), 1680–1688. https://doi.org/10.1249/MSS.0b013e318216ea37
- Morin, J. B., Jeannin, T., Chevallier, B., & Belli, A. (2006). Spring-mass model characteristics during sprint running: Correlation with performance and fatigue-induced changes. International Journal of Sports Medicine, 27(1), 158–165. https://doi.org/10.1055/s-2005-837569
- Mujika, I., Spencer, M., Santisteban, J., Gioriena, J. J., & Bishop, D. (2009). Age-related differences in repeated-sprint ability in highly trained youth football players. Journal of Sport Sciences, 27(14), 1581–1590. https://doi.org/10.1080/02640410903350281
- Nagahara, R., Haramura, M., Takai, Y., Oliver, J., Wichitaksorn, N., Sommerfield, L., & Cronin, J. (2019). Age-related differences in kinematics and kinetics of sprinting in young females. Scandinavian Journal of Medicine & Science in Sports, 29(6), 800–807. https://doi.org/10.1111/sms.13397
- Nevill, A. M., Bate, S., & Holder, R. L. (2005). Modeling Physiological and Anthropometric Variables Known to Vary with Body Size and Other Confounding Variables. American Journal of Physical Anthropology, 128(S41), 141–153. https://doi.org/10.1002/ajpa.20356
- Papaiakovou, G., Giannakos, A., Michailidis, C., Patikas, D., Bassa, E., Kalopisis, V., Kotzamanidis, C. … Kotzamanidis, C. (2009). The effect of chronological age and gender on the development of sprint performance during childhood and puberty. The Journal of Strength & Conditioning Research, 23(9), 2568–2573. https://doi.org/10.1519/JSC.0b013e3181c0d8ec
- Philippaerts, R. M., Vaeyens, R., Janssens, M., van Renterghem, B., Matthys, D., Craen, R., Malina, R. M., & Malina, R. M. (2006). The relationship between peak height velocity and physical performance in youth soccer players. Journal of Sport Science, 24(3), 221–230. https://doi.org/10.1080/02640410500189371
- Pons, E., Ponce-Bordón, J. C., Díaz-García, J., López Del Campo, R., Resta, R., Peirau, X., & García-Calvo, T. (2021). A longitudinal exploration of match running performance during a football match in the Spanish La Liga: A four-season study. International Journal of Environmental Research and Public Health, 18(3), 1133. https://doi.org/10.3390/ijerph18031133
- Quatman-Yates, C. C., Quatman, C. E., Meszaros, A. J., Paterno, M. V., & Hewett, T. E. (2012). A systematic review of sensorimotor function during adolescence: A developmental stage of increased motor awkwardness? British Journal of Sports Medicine, 46(9), 649–655. https://doi.org/10.1136/bjsm.2010.079616
- Rabita, G., Dorel, S., Slawinski, J., Saez de Villarreal, E., Courtier, A., Samozino, P., & Morin, J. (2015). Sprint mechanics in world class athletes: A new insight into the limits of human locomotion. Scandinavian Journal of Medicine in Sport & Exercise, 26(5), 583–594. https://doi.org/10.1111/sms.12389
- Ratel, S., Duche, P., Hennegrave, A., Van Praagh, E., & Bedu, M. (2002). Acid-base balance during repeated cycling sprints in boys and men. Article. Journal of Applied Physiology, 92(2), 479–485. https://doi.org/10.1152/japplphysiol.00495.2001
- Ratel, S., Duche, P., & Williams, C. (2006). Muscle fatigue during high-intensity exercise in children. Review. Sports Medicine, 36(12), 1031–1065. https://doi.org/10.2165/00007256-200636120-00004
- Ratel, S., Williams, C., Oliver, J., & Armstrong, N. (2006). Effects of age and recovery duration on performance during multiple treadmill sprints. Article. International Journal of Sports Medicine, 27(1), 1–8. https://doi.org/10.1055/s-2005-837501
- Rossi, J., Slotala, R., Samozino, P., Morin, J. B., & Edouard, P. (2017). Sprint acceleration mechanics changes from children to adolescent. Computer Methods in Biomechanics and Biomedical Engineering, 20(sup1), 181–182. https://doi.org/10.1080/10255842.2017.1382922
- Rumpf, M., Cronin, J., Mohamad, I., Mohamad, S., Oliver, J., & Hughes, M. (2015). The effect of resisted sprint training on maximum sprint kinetics and kinematics in youth. European Journal of Sport Science, 15(5), 374–381. https://doi.org/10.1080/17461391.2014.955125
- Rumpf, M., Cronin, J., Oliver, J., & Hughes, M. (2013). Vertical and leg stiffness and stretch shortening cycle changes across maturation during maximal sprint running. Human Movement Science, 32(4), 668–676. https://doi.org/10.1016/j.humov.2013.01.006
- Rumpf, M., Cronin, J., Oliver, J., & Hughes, M. (2015). Kinematics and kinetics of maximum running speed in youth across maturity. Pediatric Exercise Science, 27(2), 277–284. https://doi.org/10.1123/pes.2014-0064
- Runacres, A., Bezodis, N. E., Mackintosh, K. A., & McNarry, M. A. (2019). The reliability of force-velocity-power profiling during over-ground sprinting in children and adolescents. Journal of Sports Sciences, 37(18), 2131–2137. https://doi.org/10.1080/02640414.2019.1622316
- Samozino, P. (2018). A simple method for measuring force velocity and power capabilities and mechanical effectiveness during sprint running. In J. B. Morin (Ed.), Biomechanics of training and testing: Innovative concepts and simple field methods (pp. 237–269). Springer.
- Samozino, P., Rabita, G., Dorel, S., Slawinski, J., Peyrot, N., Saez de Villarreal, E., & Morin, J. B. (2016). A simple method for measuring power, force, velocity properties, and mechanical effectiveness in sprint running. Scandinavian Journal of Medicine & Science in Sports, 26(6), 648–658. https://doi.org/10.1111/sms.12490
- Sanderson, D. (1991). The influence of cadence and power output on the biomechanics of force application during steady state exercise in competitive and recreational cyclists. Journal of Sport Science, 9(2), 191–203. https://doi.org/10.1080/02640419108729880
- Simperingham, K., Cronin, J., Pearson, S., & Ross, A. (2017). Reliability of horizontal force–velocity–power profiling during short sprint-running accelerations using radar technology. Sports Biomechanics, 18(1), 1–12. https://doi.org/10.1080/14763141.2017.1386707
- Simperingham, K., Cronin, J., & Ross, A. (2016). Advances in sprint acceleration profiling for field-based team-sport athletes: Utility, reliability, validity and limitations. Sports Medicine, 46(11), 1619–1645. https://doi.org/10.1007/s40279-016-0508-y
- Slawinski, J., Termoz, N., Rabita, G., Morin, S., Dorel, J., Samozino, P., & Samozino, P. (2017). How 100-m event analyses improve our understanding of world-class men’s and women’s sprint performance. Scandinavian Journal of Medicine & Science in Sports, 27(1), 45–54. https://doi.org/10.1111/sms.12627
- Spencer, M., Bishop, D., & Lawrence, S. (2004). Longitudinal assessment of the effects of field-hockey training on repeated sprint ability. Journal of Science & Medicine in Sport, 7(3), 323–334. https://doi.org/10.1016/S1440-2440(04)80027-6
- Spencer, M., Lawrence, S., Rechichi, C., Bishop, D., Dawson, B., & Goodman, C. (2004). Time–motion analysis of elite field hockey, with special reference to repeated-sprint activity. Journal of Sports Sciences, 22(9), 843–850. https://doi.org/10.1080/02640410410001716715
- Spencer, M., Pyne, D., & Mujika, I. (2011). Fitness determinants of repeated-sprint ability in highly trained youth football players. Journal of Sports Physiology and Performance, 6(4), 497–508. https://doi.org/10.1123/ijspp.6.4.497
- Temfemo, A., Lelard, T., Carling, C., Mandengue, S., Chlif, M., & Ahmaidi, S. (2011). Feasibility and reliability of a repeated sprint test in children age 6 to 8 years. Pediatric Exercise Science, 23(4), 549–559. https://doi.org/10.1123/pes.23.4.549
- Welsman, J., & Armstrong, N. (2019). Interpreting aerobic fitness in youth: The fallacy of ratio scaling. Pediatric Exercise Science, 31(2), 184–190. https://doi.org/10.1123/pes.2018-0141