Abstract
Mid-lake stands of common reed grass (Phragmites australis) have persisted in the shallow Winnebago Pool Lakes of east-central Wisconsin for more than 65 years. Previous research documented that the stands are declining in area with losses up to 94%. Our goal was to characterize the ecological functions of these stands, in part to assess the implications of their loss. To do so, we studied the common reed stands of Lake Poygan during summer, 1999. Common reed stands stabilized the silty/sandy substrate. Wind velocity and wave action were reduced on the downwind sides, and fine, organic sediment accumulated leeward relative to summer prevailing winds. Water clarity was greater on the leeward side. Within stands, dissolved oxygen was less variable, and temperature tended to be cooler. The 18 species of associated aquatic plants covered three or more times the area in the leeward side than windward side. Twenty-nine species of fish were found using the perimeter of the common reed stands. We expect that stand loss will be accompanied by loss of associated wetland vegetation, and the unique combination of habitat attributes afforded by these stands. The current conditions leeward of the stands suggest that stand loss will have a broader spatial effect on ecological characteristics, including fish populations, and thus the implications may be lakewide.
Introduction
The Winnebago Pool Lakes, located in east-central Wisconsin, composed of four lakes – Winnebago, Butte des Morts, Winneconne, and Poygan – drain 16,654 square kilometers and compose 17% of Wisconsin's surface water area. Prior to the construction of two dams at the outlets of Lake Winnebago in the 1850s, the lakes were fertile riverine marshes with dense emergent vegetation, including stands of common reed (Phragmites australis) growing in scattered patches throughout the lakes (Linde Citation1975). Dam modifications gradually increased water levels in the system, raising summer levels by 1 m, and winter levels by 10–30 cm. The marshes were transformed into large, turbid open-water lakes as emergent vegetation, particularly cattails (Typha spp.), decreased, leaving common reed stands offshore in water up to 1.5 m deep (Linde Citation1975; Kahl Citation1993). These uniquely located and inundated mid-water common reed stands have persisted for decades (Gabriel and Bodensteiner Citation2002), despite altered water level regimes with winter drawdowns (Bodensteiner and Gabriel Citation2003), wave action, boating, common carp (Cyprinus carpio) feeding and spawning, and intrusive algal growths (Kahl Citation1993). However, most of the common reed stands have declined based on analysis of aerial photographs dating back to 1937 (Gabriel Citation2001; Gabriel and Bodensteiner Citation2002). The areal coverage of the East Channel stand in Lake Poygan decreased by 94%. Declines in the areas of the other three Lake Poygan stands ranged from 2% to 65%.
The perception of the ecological role and status of common reeds in wetland vegetation communities is decidedly different between Europe and North America. European literature shows great concern for the widespread decline of common reeds, based on documented and perceived benefits to food webs as primary producers and as promoters of habitat structure, water quality, and esthetics (Ostendorp Citation1993; Van der Putten 1997). Despite the apparently hardy nature of common reeds, restoration efforts in Europe have often been unsuccessful, especially under fluctuating water level regimes (Krumscheid et al. Citation1989). Therefore, research and restoration efforts have been directed toward identifying causative factors and preventing further declines (Ostendorp Citation1989). Because of its rapid colonization of disturbed habitats, persistent nature, and tendency to form monospecific stands, common reed in the US is often considered a nuisance, negatively affecting plant biodiversity as well as fish and bird habitat (Benoit and Askins Citation1999; Weinstein and Balletto Citation1999; Ailstock et al. Citation2001). As a result, the North American literature has a preponderance of publications addressing means of eradicating common reed stands (e.g., Marks et al. Citation1994; Ailstock et al. Citation2001) and controlling invasiveness (e.g., Burdick and Konisky Citation2003; Chambers et al. Citation2003; Minchinton and Bertness Citation2003; Silliman and Bertness Citation2004).
By comparison and until losses were noticeable, little effort was directed to understanding the ecological functions of mid-lake common reed stands within the Winnebago Pool Lakes. However, largely due to their unique location, a combination of attributes suggests that common reed may function as a keystone species in these lakes. In addition to typical wetland functions such as furnishing habitat and nutrition for microbial organisms and macroinvertebrates, storing nutrients, and binding substrate and preventing erosion, these stands have additional attributes stemming from their mid-water location. For example, they are free from most terrestrial predators and provide food, cover, and nesting and spawning sites for a number of waterfowl, other birds, and fishes. By reducing the fetch and stabilizing lake bottom sediments, they also prevent erosion and sediment resuspension (Nichols and Vennie Citation1991; Kahl Citation1993).
The goal of this study was to ecologically characterize the structural and functional aspects of common reed stands in the Winnebago Pool Lakes ecosystem. We measured physical and chemical characteristics in and around common reed stands in Lake Poygan. In addition, we determined the spatial relation that common reed stands have to other aquatic plants and fish assemblages that occur in Lake Poygan.
Materials and methods
Study sites
The four common reed stands in Lake Poygan, designated as East Channel, West Bay, Lone Willow, and Hindenburg Line (), encompassed 40 individual patches of common reed, and ranged in size from 6740 m2 (East Channel) to 43,048 m2 (Lone Willow) (Gabriel and Bodensteiner Citation2002). Associated median stem densities ranged between 59 and 76 stems/m2, with maximum stem densities ranging between 117 and 245 stems/m2. Mean summer water depths were 0.81–1.06 m among the stand perimeters, with extremes of 0.10–1.52 m, and mean depths within the stand interiors were 0.56–0.93 m, with extremes of 0.22–1.3 m ().
Figure 1. Locations of the four common reed stands in Lake Poygan, Wisconsin.
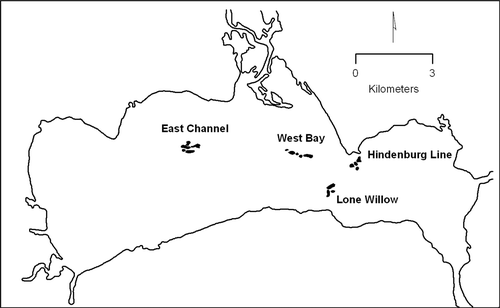
Table 1. Median (and range) stem density (no./m2) and mean (and range) water depths (m) of the four common reed stands in Lake Poygan.
Temperature, dissolved oxygen, pH, and turbidity
Twelve recording thermometers were placed 0.3 m below the surface at the upwind and downwind perimeters (in regard to prevailing wind direction) and in the center of one large patch in each of the four common reed stands. Temperature was recorded at 30 min intervals from mid-July to early September.
Two sampling transects were established in each stand for measurements to be taken 0.5 m below the surface. Transects were located from upwind direction to downwind direction across each common reed stand. Seven locations were established along each transect corresponding to 5 m outside the perimeter, at the perimeter, 25%, 50%, and 75% of the distance through the patch of common reeds, at the opposite perimeter, and 5 m outside the opposite perimeter. Six mid-day measurements as well as at least one dawn measurement of dissolved oxygen (DO) were made in situ with a DO meter at all sites. The pH was measured with a portable meter. Turbidity and Secchi depth were measured 5 m from the stands at the windward and leeward perimeter locations. The sample for turbidity was collected 0.25 m below the surface, and measured using a turbidimeter.
Wind and waves
Wind speeds 1 m above the water surface and wave heights were measured 5 m outside of each stand at the windward and leeward perimeters. Mean elevations of troughs and crests of 15 waves were measured using a portable wave staff. In total, 39 sets of measurements were made in 1- to 3-day periods, spaced over approximately 2-week intervals during a 1-month period, at the seven transect locations.
Substrate
Substrate was sampled at 36 locations, approximately 10 m outside the perimeter of each stand, using a Peterson grab. The particle size distribution by the weight of dried substrate samples was analyzed using standard sieves.
Sediment traps were installed in late July on the windward and leeward perimeters, as well as in the interior, of the four stands. Traps consisted of three adjacent, square plastic containers (10 × 10 × 10 cm3) attached to a single length of wood and positioned flat to the lake bottom. Twenty-one traps were retrieved in late August. Wet sediment depth and particle size distribution were measured for each trap. A homogenized portion of each sample was dried at 100–105°C for 24 h to determine dry weight. The dried sediment samples were then ashed at 550°C to determine percentage of organic matter.
Vegetation
A band with a minimum width of 30 m was systematically surveyed around the perimeter of each common reed patch. Within this area, each kind of emergent, floating-leaved, and submersed vegetation that was encountered was identified. The extent of each vegetation patch was estimated by measuring the distance along the longest axis and the distance perpendicular to the longest axis at the midpoint and then calculating the equivalent elliptical area. Species composition between windward and leeward locations relative to the predominant wind direction was compared using the Jaccard similarity index and coefficient of community index to compare area values (Mueller-Dumbois and Ellenberg Citation1974).
Fish
Fish were collected from each of the four stands using fyke nets and boat electrofishing in late July 1999. Four Indiana-style fyke nets (1.2 × 2.3 m2 frames, 15 m lead, 12 mm bar mesh) were set for approximately 24 h on two consecutive days in each of two common reed stands. Half of the nets were set on the upwind side and half on the downwind side. Captured fish were identified, measured (total length), and released.
In addition, 80 min of boat-electrofishing effort was exerted on each stand. Half of this effort occurred during the day and the other half after sunset. The daytime and nighttime runs were repeated in the same locations, and locations were selected to represent the habitat diversity within each stand. Captured fish, except common carp, were identified, measured (total length), and released. Carp were tallied but not captured.
Results
Water quality
Daily mean interior temperatures was cooler relative to both the windward and leeward edges in the two densest and largest stands, Lone Willow and West Bay (paired t-test, p < 0.05) (). However, interior temperature was only cooler than the leeward edge in the East Channel stand, which has generally deeper water. Conversely, interior temperature in the Hindenburg Line stand was only cooler than the windward, west-facing edge. Daily mean temperatures differed between the windward and leeward edges of three of the stands; the west-facing, windward edge was cooler at East Channel and warmer at Hindenburg Line and Lone Willow. Diel fluctuations in temperature differed between the interior and one of the edges in three of the stands; fluctuations were less than at the windward edge of East Channel and more than at the leeward edges of Hindenburg Line and West Bay, which are east- and north-facing, respectively. Diel leeward temperature fluctuations at West Bay were lower than at the windward edge, reduced along the latter, north-facing edge.
Table 2. Daily mean water temperatures and temperature ranges (°C) at windward (W), interior (I), and leeward (L) locations of Lake Poygan common reed stands.
During daylight hours, mean relative DO concentration characteristically increased through the day from a morning low of 89% of saturation up to nearly 132% during the typically sunny afternoons. Based on measurements taken along transects from windward to leeward, there was a trend of decreasing supersaturation (); DO saturation was higher at windward locations than either interior or leeward locations on the same transect (paired t-test, p < 0.05).
Table 3. Water quality measures at windward, interior, and leeward locations of Lake Poygan common reed stands.
Secchi depth did not differ between the windward and leeward edges of the common reed stands (paired t-test, p > 0.05) (). Turbidity near the surface also was unchanged from windward edge to leeward edge (Wilcoxon t-test, p > 0.05).
The pH ranged from near 8 to as high as 9.5, and there were no significant trends from the windward side to leeward side of the common reed stands (paired t-test, p > 0.05) ().
Wind and waves
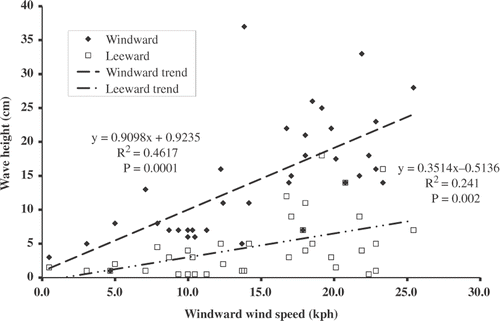
The mean wind velocity of 11.1 kph on the leeward sides of the stands was 4.1 kph less than the mean windward velocity (paired t-test, p < 0.05) (). Accordingly, mean wave height was also reduced from 14.3 to 4.6 cm (paired t-test, p < 0.05) (). Wind speed reduction from windward side to leeward side varied considerably across sites and dates, with a mean reduction of approximately 80% across all wind speeds. Likewise, difference in prevailing wave height from windward side to leeward side varied but was reduced on the leeward side by a mean of 44% across all wind speeds.
Figure 2. Reduction in wind speed from the windward side to the leeward side in relation to wind speed on the windward side of the four common reed stands.
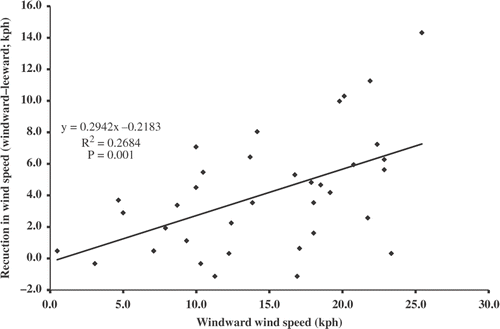
Figure 3. Reduction in wave height from the windward side to leeward side of the four common reed stands in relation to windward wind speed.
Substrate and sedimentation
A higher proportion of sand was found at windward than leeward locations (Mann–Whitney U test, p < 0.05) (). Conversely, higher proportions of very fine sand and silt were found at the leeward locations. Proportion of organic matter in the substrate did not differ between windward and leeward locations (Mann–Whitney U test, p > 0.05).
Table 4. Substrate characteristics windward and leeward of common reed stands in Lake Poygan based on summer prevailing wind direction.
Although prevailing leeward sides of the common reed stands had over twice the median depositional rates of sediment as those of windward locations, this was not statistically significant (, Mann–Whitney U test, p > 0.05). A small sample size resulted from lost equipment and an inherently high variation in sedimentation accumulation rates was noted among samples; both these factors would contribute to a lack of statistical discrimination. However, the proportion of organic matter in the accumulated sediment in the samplers was higher at the leeward locations (Mann–Whitney U test, p < 0.05).
Vegetation
In the shallow waters surrounding the common reed stands, 18 additional plant species were identified, dominated by spatterdock (Nuphar variegata) and hardstem bulrush (Scirpus acutus) (). The areal extent and diversity of plant species associated with common reed stands differed between windward and leeward locations, resulting in a Jaccard similarity coefficient of 77.8% and a coefficient of community of 23.2% for all the common reed stands. Total area covered by associated plant was at least three times greater on the leeward side than on the windward side. Associated plant species found on the windward side tended to be dominated by hardstem bulrush and river bulrush (Scirpus fluviatilis), as well as submersed plants such as wild celery (Vallisneria americana) and sago pondweed (Potamogeton pectinatus). A greater diversity of plant species was located on the leeward side and dominated by floating-leaved plants such as spatterdock and white water lily (Nymphaea odorata), as well as a variety of emergents dominated by hardstem bulrush, river bulrush, and stiff arrowhead (Sagittaria rigida).
Table 5. Vegetation associated with the windward and leeward sides of Lake Poygan common reed stands.
Differences in species composition between windward and leeward locations also varied by common reed stand (). Variation in species composition was greatest in the three larger stands, with Jaccard similarity coefficients ranging between 50% and 75%, and coefficients of community ranging between 17% and 18.8%. In contrast, the entire East Channel stand was characterized by plant species associated with windward areas, resulting in a Jaccard similarity coefficient of 100% and a coefficient of community of 43.7%.
Table 6. Similarity of vegetation between windward and leeward locations of common reed stands in Lake Poygan.
Fish
Totally, 1953 fish comprising 29 species were captured along the edges of the four common reed stands. Among most sport fishes captured, the mean sizes tended to occur in the range often retained by anglers. Potential forage species appeared to be of sizes that would render them susceptible to predation by most of the individuals of the piscivorous species that were captured ().
Table 7. Fish collected at each common reed stand in Lake Poygan.
Discussion
The mid-water common reed stands of Lake Poygan alter the mid-lake environment in a manner that facilitates the existence of a variety of plant species and also provides habitat for diverse fish species. Habitat characteristics are distinctly different downwind of common reed stands from those exposed to prevailing summer winds. These differences appear to be attributable to the protection afforded by the stands and cause different physical, chemical, and biological characteristics.
The greater vegetated area on the leeward sides, of emergent, floating-leaved, and submersed macrophytes appears to be more attributable to wind and wave activity than to water clarity, since water clarity did not differ between leeward and windward locations. Areal coverage of macrophytes was more than seven times greater on the leeward side, but proportional coverage varied by growth form and individual species. The eleven-fold increase in area of emergents and floating-leaved plants, which would be more subject to wave action, from windward to leeward versus the corresponding 1.5-fold increase in submersed plants points to inhibition of plants with leaves at or above the water surface. Further, relative areas of individual submersed species differed less than those of emergents and floating-leaved plants. While relative areas of individual submersed species were within 5% of the total areas of windward and leeward vegetations, emergents and floating-leaved plants differed by as much as 26% for spatterdock (higher proportion leeward), and 24% for river bulrush (higher proportion windward). Thus, the leeward side favored a greater area of vegetation and also was more conducive to growth of some emergent and floating-leaved species.
Other studies have found greater accumulation rates of fine mineral and organic sediments associated with coastal marshes dominated by common reed (Takeda and Kurihara Citation1988; Rooth and Stevenson Citation2000). We found substrate differed in size composition, with coarser substrate on the windward sides, which was likely a result of windward wave-induced erosion and leeward deposition of finer particles. Although these substrate differences could have caused differences in vegetation (Weisner 1987; Coops et al. Citation1994), the fact that the difference in vegetated area between windward and leeward side was distinguished by growth form more than by individual species suggests otherwise.
A major water quality problem in the Pool Lakes is their hypereutrophic state, characterized by phytoplankton blooms and high turbidity in the summer, a situation due to external nutrient loading via runoff from agricultural and urban areas (Sloey and Spangler Citation1977; Lillie and Mason Citation1983) and probably exacerbated by internal loading when phosphorus bound to sediment particles is suspended from the substrate by wave action. Farnsworth and Meyerson (Citation2003) found that common reed takes up nutrients more efficiently than other species such as broadleaf cattail, due in part to earlier leaf emergence and longer leaf longevity, greater shoot length and biomass per ramet, as well as higher levels of standing leaf area, total leaf area per plant, and total foliar chlorophyll. Because of these characteristics, common reeds function as important sinks for nutrients such as nitrogen. The reduced fetch, downwind protection, and the substrate stabilization effected by common reed stands may also counteract the internal loading of phosphorus.
The disparity of conclusions about the effects of common reed on fish habitat indicates that the location of the stand affects its role. For example, Ostendorp (Citation1993) maintained that the structure of lakeshore common reed stands is beneficial to fish, while Weinstein and Balletto (Citation1999) contended that the invasion of common reed in coastal marshes negatively affects fish habitat by changing features such as the hydroperiod, drainage densities, and flows and exchanges of organic materials. By comparison, the abundance and diversity of aquatic vegetation that we found associated with mid-lake common reed stands and the variety of growth forms and resources that they provide are conducive to the presence of several kinds and age classes of fishes. Many of the fishes that we collected were large individuals and many were predators. Coupled with the fact that large numbers of emerald shiners and smaller gizzard shad were also collected in these areas, this strongly suggests that common reed habitat is being used for foraging by larger, piscivorous fishes, including some of the most valued recreational species such as walleye, channel catfish, and yellow perch. The presence of smaller individuals (e.g., sunfish family members including pumpkinseed, bluegill, black crappie, and largemouth bass) suggests that young fish are also using this area for shelter, as the three-dimensional structure supplied by the common reed stems likely provides both protection from larger piscivorous fish and substrate for macroinvertebrate prey (Ostendorp Citation1993).
DO is one factor that may limit the suitability of this habitat for fish, but the permeable structure of the common reed stems and the mid-lake location appear to mitigate the occurrence of hypoxic conditions that have been found associated with vegetated shoreline aquatic habitats, perhaps by allowing greater water circulation. In nearshore habitats, we found DO concentrations at dawn of less than 1 mg/L and less than 10% of saturation at corresponding times of the year (Gabriel and Bodensteiner Citation2000). In areas heavily vegetated with aquatic macrophytes, DO concentrations of 0.1 mg/L were recorded. The cooler water temperatures found in the interior or leeward of some of the common reed stands also help alleviate the occurrence of hypoxia, as oxygen is more soluble in cooler water and fish metabolism is lower.
Primarily due to their unique, mid-lake location, common reed stands in Lake Poygan provide a number of ecological benefits, in sharp contrast to the majority of studies in North America, which document negative impacts and emphasize the invasiveness of common reed. We expect that losses of common reed in Lake Poygan, and possibly the other Winnebago Pool Lakes, will be accompanied by loss of associated vegetation and the unique combination of habitat attributes afforded by these stands. The different ecological conditions found windward and leeward of the stands suggest that continued stand losses may have a broader spatial effect on ecosystem characteristics with lake-wide implications for dozens of species of plants and animals.
Acknowledgements
This study was funded by a Wisconsin Department of Natural Resources Lakes Protection Grant, the Lake Poygan Sportsman's Club, and a Faculty Development Grant from the University of Wisconsin-Oshkosh. We thank D. Rudebeck, A. Techlow, M. Sesing, K. Kamke, M. Schrage, R. Bahnfleth, C. Lancaster, M. McCaffrey, B. Rissky, and K. Townsend.
Related Research Data
References
- Ailstock , SM , Norman , MC and Bushmann , JP . 2001 . Common reed (Phragmites australis): control and effects upon biodiversity in freshwater nontidal wetlands . Restoration Ecology , 9 : 49 – 59 .
- Benoit , LK and Askins , RA . 1999 . Impact of the spread of Phragmites on the distribution of birds in Connecticut tidal marshes . Wetlands , 19 : 194 – 208 .
- Bodensteiner , LR and Gabriel , AO . 2003 . Response of mid-water common reed stands to physical environmental conditions in Lake Poygan, Wisconsin . Aquatic Botany , 76 : 49 – 64 .
- Burdick , DM and Konisky , RA . 2003 . Determinants of expansion for Phragmites australis, common reed, in natural and impacted coastal marshes . Estuaries , 26 : 407 – 416 .
- Chambers , RH , Osgood , DT , Bart , DJ and Montalto , F . 2003 . Phragmites australis invasion and expansion in tidal wetlands: interactions among salinity, sulfide and hydrology . Estuaries , 26 : 398 – 406 .
- Coops , H , Geilen , N and van der Velde , G . 1994 . Distribution and growth of the helophyte species Phragmites australis and Scirpus lacustris in water depth gradients in relation to wave exposure . Aquatic Botany , 48 : 273 – 284 .
- Farnsworth , EJ and Meyerson , LA . 2003 . Comparative ecophysiology of four wetland plant species along a continuum of invasiveness . Wetlands , 23 : 750 – 762 .
- Gabriel , AO . 2001. Assessment of stress-response in common reed stands in the Winnebago Pool Lakes, Wisconsin. Wisconsin Department of Natural Resources Lake Planning Grant Report. Oshkosh (WI), USA
- Gabriel , AO and Bodensteiner , L . 2000. Restoration and ecosystem functions of common reed stands in Lake Poygan, Wisconsin. Wisconsin Department of Natural Resources Lake Protection Grant Report. Oshkosh (WI), USA
- Gabriel , AO and Bodensteiner , LR . 2002 . Historical changes in mid-water stands of common reed in the Winnebago Pool Lakes, Wisconsin . Journal of Freshwater Ecology , 17 : 563 – 573 .
- Kahl , R . 1993. Aquatic macrophyte ecology in the Upper Winnebago Pool Lakes, Wisconsin. Wisconsin Department of Natural Resources Technical Bulletin 182. Madison (WI), USA
- Krumscheid , P , Stark , H and Peintinger , M . 1989 . Decline of reed at Lake Constance (Obersee) since (1967) based on interpretations of aerial photographs . Aquatic Botany , 35 : 57 – 62 .
- Lillie , RA and Mason , JW . 1983. Limnological characteristics of Wisconsin lakes. Department of Natural Resources Technical Bulletin No. 138. Madison (WI), USA
- Linde , A . 1975. Report on preliminary Fox and Wolf River Lakes vegetation studies. Fox-Wolf Lakes Habitat Deterioration Exploratory Studies. Wisconsin Department of Natural Resources, Oshkosh (WI), USA
- Marks , M , Lapin , B and Randall , J . 1994 . Phragmites australis (P. communis): threats, management, and monitoring . Natural Areas Journal , 14 : 285 – 294 .
- Minchinton , TE and Bertness , MD . 2003 . Disturbance-mediated competition and the spread of Phragmites australis in a coastal marsh . Ecological Applications , 13 : 1400 – 1416 .
- Mueller-Dumbois , D and Ellenberg , H . 1974 . Aims and methods of vegetation ecology , New York : John Wiley and Sons .
- Nichols , SA and Vennie , JG . 1991. Attributes of Wisconsin lake plants. Wisconsin Geological and Natural History Survey Information Circular 73. Madison (WI), USA
- Ostendorp , W . 1989 . Die-back of reeds in Europe – a critical review of literature . Aquatic Botany , 35 : 5 – 26 .
- Ostendorp , W . 1993 . “ Reed bed characteristics and significance of reed in landscape ecology ” . In Lakeshore deterioration and restoration works in Central Europe , Edited by: Ostedorp , W and Krumsheid-Plankert , P . 149 – 161 . Stuttgart, Germany : Fischer-Verlag .
- Rooth , JE and Stevenson , JC . 2000 . Sediment deposition patterns in Phragmites australis communities: implications for coastal areas threatened by rising sea-level . Wetlands Ecology and Management , 8 : 173 – 183 .
- Silliman , BR and Bertness , MD . 2004 . Shoreline development drives invasion of Phragmites australis and the loss of plant diversity on New England salt marshes . Conservation Biology , 18 : 1424 – 1434 .
- Sloey , WE and Spangler , FL . 1977. The trophic status of the Winnebago Pool Lakes. Fox Valley Water Quality Planning Agency Report, Neenah (WI), USA
- Takeda , S and Kurihara , Y . 1988 . The effects of the reed, Phragmites australis (Trin.), on substratum grain-size distribution in a salt marsh . Journal of the Oceanographical Society of Japan , 44 : 103 – 112 .
- Van der Putten , WH . 1997 . Die-back of Phragmites australis in European wetlands: an overview of the European Research Programme on reed die-back and progression (1993-1994) . Aquatic Botany , 59 : 263 – 275 .
- Weinstein , MP and Balletto , JH . 1999 . Does the common reed, Phragmites australis, affect essential fish habitat? . Estuaries , 22 : 793 – 802 .
- Weisner , SEB . 1987 . The relation between wave exposure and distribution of emergent vegetation in a eutrophic lake . Freshwater Biology , 18 : 537 – 544 .