Abstract
We studied the population dynamics of Brachionus angularis during summer and autumn in 2008 in subtropical, eutrophic Lake Jinghu, China, and the relationships between rotifer population parameters and water temperature, pH, dissolved oxygen, transparency, chlorophyll a concentration, and potential competitors and predators with and without a time lag of 4 days. Brachionus angularis appeared on 24 July and disappeared on 21 October. No significant correlations were found between the concentration of chlorophyll a and population parameters, but B. angularis density was positive correlated with the density of other microfilter-feeder rotifers, indicating that the B. angularis population in Lake Jinghu was not food-limited during the study period. Since there were no correlations between the density of omnivorous rotifers (e.g., Asplanchna spp.) and population parameters of B. angularis and since very few cladocerans and copepods were present, predator pressure on B. angularis did not exist. Among all the abiotic factors involved in this study, only temperature was correlated positively with the population density and the birth rate of B. angularis and had a possible effect on the proportion of mictic females in the population at the end of study.
Introduction
It is well known that predation (top–down forces) and resource supply (bottom–up forces) are important in the regulation of population dynamics and community structure in freshwater plankton (Sommer Citation1989). Many previous studies have found that individual numbers or biomasses of organisms in aquatic systems are sensitive to changes in both resource supply and abundance of their natural enemies (Carpenter et al. Citation1987; Power Citation1990; Špoljar et al. Citation2005). However, Hunter and Price (Citation1992) argued that ecologists should seek to identify the specific conditions determining the relative importance of these forces. Notably, both the analyses of long-term data and mesocosm experiments suggest that temperature and not food is the dominant factor driving interannual variability of Daphnia population dynamics during spring (Straile Citation2000; Schalau et al. Citation2008).
Rotifers are one of the most important constituents of freshwater zooplankton communities. Regulation of the seasonal variation of rotifer populations has been attributed to both abiotic factors, including temperature, pH, dissolved oxygen (DO), etc., and biotic factors such as food resources, competition, and predation (Dumont Citation1977; Hofmann Citation1977). Temperature was the most important factor that influenced the different parameters in a rotifer experimental population (Xi and Huang Citation2004), and food resource has been receiving much attention as a driving force regulating seasonal variation of rotifer natural populations (Urabe Citation1992; Merriman and Kirk Citation2000; López et al. Citation2007). In addition, rotifers can be effectively depressed by crustacean zooplankton and omnivorous rotifers (e.g., Asplanchna spp.) through exploitative competition and predation (Henssen and Nilssen Citation1985; Pace et al. Citation1998; Brandl Citation2005).
The eurythermal and cosmopolitan rotifer B. angularis inhabits a variety of freshwater ecosystems and is widely used in aquaculture and aquatic ecotoxicology (Gama-Flores et al. Citation2004; Hu and Xi Citation2006a, Citation2006b). Although a large number of studies have focused on the effects of various environmental factors on experimental population dynamics (Walz Citation1987; Gama-Flores et al. Citation2004; Hu and Xi Citation2006a, Citation2006b), little is known about the ecological mechanisms of natural population dynamics of B. angularis. In subtropical, shallow, eutrophic Lake Jinghu, China, B. angularis is one of the common planktonic rotifers and has a maximal density peak in September, which declines rapidly from the beginning of October (Wen et al. Citation2006). In this study, our aims were to (1) investigate the population dynamics of B. angularis during summer and autumn, and analyze the relationships between abiotic and biotic factors and population parameters in Lake Jinghu and (2) identify key factor(s) regulating the temporal variation of the B. angularis population.
Materials and methods
Lake Jinghu is a lake of 0.15 km2 located near the middle reach of the Yangtze River and is comprised of two sections named the small lake region and the big lake region (Wen et al. Citation2006; Qian et al. Citation2007). The planktonic rotifers were sampled at two fixed stations at about 4-day intervals during the summer and autumn in 2008. Stations I and II were situated in the centers of the small and big regions of the lake, respectively. Water depth at the two stations usually fluctuated from 1.0 to 1.5 m.
On each sampling date, a quantitative sample for estimating rotifer density was obtained from each station by filtering (25 µm mesh net) a 15 L sample of the integrated lake water (5 L of water from surface to bottom at 0.5 m intervals). The retained rotifers were preserved in 4% sucrose formaldehyde (Haney and Hall Citation1973) and were concentrated to 30 mL, and 10% of sample was examined under 100 × magnification (López et al. Citation2007). Qualitative samples for identification of rotifer species were collected by filtering 40 L of lake water through a 64 µm nylon sieve. Rotifer densities were determined for discrete species, and species identification was carried out on living materials according to Koste (1978).
The egg ratio of B. angularis was recorded and included attached and free eggs; resting eggs were also recorded and counted according to their morphology, as described by Lin et al. (Citation1997). Both the possible dislodgement of eggs from females and loss of eggs during the filtering, as reported by Orcutt and Pace (Citation1984) and Likens and Gilbert (Citation1970), were not checked. The proportion of mictic females in the B. angularis population was estimated according to Xi and Huang (Citation2000). The instantaneous birth rate (b, day−1), the instantaneous population growth rate (r, day−1), and the instantaneous death rate (d, day−1) were calculated with the following formulae: b = ln (E + 1)/D, where E is the number of amictic eggs per female, and D the egg development time in days determined by Baker (Citation1979); r = ln N t − ln N t −1/t, where N t and N t −1 are population densities at times t and t−1, respectively; and, d = b−r (Edmonson Citation1965).
Simultaneously with the rotifer sampling, physicochemical conditions in each area were measured. Surface water temperature was measured using a mercury thermometer, and water transparency as well as water depth were determined using Secchi disk. DO was determined with a HI-9143 DO meter (HANNA, Italy). For measuring chlorophyll a (chl a) concentration, 1.0 L of water of another 15 L integrated lake water was sampled in two fractions ‐ one filtered through a 25 µm plankton net and one unfiltered. Chl a content was measured using a spectrophotometer and the standard acetone extraction method (Huang Citation1999).
Results
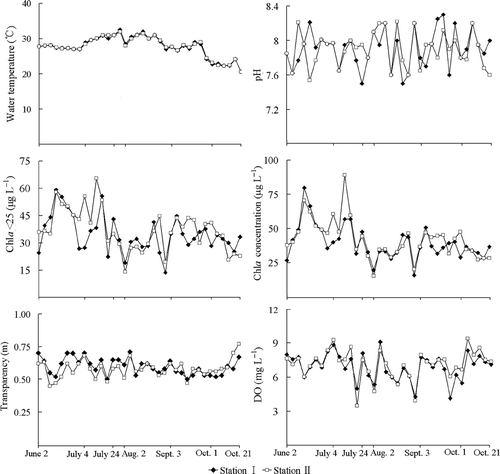
Between 24 July and 21 October 2008 when B. angularis was recorded, the water temperature ranged from 20.5°C to 32.5°C. There were little differences in the water temperature, transparency, and DO content between the two stations. The difference in temperature between the surface and the bottom layers of the water column was less than 0.5°C. The pH of the water at both stations fluctuated from 7.5 to 8.5. The concentrations of chl a in water samples unfiltered (chl a) and filtered (chl a < 25 µm) were high, with average values at both stations exceeding 30 µg L−1, and varied considerably but with no discernable patterns between the two stations or within each station (). The concentrations of nutrients including total nitrogen and total phosphorus were not involved in this study because they showed small variation between summer and autumn in Lake Jinghu (Qian et al. Citation2007).
Figure 1. Physicochemical conditions at the two stations of Lake Jinghu between July 24 and October 21 in 2008.
The rotifer B. angularis appeared on 24 July and disappeared on 21 October, and its population density varied between 1.0 ind. L−1 and 72.0 ind. L−1 at station I and ranged from 1.0 ind. L−1 to 100.0 ind. L−1 at station II. Generally, the population density was highest during the period from 28 July to 7 September and then decreased at both stations except the peak on 1 October at station I (). When the density of B. angularis declined in October, individuals carrying resting eggs were observed at both stations, and the proportion of mictic females (Y,%) was correlated with water temperature (X,°C) as follows: station I: Y 1 = −0.309X 1 + 7.29 (R 2 = 0.50, P = 0.071); station II: Y 2 = 0.284X 2 − 6.16 (R 2 = 0.65, P = 0.053).
Figure 2. Dynamics of population density, egg ratio, death rate, birth rate, and growth rate of B. angularis at the two stations of Lake Jinghu between 24 July and 21 October 2008.
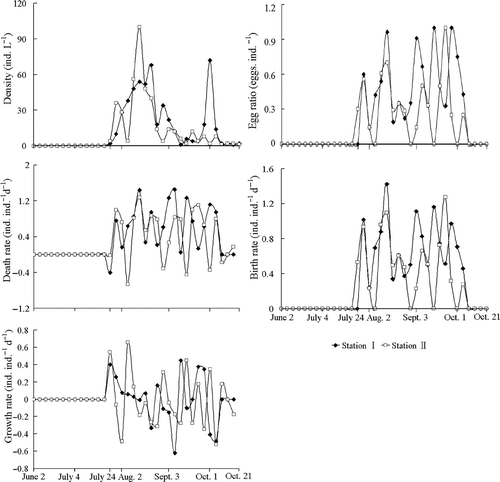
The density of B. angularis at station II was positively correlated with both the egg ratio and the birth rate and negatively associated with the death rate without a time lag, but it presented a positive relationship with the population growth rate with a time lag. No significant correlations were found between the population density and the egg ratio, the birth rate, the death rate, or the population growth rate at station I both with or without a time lag ().
Table 1. Product–moment correlations between population density and egg ratio (e), population growth rate (r), birth rate (b), and death rate (d) of B. angularis with and without a time lag of one sampling interval (four days).
The density of B. angularis at station II showed positive relationship with temperature and with the densities of the other microfilter-feeder rotifers, including Anuiaeopsis fissa, Filinia longiseta, Brachionus forficula, Brachionus diversicornis, and Keratella cochlearis observed during the study period both with and without a time lag, while at station I the density of B. angularis was positively correlated with both the density of the other microfilter-feeder rotifers without a time lag and temperature with a time lag ().
Table 2. Correlation coefficients between population density and environmental variables.
Among the eight abiotic and biotic factors involved in this study, temperature alone was correlated positively with birth rate at station I and with birth rate and egg ratio at station II without a time lag.
Discussion
Monogonont rotifers reproduce by cyclical, ameiotic parthenogenesis, resulting in high production rates and population densities in favorable conditions such as appropriate temperature, food resource, and little prey pressure. However, unfavorable conditions decrease the population densities of rotifers and induce the production of non-planktonic resting eggs which are critical for rotifer survival from one time to the next period (Andrew and Fitzsimons Citation1992). A negative death rate of natural rotifer populations is often apparent, which partly indicates that the source of a rotifer population is most likely from hatching of resting eggs in the sediments rather than ameiotic parthenogenesis (Andrew and Fitzsimons Citation1992). The numbers of hatching individuals (8.3 ind./200 mL sediment) of B. angularis from resting eggs in the sediments of Lake Jinghu at the inducing temperature of 30°C were much greater than that at 10°C and 20°C (1.0 ind./200 mL sediment) (Zhang et al. Citation2008). In this study, the water temperature of 31°C and the negative death rates on 24 July when B. angularis began to occur at the two stations suggest that the source of this rotifer might be from the hatching of resting eggs in the sediments.
Algae, heterotrophic nanoflagellates, bacteria, and picoplankton are major food resources for rotifers and are generally abundant in nutrient-rich environments (Hwang and Heath Citation1999; Yoshida et al. Citation2003; Auer et al. Citation2004). Chl a may be a representative of edible algae or other food resources accompanied with it (Auer et al. Citation2004), and bacterial productivity may be positively correlated with nutrient loading and chl a concentration (Hwang and Heath Citation1997). Together with B. angularis, the other five rotifer species observed during the study period (A. fissa, F. longiseta, B. forficula, B. diversicornis, and K. cochlearis) could be regarded as microfilter-feeder rotifers based on their food-collecting mechanisms and the size of consuming food particles (particles ≤ 20 µm; Pourriot Citation1977; Špoljar et al. Citation2005; Virro et al. Citation2009). Overall, in eutrophic Lake Jinghu with high chl a concentration, no correlations were found between the concentrations of two fractions of chl a and population parameters including population density, egg ratio, birth rate, death rate, and growth rate, indicating that the B. angularis population in Lake Jinghu was not food-limited during the study period. Besides, the positive correlation of the population density of B. angularis with the density of all the other microfilter-feeder rotifers also confirmed this assumption.
Inverse relations between population density and egg ratio, population growth rate, and birth rate and direct relations between population density and death rate suggest intraspecific competitive interactions (DeMott Citation1983; Ooms-Wilms et al. Citation1999; Sarma Citation2005). Density-dependent population growth seems plausible for a rotifer population with density exceeding 500 ind. L−1 (Ooms-Wilms et al. Citation1999). In contrast, Snell et al. (Citation2001) found that a density effect can exist even when the natural population density of B. angularis was lower than 10 ind. L−1. In this study, the average density of B. angularis was lower than 500 ind. L−1 but higher than 10 ind. L−1. In addition, significant negative correlations between population density and egg ratio, growth rate, and birth rate were lacking. Therefore, if intraspecific competition occurred within the B. angularis population in Lake Jinghu, it remains uncertain.
Rotifers suffer predation by copepods, planktiorous fishes, and macroinvertebrates such as Chaoborus spp., and competition (both interference and exploitative) from cladocerans (Williamson Citation1983; Williamson and Butler Citation1986; Gilbert Citation1988; Yoshida et al. Citation2003). Thus, rotifer abundance can be reduced when mesozooplankton is abundant. Chaoborus spp. were not recorded in this study because rotifer samples were collected in nearly mid-day when this invertebrate predator had likely migrated to lower strata (Havens et al. Citation2007), and fishes were not considered as important predators to rotifers because rotifers are too small to be grazed by most open-water planktivores although they may comprise an important part of the diet of larval fish (Bartsch et al. Citation2004). In Lake Jinghu, predator pressure was excluded because very few cladocerans and copepods were observed in this study and there were no correlations between the density of the omnivorous rotifers Asplanchna spp. and population parameters of B. angularis.
Temperature is not only playing a key role in restricting the occurrence and influencing seasonal succession of rotifer species (Herzig Citation1987) but also has a great effect on embryonic and post-embryonic developments of rotifers (Herzig Citation1983). The population of B. angularis approaches its thermal tolerance limits at 25°C (Walz et al. Citation1989) and the Wuhu strain cannot survive at temperatures exceeding 30°C (Hu et al. Citation2006b). Brachionus angularis reaches maximum fertility at about 18°C (Radwan Citation1980) and obtains its highest gross and net reproductive rates, growth, and birth rates at 20°C, but the shortest embryonic development time at 25°C (Walz Citation1987). In this study, water temperature was correlated positively with population density and birth rate of B. angularis in the two areas of Lake Jinghu, and the population growth rate at 26°C in laboratory was significantly higher than that at 30°C (unpublished data), implying that temperature limitation might be possible. Clearly, the unfavorable high temperature in summer might restrain the population growth of B. angularis in Lake Jinghu, while the rapid population decline in autumn might be attributed to the production of resting eggs induced by unstable water temperature during this season.
Acknowledgements
This Indent study was supported by the Natural Science Foundation of China (30770352), Foundation of PhD Program of Ministry of Education of the Peoples’ Republic of China (20093424110002), Foundation of Key Laboratory for the Conservation and Utilization of Important Biological Resources in Anhui province, Foundation of Key Laboratory of Aquatic Eutrophication and Control of Harmful Algal Blooms of Guangdong Higher Education Institutes, the Foundation for Young Talents in College of Anhui Province (2009SQRZ029), and the Grant for Youth of Anhui Normal University (2007xqn74).
Related Research Data
References
- Andrew , TE and Fitzsimons , AG . 1992 . Seasonality, population dynamics and production of planktonic rotifers in Lough Neagh, Northern Ireland . Hydrobiologia , 246 : 147 – 164 .
- Arndt , H . 1993 . Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliate) – a review . Hydrobiologia , 255/256 : 231 – 246 .
- Auer , B , Elzer , U and Arndt , H . 2004 . Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: influence of resource and predation . Journal of Plankton Research , 26 : 397 – 709 .
- Baker , RL . 1979 . Birth rate of planktonic rotifers in relation to food concentration in a shallow eutrophic lake in western Canada . Canadian Journal of Zoology , 57 : 1206 – 1214 .
- Bartsch , LA , Richardson , WB and Sandheinrich , MB . 2004 . Zebra mussels (Driessena polymorpha) limit food for larval fish (Pimephales promelas) in turbulent systems: a bioenergetics analysis . Hydrobiologia , 495 : 59 – 72 .
- Brandl , Z . 2005 . Freshwater copepods and rotifers: predators and prey . Hydrobiologia , 546 : 475 – 489 .
- Carpenter , SR , Kitchell , JF , Hodgson , JR , Cochran , PA , Elser , JJ , Elser , MM , Lodge , DM , Kretchmer , D , He , X and Vonende , CN . 1987 . Regulation of lake primary productivity by food web structure . Ecology , 68 : 1863 – 1876 .
- DeMott , WR . 1983 . Seasonal succession in a natural Daphnia assemblage . Ecological Monographs , 53 : 321 – 340 .
- Dumont , HJ . 1977 . Biotic factors in the population dynamics of rotifers . Archiv fur Hydrobiologie , 8 : 98 – 122 .
- Edmonson , WT . 1965 . Reproductive rates of planktonic rotifers as related to food and temperature in nature . Ecological Monographs , 35 : 61 – 111 .
- Gama-Flores , JL , Sarma , SSS and Nandini , S . 2004 . Acute and chronic toxicity of the pesticide methyl parathion to the rotifer Brachionus angularis (Rotifera) at different algal (Chlorella vulgaris) food densities . Aquatic Ecology , 38 : 27 – 36 .
- Gilbert , JJ . 1988 . Suppression of rotifer populations by Daphnia: a review of the evidence, the mechanisms, and the effects on zooplankton community structure . Limnology and Oceanography , 33 : 1286 – 1303 .
- Haney , J and Hall , D . 1973 . Sugar-coated Daphnia: a preservation technique for Cladocera . Limnology and Oceanography , 18 : 331 – 333 .
- Havens , KE , East , TL and John , RB . 2007 . Zooplankton response to extreme drought in a large subtropical lake . Hydrobiologia , 589 : 187 – 198 .
- Henssen , DO and Nilssen , JP . 1985 . Factors controlling rotifer abundances in a Norwegian eutrophic lake: an experimental study . Annales de limnologie , 21 : 97 – 105 .
- Herzig , A . 1983 . Comparative studies on the relationship between temperature and duration of embryonic development of rotifers . Hydrobiologia , 104 : 237 – 246 .
- Herzig , A . 1987 . The analysis of planktonic rotifer populations: plea for long-term investigations . Hydrobiologia , 14 : 163 – 180 .
- Hofmann , W . 1977 . The influence of abiotic environmental factors on population dynamics in planktonic rotifers . Archiv fur Hydrobiologie , 8 : 77 – 83 .
- Hu , H-Y and Xi , Y-L . 2006a . Differences in population growth and morphometric characteristics of three strains of Brachionus angularis . Journal of Freshwater Ecology , 21 : 101 – 108 .
- Hu , H-Y , Xi , Y-L and Geng , H . 2006b . Effects of temperature on life history strategies of three strains of Brachionus angularis . Acta Hydrobiologica Sinica , 28 : 284 – 288 . (in Chinese with English abstract)
- Huang , X-F . 1999 . Survey, observation and analysis of lake ecology , Beijing : Chinese standard press .
- Hunter , MD and Price , PW . 1992 . Playing chutes and ladders: heterogeneity and the relative roles of bottom-up and top-down forces in natural communities . Ecology , 73 : 724 – 732 .
- Hwang , SJ and Heath , TR . 1997 . Bacterial productivity and protistan bacterivory in coastal and offshore communities of Lake Erie . Canadian Journal of Fisheries and Aquatic Sciences , 54 : 788 – 799 .
- Hwang , SJ and Heath , TR . 1999 . Zooplankton bacterivory at coastal and offshore sites of Lake Erie . Journal of Plankton Research , 21 : 699 – 719 .
- Likens , GE and Gilbert , JJ . 1970 . Notes on quantitative sampling of natural populations of planktonic rotifers . Limnology and Oceanography , 15 : 816 – 820 .
- Lin , L , Zhou , J and Huang , X-F . 1997 . The morphology of resting eggs of some rotifers in Lake Donghu, Wuhan . Acta Hydrobiologica Sinica , 21 : 334 – 240 . (in Chinese with English abstract)
- López , C , Soto , LM , Dávalos-Lind , L and Lind , O . 2007 . Summer dynamics of egg-ratio of the rotifer Keratella cochlearis (Gosse, 1851) in a eutrophic reservoir: a field study on affecting factors . Hydrobiologia , 589 : 175 – 185 .
- Merriman , JL and Kirk , KL . 2000 . Temporal patterns of resource limitation in natural populations of rotifers . Ecology , 81 : 414 – 149 .
- Ooms-Wilms , AL , Postema , G and Gulati , RD . 1999 . Population dynamics of planktonic rotifers in Lake Loosdrecht, the Netherlands, in relation to their potential food and predators . Freshwater Biology , 42 : 77 – 97 .
- Orcutt , J and Pace , M . 1984 . Seasonal dynamics of rotifer and crustacean zooplankton population in a eutrophic, monomictic lake with a note on rotifer sampling techniques . Hydrobiologia , 119 : 73 – 80 .
- Pace , ML , Cole , JJ and Carpenter , SR . 1998 . Trophic cascades and compensation: differential responses of microzooplankton in whole-lake experiments . Ecology , 79 : 138 – 152 .
- Pourriot , R . 1977 . Food and feeding habits of Rotifera . Archiv fur Hydrobiologie , 8 : 243 – 260 .
- Power , ME . 1990 . Resource enhancement by indirect effects of grazers: armored catfish, algae, and sediment . Ecology , 71 : 897 – 904 .
- Qian , F-P , Xi , Y-L , Wen , X-L and Huang , L . 2007 . The influence of eutrophication on community structure and species diversity of rotifers in five lakes . Biodiversity Science , 15 : 344 – 355 . (in Chinese with English abstract)
- Radwan , S . 1980 . The effect of some effect of some biotic and abiotic factors on the fertility of planktonic rotifer species . Hydrobiologia , 73 : 59 – 62 .
- Sarma , SSS , Gulati , R and Nandini , S . 2005 . Factors affecting egg-ratio in planktonic rotifers . Hydrobiologia , 546 : 361 – 373 .
- Schalau , K , Rinke , K , Straile , D and Peeters , F . 2008 . Temperature is the key factor explaining interannual variability of Daphnia development in spring: a modeling study . Oecologia , 157 : 531 – 543 .
- Snell , TW , Dingmann , BJ and Serra , M . 2001 . Density-dependent regulation of natural and laboratory rotifer population . Hydrobiologia , 446/447 : 39 – 44 .
- Sommer , U . 1989 . Plankton ecology: succession in plankton communities , Berlin : Springer-Verlag .
- Špoljar , M , Habdija , I , Primc-Habdija , B and Sipos , L . 2005 . Impact of environmental variables and food availability on rotifer assemblage in the Karstic Barrage Lake Visovac (Krka River, Croatia) . International Review of Hydrobiology , 90 : 555 – 579 .
- Straile , D . 2000 . Meteorological forcing of plankton dynamics in a large and deep continental European lake . Oecologia , 122 : 44 – 50 .
- Urabe , J . 1992 . Midsummer succession of rotifer plankton in a shallow eutrophic pond . Journal of Plankton Research , 14 : 851 – 866 .
- Virro , T , Haberman , J , Haldna , M and Blank , K . 2009 . Diversity and structure of the winter rotifer assemblage in a shallow eutrophic northern temperate Lake Võrtsjärv . Aquatic Ecology , 43 : 755 – 764 .
- Walz , N . 1987 . Comparative population dynamics of the rotifers Brachionus angularis and Keratella cochlearis . Hydrobiologia , 147 : 209 – 213 .
- Walz , N , Gschloessl , T and Hartmann , U . 1989 . Temperature aspects of ecological bioenergetics in Brachionus angularis (Rotatoria) . Hydrobiologia , 186/187 : 363 – 369 .
- Wen , X-L , Xi , Y-L , Zhang , L and Lu , X-J . 2006 . Analysis of community structure of Rotifera and ecological assessment of water quality in Lake Jinghu, Wuhu city . Acta Hydrobiologica Sinica , 30 : 152 – 158 . (in Chinese with English abstract)
- Williamson , CE . 1983 . Invertebrate predation on planktonic rotifers . Hydrobiologia , 104 : 385 – 396 .
- Williamson , CE and Butler , NM . 1986 . Predation on rotifers by the suspension-feeding calanoid copepod Diaptomus pallidus . Limnology and Oceanography , 31 : 393 – 402 .
- Xi , Y-L and Huang , X-F . 2000 . Effect of culture water replacement on resting egg formation of freshwater rotifer, Brachionus calyciflorus . Journal of Lake Science , 12 : 47 – 52 . (in Chinese with English abstract)
- Xi , Y-L and Huang , X-F . 2004 . Temperature effect on the life history of three types of Brachionus calyciflorus females . Chinese Journal of Oceanology and Limnology , 22 : 192 – 197 .
- Yoshida , T , Urabe , J and Elser , JJ . 2003 . Assessment of ‘top-down’ and ‘bottom-up’ forces as determinants of rotifer distribution among lakes in Ontario . Canada Ecological Research , 18 : 639 – 650 .
- Zhang , L , Xi , Y-L and Liu , G-Y . 2008 . Comparative studies on hatching of rotifera resting eggs in the sediments of different basins in Lake Jinghu . Acta Hydrobiologica Sinica , 32 : 6 – 12 . (in Chinese with English abstract)