Abstract
Standard bioassessment protocols compare macroinvertebrate communities in potentially impacted streams to communities in reference streams. However, in many landscapes ‘pristine’ reference streams may not exist and researchers may be forced to use the best available alternatives. The objective of this study was to evaluate macroinvertebrate communities in potential reference streams in the highly impacted Piedmont physiographic region of Georgia. We determined the influence of season, substrate, and reach-scale habitat type on macroinvertebrate assemblages in five forested, headwater streams that were minimally disturbed by current anthropogenic activities. The five streams represented three different reach-scale substrate compositions (gravel, sand, and bedrock). We predicted that macroinvertebrate communities in these streams would be influenced by season and by substrate composition. Streams were sampled in May, August, and November 2003, using a corer to sample riffles and a Hess sampler to sample runs and pools. Four of the five streams studied exhibited distinct seasonal variation, which exceeded spatial variation among those streams. Invertebrate community compositions among streams were not consistently predicted by reach-scale substrate in our study. The communities in the two sand streams were unique but generally dissimilar to each other. The two gravel streams supported similar communities across seasons. Riffle and wood habitats supported distinct invertebrate communities that were dissimilar to the run and pool habitats. Our data indicated that potential reference streams can vary widely in invertebrate community compositions and we cannot always predict that variability simply by substrate alone. Thus, selecting reference streams for bioassessment projects in the Georgia Piedmont may be complex.
Introduction
Aquatic macroinvertebrates have been widely and successfully used to assess the biological integrity of streams (Rosenberg et al. Citation2008). Most approaches to bioassessment require a measure of reference conditions which can be a site-specific or regional standard (Barbour, Diamond et al. Citation1996; Reynoldson et al. Citation1997). Site-specific references consist of samples taken within an impaired stream, typically upstream from the area of disturbance. Regional references involve sampling relatively unimpaired streams within a similar region and habitat type. In headwater streams, a range of approaches for determining reference conditions has been employed, ranging from rigorous statistical regional approaches to historical or paleo-ecological data to best professional judgment (Reynoldson et al. Citation1997). Measures of macroinvertebrate taxa richness, especially of the EPT taxa (Ephemeroptera, Plecoptera, Trichoptera), are considered particularly useful in establishing reference conditions in the Southeastern US (Barbour, Gerritsen et al. Citation1996) and elsewhere (Reynoldson et al. Citation1997).
The selection of reference conditions is important because any impairment of streams will be determined by comparing with the reference condition. Most bioassessment protocols specify time of year for sampling due to seasonal variation but these index periods may vary. For example, the index period for Georgia according to the Standard Operating Procedures of the Georgia Department of Natural Resources is mid-September through February, but Georgia Adopt-A-Stream advises to sample every three months to incorporate one sample from each season.
In some areas, it may be difficult to find reference streams that are devoid of significant impacts from human activities. The Piedmont physiographic region of Georgia in particular has a long history of human management. During the 1800s, European settlers almost completely deforested the region for agricultural production of tobacco and cotton (Trimble Citation1974). By 1930, more than 80% of the original forests in this area have been cleared (Bond and Spillers Citation1935). Agricultural practices of the 1800s and early 1900s led to the deposition of an average of 1.6 m of sediment in the floodplains of the Murder Creek Basin, Georgia, and Jackson et al. (Citation2005) estimated that this stream would need 6–10 millenia to transport that amount of sediment out of the watershed. High concentrations of suspended sediments have been shown to alter macroinvertebrate community structure because of poor substrate quality (Richards et al. Citation1993). From the 1930s–1980s, agricultural land use decreased from 30% to 12% of the landscape, while coniferous forest increased from 5% to 41% (Turner and Ruscher Citation1988). While shifts in land use from agriculture to forest may improve the quality of stream ecosystems, prior land use may still have an impact on current aquatic macroinvertebrate communities (Harding et al. Citation1998). Recently, the Georgia Piedmont has been subjected to expansive urbanization, and this development is now considered the primary threat to stream integrity (Roy et al. Citation2003; Walter et al. Citation2009).
In the Georgia Piedmont no stream may exist that has not been historically impacted by human intervention. Additionally, work on small headwater streams (drainage area <5 km2) of the Georgia Piedmont is limited and regional reference conditions for Piedmont headwater streams have yet to be described. Reference standards should be developed to assess on-going impacts to streams in this region. However, because ‘pristine’ reference streams may not exist, we may instead be forced to select the best alternatives available, recognizing that even in watersheds that have been forested for more than 50 years, macroinvertebrate communities might reflect previous degradation. These potential reference standards must account for the current variability in stream habitat and biota among watersheds.
This study was designed to quantify the influence of season, reach-scale substrate, and local substrate on macroinvertebrate communities in headwater streams of the Georgia Piedmont that currently are protected and are set in largely forested catchments. Each of these streams had the potential to be a suitable reference standard for the region. We expected that macroinvertebrate communities in these streams would vary with season and that streams with similar substrate compositions would contain similar aquatic macroinvertebrate communities, while those with different substrates would differ significantly. We also expected that different substrates within individual streams would support different macroinvertebrate assemblages. We selected five forested reference streams varying in dominant substrate type (two sand streams, two gravel streams, and one bedrock stream) and sampled macroinvertebrates in spring, summer, and fall.
Methods
Study sites
We selected five sites in the Piedmont region of the Upper Oconee River Watershed (). Streams were selected based on substrate composition (gravel, sand, and bedrock). The sites were all chosen in areas that are owned and protected by government agencies. We believe that these streams are likely candidates for reference streams because they are located in catchments that had been protected for 40–100 years. The streams were categorized by substrate characteristics and the descriptive metrics for each stream are shown in . Gravel-1 and Gravel-2 were located in the Oconee National Forest, Putnam County. Sand-1 and Sand-2 were both located near each other in the Oconee National Forest of Greene County. Bedrock-1 was located in the Rock Eagle 4-H Center of Putnam County. A second-bedrock stream was selected in the Georgia State Botanical Garden (Churchel and Batzer Citation2006), but was later rejected as a potential reference because organic pollution from groundwater was discovered.
Figure 1. Study site locations in the Oconee River Basin, Piedmont physiographic region, Georgia, USA.
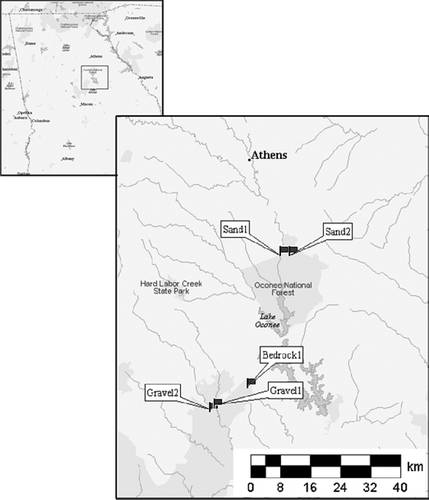
Table 1. Descriptive metrics of the five study streams located in the Piedmont region of the Upper Oconee River Watershed, Georgia, USA.
Sampling methods
Streams were sampled in May, August, and November 2003. In the gravel and sand streams three randomly selected locations for each of the three habitats present (riffles, runs, and pools) were sampled for a total of nine samples per stream. In the bedrock stream, four randomly selected locations for each of the two habitats present (riffles and runs) were sampled for a total of eight samples. We opted to use quantitative-sampling methods rather than qualitative or semi-quantitative approaches such as dip netting or kick sampling (Merritt et al. Citation2008). A Hess sampler (860 cm2, 250 µm mesh) was used to sample runs and pools by forcing the sampler into the streambed and washing the top 15 cm of substrate and water into the mesh collection bag. Because water depths in riffles were typically too shallow to efficiently use the Hess sampler, we used a core sampler (27.5 cm2) to sample riffles, removing all of the contents of the sampler down to 7.5 cm. The water in the core was removed and filtered through a 250 µm sieve and the contents were added to the rest of the collection. Even in the bedrock stream, riffles had sufficient gravel and sand to permit the use of the corer. Due to the relatively small size of the corer compared to the Hess sampler, three replicates were taken in each riffle and combined into one composite sample. Wood samples were taken by randomly collecting approximately 30 pieces of woody debris that were at least 1 cm in diameter. Large wood snags were rare in all study streams and were not sampled.
All samples were preserved in 95% ethanol and elutriated in the lab to separate the organic and inorganic material. Large samples were sub-sampled as needed and macroinvertebrates were removed from the organic material. Wood pieces were scrubbed to remove all organisms and all macroinvertebrates were removed from debris. Macroinvertebrates were identified to genus or the lowest taxonomic category possible (Pennak Citation1989; Peckarsky et al. Citation1990; Epler 1996; Merritt and Cummins Citation1996).
Pebble counts were carried out in each stream to justify our pairing of streams by substrate. Following the method developed by Wolman (Citation1954), this study was divided into 10 transects and 10 substrate particles were randomly selected and measured at each transect. Particles were only sampled from within the stream channel and the intermediate axis of each grain was measured and placed into size categories.
Statistical analyses
Invertebrate communities were analyzed for differences among the three months sampled (May, August, and November), the three substrates (gravel, sand, and bedrock), and the four habitats (riffles, runs, pools, and wood). We analyzed the presence or absence of taxa in all five streams by month using a cluster analysis with a Sorenson (Bray-Curtis) distance measure and a flexible beta group linkage method (α = −0.25). All habitats were combined for this analysis. To further examine the differences in these groupings, average number of taxa and EPT taxa were calculated for each habitat by month for all five streams. Overall number of taxa and EPT taxa were calculated for each stream by combining habitats. To evaluate differences between habitats in each stream, we used a non-metric multidimensional scaling (NMS) analysis, again using a Sorenson distance measure. All months were combined for this analysis. Like cluster analyses, NMS provides a means to visualize variation among samples using whole community data but in this case rather than a two-dimensional perspective, a multiple dimensional perspective in ordination space is employed. Cluster and NMS analyses were completed using PC-ORD Version 4.1 (MJM Software Design, Glenden Beach, OR, USA). Rare taxa (those that occurred in only one sample) were not included in the analysis. Those individuals that could not be identified were dealt in two ways. If there was an individual that could not be identified and only one identifiable subgrouping existed (e.g., one genus in a family), the unknown individual was combined with the others using the higher taxon (e.g., family). If there was an individual that could not be identified and multiple groupings existed (e.g., several genera from a family), the individual was deleted from the analysis.
Results
We collected and identified a total of 105,318 invertebrates from 91 taxa, including 15 rare taxa that were excluded from statistical analysis (Appendix). The most diverse orders were Diptera, with 15 families and 21 genera and Trichoptera, with 11 families and 14 genera. Average richness was 51 taxa per stream with a range of 40–58 taxa per stream.
Cluster analysis of invertebrate communities generally separated the five streams into groups based either on stream or on the month sampled (). Sand-2 was largely unique from the other streams; the May and November samples from Sand-2 clustered weakly together, but the August Sand-2 sample was unlike any other. The May sample from Sand-1 clustered with the May and November Sand-2 samples. Thereafter, Sand-1 samples clustered with samples from other streams collected during the same month. Bedrock-1, Gravel-1, and Gravel-2 samples from May constituted one grouping and August and November samples from Bedrock-1, Gravel-1, Gravel-2, and Sand-1 constituted another grouping.
Figure 2. Cluster analysis of invertebrate communities at five streams with each month treated separately (G = Gravel, S = S, and B = Bedrock). Samples were collected in May, August, and November, 2003, and community structures for each sample reflect replicate sampling of all sub-habitats present (riffles, pools, runs, and wood).
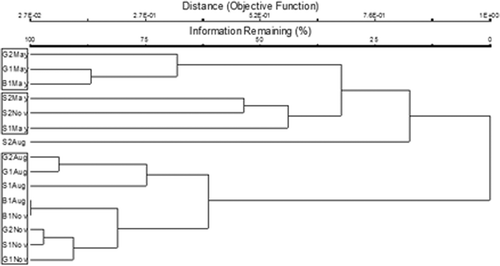
Sand-2 was unique from the other streams mainly due to the rarity of taxa present. Pisidium, Lepidostoma, Stenelmis, Dubiraphia, Lype, Chrysops, Nigronia, Seira, Macronychus, Tipula, and Leptophlebiidae were present in Sand-2 during only one month, but were present in at least 60% of the samples from other streams. Anchytarsus was present in all other samples, but was absent from all Sand-2 samples. Sand-2 contained fewer total taxa and somewhat fewer EPT taxa than the other four streams during most sample months (). While both of these metrics generally increased in the other streams from May to August, a decrease was observed in Sand-2 with only 20 taxa and three EPT taxa collected in August.
Table 2. Average number of taxa and EPT taxa in five streams for each habitat type and for all habitats combined (Ri = riffles, Ru = runs, P = pools, and W = wood).
Most variation among the other four streams was due to seasonal change. Taxa found only in the May samples included: Alloperla, Sialis, and Dineutus. From May to August, the total number of taxa and number of EPT taxa in Gravel-1, Gravel-2, Sand-1, and Bedrock-1 increased by an average of 29.6% and 75.0%, respectively (). From August to November, smaller increases were observed in the gravel streams, while these metrics decreased in Sand-1 and Bedrock-1. Twenty taxa were absent in May but present in August and November, with the most common being Anisocentropus, Calopteryx, Diplectrona, Dixa, Hemerodromia, and Hexagenia.
The NMS ordination of communities indicated two main groupings (). The wood samples, except for Bedrock-1, clustered together as having a distinct community composition. The riffle samples for all five streams grouped together. However, the run and pool samples were intermixed in ordination space. For some streams, samples clustered together regardless of habitat. The riffle, run, and pool samples from Sand-1 grouped closely together as having similar communities, and even the wood sample for Sand-1 was in close proximity ().
Figure 3. NMS ordination of sub-habitats based on invertebrate community composition. Final stress = 22.1 (for two-dimensional solution). Replicate samples were collected from riffles, runs, pools, and/or wood from five streams Gravel (G) 1, Gravel 2, Sand (S) 1, Sand 2, and Bedrock (B) 1. Collections were made in May, August, and November, 2003, and pooled (by sub-habitat) for this analysis.
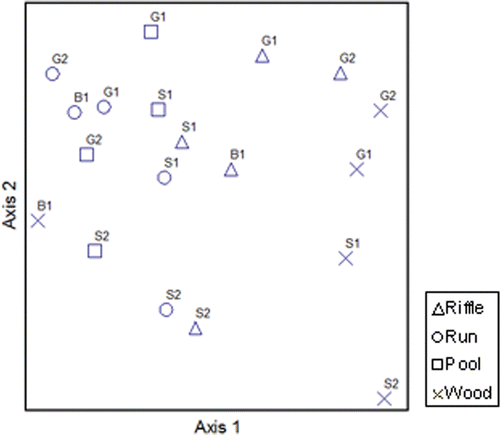
Among the habitats richness was highest in the riffles with an average of 37 taxa and lowest in the wood habitats with an average of 27 taxa. The grouping of wood habitats probably occurred due to low richness. Odontomyia and Scirtidae were the only taxa present in the wood samples that were not present in the other habitats. Twenty-nine taxa found elsewhere were not collected from wood. The grouping of riffle habitats was likely due to high richness. Four taxa were found only in riffle habitats: Nemouridae, Neophylax, Eurylophella, and Sminthuridae.
Discussion
All of the streams used in this study are likely candidates as reference streams. They were all in areas that are managed by government agencies and had been protected for 40–100 years. Their catchments contained largely undeveloped, mature-growth forest, and no roads crossed the streams upstream from the sampling reaches. We had anticipated that the sand streams might reflect historical sedimentation but environmentally sensitive EPT taxa were still common in these habitats. Most variations among streams were generated by invertebrates other than the EPT taxa.
The Gravel-1, Gravel-2, Sand-1, and Bedrock-1 streams exhibited distinct seasonal variation and this temporal variation exceeded spatial variation for those streams. In the Sand-2 stream, some temporal variation was evident, but this change did not track the pattern observed in the Gravel-1, Gravel-2, Sand-1, and Bedrock-1 streams. Within the streams that exhibited strong temporal variation, the gravel streams were the only category that clustered together for each month.
We had expected that streams with similar substrate composition would have similar invertebrate communities and, therefore, similarities in reach-scale substrate would be a crucial factor in choosing a reference stream. Fish assemblages in other Piedmont streams have been shown to vary based on differences in physical parameters, including substrate composition (Walters et al. Citation2003). In other regions, macroinvertebrate communities are more strongly influenced by substrate composition than other physical characteristics (Richards et al. Citation1993; Stead et al. Citation2003; Buss et al. Citation2004). However, invertebrate community composition among streams did not group strongly by substrate in our study. The community in Sand-2 was clearly unique from the other streams and was also generally dissimilar to Sand-1.
The unique character of riffle habitats was likely due to higher numbers of taxa being present, including some unique taxa. Other researchers have observed higher total macroinvertebrate abundances and higher individual taxa abundances in riffles than in other habitats sampled (Buss et al. Citation2004; Halwas et al. Citation2005). As in many streams, riffle habitats in Piedmont headwater streams should be prioritized for sampling. The unique character of communities on wood developed primarily because fewer taxa occurred on wood and not because the material supported numerous unique taxa. Unlike in sandy streams of the Coastal Plain (Drury and Kelso Citation2000; Rinella and Feminella Citation2005), wood appears to be a less-important structuring feature for macroinvertebrates in Piedmont headwater streams.
The value of headwater streams has been widely recognized but little research in these systems has been conducted in the Georgia Piedmont region. This study not only provides an extensive invertebrate taxa list for five Piedmont streams, it also shows how variable communities are among streams and within individual streams. A goal of this study was to explore reference criteria to bolster efforts to monitor anthropogenic impacts of similar streams in this region. However, our data indicate that potential reference streams can vary widely in invertebrate community compositions and that variability cannot be predicted by substrate alone. Pairing of streams based on substrate composition or proximity may not always be appropriate. Seasonality can be pronounced and should be considered in sampling programs. Sampling only in one season would not provide a complete picture of the macroinvertebrate communities in these streams. Given the large natural temporal and spatial variation, only anthropogenic impacts that exceed the large natural variability might be detectable using macroinvertebrates.
Including numerous additional reference streams to our data set might compensate for uncertainty induced by natural variability but whether sufficient high-quality streams are available in the region is questionable. Multimetric and multivariate approaches to bioassessment typically require an array of reference sites to characterize the biological condition of a region (Reynoldson et al. Citation1997). With so few minimally impacted streams being present in the Georgia Piedmont and the high variability in the ones that exist, those approaches might not be practical. Perhaps approaches that use site-specific, rather than regional, references might be most appropriate for macroinvertebrate bioassessment in the Georgia Piedmont. Knowing how invertebrate communities might naturally vary within streams, as described in this study, would be valuable information for a site-specific approach.
Acknowledgments
This project was funded by the U.S. Environmental Protection Agency and the Hatch Fund. We would like to thank Mary Freeman and the reviewers for their comments and suggestions. We would also like to thank Jennifer Henke, Rich Schultheis, and Ginny Lipscomb for their assistance in the field and in the lab.
References
- Barbour , MT , Diamond , JM and Yoder , CO . 1996 . “ Biological assessment strategies: applications and limitations ” . In Whole effluent toxicity testing: an evaluation of methods and prediction of receiving system impacts , Edited by: Grothe , DR , Dickson , KL and Reed-Judkins , DR . 245 – 270 . Pensacola , FL : SETAC Press .
- Barbour , MT , Gerritsen , J , Griffith , GE , Frydenborg , R , McCarron , E , White , JS and Bastain , ML . 1996 . A framework for biological criteria for Florida streams using benthic macroinvertebrates . Journal of the North American Benthological Society , 15 : 185 – 211 .
- Bond , WE and Spillers , AR . 1935 . Use of land for forests in the lower Piedmont region of Georgia , Occasional paper 53. [Asheville (NC)] : Southern Forest Experiment Station .
- Buss , DF , Baptista , DF , Nessimian , JL and Egler , M . 2004 . Substrate specificity, environmental degradation and disturbance structuring macroinvertebrate assemblages in neotropical streams . Hydrobiologia , 518 : 179 – 188 .
- Churchel , MA and Batzer , DP . 2006 . Recovery of aquatic macroinvertebrate communities from drought in Georgia Piedmont headwater streams . American Midland Naturalist , 156 : 259 – 272 .
- Drury , DM and Kelso , WE . 2000 . Invertebrate colonization of woody debris in coastal plain streams . Hydrobiologia , 434 : 63 – 72 .
- Epler , JH . 1996 . Identification manual for the water beetles of Florida (Coleoptera: Dryopidae, Dytiscidae, Elmidae, Gyrinidae, Haliplidae, Hydraenidae, Hydrophilidae, Noteridae, Psephenidae, Ptilodactylidae, Scirtidae) , 257 Tallahassee , FL : Florida Department of Environmental Protection .
- Halwas , KL , Church , M and Richardson , JS . 2005 . Benthic assemblage variation among channel units in high-gradient streams on Vancouver Island, British Columbia . Journal of the North American Benthological Society , 24 : 478 – 494 .
- Harding , JS , Benfield , EF , Bolstad , PV , Helfman , GS and Jones , III . 1998 . EDB. , Stream biodiversity: the ghost of land use past. Proceedings of the National Academy of Science. 95:14843–14847
- Jackson , CR , Martin , JK , Leigh , DS and West , LT . 2005 . A southeastern Piedmont watershed sediment budget: evidence for a multi-millennial agricultural legacy . Journal of Soil and Water Conservation , 60 : 298 – 310 .
- Merritt , RW and Cummins , KW . (editors). 1996. An introduction to the aquatic insects of North America. 3rd ed. Dubuque (IA): Kendall/Hunt Publishing Co., 862 pp
- Merritt , RW , Cummins , KW , Resh , VH and Batzer , DP . 2008 . “ Sampling aquatic insects; collection devices, statistical considerations, and rearing procedures ” . In An introduction to the aquatic insects of North America , 4th ed , Edited by: Merritt , RW , Cummins , KW and Berg , MB . 15 – 37 . Dubuque , IA : Kendall/Hunt Publishers .
- Peckarsky , BL , Fraissinet , PR , Penton , MA and Conklin Jr , DJ . 1990 . Freshwater Macroinvertebrates of Northeastern North America , 442 Ithaca , NY : Cornell University Press .
- Pennak , RW . 1989 . Fresh-Water Invertebrates of the United States: Protozoa to Mollusca , 3rd ed , 656 New York , NY : John Wiley and Sons, Inc .
- Reynoldson , TB , Norris , RH , Resh , VH , Day , KE and Rosenberg , DM . 1997 . The reference condition: a comparison of multimetric and multivariate approaches to asses water-quality impairment using benthic macroinvertebrates . Journal of the North American Benthological Society , 16 : 833 – 852 .
- Richards , C , Host , GE and Arthur , JW . 1993 . Identification of predominant environmental factors structuring stream macroinvertebrate communities within a large agricultural catchment . Freshwater Biology , 29 : 285 – 294 .
- Rinella , DJ and Feminella , JW . 2005 . Comparison of benthic macroinvertebrates colonizing sand, wood, and artificial substrates in a low-gradient stream . Journal of Freshwater Ecology , 20 : 209 – 220 .
- Rosenberg , DM , Resh , VH and King , RS . 2008 . “ Use of aquatic insects in biomonitoring ” . In An introduction to the aquatic insects of North America , Edited by: Merritt , RW , Cummins , KW and Berg , MB . 123 – 138 . Dubuque , IA : Kendall/Hunt .
- Roy , AH , Rosemond , AD , Leigh , DS , Paul , MJ and Wallace , JB . 2003 . Habitat-specific responses of stream insects to land cover disturbance: biological consequences and monitoring implications . Journal of the North American Benthological Society , 22 : 292 – 307 .
- Stead , TK , Schmid-Araya , JM and Hildrew , AG . 2003 . All creatures great and small: patterns in the stream benthos across a wide range of metazoan body size . Freshwater Biology , 48 : 532 – 547 .
- Trimble , SW . 1974 . Man induced soil erosion on the Southern Piedmont, 1700–1970 , 188 Ankeny , IA : Soil Water Conservation Society .
- Turner , MG and Ruscher , CL . 1988 . Changes in landscape patterns in Georgia, USA . Landscape Ecology , 1 : 241 – 251 .
- Walters , DM , Leigh , DS , Freeman , MC , Freeman , BJ and Pringle , CM . 2003 . Geomorphology and fish assemblages in a Piedmont river basin, U.S.A . Freshwater Biology , 48 : 1950 – 1970 .
- Walters , DM , Roy , AH and Leigh , DS . 2009 . Environmental indicators of macroinvertebrates and fish assemblage integrity in urbanizing watersheds . Ecological Indicators , 9 : 1222 – 1233 .
- Wolman , MG . 1954 . A method of sampling coarse river bed material . Transactions of the American Geophysical Union , 35 : 951 – 956 .