
Abstract
Most studies of organic matter transport have been conducted on small headwater streams and have found that floods strongly influence downstream transport. We studied five sites on four rivers, each with different hydrographs and flood frequencies, in the Adirondack Mountains, New York, USA, during the summer and autumn of 2005. Sites on three rivers had more natural hydrographs, whereas recreational releases in the Indian River increased discharge 10-fold and to a lesser extent in the upper Hudson River below their confluence. Fine and coarse particulate organic matter (FPOM and CPOM, respectively) transport decreased from June to August at base flow in most rivers, concurrent with summer decreases in discharge; however, during releases CPOM transport increased in the Indian River and at the downstream Hudson River site below the confluence with the Indian River. Also, in the summer there was a shift from terrestrial POM in transport in June to more filamentous algae in July and August because of sloughing of periphyton. In October, POM transport increased at most sites from August levels because of autumnal leaf inputs. In summer 2006, we sampled all sites during and after a natural flood, and both FPOM transport and CPOM transport were highest in rivers with more natural hydrographs (>10,000% increase from base-flow transport levels after the flood) than in the Indian and downstream Hudson sites (∼1000% increase or less). Our results suggest that recreational releases do reduce POM sources from floodplain and benthic habitats, based on lower POM transport during a natural flood.
Introduction
Particulate organic matter (POM) varies in streams and rivers, depending on whether it is of allochthonous origin from the riparian corridor or is part of an autochthonous aquatic plant, algal or bacterial community (Minshall Citation1978; Vannote et al. Citation1980). Sources differ in importance depending on the longitudinal position along a river system, season (especially in temperate regions) and type of riparian vegetation (Vannote et al. Citation1980; Minshall et al. Citation1985; Webster and Meyer Citation1997; Tank et al. Citation2010). Forested headwater streams typically are dominated by seasonal allochthonous inputs of coarse particulate organic matter (CPOM), but dissolved organic matter may also be significant (Vannote et al. Citation1980; Webster and Meyer Citation1997; Tank et al. Citation2010).
The downstream transport of POM is dependent on the particle size with smaller size fractions (<1.0 mm) being influenced mostly by hydrology (Thomas et al. Citation2001), whereas CPOM (>1.0 mm) transport is determined by hydrology and abundance of retention devices (Naiman Citation1982; Wallace et al. Citation1995; Golladay Citation1997). Macroinvertebrate shredding activities can influence the amount of POM in transport through conversion of CPOM into fine particulate organic matter (FPOM < 1.0 mm) as well as the capture of FPOM by filter-feeding collectors (Wallace and Webster Citation1996). Forested headwater streams typically have lower concentrations of FPOM in suspension compared to CPOM because of greater CPOM inputs from riparian areas (Naiman and Sedell Citation1979; Vannote et al. Citation1980). As POM is transported downstream, it decreases in size and becomes more refractory, resulting in a decrease in the CPOM-to-FPOM ratio at greater distances from the headwaters (Webster et al. Citation1995). Most studies of organic matter transport dynamics have occurred in low-order streams (Neatrour et al. Citation2004; Wipfli et al. Citation2007) and demonstrate a seasonal influence related to input dynamics and changes in discharge (Wallace et al. Citation1982, Citation1995; Jones and Smock Citation1991; Atkinson et al. Citation2009). Yet there is growing interest in understanding how well processes observed in small streams can be transferred (i.e. scaled) to understand larger stream and river ecosystems (e.g. Tank et al. Citation2008).
Flood-type disturbances have been shown to increase organic matter transport through scouring processes that mobilize POM from the stream bed as well as from inundated floodplains and adjacent riparian areas (Wallace et al. Citation1995; Jakob et al. Citation2003; Neatrour et al. Citation2004; Atkinson et al. Citation2009). However, the relationship between discharge and organic matter transport is complex because of variations in flow conditions (magnitude and frequency of floods), season of occurrence, hysteresis, particle sizes present, and their relative availability. All of these factors influence the amount of organic matter in seston (Cuffney and Wallace Citation1989; Webster et al. Citation1990, Citation1995, Citation1999; Atkinson et al. Citation2009; Tank et al. Citation2010). As the frequency of flood-type disturbances increases, seston concentrations sometimes decrease because of exhaustion of POM sources due to the frequent scouring events (Pozo et al. Citation1994).
We studied five sites on four rivers in the Adirondack Mountains of New York, USA, that varied in flood frequency, hydrology, and geomorphology. The Indian River experiences recreational flow releases from the Abanakee Dam (four times/week from April to October) that create bankfull flood events. These recreational flow releases support a whitewater rafting industry, although their frequency and duration during the summer months are similar to flow releases associated with hydro-peaking from summer month power production. The Indian River flows into the Hudson River near its headwaters; therefore, the Hudson River below their confluence is also influenced by these recreational releases. We also sampled two other rivers with more natural hydrographs to compare to the Indian and Hudson Rivers. Our objectives were to compare organic matter transport dynamics in human-altered and free-flowing rivers (1) at base flow throughout the summer and in the autumn in rivers with different flood frequencies, (2) between base flow and recreational releases for sites experiencing releases, and (3) among all sites during and after a natural flood event.
Methods
Study sites
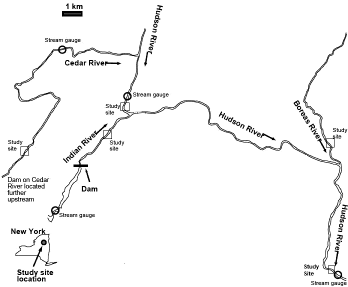
The study was conducted in the Indian, upper Hudson, Cedar, and Boreas Rivers in the Adirondack Mountains, New York, USA, from June to October 2005 and in June and July 2006 (Figure ). Study sites were selected to provide a range of flow regimes, both natural and anthropogenic. All sites are located in an area of metamorphic rock overlain with glacial till deposited during the retreat of the Wisconsin glaciation, c. 10,000 yr BP. Channels in the region were formed in alluvial material and include erratics distributed throughout the valleys with particularly frequent large boulders (>1 m diameter) in all the study sites. Regional hydrology is driven by snowmelt peak flows in April to May, low flow in July and August interrupted by local thunderstorms, and large flows via frontal systems often associated with tropical depressions in the autumn. All rivers flow through forested drainage basins containing mostly mixed hardwood trees (represented by the genera Acer, Fagus, Prunus, Betula, and Fraxinus) along with some hemlock (Tsuga canadensis) and scattered spruce trees (Picea spp.).
Figure 1. Map showing five sample sites on four rivers in the Adirondack Mountains of New York, USA.
The Indian River downstream of the Abanakee Dam (a 5-m-low-head dam with a surface spillway) experiences recreational releases that were originally restricted to the spring and autumn months (1980s) when river levels were already high from snowmelt and rain storms. In the mid-1990s, releases were allowed throughout the summer; thus, these releases have been occurring for over a decade prior to our study. These recreational flow releases elevate discharge in the Indian River regularly throughout the summer months, creating an extremely high frequency of short-duration disturbances increasing discharge from a base flow of 2.5–4.0 m3/s to 35–43 m3/s (Figure ), which is very close to the geomorphic bankfull discharge in the segment but less than observed peak discharges at the site (e.g. 97 m3/s on 29 June 2006). Flow releases last from 1.5 to 2.0 hours, and occur 4 days each week (Tuesday, Thursday, Saturday, and Sunday) from April to October to support the whitewater rafting industry (see Fuller et al. Citation2011).
Figure 2. Photographs depicting the Indian River below the Abanakee Dam at base flow (left-hand side) and during a recreational release (right-hand side).

Our sample site on the Indian River was located approximately 3 km downstream from the Abanakee Dam (Figures and ) in a low-gradient reach of the Indian River (gradient of sample site 0.0031 compared to 0.02–0.03 elsewhere along the segment; Table ). The lower gradient reach allowed us to collect samples and make hydraulic measurements during the flow releases, which was not possible elsewhere along the river. There are a few, small intermittent streams entering the Indian River upstream of our sample site. Because the dam retains the upstream-derived sediment load and there is negligible bank erosion along the reach, these small tributaries are the primary sources of any sediment downstream of the dam. The width of the channel is 30–50 m in this area, with a large island upstream from the site where we sampled. The substrata consist of boulders, cobbles, and pebbles. The depth is 30–50 cm at base flow with water levels rising 40 cm or more during a release (Table ; see Fuller et al. Citation2011).
Table 1. Location and physical parameters of study sites on four rivers in the Adirondack Mountains, New York, USA.
The Indian River flows into the Hudson River, so the Hudson River upstream of the confluence has a natural hydrologic regime but below the confluence has a manipulated regime. We sampled two sites on the Hudson River. The first site (upstream Hudson) was approximately 500 m upstream from the confluence with the Indian River, with substrata consisting of large boulders, cobbles, pebbles, and sand; the channel width was approximately 80 m and depths ranged from 15 to 100 cm depending on proximity to large boulders (Figure ; Table ). This site acted as a reference site for comparison with a downstream site on the Hudson River (designated ‘downstream Hudson’) located approximately 15 km downstream from the confluence with the Indian River. This downstream site on the Hudson River is influenced by the recreational releases from the Abanakee Dam but to a lesser extent because of the dampening effect of the larger volumes of water in the Hudson River, resulting in an ∼35% increase in discharge during a release (from approximately 73 m3/s to 100 m3/s) compared to the order of magnitude increase in the Indian River. The substrata at this downstream site consist of boulders, cobbles, and pebbles but less sand than upstream; the channel is approximately 80 m wide in most areas and the depth ranges from 15 to 150 cm (Table ).
The Cedar River site was chosen as a site with an upstream impoundment but no flow releases (Figure ; Table ). There is a run-of-river dam approximately 4 km upstream from the sample site and while this run-of-river dam does not have water releases, it does slightly attenuate the rise and fall of discharge during flood events. The sampling site on the Cedar River is located upstream of a significant geologic constraint that reduces the gradient to 0.0021. This reduced gradient also reduces shear stress exerted by flows and thus the site has accumulated pebbles and sand, in addition to the boulder and cobble characteristic of the glaciated region. In this way, the sampling site on the Cedar River is comparable to the reduced-slope reach site on the Indian River. The width of the river is 35 m and the depths range from 20 to 70 cm.
Finally, the Boreas River site was chosen as a site to represent a completely unregulated river and one with high gradient and high natural disturbance potential (Figure ; Table ). The site has a gradient of 0.0111, with velocities exceeding 3 m/s during flood events. The bed sediment is large and comparable to the upstream Hudson site, with substrata being dominated by boulders and cobbles with very little fine gravel or sand. The width of the river was 20 m and depths ranged from 20 to 70 cm.
Particulate organic matter transport
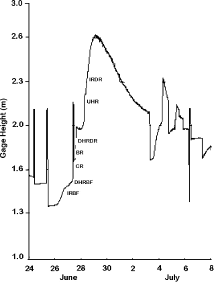
Samples were collected at all study sites in June, July, August and October in 2005. In addition, in June 2006 we sampled each site during a 3-day rainstorm event, collecting samples on the rising limb of this major flood (Figure ); we sampled all sites again between 16 and 20 July 2006 when flood waters receded in the Indian River enough to allow us to safely sample. Under normal base-flow conditions, five drift nets (1350 cm2 aperture opening, length 1 m, and mesh openings 363 μm) were placed along a transect across the river at each site for 30 min. Current velocities at each net were recorded using a Marsh McBirney Flow-Mate (Hach Corporation, http://www.hach.com). During releases in the Indian River and at the downstream Hudson River site, five replicate samples were collected all within 3 m of the shoreline for safety reasons; a modified kick net (1500 cm2 aperture opening, length 1 m, and mesh opening 363 μm) was held in the river for 2 min; current velocities were measured at the same place where drift samples were taken. All POM samples were preserved with 5% formalin in 500 ml glass jars. In the laboratory, the samples were filtered through a 250 μm sieve and the macroinvertebrates were removed from the organic matter. The organic matter was separated using sieves of length 1 mm and 250 μm mesh sizes and organic matter from each sieve was placed in pre-weighed crucibles, and then dried at 60°C for at least 48 hours. The crucible with organic matter, >1 mm in length (CPOM transport), was then weighed on an electronic balance (±0.01 mg; Sartorius 1712 MP8, http://www.sartorius.com). The crucible with organic matter from the 250 μm mesh sieve (FPOM transport) was weighed, ashed in a muffle furnace at 550°C for 1 hour, and weighed again.
Figure 3. Gage height during late June and early July in 2006 during a natural flood. The letters on the rising limb of the hydrograph indicate when samples were taken at each site during the flood (BR = Boreas River; CR = Cedar River; UHR = upstream Hudson River; DHRBF = downstream Hudson River at base flow; DHRDR = downstream Hudson River during a release; IRBF = Indian River at base flow – prior to opening the flood gate; IRDR = Indian River during a release – after opening the flood gate).
Data analysis
Values for each parameter (FPOM and CPOM transport) were compared separately among all sites and sample dates during 2005 using a two-way analysis of variance (JMP statistical software; http://www.jmp.com). Similarly, FPOM and CPOM transport values in 2006 were compared separately among sites and stages (during and after the flood) using a two-way analysis of variance; after finding significant differences among sites and dates (or stages), a one-way analysis of variance was conducted followed by a Tukey multiple comparison test (α = 0.05) to determine differences among sites and months. All presented results are at p < 0.05 unless indicated otherwise. When data were not normally distributed, a log10(x + 1) transformation was performed to obtain a more normal distribution.
Results
During the summer and autumn there were significant differences in FPOM transport among sites (F6, 112 = 45.58; p < 0.0001) and months (F3, 112 = 23.43; p < 0.0001), and there was a significant interaction between sites and months (F18, 112 = 20.11; p < 0.0001; Table ). The strong interaction was likely because of high FPOM transport values in the Cedar River in June but lower values for all other sample dates (). In June 2005, the Cedar River had similar FPOM amounts in transport to the Indian and downstream Hudson Rivers during a release; these amounts were significantly greater (F6, 26 = 5.005; p < 0.0016) than amounts in the Indian and downstream Hudson Rivers at base flow as well as in the Boreas and upstream Hudson Rivers all of which were similar (Figure ). In July, there was less difference among sites in suspended FPOM (F6, 26 = 8.323; p < 0.0001) with only the downstream Hudson River site during a release having significantly higher FPOM in transport than other sites at base-flow conditions (Table ); the Indian River during a release had intermediate amounts of FPOM in transport, but was still similar to all other sites (Figure ). In August, the Indian and downstream Hudson River sites during a release again had significantly higher suspended FPOM than all other sites (F6, 26 = 181.3; p < 0.0001); all other sites had equal amounts of FPOM in transport (Table ; Figure ). Similarly, in October the Indian River during a release had significantly higher amounts of FPOM in transport (F6, 26 = 65.94; p < 0.0001) than all other sites, which were all equal.
Table 2. Two-way ANOVA results comparing the transport of FPOM and CPOM among sites and months in the Adirondack Mountain Rivers in 2005 and among sites at different stages (during a natural flood and base-flow conditions) in 2006.
Table 3. One-way ANOVA results and multiple comparisons among sites in the Adirondack Mountains, New York, USA, for FPOM and CPOM in transport during base flow in 2005 and among sites at different stages (during a natural flood and base-flow conditions) in 2006. Different letters in a row indicate a significant difference (p < 0.05) among sites (BR = Boreas River; CR = Cedar River; UPHR = upstream Hudson River; DHRBF = downstream Hudson River base flow; IRBF = Indian River base flow; DHRD = downstream Hudson River during a release; IRDR = Indian River during a release).
Figure 4. Monthly mean dry weights of FPOM (A) and CPOM (B) in transport at five sites on four rivers in the Adirondack Mountains, New York, USA, from June to October 2005 (BF = base-flow conditions; DR = during a release; lines at the top of bars are +1 SE).
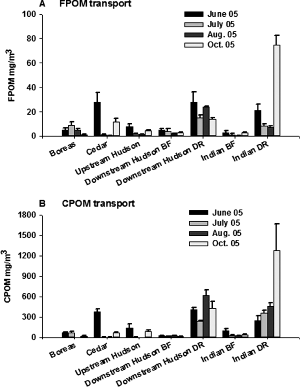
CPOM transport followed the same general trends as observed for FPOM transport with significant differences among sites (F6, 112 = 27.79; p < 0.0001) and months (F3, 112 = 511; p < 0.003; Table ). Also, there was a strong interaction between sites and months (F18, 112 = 6.43; p < 0.0001), again due to high transport values in the Cedar River in June but lower values for all other sample dates (Figure , Table ). In June 2005, CPOM transport was significantly higher in the Cedar River and during releases in the Indian and downstream Hudson Rivers (F6, 26 = 13.23; p < 0.0001; Figure ); all other sites had lower and similar CPOM transport values (Table , Figure ). During July and August (F6, 28 = 81.973; p < 0.0001 and F6, 26 = 50.228; p < 0.0001, respectively), CPOM transport was significantly higher in the Indian and downstream Hudson River sites during a release compared to base-flow conditions and all other river sites (Table ). In October, a similar trend was observed (F6, 26 = 8.88; p < 0.0001), although amounts of CPOM in transport increased at most sites compared to their August values because of leaf inputs from riparian forests.
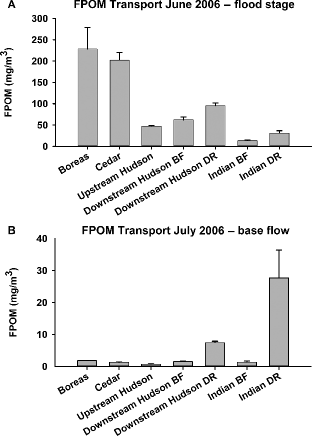
During June and July of 2006, there were significant differences in FPOM transport among sites (F6, 56 = 15.56; p < 0.0001) and between stages (F1, 56 = 131.32; p < 0.0001) and there was a significant interaction between sites and stages (F6, 56 = 19.06; p < 0.0001; Table 2 and 3; Figure ). The strong interaction term is likely the result of much lower FPOM transport values in the Boreas and Cedar Rivers at base flow and much higher transport during the flood (Figure ). At base-flow conditions after the flood, FPOM transport was significantly higher (F6, 28 = 200.41, p = 0.0001) during releases in the Indian and downstream Hudson Rivers compared to all other sites (Table , Figure ) with the lowest FPOM transport at the upstream Hudson site. However, during the flood (F6, 28 = 51.69, p = 0.0001), the highest FPOM transport was in the Cedar and Boreas Rivers (p < 0.05), with lower values at all other sites; the lowest transport values occurred in the Indian River prior to the raising of the flood gate as well as after the flood gate was raised. Per cent increases in FPOM transport during the flood versus base flow were >12,000% for the Boreas and Cedar Rivers compared to <1,000% in the Indian River and downstream Hudson site after the Abanakee Dam flood gate was raised (Table ).
Table 4. Per cent increase in FPOM and CPOM transport during the June 2006 flood compared to transport during base-flow conditions in July 2006 in four rivers in the Adirondack Mountains, New York, USA. The higher increases in the Boreas, Cedar and upstream Hudson Rivers reflect effects on POM transport in rivers not experiencing frequent recreational releases versus the downstream Hudson and Indian River sites where recreational releases occur 4 days/week from April to October (BF = base-flow conditions; DR = during a recreational release).
Figure 5. Mean dry weights of FPOM in transport during a natural flood in June 2006 (A) and following the flood in July 2006 at base-flow conditions (B) for five sites on four rivers in the Adirondack Mountains, New York, USA (BF = base-flow conditions; DR = during a release; lines at the top of bars are +1 SE).
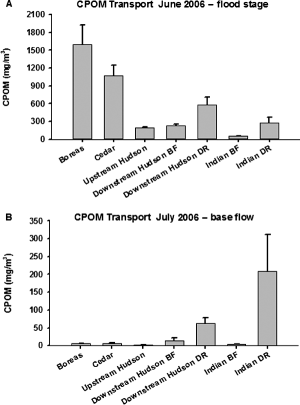
CPOM transport dynamics showed a similar trend to FPOM transport with significant differences among sites and stages (F6, 56 = 10.49, p < 0.0001; F1, 56 = 108.43; p < 0.0001, respectively; Table ; Figure ); also, there was a significant interaction between sites and stage (F6, 56 = 12.01; p < 0.0001). Again, the significant interaction term is likely because of the low CPOM transport values in the Cedar and Boreas Rivers at base flow versus their very high transport values during the flood. At base flow after the flood (F6, 28 = 47.75, p < 0.0001), CPOM transport values were significantly higher in the Indian and downstream Hudson Rivers during a release, and all other sites were similar except the upstream Hudson River site, which was significantly lower (Figure ). During the flood in late June (F6, 28 = 46.21, p < 0.0001), CPOM transport was significantly higher in the Boreas and Cedar Rivers, intermediate at the two Hudson River sites, and lowest at the Indian River site. Per cent increase in CPOM transport between the flood stage and base flow was highest in the Boreas, Cedar, and upstream Hudson sites (>12,000%), whereas the Indian River and downstream Hudson sites had much lower per cent increases in CPOM transport during the flood (120–3300%; Table ).
Figure 6. Mean dry weights of CPOM in transport during a natural flood in June 2006 (A) and following the flood in July 2006 at base-flow conditions (B) for five sites on four rivers in the Adirondack Mountains, New York, USA (BF = base-flow conditions; DR = during a release; lines at the top of bars are +1 SE).
Discussion
Most studies of organic matter transport in streams and rivers have found that FPOM in transport is much more abundant than CPOM at base-flow conditions and that only during flooding events does CPOM transport exceed FPOM transport (Wallace et al. Citation1982; Webster and Meyer Citation1997; Tank et al. Citation2010). CPOM that enters streams is trapped by various retention devices and converted to FPOM via invertebrate feeding, abrasion, and microbial activity, with these processes being especially important in forested headwater streams. The hydrologic connectivity of headwater streams with downstream reaches results in FPOM being more easily transported than CPOM to the lower reaches of a river network (Webster and Meyer Citation1997; Wipfli et al. Citation2007). We suspect that our higher CPOM in transport at base-flow conditions is likely the result of not quantifying the smaller FPOM size fractions (<100 μm) that are often the dominant FPOM size fractions in transport (Naiman Citation1982); our net mesh size (363 μm) did not capture these more dominant, smaller size fractions of FPOM.
We observed a general decrease in transport of both FPOM and CPOM from June to August during base-flow conditions which reflects a close relationship between transport and discharge. In October, we observed higher CPOM transport with seasonal leaf abscission contributing greater amounts of leaf material to the river channels. Others have observed similar seasonal increases in CPOM transport in streams bordering deciduous forests (Benfield Citation1997; Webster et al. Citation1999); however, these studies occurred in small headwater streams. Wipfli et al. (Citation2007) suggested that if retention is high in tributary streams, the pulse of CPOM inputs to larger rivers may be delayed until snowmelt in the spring. Our June CPOM transport values were among the highest at most of our sites, perhaps suggesting a seasonal delay in CPOM transport from their tributary streams to these rivers. Boulders can act as important retentive devices (Nakajima et al. Citation2006) and the high boulder densities in all our rivers could have influenced transport of CPOM. However, during a release in the Indian River in October, CPOM transport was much higher than in the summer months, suggesting mobilization of CPOM from debris dams as well as their potential transport from the floodplain. The downstream Hudson River did not show the same increase in CPOM transport during this month, but discharge in the Hudson River increases by only about 30% during a release compared to a 5–10-fold increase in discharge in the Indian River.
The Indian River showed a progressive increase in CPOM transport throughout the summer, which is not what one would expect in a frequently flooded river. Other studies have shown a decrease in POM transport in frequently flooded rivers because of a proposed exhaustion of organic matter entering from the floodplain and a flushing of easily mobilized benthic organic matter (Atkinson et al. Citation2009). We observed the opposite trend, but this is probably because most of the CPOM throughout the summer was a filamentous algal mat that was sloughed with each regulated release. The algal mats were common in low shear stress areas in the Indian River (Fuller et al. Citation2010, Citation2011) and persisted throughout the period of regulated releases; other rivers had lower nitrate concentrations than the Indian River which likely contributed to the dense algal mats that were found only in low shear stress areas in the Indian River (Fuller et al. Citation2011). Visual examination of POM in drift nets revealed primarily remnants of the filamentous algal mat throughout the summer in the Indian River, whereas other rivers had predominantly terrestrial debris, including seeds, flowers, catkins, and leaves. There was little evidence of filamentous algae in other rivers except at the downstream Hudson site during a release where filamentous algae were also observed but not in the same relative abundance as in the Indian River. Thus, our increased transport of CPOM during the summer in the Indian River was due to an autochthonous source of organic matter and did not occur because of allochthonous inputs.
We did not observe an exhaustion of organic matter in transport from either a benthic source or the floodplain in the Indian River despite the frequent regulated releases, suggesting that these releases have little impact on organic matter transport dynamics. However, during a large, natural flood, we did observe much lower FPOM and CPOM transport in the Indian River compared to other rivers, which does suggest a reduction in stores of organic matter versus those available in the other rivers. The Cedar and Boreas Rivers had much higher FPOM and CPOM transport during the flood than the Indian River, and the increase in POM transport during the flood in the downstream Hudson River was intermediate to these sites. Also, organic matter in transport in the Indian River during this natural flood was a combination of both autochthonous and allochthonous material in contrast to the leaves, sticks, and other allochthonous debris more commonly represented in the seston of the other rivers. The Indian and downstream Hudson Rivers had much lower per cent increases in POM transport during the floods than rivers with more natural hydrographs, suggesting that the regulated releases probably do decrease the availability of both benthic and floodplain POM for transport. Personal observations along the near-shore areas of the Indian River showed a distinct delineation of the high water mark present during releases which was marked by an absence of terrestrial debris from these near-shore areas.
In summary, we observed a seasonal decrease in POM transport in rivers with natural hydrographs over the summer that followed natural decreases in discharge. In contrast, there was a continuous increase in CPOM transport during the recreational releases in the Indian River throughout the summer and into the autumn. Summer CPOM transport increases were largely from the sloughing of filamentous algae, whereas the autumnal peak in CPOM transport was a combination of both allochthonous and autochthonous sources. These increases in POM transport suggest a lack of exhaustion of resources from the recreational releases, but when POM transport during a natural flood is compared among rivers, it was clear that the sites experiencing the recreational releases had lower total amounts of POM in transport than rivers with natural hydrographs. These results suggest that frequent flooding from recreational releases does decrease availability of POM in medium-sized rivers, but autochthonous sources of organic matter in these more autotrophic systems may mask the true effects of frequent floods on transport of POM resources in these rivers.
Acknowledgements
The authors acknowledge the assistance of Dr B. Baldigo for his assistance in placement of a USGS gaging station on the Indian River and Independent Paper Inc., Glens Falls, New York, for providing access to the upstream Hudson River site. Funding was provided by NSF grant DEB 04150365. Additional funding was provided to RLF by the Colgate University Research Council and a University of North Carolina Junior Faculty award was conferred on MWD. Two anonymous reviewers provided helpful comments on an early version of this paper.
References
- Atkinson CL, Golladay SW, Opsahl SP, Covich AP. 2009. Stream discharge and floodplain connections affect seston quality and stable isotopic signatures in a coastal plain stream. Journal of the North American Benthological Society. 28:360–370.
- Benfield EF. 1997. Comparisons of litterfall input to streams. Journal of the North American Benthological Society. 16:104–108.
- Cuffney TD, Wallace JB. 1989. Discharge-export relationships in headwater streams – the influence of invertebrate manipulations and drought. Journal of the North American Benthological Society. 8:331–341.
- Fuller RL, Doyle S, Levy L, Owens J, Shope E, Vo L, Wolyniak E, Small MJ, Doyle MW. 2011. Impact of regulated releases on periphyton and macroinvertebrate communities: the dynamic relationship between hydrology and geomorphology. River Research and Applications. 27:630–645.
- Fuller RL, Griego C, Meuhlbauer JD, Dennison J, Doyle MW. 2010. Response of stream macroinvertebrates in flow refugia and high scour areas to a series of floods: a reciprocal replacement study. Journal of the North American Benthological Society. 29:750–760.
- Golladay SW. 1997. Suspended particulate organic matter concentration and export in streams. Journal of the North American Benthological Society. 16:122–131.
- Jakob C, Robinson CT, Uehlinger U. 2003. Longitudinal effects of experimental floods on stream benthos downstream from a large dam. Aquatic Sciences. 65:223–231.
- Jones JB, Smock LA. 1991. Transport and retention of particulate organic matter in two low-gradient headwater streams. Journal of the North American Benthological Society. 10:115–126.
- Minshall GW. 1978. Autotrophy in stream ecosystems. Bio Science. 28:767–771.
- Minshall GW, Cummins KW, Peterson RC, Cushing CE, Bruns DA, Sedell JR, Vannote RL. 1985. Developments in stream ecosystem theory. Canadian Journal of Fisheries and Aquatic Sciences. 42:1045–1055.
- Naiman RJ. 1982. Characteristics of sediment and organic carbon export from pristine boreal forest watersheds. Canadian Journal of Fisheries and Aquatic Sciences. 39:1699–1718.
- Naiman RJ, Sedell JR. 1979. Characterization of particulate organic matter transported by some Cascade Mountain streams. Journal of the Fisheries Research Board of Canada. 36:17–31.
- Nakajima, T, Asaeda T, Fujino T, Nanda A. 2006. Coarse particulate organic matter distribution in the pools and riffles of a second order stream. Hydrobiologia. 559:275–283
- Neatrour MA, Webster JR, Benfield EF. 2004. The role of floods in particulate organic matter dynamics of a southern Appalachian river–floodplain ecosystem. Journal of the North American Benthological Society. 23:198–213.
- Pozo J, Elosegui A, Basaguren A. 1994. Seston transport variability at different spatial and temporal scales in the Aguera watershed (North Spain). Water Research. 28:125–135.
- Tank JL, Rosi-Marshall EJ, Baker MA, Hall RO. 2008. Are rivers just big streams? A pulse method to quantify nitrogen demand in a large river. Ecology. 89:2935–2945.
- Tank JL, Rosi-Marshall EJ, Griffiths NA, Entrekin SA, Stephen ML. 2010. A review of allochthonous organic matter dynamics and metabolism in streams. Journal of the North American Benthological Society. 29:118–146.
- Thomas SA, Newbold JD, Monaghan MT, Minshall GW, Georgian T, Cushing CE. 2001. The influence of particle size on seston deposition in streams. Limnology and Oceanography. 46:1415–1424.
- Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences. 37:130–137.
- Wallace JB, Ross DH, Meyer JL. 1982. Seston and dissolved organic carbon dynamics in a southern Appalachian stream. Ecology, 63:824–838.
- Wallace JB, Webster JR. 1996. The role of macroinvertebrates in stream ecosystem function. Annual Review of Entomology. 41:115–139.
- Wallace JB, Whiles MR, Eggert S, Cuffney TF, Lugthart GJ, Chung K. 1995. Long term dynamics of coarse particulate organic matter in three Appalachian mountain streams. Journal of the North American Benthological Society. 14:217–232.
- Webster JR, Benfield EF, Ehrman TP, Schaeffer MA, Tank JL, Hutchens JJ, D’Angelo DJ. 1999. What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. Freshwater Biology. 41:687–705
- Webster JR, Golladay SW, Benfield EF, D’Angelo DJ, Peters GT. 1990. Effects of forest disturbance on particulate organic matter budgets of small streams. Journal of the North American Benthological Society. 9:120–140.
- Webster JR, Meyer JL. 1997. Organic matter budgets for streams: a synthesis. Journal of the North American Benthological Society. 16:141–161.
- Webster JR, Wallace JB, Benfield EF. 1995. Organic processes in streams of the eastern United States. In: Cushing CE, Cummins KW, Minshall GW, editors. River and stream ecosystems: ecosystems of the world, volume 22. New York (NY): Elsevier. p. 117–187.
- Wipfli MS, Richardson JR, Naiman RJ. 2007. Ecological linkages between headwaters and downstream ecosystems: transport of organic matter, invertebrates, and wood down headwater channels. Journal of the American Water Resources Association. 43:72–85.