ABSTRACT
Despite the fact that fossil crocodylians have been recovered from the Panama Canal Zone starting with initial excavations in 1912, detailed studies have been lacking. Recent excavations of the canal have resulted in new discoveries of many vertebrate fossils, including the first known Miocene crocodylian skulls from Central America. These fossil skulls from the early–middle Miocene represent two new taxa with distinct morphology that is shared with extinct and extant caimans (Caimaninae). A cladistic analysis of 32 alligatorid and three outgroup taxa, scored for 75 characters, resulted in 1210 equally most parsimonious cladograms, all of which suggest that Culebrasuchus mesoamericanus, gen. et sp. nov., is the sister taxon to all previously known Caimaninae. Additionally, the analysis suggests that Centenariosuchus gilmorei, gen. et sp. nov., is the sister taxon to a caimanine clade that includes Purussaurus from the Miocene of South America. In fact, teeth very similar to those of Purussaurus have also been recovered from the Panama Canal. Given these South American affinities, we suggest that these early caimanines dispersed across saltwater. This is a potentially surprising result, because all extant alligatorids lack the salt glands that would have been necessary for the marine dispersal required to reach Central America during the Miocene. Unlike Miocene mammals that all have North American affinities, the Miocene crocodylians of Panama represent a ‘melting pot’ with taxa of disparate origins living together at the southern extreme of Central America.
SUPPLEMENTAL DATA—Supplemental materials are available for this article for free at http://www.tandfonline.com/UJVP
INTRODUCTION
The fossil record of crocodylians in Central America is limited, with only five known sites. All of the four non-Panamanian fossil crocodylian localities of Central America are younger than 6 Ma. Previously, the oldest records were from the upper Miocene or early Pliocene deposits of southeastern Costa Rica and attributed to Gavialosuchus americanus (Laurito and Valerio, Citation2008), providing the first evidence of this otherwise North American species in Central America. In addition, fossils including crocodylian crania and osteoderms were recovered from a late Pliocene–early Pleistocene site in El Indio, Costa Rica, which were attributed to extant Crocodylus (Mead et al., Citation2006). Additionally, a partial maxilla and isolated teeth from the early–middle Pleistocene were attributed to Crocodylus acutus from El Salvador (Cisneros, 2005), and a maxilla from the middle Pleistocene of Guatemala was described as a new subspecies of the extant Crocodylus moreletii (Mook, Citation1959). As such, most of what should arguably be a rich history of Central American crocodylians has been poorly or completely unknown from the fossil record, especially prior to the late Pliocene.
Previously, fossil crocodylians from Panama have only been mentioned in the literature (Whitmore and Stewart, Citation1965; MacFadden, Citation2006; Kirby et al., Citation2008) and very briefly described (Gilmore, Citation1912; Gillette, Citation1984; Hastings et al., Citation2009), but never figured or placed in any taxonomic context. Fossil crocodylians were first discovered in Panama during the canal's excavation in 1912. With recent excavations associated with expansion of the canal, new fossils have been recovered by crews from the Smithsonian Tropical Research Institute (STRI) and the Florida Museum of Natural History (FLMNH), in collaboration with the Panama Canal Authority (ACP). The most notable finds include two new fossil crocodylian skulls, the first and oldest known thus far from Central America. Each represents a new species from different parts of caimanine evolutionary history, and provides new insight into the origin and dispersal capabilities of this group. Here we describe and place within temporal and biogeographic contexts the Miocene crocodylian collections of the Panama Canal Zone made over the last century.
Crocodylian fossils have been recovered from four different formations within the Canal Zone (). These include the late Oligocene/early Miocene Las Cascadas Formation, early Miocene Culebra Formation, early or middle Miocene Cucaracha Formation, and the late Miocene Gatun Formation.
FIGURE 1 Fossil crocodylian localities (black circles) in the Panama Canal Zone, Panama, as represented by both a paleomap (modified from CitationMontes et al., 2012) and modern political map.
Although it has long been known that Panama was not connected to South America during the Miocene, it has recently been suggested that only a narrow gap, approximately 200 km wide, separated it from South America (Farris et al., Citation2011; CitationMontes et al., 2012), contradicting earlier views of a much larger gap during the early Miocene between Panama and South America (Coates et al., Citation2004). Despite this new evidence for a narrower seaway, the mammalian faunas of these formations have distinctly North American affinities (MacFadden, Citation2006).
Institutional Abbreviations
ChM, Charleston Museum, Charleston, South Carolina; SMU, Southern Methodist University, Shuler Museum of Paleontology, Dallas, Texas; UF, University of Florida, Florida Museum of Natural History, Gainsville, Florida; USNM, United States National Museum, Smithsonian Institution, Washington, D.C.
Terminology and Abbreviations
Teeth and alveoli are abbreviated with ‘pm’ for the premaxilla, ‘m’ for the maxilla, and ‘d’ for the dentary and with numbers indicating position in the jaw, 1 being the most anterior. For example, the first tooth/alveolus of the maxilla is ‘m1’ and the fourth tooth/alveolus of the dentary is ‘d4.’ Figures in Supplement Data (available online at www.tandfonline.com/UJVP) have the prefix ‘S.’
SYSTEMATIC PALEONTOLOGY
CROCODYLIA Gmelin, Citation1789
ALLIGATORIDAE Gray, Citation1844
CAIMANINAE Brochu, Citation2003 (following Norell, Citation1988)
CULEBRASUCHUS MESOAMERICANUS, gen. et sp. nov.
(–)
Holotype and Only Known Specimen
UF 244434, skull including left jugal, maxilla, partial premaxilla, right lacrimal and partial prefrontal, nearly complete left dentary with partial angular and surangular, anterior right dentary and partial right maxilla, braincase, 20 teeth in articulation, two additional associated teeth, and three cervical vertebrae.
Locality and Horizon
Type locality is within the Panama Canal Zone at the El Lirio Norte site (9.0535°N, 79.6571°W). The holotype was recovered from the upper part of the Culebra Formation, and is likely early Miocene in age (19.83–19.12 Ma).
Etymology
Genus named after the Culebra Formation (culebra is Spanish for snake) from which the holotype and only known fossil was discovered and ‘suchus’ for crocodile. Species name for Central America (‘meso’ from Greek ‘mesos’ for middle or central and ‘americanus’ meaning from America), the region where the holotype and only known fossil was discovered.
Diagnosis
Uniquely possesses in combination: well-exposed supraoccipital on the dorsal surface that excludes the parietal from the posterior margin of the skull table; surangular-angular suture that meets at the posterior rim of the external mandibular fenestra; linear dentary from d4 to the posterior-most alveolus; enlarged external mandibular fenestra through which the foramen intermandibularis caudalis is visible; flat, not upturned, medial orbital margin; a frontal that lacks anteroposteriorly oriented ridges; splenial that ceases anteriorly dorsal to the Meckelian groove, without entering the symphysis; wide supratemporal fenestrae that have only a minor overhang along the medial wall; exoccipitals that terminate dorsal to the basioccipital tubera; medial eustachian foramen with deeply divergent anterior and posterior branches; m3 the largest maxillary tooth/alveolus; and d4 tooth that occludes within a pit in the maxillary.
Description
Cranial Openings
A small portion of the left margin of the external nares is preserved in the holotype of Culebrasuchus mesoamericanus (UF 244434; ). The margin is oriented dorsally, rather than anterodorsally, resulting in an upward-facing nostril (Fig. S1). Part of the lateral margin of the left orbit and the anterior margin of the right orbit are also preserved (). The anterior margin of the orbit is transversally oriented, and lacks forward indentation. No portion of the suborbital fenestrae or choanae is preserved.
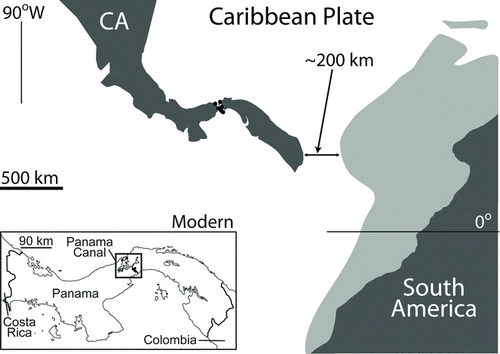
FIGURE 2 Holotype of Culebrasuchus mesoamericanus, gen. et sp. nov. (UF 244434), from the early Miocene Culebra Formation of Panama. A and B, lateral view of the rostrum and rear mandible; C and D, dorsal view of the rostrum and rear mandible. Abbreviations: an, angular; d, dentary; d4, fourth dentary tooth; dt, dentary tooth; emf, external mandibular fenestra; f, frontal; fic, foramen intermandibularis caudalis; j, jugal; m, maxilla; m3, third maxillary tooth; m7–9, seventh through ninth maxillary teeth; m10–12, tenth through twelfth maxillary teeth; nf, nasal fragment; pm, premaxilla; qj, quadratojugal; sa, surangular. Scale bar equals 10 cm.
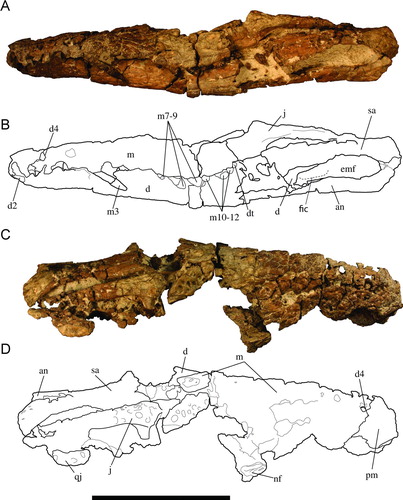
FIGURE 3 Holotype of Culebrasuchus mesoamericanus, gen. et sp. nov. (UF 244434), from the early Miocene Culebra Formation of Panama. A, skull table and interorbital region in dorsal view; B, line drawing of image in A. Abbreviations: f, frontal; fps, frontoparietal suture; lc, lacrimal; or, orbit; p, parietal; prf, prefrontal; q, quadrate; so, supraoccipital; sq, squamosal; stf, supratemporal fenestra. Scale bar equals 10 cm.
The ventral margin of the left infratemporal fenestrae is preserved in UF 244434 and is bound by the jugal (). This fenestra is relatively large and anteroposteriorly elongate. The supratemporal fenestrae are relatively robust, and roughly circular in shape (). The fenestrae are preserved well along the rest of the margins, and are medially and posteromedially bound by the parietal, and at least posterolaterally bound by the squamosals. The anteromedial corner of the fenestrae is smooth, lacking any fossa. The medial parietal wall is imperforate, lacking foramina (Supplementary Fig. S1).
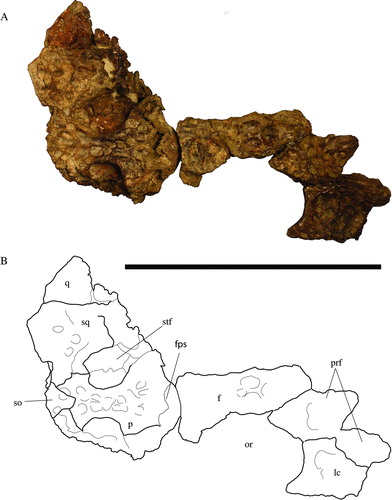
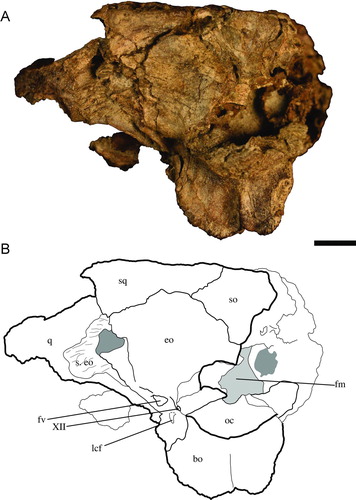
The foramen magnum is bound almost entirely by the exoccipitals (). Only the ventral-most margin is bound by the basioccipital. The dorsal lip of the exoccipital that forms the foramen vagus is preserved well and is rather robust in UF 244434. Two small foramina, present between a foramen vagus and the occipital condyle, are separated by small, vertically oriented bridges that are presumably for anterior and posterior passages of cranial nerve XII (hypoglossal nerve). Whereas the anterior foramen is bound entirely by the exoccipital, the posterior foramen is bound dorsally by the exoccipital and ventrally by the basioccipital.
FIGURE 4 Holotype of Culebrasuchus mesoamericanus, gen. et sp. nov. (UF 244434), from the early Miocene Culebra Formation of Panama. A, skull in occipital view; B, line drawing of skull. Abbreviations: bo, basioccipital; eo, exoccipital; fm, foramen magnum; fv, foramen vagus; lcf, lateral carotid foramen; oc, occipital condyle; q, quadrate; so, supraoccipital; s. eo, suture for exoccipital; sq, squamosal; XII, foramen for cranial nerve XII. Scale bar equals 1 cm.
Premaxilla
Part of the left premaxilla is preserved in UF 244434 (). The premaxilla-maxilla suture extends along the dorsal surface and passes anterior to the perforation for articulation of the enlarged fourth dentary tooth. A well-preserved alveolus for pm5 would have supported a relatively reduced tooth. Another pit is located between pm3 and pm4 for occlusion of d2.
Maxilla
Most of the left maxilla and part of the right maxilla are preserved in UF 244434. They are dorsally pitted and poor preservation obscures contacts with the nasal and lacrimal. In lateral view, the surface of the rostrum curves smoothly from the orbits to the tip of the snout (), with no hint of a medial dorsal boss. The maxilla has a straight lateral margin (). Although the exact nature of the maxillary-jugal contact is obscured by deformation, the maxilla appears to have contacted the jugal posteriorly. The largest maxillary alveolus is that of m3, as shown by both the preserved alveoli and the enlarged tooth preserved in this position (, S1). The alveoli gradually decrease in size posterior to the third tooth position (Fig. S1). Although only five empty alveoli and seven containing teeth are preserved in UF 244434, the posterior end of the maxilla is not fully preserved, so there may have been more. The maxillary toothrow is almost straight with a slight medial bend anteriorly (Fig. S1).
Lacrimal
A portion of the right lacrimal is preserved in UF 244434 and is roughly triangular in shape, tapering anteriorly (). The anterior-most tip of the lacrimal is not preserved, but what is there strongly indicates that the lacrimal would not have extended as far anterior as the prefrontal. Although not certain, the preserved portion also seems to indicate that the lacrimal would not have a strongly indented maxillary suture. The lacrimal makes up part of the anterior margin of the orbit and is transversally oriented, without an anteriorly directed indentation. The lacrimal does not appear to contact the nasal and lacks any hint of canthi rostralii.
Prefrontal
The right prefrontal and its attachment with the lacrimal are preserved in UF 24434 (). The anterior-most extent is not preserved, and thus its relative length as compared with the frontal, nasal, and lacrimal cannot be determined. The dorsal portion of the prefrontal pillar is present on the ventral side, expands dorsally, and is solid without a prefrontal recess (Fig. S1). The prefrontal-lacrimal contact is oriented anterolaterally-posteromedially. Although the prefrontals are at least separated by the frontal posteriorly, the contact with the nasal is not preserved. The prefrontal has a subtle, semicircular ridge in the anteromedial corner of the orbit ().
Jugal
The intersection of sutures for the jugal, lacrimal, and maxilla forms an anteriorly pointing triangle. The postorbital bar is slender and inset medially from the lateral surface of the jugal (). The contact with the quadratojugal is not preserved. The jugal bar is thin and elongate without strong curvature along the dorsal margin.
Frontal
The medial orbital margins of the frontal are not upturned, creating a relatively flat plane along the dorsal surface (, S1). Pitting is shallow and wide and continuous up to the edge of the orbits, without any notable ridges.
Parietal
Most of the central, planar skull table is composed of the parietal (). The medial supratemporal fenestral walls are indented toward the midline, and the dorsal lip is robust and relatively smooth (Fig. S1). Although the supraoccipital-squamosal suture excludes the parietal from contact with the posterior margin of the skull table, it does come very close to the posterior margin, ending less than 4 mm from the edge.
Supraoccipital
The supraoccipital is broadly exposed on the dorsal skull surface and is triangular in shape (Fig. S1). Exposure of the supraoccipital on the occipital surface is also triangular and much larger than its dorsal exposure and the lateral suture with the squamosal does not reach the dorsal surface of the skull table ().
Laterosphenoid
In UF 244434, the anterior face of the braincase is formed by the left and right laterosphenoids. In dorsal view, the laterosphenoid contacts the parietal. Because only the dorsal portion of the lateral laterosphenoid bridge is preserved, the pterygoid participation to the ventral base is unknown.
Squamosal
The posterolateral section of the skull table and the posterolateral margin of the supratemporal fenestra are formed by the squamosal (). Both the squamosal-supraoccipital and squamosal-parietal sutures are short on the dorsal surface of the skull. The squamosal and supraoccipital exclude the parietal from the posterior margin of the skull table for a very short distance. The squamosal and parietal meet along the posterior wall of the supratemporal fenestrae. The lateral surface is not preserved well enough to determine if the groove for the external ear valve musculature would have flared anteriorly. The posterior margin of the otic aperture is invaginate.
Exoccipital
Most of the occipital surface of the cranium and nearly the entire margin of the foramen magnum are formed by the exoccipitals. The exoccipital-basioccipital suture is well preserved on the left side and clearly shows that the exoccipital terminates dorsal to the basioccipital tubera (Fig. S1). The lateral carotid foramen opens lateral to the basisphenoid.
Basioccipital
Most of the occipital condyle is formed by the basioccipital. The basioccipital tubera flare out laterally with a strong vertically oriented ridge extending dorsally from their ventral meeting point ().
Basisphenoid
Located between the basioccipital and the preserved portions of the laterosphenoid. The basisphenoid is largely obscured from lateral view by the overlapping laterosphenoid, and appears thin ventral to the basioccipital and is not exposed extensively anterior to the foramen ovale. A bridge that forms the division between anterior and posterior branches of the medial eustachian foramen is not evident, suggesting that it is located deep within the foramen beyond the area that has been prepared so far (Fig. S1).
Pterygoid
An isolated partial right pterygoid was recovered with the skull, although its point of attachment to the braincase was not preserved. It includes a well-developed torus transiliens that has a triangular swelling toward the anterior end. The dorsal surface is concave and the broad ventral surface is convex with a large articulation for the ectopterygoid.
Quadratojugal
A small portion of the left quadratojugal is preserved in place (), but its contact with the jugal is not preserved. The lateral surface has similar wide pitting as that of the jugal.
Quadrate
A portion of the left quadrate is preserved. Its ventral surface does not have the expected linear crests for attachment of jaw musculature. Even allowing for preservational effects, the crests must have been severely reduced to account for the smooth ventral surface present on UF 244434.
Dentary
At least 10 partial to complete teeth were preserved in place, as well as one other empty alveolus, in the left dentary, and three alveoli with teeth and three alveoli with partial/insipient teeth are present on the right side of UF 244434 (). Although uncertain, because the posterior portion is obscured, the dentary tooth count was likely greater than 11. A small sliver of an anterodorsally projected (not procumbent) d1 is preserved within the anterior part of the dentary. The d4 is much larger than d3, and the alveoli are not confluent. The dentary is straight from d4 to the posterior-most tooth position (Fig. S1). A portion of the dentary symphysis was also preserved, but the number of teeth participating is unclear. The dentary forms the anterior margin of a large external mandibular fenestra.
Splenial
The posterior part of the left splenial preserves a smooth medial surface. The anterior portion of the right splenial preserves its rostral-most tip, which does not contact the symphysis, and instead comes to a point slightly posterior to the symphysis, dorsal to the meckelian groove (Fig. S1).
Angular
The angular-surangular suture meets at the posterior margin of the large external mandibular fenestrae (EMF). The angular forms the ventral half of the EMF and extends ventrally along the mandible. The angular joins the dentary at the anteroventral corner of the EMF. The dorsal extension bracketing the foramen intermandibularis caudalis is missing in UF 244434 and thus its anterior extent relative to the ventral extension cannot be assessed. Supporting rock matrix currently fills the EMF, but preservation on the medial side of the mandible clearly indicates that the foramen intermandibularis caudalis would be visible through the EMF in lateral view ().
Surangular
Extends from the posterior margin to the anterior one-third of the dorsal margin of the EMF (Fig. S1). No parts of the retroarticular process or glenoid fossa are preserved.
Maxillary Dentition
A greatly enlarged m3 is straight, with well-defined carinae forming distinct lingual and labial surfaces. The crown of m7 is reduced and straight, in contrast to those of m8 and m9, which are much more rounded and spade-shaped, with crenulations near the apex (). The crowns of m8–9 also have well-defined carinae. The crowns of m10–12 are progressively blunter and more rounded.
Dentary Dentition
The crowns of the dentary teeth are similar to those of the maxilla in being sharp and pointed anteriorly, grading into progressively blunter, rounded crowns posteriorly. The crowns of d2–5 are long, thin, and dorsally oriented. The posterior-most dentary tooth preserved (position uncertain) is very blunt and rounded, and anteroposteriorly elongated.
Cervical Vertebrae
Three cervical vertebrae were recovered in articulation in UF 244434 and are likely cervicals 4–6 (Fig. S1). The neurocentral sutures of these vertebrae are partially fused. No centrum was preserved with cervical 6. The prezygapophyses are dorsoventrally tall and have a broad ventral contact with the centrum.
Comparisons
Cranial and Mandibular Openings
Although only a small portion of the external nares of Culebrasuchus mesoamericanusis preserved, it does not appear to have the overly enlarged and widened aperture seen in Purussaurus mirandai (Aguilera et al., Citation2006). Like that of all caimanines, the external nares of C. mesoamericanus are oriented dorsally (Fig. S1), as opposed to anterodorsally as in basal alligatorines (Brochu, Citation1999).
The supratemporal fenestrae of Culebrasuchus have dermal bones that do not overhang the rim at maturity (, S1), as for most caimanines (Brochu, Citation1999). The same condition as Culebrasuchus was scored for Eocaiman cavernensis (Brochu, Citation1999), and Brochu (Citation2010) mentioned that E. cavernensis likely had wide fossae, distinguishing it from Tsoabichi greenriverensis and all other caimanines for which this character is known. Unlike extant Caiman and Melanosuchus that have a foramen in the medial parietal wall of the supratemporal fenestra (Brochu, Citation1999), the wall is imperforate in Culebrasuchus (, S1), making it similar to that of Alligatorinae (Brochu, Citation1999).
Culebrasuchus mesoamericanus is similar to derived Alligator in having a very large EMF and, when the medial and lateral sides of the preserved left rear jaw are compared, a laterally visible foramen intermandibularis caudalis (FIC) (). In contrast, all caimanines except Purussaurus neivensis have a reduced EMF (Brochu, Citation1999). The EMF is preserved in E. cavernensis and was coded with the caimanine condition of a smaller EMF through which the FIC is not visible laterally (Brochu, Citation1999). The EMF is preserved in E. palaeocenicus, but the FIC is not, so this character cannot be assessed. Unlike that of Allognathosuchus polyodon, Allognathosuchus mooki, and Procaimanoidea kayi, Culebrasuchus has a surangular-dentary suture that intersects the EMF anterior to the posterodorsal corner of the EMF, as in the rest of Alligatoridae.
Premaxilla
Due to the incomplete anterior end of the skull of Culebrasuchus, we could not assess the number of premaxillary alveoli to know whether it had only four teeth as in Paleosuchus, or five as in all other alligatorids (Brochu, Citation1999).
Maxilla
The maxilla of Culebrasuchus mesoamericanus is dorsoventrally flat, not altirostral as in both species of Paleosuchus, and lacks the canthi rostralii characteristic of Caiman lutescens, Caiman latirostris, and both species of Melanosuchus (Brochu, Citation1999). The pit for the occlusion of the left fourth dentary tooth is worn through completely such that the tooth occludes through the dorsal side in UF 244434. Thus, Culebrasuchus shows a well-developed pit for occlusion that it shares in common with all other Alligatoridae. The largest maxillary alveolus in Culebrasuchus is the m3 (, S1). In nearly all other alligatorids, it is instead m4, with the only exception being the homodont teeth of the extinct caimanine Mourasuchus. Like most caimanines, Culebrasuchus has all dentary teeth occluding lingual to the maxillary tooth row (Brochu, Citation1999). The exceptions to this are Caiman yacare and Caiman crocodilus, which have an occlusal pit between m7 and m8 (Brochu, Citation1999).
Prefrontal
The prefrontal pillar of Culebrasuchus mesoamericanus is solid (Fig. S1), rather than containing a large pneumatic sinus as in Alligator mississippiensis and Alligator mefferdi. Culebrasuchus has a modest, semicircular prefrontal ridge, comparable to that seen in Caiman yacare and Alligator olseni. This ridge contrasts with the very prominent ridge seen in Mourasuchus (Brochu, Citation1999).
Frontal
The medial margin of the orbit of Culebrasuchus mesoamericanus, constituted by the frontal, is flat and not upturned as in all Caiman and Melanosuchus species (Fig. S1; Brochu, Citation1999). The flat margin is similar to that of Stangerochampsa and Brachychampsa as well as basal alligatorines (Brochu, Citation1999). The dorsal surface of the frontal of C. mesoamericanus lacks the anteroposteriorly oriented ridges between the orbits that are characteristic of Tsoabichi greenriverensis (Brochu, Citation2010).
Supraoccipital
Dorsal exposure of the supraoccipital on the skull table, excluding the parietal from contact with the posterior margin, is similar to that of extant Caiman and Melanosuchus (Brochu, Citation1999). The degree to which the parietal is excluded in Culebrasuchus mesoamericanus is small relative to that of extant Caiman, in that the parietal comes within 4 mm of contacting the posterior margin. Conversely, all alligatorines have minimal dorsal exposure of the supraoccipital on the skull table, and in most species it is completely restricted to the occipital surface (Brochu, Citation1999). The condition in Culebrasuchus is intermediate between the partial inclusion of the parietal as in Tsoabichi greenriverensis, and the complete exclusion seen in extant Caiman (Brochu, Citation2010).
Squamosal
Culebrasuchus mesoamericanus has a broad squamosal-parietal contact within the supratemporal fenestrae, similar to all known alligatorids (Brochu, Citation1999). Conversely, these bones in the non-alligatorid Brachychampsa montana do not come in contact at all within the fenestrae (Brochu, Citation1999).
Exoccipital
Culebrasuchus mesoamericanus is similar to all alligatorines, Stangerochampsa, and Brachychampsa (Brochu, Citation1999) in having exoccipitals that terminate dorsal to the basioccipital tubera (Fig. S1). This is quite different from the long, ventrally directed descending processes that appear in all caimanines (Brochu, Citation1999, Citation2010).
Basioccipital
Culebrasuchus mesoamericanus is similar to Alligator mississippiensis in having basioccipital tubera that flare out laterally, not as ventrally directed as those of extant Caiman and Paleosuchus.
Basisphenoid
The divide between anterior and posterior branches of the medial eustachian foramen in Culebrasuchus mesoamericanus must occur deep within the foramen as in extant Alligator. No hint of the transversally oriented bridge is apparent in Culebrasuchus as is often the case for extant Caiman and Paleosuchus (Brochu, Citation1999).
Dentary
The straight condition of the dentary of Culebrasuchus mesoamericanus from the d4 alveolus to the posterior-most tooth position (Fig. S1) is different from the gently curved condition seen in all extant caimanines, both species of extant Alligator, Purussaurus, Eocaiman, and Necrosuchus (Brochu, Citation1999). The only alligatorid that shares this condition is Mourasuchus, which has much more elongated and ‘U’-shaped dentaries, but are nonetheless much closer in their shape than either is to any other alligatorid.
Splenial
The splenial of Culebrasuchus mesoamericanus clearly does not extend into the symphysis and stops anteriorly, dorsal to the Meckelian groove (Fig. S1). This is similar to that seen in all of Caimaninae, for which the character is known, as well as extant Alligator and Alligator mefferdi (Brochu, Citation1999).
Angular
In contrast to the condition found in nearly all caimanines, the angular-surangular suture of Culebrasuchus mesoamericanus does not pass broadly along the ventral margin and instead meets the external mandibular fenestra at the posterior angle (Fig. S1). The condition of Culebrasuchus is instead similar to that of all alligatorines, the extinct caimanine Mourasuchus, Stangerochampsa, and Brachychampsa (Brochu, Citation1999).
Dentition
The overall shape, carinae, subtle dorsoventral ribbing, and posteriorly increasing bluntness of the tooth crowns of Culebrasuchus mesoamericanus are very similar to those of extant Caiman and Paleosuchus.
CENTENARIOSUCHUS GILMOREI, gen. et sp. nov.
(, )
Holotype
UF 262800, partial skull including braincase, dorsal skull table, both quadrate condyles, frontal, and two isolated teeth. Also associated were a cervical vertebra and two partial osteoderms. This specimen was collected from the same locality as the paratype, but two years later in March 2011 by U. Denetclaw.
Paratype
UF 245503, left premaxilla and partial right maxilla, including five teeth in situ. The specimen was collected by A. Rincon in April 2009. Given the close proximity and similar size, it is likely that the skull and snout were associated, but due to the lengthy period between collections and slightly different lithology, they have been given different catalogue numbers.
Type Locality and Horizon
The type fossils were recovered from the lower part of the early or middle Miocene Cucaracha Formation at the Hodges Microsite at the Hodges Hill locality within the Panama Canal Zone, Panama (9.04768847°N, 79.653814°W). At the locality, the upper part of the Cucaracha Formation is characterized by red, green, and purple mudstones interbedded with coarse- to medium-grained lithic sandstones and volcaniclastic pebble conglomerates. The fossiliferous lenticular layer is about 1 m thick, and is composed by normally graded, poorly to subrounded matrix-supported granule conglomerates. The two type fossils (UF 245503 and UF 262800) were collected along the base of this layer, where lenticular centimetric layers of olive-gray claystone granules are commonly present. The distinctive preservation of the fossil remains, which is characterized by poorly transported cranial material with no evidence of intense abrasion, as well as the texture of the basal conglomerates suggest sporadic development of low traction deposits at the base of the fossiliferous layer, likely representing sporadic crevasse channel deposits.
Diagnosis
Centenariosuchus gilmorei differs from Culebrasuchus mesoamericanus in having m4 as the largest maxillary alveolus, upturned medial orbital margins, exoccipitals with ventrally directed processes that reach the basioccipital tubera, a foramen in the medial wall of the supratemporal fenestra, and constricted supratemporal fenestrae. It differs from Eocaiman cavernensis in having a straight lateral margin of the suborbital fenestra and constricted supratemporal fenestrae. It differs from Tsoabichi greenriverensis in having a naris that projects dorsally, a large supraoccipital dorsal exposure, and upturned medial orbital margins. It differs from Paleosuchus spp. in having a premaxilla with five teeth, an incisive foramen between the first two premaxillary teeth, and a large supraoccipital dorsal exposure. It differs from Purussaurus spp. in having a small, narrow incisive foramen. It differs from Mourasuchus spp. in having a circular naris and enlarged m4. It differs from Orthogenysuchus olseni in having a small, narrow incisive foramen and a circular naris. It differs from Melanosuchus spp. in the lack of vomer exposure on palate and the lack of canthi rostrali. It differs from Caiman latirostris and Caiman lutescens in lacking canthi rostrali. It differs from Caiman yacare and Caiman crocodilus in having dentary teeth that occlude lingual to the maxillary dentition. It further differs from all Caiman and Melanosuchus species in having a suborbital fenestra with a straight lateral margin.
Etymology
Genus name in honor of the centennial anniversary of the excavation of the Panama Canal (2014) and ‘suchus’ for crocodile. Species name for Charles W. Gilmore, the first to publish findings of crocodylian fossils within the Panama Canal in 1912.
Description
Premaxilla
Five distinct alveoli are present on the premaxilla (UF 245503; ). The first and second are by far the smallest and are separated by a large pit for occlusion of the first dentary tooth. The alveoli of pm3–5 are larger, with two small medial depressions for occlusion of d2–3 (). A small shelf posterior to the pm5 alveolus indicates a deep pit for occlusion of an enlarged tooth in the mandible, likely the d4. The incisive foramen is narrow and teardrop-shaped, consisting of less than 25% of the total transverse width, and appears to bisect the left and right pm1 alveoli. The external nares face dorsally and are concave posteriorly, forming a medial point where sutures indicate it likely joined the nasal (). The ventral posteromedial margin forms a right angle, suggesting that the vomer was not exposed on the palatal surface. In ventral view, the premaxilla-maxilla suture is roughly linear.
Maxilla
A large portion of the right maxillary tooth row is preserved in the paratype UF 245503. The premaxillary-maxillary suture clearly passes through a large depression for the occlusion of d4. The crowns of m1–4 increase in size posteriorly, with m4 being the largest in the preserved tooth row (). A total of 12 alveoli are preserved at least in part, and the maxilla likely possessed a total of 14 alveoli. Occlusal pits lingual to the maxillary alveoli indicate that the lower dentition occluded lingual to the upper dentition (). The anterior and lateral edges of the suborbital fenestra are preserved on UF 245506. The lateral margin of the suborbital fenestra is straight and not bowed medially. The articular surface for the jugal indicates a strong overlap ventral to the orbit of these two bones.
Frontal
The nearly complete frontal of UF 262800 clearly possesses upturned medial margins of the orbits. The prefrontal articular surface indicates that the frontal was excluded from the anterior margin of the orbit (). Although only a small portion of the frontoparietal suture is preserved, it was likely entirely on the dorsal skull surface. Not enough of the center of the suture is preserved to discern whether it was concavoconvex or linear.
Skull Table
Very flat with shallow consistent pitting (). The posterior margin is transversally straight. The supratemporal fenestrae of UF 262800 are severely constricted with strong dorsal overhang. Closure of the right fenestra is more complete than the left. The medial wall within the right fenestra has a small perforation. The fenestrae are bound medially by the parietal, laterally by the squamosal, and anterolaterally by the postorbital.
Parietal
The parietal is broad, flat, and well ornamented (). It contacts the squamosal at the posterior-most edge of the supratemporal fenestra. The parietal-supraoccipital suture is bowed slightly anteriorly, excluding the parietal from contact with the posterior edge of the skull table.
Squamosal
Left and right squamosals are well preserved in UF 262800 and make up the dorsal and posterior portions of the invaginate external otic apertures. The squamosal groove for external ear valve musculature does not flare anteriorly. Squamosals curve to posteriorly directed points () that descend posteroventrally to the squamosal-quadrate suture. The squamosal exposure is limited in occipital view ().
Postorbital
A small part of the left postorbital is preserved in UF 262800, including the dorsal portion of the postorbital bar. The postorbital participates with the anterodorsal corner of the infratemporal fenestra and contacts the quadrate.
Supraoccipital
The supraoccipital-squamosal suture is long and anteroposteriorly directed (). The supraoccipital is broadly exposed on the dorsal skull table surface, where it is deeply pitted. Its exposure on the occipital surface is roughly pentagonal, due to enlargement ventral to the dorsal suture with the squamosal (). It participates to the medial margins of the posttemporal fenestrae.
Exoccipital
This element is broadly exposed on the lateral surface and recessed anteriorly, forming a strong dorsal lip to the foramen magnum (). The exoccipitals terminate ventrally along the basioccipital tubera and contain the posterior carotid foramen and medially bound the foramen vagus.
Quadrate
Both quadrates are well preserved in UF 262800. The quadrate makes up the ventral floor of the external otic recess and has a broad contact with the squamosal. The foramen aerum is dorsally and medially placed above the quadratic condyle. The medial condyle of the quadrate is smaller than the lateral (). The quadrate constitutes the lateral margin of the foramen vagus. Ventrally, the quadrate possesses two strong crests (A’–A and B of Iordansky, Citation1973).
FIGURE 5 Holotype (UF 245503) and paratype (262800) of Centenariosuchus gilmorei, gen. et sp. nov., from the early–middle Miocene Cucaracha Formation of Panama. A and B, skull and snout in dorsal view; C and D, skull and snout in ventral view. Abbreviations: bo, basioccipital; bsp, basisphenoid; d1op, occlusal pit for first dentary tooth; eo, exoccipital; f, frontal; fo, foramen ovale; en, external nares; m, maxilla; m1–3, first through third maxillary teeth; m4, fourth maxillary tooth; mef, medial eustachian foramen; ncs, neurocentral suture; oc, occipital condyle; op, occlusal pits; or, orbit; p, parietal; pm, premaxilla; pm1–2, first and second premaxillary alveoli; pm3–5, third through fifth premaxillary alveoli; po, postorbital; q, quadrate; qc, quadrate condyle; so, supraoccipital; sof, suborbital fenestra; sq, squamosal; stf, supratemporal fenestra. Scale bar equals 10 cm.
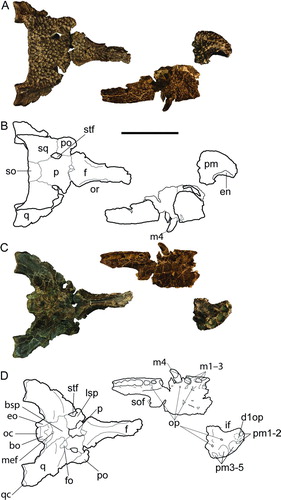
FIGURE 6 Centenariosuchus gilmorei, gen. et sp. nov., from the early–middle Miocene Cucaracha Formation of Panama. A and B, skull in occipital view (UF 262800); C and D, cervical vertebra in lateral view (UF 262800); E, osteoderm in dorsal, ventral, and lateral views (UF 262800); F, premaxilla in left lateral view (UF 245503). Abbreviations: acr, articular surface for cervical rib; bo, basioccipital; eo, exoccipital; fm, foramen magnum; fv, foramen vagus; en, external nares; hk, haemal keel; ncs, neurocentral suture; oc, occipital condyle; pozy, postzygapophyses; przy, prezygapophyses; q, quadrate; qc, quadrate condyle; so, supraoccipital; sq, squamosal. Scale bar for A–B equals 10 cm, for C–E equals 1 cm, for F equals 5 cm.
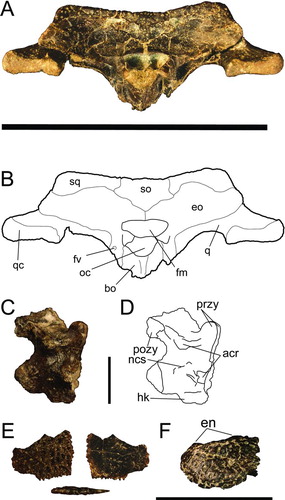
Basioccipital
The occipital condyle is caudally directed. The basioccipital tubera are reduced and narrow ventrally with the exoccipital flanges. A pronounced dorsoventral crest is present along the ventral occipital surface below the occipital condyle (). The basioccipital forms the posterior margin of the medial eustachian foramen.
Basisphenoid
Although the pterygoids were not preserved, their articular surfaces indicate that the basisphenoid was narrowly exposed between the basioccipital and pterygoids. The posterior margin of the medial eustachian foramen is composed of the basisphenoid (). The anterior and posterior branches of the medial eustachian foramen diverge deeply within this foramen.
Laterosphenoid
Most of the lateral braincase wall is formed by the laterosphenoid, which rises dorsally to meet the parietal, forming the medial margin of the foramen ovale and the imperforate anteroventral floor of the supratemporal fenestra (). The pterygoid-laterosphenoid suture is not preserved, so whether or not the pterygoid contributes to the lateral laterosphenoid bridge cannot be assessed.
Dentition
Preserved dentition includes right m1–4, an additional maxillary tooth (either m6 or m7), and two isolated teeth (). The tooth crowns are generally smooth, with carinae defining lingual and labial surfaces. They are generally spade-shaped and small, with the posterior-most being more rounded. One of the isolated teeth bears subtle striations and is more narrow and thin than the ones preserved in the maxilla.
Cervical Vertebra
The single complete cervical vertebra associated with UF 262800 bears a clear hypapophysis with articular surfaces for the cervical rib still present, likely indicating the eighth position behind the skull. The neurocentral suture is nearly fused ().
Osteoderm
A partial osteoderm associated with UF 262800 preserves one corner with a rectangular shape (). The pits are shallow and the bone is very thin (<2 mm thick). The osteoderm is flat without a bend or curve and is smooth and imperforate on the ventral side.
Comparisons
Premaxilla
As for most Alligatoridae, the premaxilla of Centenariosuchus gilmorei has five alveoli (). In contrast, Paleosuchus has only four (Brochu, Citation1999). The incisive foramen is narrow in Centenariosuchus, similar to that of all caimanines except Purussaurus spp. and Orthogenysuchus olseni, which have enlarged, wider foramina (Brochu, Citation1999). Furthermore, the incisive foramen of Centenariosuchus shows that the left and right pm1 alveoli were separated, as in most Caimaninae except Paleosuchus, which has a foramen that abuts the premaxillary teeth (Brochu, Citation1999). The preserved posteroventral portion of UF 245503 indicates that the vomer was not exposed on the palatal surface in Centenariosuchus (), much like all Caimaninae except Melanosuchus spp. (Brochu, Citation1999).
Maxilla
The lower teeth occlude lingual to the upper teeth in Centenariosuchus gilmorei () and most caimanines, with the exception of Caiman yacare and Caiman crocodilus, which possess interalveolar occlusal pits between at least one pair of adjacent maxillary teeth (Brochu, Citation1999). As in nearly all alligatorids, the largest maxillary alveolus of Centenariosuchus is m4, as opposed to m3 in Culebrasuchus. The number of maxillary teeth of Centenariosuchus (approximately 14) is comparable to that of all other alligatorids (14–16; Iordansky, Citation1973). The lateral edge of the suborbital fenestra is straight in Centenariosuchus and all caimanines except all species of Caiman, Melanosuchus, and Eocaiman (Brochu, Citation1999).
Frontal
The medial margins of the frontal of Centenariosuchus are upturned as in all Caimaninae except Tsoabichi greenriverensis (Brochu, Citation2010) and Culebrasuchus mesoamericanus.
Skull Table
The supratemporal fenestrae of Centenariosuchus are severely constricted with asymmetrical closure (), much like those of extant Caiman. This contrasts with the wide, open fenestrae of Culebrasuchus mesoamericanus and Eocaiman cavernensis (Brochu, Citation2010). A small perforation present in the medial wall of the supratemporal fenestra of Centenariosuchus is similar to that of extant Caiman, but not Culebrasuchus.
Parietal
The parietal-supraoccipital suture in Centenariosuchus gilmorei is similar to that of nearly all other caimanines in extending along the dorsal surface such that the parietal is excluded from the posterior wall of the skull table (). The exceptions are extant Paleosuchus and the Eocene T. greenriverensis (Brochu, Citation2010) in which the supraoccipital forms a narrow wedge that penetrates the parietal, but does not exclude it from the posterior wall.
Squamosal
As for other caimanines, the squamosal of Centenariosuchus has a strongly invaginate external otic aperture and pitting present along the entire dorsal surface. Both Centenariosuchus and extant Caiman have a relatively reduced groove for external ear valve musculature, as opposed to the dorsoventrally thickened groove of extant Paleosuchus. Occipital exposure of the squamosal in Centenariosuchus () and Paleosuchus differs from the broad exposure of extant Caiman.
Postorbital
The postorbital-squamosal suture of Centenariosuchus gilmorei and all Alligatoridae passes medially ventral to the skull table, and the postorbital contributes to the anterolateral corner of the supratemporal fenestra (Brochu, Citation1999).
Supraoccipital
The supraoccipital on the skull table is pentagonal and similar to that of most Caimaninae, except for the thin wedge present in Paleosuchus and Tsoabichi (Brochu, Citation2010). The extent of dorsal expression precludes the parietal fully, even more so than the exclusion seen in Culebrasuchus ().
Exoccipital
Much like all other caimanines, except Culebrasuchus mesoamericanus (, S1), the exoccipital of Centenariosuchus gilmorei sends a slender process ventrally, contacting the basioccipital tubera (, ). The well-defined dorsal lip of the foramen magnum formed by the exoccipitals is more consistent with that of Caiman than the more reduced lip of Paleosuchus.
Quadrate
The crests for articulation of jaw closure muscles present in UF 262800 (A’–A and B of Iordansky, Citation1973) are most similar to those of extant Caiman in height, length, and position on the quadrate (). In Paleosuchus, these ridges are far more reduced. Crest A’–A of Melanosuchus is well developed (more so than Caiman or Centenariosuchus) but crest B is reduced to a small rugosity.
Basioccipital
The long and thin dorsoventral crest present along the ventral occipital surface of the basioccipital of Centenariosuchus gilmorei () is very similar to that of extant Caiman. In contrast, Melanosuchus has a widened rugosity, not a ridge.
Basisphenoid
As in other caimanines, particularly the smaller Paleosuchus and Caiman, the anterior and posterior branches of the medial eustachian foramen diverge deeply in Centenariosuchus gilmorei.
Laterosphenoid
The lateral laterosphenoid bridge in extant caimanines, as well as the non-alligatorid Brachychampsa montana, is composed only of the laterosphenoid, whereas extant Alligator has an ascending process of the pterygoid that participates in the lateral laterosphenoid bridge (Brochu, Citation1999). The lateral bridge of UF 262800 looks to be exclusively laterosphenoid, but because the pterygoid was not preserved, the caimanine condition could not be confirmed.
Dentition
The recovered tooth crowns of Centenariosuchus gilmorei are spade-shaped, with clear anterior and posterior carinae. They are most similar to those of extant Paleosuchus, both in size and shape throughout the jaw. They differ from those of Melanosuchus niger in being much shorter and less pointed. They differ from those of Purussaurus spp. in having clear points, lacking blunted tips, being much more lingually compressed, and having well-defined carinae.
Cervical Vertebra
The cervical vertebra associated with UF 262800 bears a nearly closed neurocentral suture (), implying that the individual was very near morphological maturity (Brochu, Citation1996). As a result, the characters used to diagnose this new taxon are considered valid for an adult individual.
Osteoderm
The osteoderm associated with UF 262800 is small and flat, with no hint of a bend (). There is also no indication of the imbricating shelf typical of extant Paleosuchus. Among extant caimanines, the osteoderm of Centenariosuchus gilmorei most closely resembles those of Melanosuchus niger in general pitting pattern and thickness.
cf. CENTENARIOSUCHUS GILMOREI
Referred Specimen
UF 245593, nearly complete right angular. Specimen was collected at the Centenario Bridge locality (9.03011452°N, 79.636786°W), Cucaracha Formation, within the Panama Canal Zone, Panama.
Remarks
This specimen was found in the same formation, but at a different locality from the holotype and paratype of C. gilmorei (Fig. S2). The preserved margins of the external and internal mandibular fenestrae suggest a reduced external mandibular fenestra, as in all Caimaninae (Brochu, Citation2010). The external mandibular fenestra of Culebrasuchus and all of Alligatorinae is instead enlarged, such that the internal mandibular fenestra is visible (Brochu, Citation2010). Based on differences from the only other known caimanine from Panama, and the similarity in location and age, we here refer this angular as cf. Centenariosuchus gilmorei. We use the qualifier ‘cf.’ due to the lack of an angular in the holotype or paratype specimens.
cf. PURUSSAURUS sp. Barbosa-Rodrigues, Citation1892
()
Referred Specimens
UF 259879, isolated tooth from the early Miocene Culebra Formation of the Panama Canal Zone, Panama (9.02140203°N, 79.620369°W). UF 244335, isolated tooth from El Lirio Norte site also from the Culebra Formation (9.0535°N, 79.6571°W).
Description
These two isolated teeth are very distinct from the other teeth of the Panama Canal and Gavialosuchus (reported from Costa Rica by Laurito and Valerio, Citation2008; see below) in being larger, thicker, and blunter (). In fact, for the early Miocene tropics, these tooth crowns are indistinguishable from those described for Purussaurus, otherwise only known from South America (Aguilera et al., Citation2006). Because this taxon is only represented to date by two isolated teeth, its taxonomic affinities remain uncertain.
FIGURE 7 Teeth of cf. Purussaurus sp. from the Culebra Formation and Gavialosuchus americanus for comparison. A and B, UF 259879 in lingual and posterior views; C and D, UF 244335 in lingual and posterior views; E, Gavialosuchus tooth from Alachua County, Florida (UF 264830). Scale bar equals 1 cm.

EUSUCHIA Huxley, Citation1875
Incertae sedis
‘Headless Skeleton’
A ‘headless skeleton’ (USNM 23142) collected in 1962 and mentioned by Whitmore and Stewart (Citation1965) and Ferrusquía-Villafranca (Citation1977) has gone undescribed and unfigured ever since. It includes 10 vertebrae of varying completeness, eight partial osteoderms, possible rib fragments, unidentifiable bone, matrix, and at least one testudine plastral fragment mixed into the collection. These associated fossils were all collected from the Cucaracha Formation within the Panama Canal Zone, Panama (). The vertebrae are strongly procoelous and mostly represent posterior dorsals, close to the sacrum (Fig. S3). These wide vertebrae have large centra, indicating that the individual was large and mature. All vertebrae associated appear to pertain to a single individual.
The associated osteoderms are all relatively flat, with wide shallow pits (Fig. S4). There is no hint of a crest or a dorsoventrally angled bend to any of the osteoderms. Most have a rim of non-pitted surface around the rectangular edges. These osteoderms are not consistent with those of either Alligatoridae or Crocodylinae. They are, however, very consistent with those of Gavialosuchus americanus, from Florida (Fig. S4). It cannot be ruled out, however, that the osteoderms may instead belong to a gavialid and as such a definitive family-level classification cannot be determined. Due to the less diagnostic nature of crocodylian osteoderms and vertebrae, these associated fossils are not assigned beyond Eusuchia indet.
Culebra Block
An articulated series of three vertebrae with partial osteoderms was collected together within a large nodule in 1911 during the canal's initial excavation (Gilmore, Citation1912; USNM 7091; Fig. S5). These three posterior dorsal vertebrae are strongly procoelous and associated osteoderms are fragmentary, but appear flat with shallow, wide pitting. This specimen again is very similar to Gavialosuchus from Florida, but affinities with South American gryposuchines cannot be ruled out.
Vertebrae
Seventeen isolated vertebrae have been recovered from the Cascadas, Culebra, and Cucaracha formations (Fig. S5). These vertebrae range from very small, young individuals (centrum length: 23.2 mm) to very large (centrum length: 65.3 mm). Thirteen vertebrae are dorsals, two are cervicals, and at least two poorly preserved vertebrae are from the tail. All represent eusuchians with strongly procoelous centra.
Maxilla
An isolated partial maxilla (UF 244336) was discovered from the Culebra Formation, of the El Lirio Norte site (9.0535°N, 79.6571°W). This maxilla has part of the articulation surface for the jugal and represents a posterior section of the toothrow (Fig. S6). The four preserved partial teeth and alveoli are homodont and thus tooth position could not be discerned. The lateral surface is shallowly ornamented with small pits and gives no indication of canthi rostrali. The general size suggests that it belonged to a small individual.
Symphyseal Sections
Three dentary fragments from different individuals clearly indicate the presence of at least one longirostrine taxon in the early Miocene of Panama. Two of these (UF 244435 and 259873) have both left and right portions of the symphysis, and the third (UF 259875) represents a single right side, but with clear markings for symphyseal attachment (). None of the specimens include a splenial, indicating that they come from the anterior part of the rostrum. UF 259873 has no clear suture for the fusion of left and right dentaries. A probable suture extends along the midline both dorsally and ventrally in UF 244435, likely indicating the fusion of left and right dentaries. In all three, interalveolar length is comparable to alveolar length. Alveoli are facing anterolaterally in all specimens.
FIGURE 8 Symphyseal segments of longirostrine crocodylians recovered from the Culebra and Cucaracha formations within the Panama Canal Zone. A, fused dentary symphysis (UF 259873) in dorsal and lateral views; B, partial right dentary (UF 259875) from symphyseal portion of mandible in dorsal, lateral, and medial views; C, fused dentary symphysis (UF 244435) in dorsal and lateral views. Arrows on left side indicate anterior and posterior directions. Scale bar equals 5 cm.

The dentary fragments differ from those of the longirostrine Charactosuchus in having alveoli with low walls, close to the mandibular margin (Langston, Citation1965). Charactosuchus has much more emarginated alveoli. The Panamanian specimens differ from those typical of Miocene-aged South American gharials such as Gryposuchus in being much narrower anteroposteriorly and with wider interalveolar spacing (Langston and Gasparini, Citation1997). Hesperogavialis, from Venezuela, has wider interalveolar spacing, but much more dorsally facing alveoli that are further inset from the lateral dentary margin (Bocquentin-Villanueva and Buffetaut, Citation1981). The dentaries are similar to Gavialosuchus americanus in interalveolar spacing, orientation of dentition, and presence of a smooth margin. The Panamanian dentaries are narrower than those of Gavialosuchus carolinensis (Erickson and Sawyer, Citation1996). We cannot exclusively rule out gryposuchines and because these taxa are of very different placements within Crocodylia, they remain identified only as Crocodylia indet.
Teeth
Crocodylian teeth from the Cascadas and Culebra formations consist of both long, pointed and broad, spade-shaped forms (Fig. S7). These are neither particularly large nor small, and are similar to teeth found in many species of crocodylians.
A fossil site from the Hodges Hill locality (Cucaracha Formation; 9.04768847°N, 79.653814°W) has recovered numerous microfossils and some of the most abundant fossils recovered are very small (<1 cm) crocodylian teeth (Fig. S7). These teeth are from varying positions in the jaw, but all are less than 1 cm in length, with no larger preserved individuals. From comparisons with juvenile extant Alligator, the sizes of the teeth from this site indicate that they belonged to young individuals less than one year old. Other sites from the Cucaracha Formation have larger teeth that are more similar to those of the Culebra and Cascadas formations.
Gillette (Citation1984) briefly described crocodylian teeth from the Gatun Formation. The eight teeth were collected via screen washing and are thus far the only known crocodylian teeth from the Gatun Formation. The teeth are consistent with others found within the Cucaracha and Culebra formations. The Gatun teeth are recurved to varying degrees, triangular, and possess subtle striations (Fig. S8). Each tooth bears a distinct carina defining the lingual and labial surfaces. The varying degree of lingual/buccal separation of the carinae and lingual curvature suggest that the teeth were from different sections of the jaw. The similarity in size between all of these specimens suggests at least the possibility that they came from the same individual. None of these teeth resemble the spade-shaped teeth found in the new caimanine taxa of the Cucaracha and Culebra formations.
An isolated tooth (UF 259874) from the same stratigraphic section as the right dentary (UF 259875) is distinct from all other fossil Panamanian crocodylian teeth (). It is very long (46 mm), recurved, thin, and has pronounced carinae. This isolated tooth is much larger and longer than any other fossil crocodylian tooth from Panama and its recurved, thin shape is entirely consistent with teeth of Gavialosuchus americanus (). These long, recurved teeth as well as the longirostrine skull shape are thought to be associated with piscivory (Rayfield et al., Citation2007). This tooth likely belongs to a longirostrine crocodylian, possibly the same as for the dentary fragments described above.
FIGURE 9 Fossil crocodylian teeth. A, elongate and recurved tooth (UF 259874) from the early Miocene Culebra Formation in posterior view; B, premaxillary tooth of Gavialosuchus americanus (UF 211283) from Florida for comparison. Scale bar equals 1 cm.

Ribs
Partial rib fragments are commonly found as fossils within the Panama Canal Zone. One right second dorsal rib in particular from the Cucaracha Formation, Cartagena Hill locality (UF 245506; 9.02273066°N, 79.623633°W), was very well preserved and clearly belonged to a large crocodylian, and has a strongly divergent tuberculum and capitulum (). The rib is very similar in overall morphology to extant Alligator, only larger and thicker.
FIGURE 10 Crocodylian postcrania and coprolite from Miocene formations of the Panama Canal Zone. A, large right distal femur (USNM 7283) from the late Miocene Gatun Formation in lateral view; B, complete right femur (USNM 171018) from the early or middle Miocene Cucaracha Formation in lateral view; C, right distal humerus (UF 244332) from the early Miocene Culebra Formation in lateral view; D, anterior thoracic rib, position 2 (UF 245506), from the early or middle Miocene Cucaracha Formation in posterior view; E, partial osteoderm (UF 244331) from the early Miocene Culebra Formation in dorsal view; F, right proximal radius (UF 259874) from the early Miocene Culebra Formation in lateral view; G, coprolite (USNM 23177) from the early or middle Miocene Cucaracha Formation in lateral view; H, same as F in proximal articular view. Scale bar for A–D equals 10 cm; scale bar for E–H equals 5 cm.
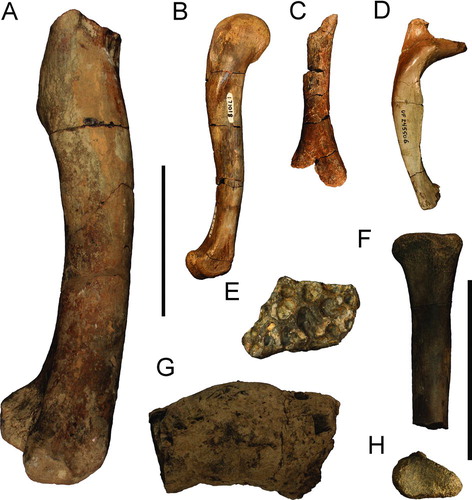
Humerus
A single partially preserved right distal humerus (UF 244332; ) was discovered at the El Lirio Norte site from the Culebra Formation (9.0535°N, 79.6571°W). This specimen is long and thin and has part of the medial condyle preserved. The narrowest diameter is 55 mm. The deltopectoral crest is missing, making further identification difficult.
Radius
A single right proximal radius (UF 259874) was discovered from the Centenario Bridge site of the Cucaracha Formation (). This radius is thinner than that of similarly sized Alligator and bears a small, but strong crest on the lateral side, just below the articular surface for the M. humeroradialus (Meers, Citation2003).
Femora
An isolated crocodylian femur from the Gatun Formation (USNM 7283) was reported by Gilmore (Citation1912). The femur is clearly from a very large crocodylian (). The preserved portion of this femur is over 30 cm in length, and is missing the entirety of the proximal articular head as well as the fourth trochanter. Based on the regressions of Farlow et al. (Citation2005), dimensions of the distal femoral condyle predict a total body length of 4.74–4.97 m. Given the marine nature of the lithology of the site, Gavialosuchus seems the most appropriate match, because it is known to have attained great size and lived within this habitat (Erickson and Sawyer, Citation1996). However, an isolated femur is not diagnostic enough to assert such a definitive identification, and it is instead referred to Crocodylia indet.
In addition to the Gatun femur, two additional isolated femora have been recovered from the Cucaracha Formation. A complete femur was discovered at the same site as the ‘headless skeleton’ (). It is unclear whether or not it can be associated with the ‘skeleton’ and was catalogued separately. Based on the size of the femur relative to the vertebrae of the ‘skeleton,’ it likely represents a different individual, and possibly another taxon entirely. The femur has a well-developed fourth trochanter and complete proximal and distal articular surfaces. The second femur from the Cucaracha Formation greatly resembles the first, only different in its notably smaller size.
Osteoderms
Four additional partial osteoderms have been catalogued thus far from the Culebra and Cucaracha formations as well as an additional four osteoderm fragments. These osteoderms are much more difficult to identify, but have more varied and deeper pitting, which is not typical of Gavialosuchus or gryposuchines.
An additional isolated osteoderm (UF 244331; ) from the Culebra Formation (El Lirio Norte locality; 9.0535°N, 79.6571°W) shows the same shallow wide pitting found in the Cucaracha ‘skeleton’ osteoderms, and may pertain to the same taxon.
Coprolites
Two ‘coprolites’ were recovered from the same site as the ‘headless skeleton.’ Both are smooth, rounded, and have little morphology. One is broken, revealing preserved cancellous bone, indicating that it is likely not a coprolite, but instead fragmentary bone surrounded by matrix. This identification is due in part to the poor likelihood of recognizable bone structure surviving the crocodylian digestive system as well as the fossilization process (Fisher, Citation1981). The other specimen may indeed be a coprolite of a young adult crocodylian ().
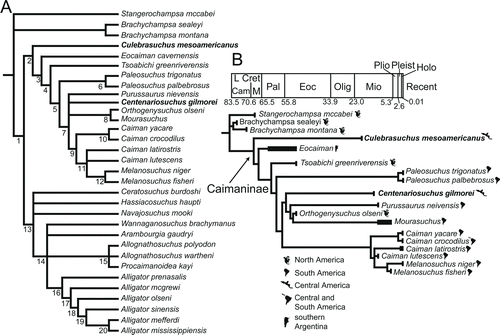
FIGURE 11 Cladograms generated from cladistic analysis of 75 characters for 32 taxa of Alligatoridae and three outgroup taxa. A, strict consensus cladogram of 1210 trees; B, 50% majority rule consensus cladogram from analysis of Caimaninae and their outgroup, placed in stratigraphic and geographic contexts. Black boxes indicate known stratigraphic occurrences.
PHYLOGENETIC ANALYSIS
The current analysis is modified from Brochu (Citation1999). It differs in that we reduced the number of taxa to the family Alligatoridae plus three outgroup taxa (Stangerochampsa mccabei, Brachychampsa montana, and Brachychampsa sealeyi). As such, many of the original 164 characters were rendered uninformative and we reduced the matrix to 75 parsimoniously informative characters. Tsoabichi greenriverensis was added to the analysis given its relevance to the fossil crocodylians of Panama based on information from Brochu (Citation2010). Furthermore, we added the two new species, Culebrasuchus mesoamericanus and Centenariosuchus gilmorei. Character descriptions are provided in Appendix 1 and the data matrix is provided in Appendix 2.
We were able to score additional character states (characters 24, 28, and 35) for Eocaiman that were not included in the previous analysis (Brochu, Citation1999) based on the more recent description of Eocaiman palaeocenicus (Bona, Citation2007) from the Paleocene of southern Argentina.
Because only a single character could be scored for Caiman tremembensis, Necrosuchus ionensis, and Notocaiman stromeri based on the existing literature (character 38, gently curved dentary), they were not included in the current analysis. In all, 32 taxa including the three outgroup taxa were included. A heuristic search was conducted on the matrix using PAUP 4.0b10 (Swofford, Citation2004) with 10,000 replicates.
The analysis resulted in 1210 equally most parsimonious cladograms with a tree length of 142 steps (consistency index = 0.648; retention index = 0.848; homoplasy index = 0.352; rescaled consistency index = 0.550). The large number of cladograms is due largely to an unresolved polytomy between basal members of Alligatorinae ().
In the strict consensus (), Culebrasuchus mesoamericanus is the sister to Caimaninae (sensu Brochu, Citation2010). Eocaiman is sister to all other caimanines and Tsoabichi greenriverensis is the sister to crown group Caimaninae. This position for T. greenriverensis differs from the results of Brochu (Citation2010), which had this taxon as sister to Paleosuchus (extant dwarf caiman). Centenariosuchus gilmorei is situated in a polytomy with Purussaurus neivensis, the paired grouping of Orthogenysuchus olseni and Mourasuchus, and a clade uniting all Caiman and Melanosuchus. The 50% majority rule consensus cladogram () shows more resolution, with Centenariosuchus sister to a clade including P. neivensis, Orthogenysuchus olseni, and Mourasuchus. As in Brochu (Citation2010), Caiman is rendered paraphyletic with respect to the Melanosuchus clade ().
Given the uncertain condition of the frontoparietal suture in UF 262800, this character (character 44) was not coded for Centenariosuchus. However, in two subsequent analyses we ran it with the character scored as either linear or concavoconvex. When the character was scored as linear, the strict consensus placed Centenariosuchus in the same position as the 50% majority rule consensus. When the character was scored as concavoconvex, Centenariosuchus was instead sister to Purussaurus neivensis.
BIOGEOGRAPHIC ANALYSIS
We used S-DIVA (Yu et al., Citation2010a) on our phylogenetic results to investigate the paleobiogeographic implications of the new fossil taxa. S-DIVA generated an independent consensus tree utilizing the geographic distribution of the taxa, as binned into six different broad regions (Fig. S9). Due to the limitations of the program, polytomies needed to be resolved to the most often recurring topology (Yu et al., Citation2010b). Furthermore, the program cannot discern contiguous from non-contiguous regions, and thus treats combined regions such as AB the same as AF, even though in reality A and B are contiguous whereas A and F are separated by a large dispersal barrier (Kodandaramaiah, Citation2010). However, despite its limitations the program can provide some support for the dispersal methods already implied by the phylogenetic analysis. The S-DIVA consensus greatly resembles that of the PAUP 50% majority rule consensus (Fig. S9). The only difference is a more resolved relationship among Caiman, with C. latirostris and C. lutescens pairing and being sister to the Melanosuchus clade, as opposed to being in an unresolved polytomy.
Dispersals for all of the outgroups as well as the initial split between Alligatorinae and Caimaninae cannot be resolved between being located exclusively in North America and being present throughout North, Central, and South America. Five equally parsimonious distributions for the common ancestor of Caimaninae were recovered, all of which include Central America. Four possible distributions include North America and both northern and southern South America are included in three out of five of the scenarios. The analysis shows clear presence of the ancestor of Eocaiman as being present in southern South America, with a 2/3 likelihood of also being present in northern South America and North America. A clear bifurcated population of crown group Caimaninae and Tsoabichi greenriverensis is suggested, located in both North America and northern South America. One of these populations would have given rise to T. greenriverensis, the other to crown group caimanines. Crown caimanines remain an entirely northern South American clade except for two dispersal events to Central America by the ancestor of Centenariosuchus gilmorei and an independent dispersal to North America by the ancestor of Orthogenysuchus olseni.
DISCUSSION
Diagnosis of Caimaninae
Brochu (Citation2010) found four unambiguous synapomorphies that are shared by all then-known Caimaninae: (1) splenials that do not meet at the midline, but end anteriorly dorsal to the Meckelian groove; (2) a surangular-angular suture that passes broadly along the ventral margin of the external mandibular foramen; (3) supraoccipital exposure on the dorsal skull table; and (4) long processes extending ventrally to the basioccipital tubera. Of these, Culebrasuchus mesoamericanus shares two (1 and 3). We follow an expanded concept of Caimaninae that can be formally defined under the guidelines of Phylocode (Cantino and de Queiroz, Citation2010) as the clade stemming from the common ancestor of Culebrasuchus mesoamericanus and Caiman crocodilus (). Traits likely characterizing the common ancestor of this group include (1) splenials that do not meet at the midline, but end anteriorly dorsal to the Meckelian groove; and (2) supraoccipital exposure on the dorsal skull table.
Biogeography
Results from our biogeographic analysis support either an entirely North American alligatorid ancestral population, or one that was present across all of the New World. Given the known fossil record of alligatorid outgroups exclusively in North America, the former hypothesis with a southern dispersal seems likely during, or shortly after, the Late Cretaceous. Particularly for the Late Cretaceous, a land bridge has been proposed due to the dispersal of several terrestrial taxa from North America to South America, including snakes, didelphid marsupials, condylarths, hadrosaurid ornithopods, titanosaurid sauropods, and at least one theropod (Rage, Citation1986). The likely dispersal route was along the Greater Antilles Arc, now Cuba and Puerto Rico, at the time located between southern North America and northern South America (; Hauff et al., Citation2000; Iturralde-Vinent, Citation2006; Proenza et al., Citation2006). Caimanine dispersal into Central America prior to the early Paleocene is supported by our analysis (Fig. S9). How widespread this clade was remains unclear, only their existence in Central America, likely near the Yucatan Peninsula, is shown by the biogeographic analysis. Dispersal from Central America to southern Argentina by the Paleocene likely occurred, probably while maintaining a presence in northern/central South America ().
FIGURE 12 Biogeographic dispersal of caimanines placed in a temporal context from the Late Cretaceous to Recent. Paleomaps are modified from Scotese (Citation2001). Modern distributions were compiled from Ross (Citation1998).
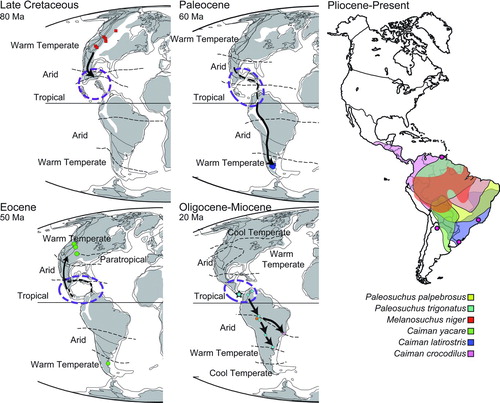
Although a population remained in southern Argentina, the persistent northern/central South American taxa dispersed back to North America in the Eocene, giving rise to Tsoabichi. A possible island chain between the Americas during the Eocene may help explain this back dispersal to North America, although so far uplift is only known to have occurred by the late Eocene, rather than the early Eocene (Iturralde-Vinent, Citation2006; Herrera et al., Citation2012; CitationMontes et al., 2012). A study focusing on New World frog genetics found evidence supporting an Eocene dispersal of these non-marine organisms, suggesting a possible land dispersal method was available at this time (Heinicke et al., Citation2007). There is an alternative explanation, although not supported by as many topologies, that an ancestor of Tsoabichi remained in North America since before the caimanine-alligatorine split. Following dispersal of the Tsoabichi lineage, Caimaninae diversified into five different groups, but with most representatives absent from the fossil record until the Miocene. Two independent lineages migrated north, one giving rise to Centenariosuchus gilmorei in Panama and the other giving rise to Orthogenysuchus from the Eocene of Wyoming (). These two do not share a direct relationship within Caimaninae, suggesting independent dispersals () and further indicating a still unknown early diversification of Caimaninae (). Notably, the timing of this diversification () creates a ghost lineage for Paleosuchus (extant dwarf caiman), Centenariosuchus, Purussaurus, and Mourasuchus back into the Eocene. However, the fossil record of caimanines is minimal during the Eocene–Oligocene of South America, and the fossils of these lineages have likely not yet been discovered. The ghost lineage for the Paleosuchus clade is extensive, but possible fossils from the Miocene of Peru may help fill in this gap (Salas-Gismondi et al., Citation2007). The biogeographic analysis supports a dominantly northern/central South American distribution of Caimaninae.
The Miocene has a large and diverse record of caimanines from the New World Tropics (). Meanwhile, the more primitive population of Central America is preserved in the early Miocene of Panama, as represented by Culebrasuchus mesoamericanus, and later on by the more derived immigrant C. gilmorei. Considering Panama was mostly underwater for much of its history, the ancestor of Culebrasuchus likely lived in more northern parts of Central America, possibly southern Mexico (Iturralde-Vinent, Citation2006; Bayona et al., 2010; Herrera et al., Citation2012). Once Panama surfaced and formed a continuous landmass in the Miocene (Kirby et al., Citation2008), this population likely immigrated southward, reaching what is now the Panama Canal Zone. Caimanine diversity decreases after the Miocene, with most forms going extinct (Riff et al., Citation2010). These now extinct lineages would have diversified into the extant caimans, dispersing and establishing populations throughout the Amazon Basin (). Additionally, a population of Caiman crocodilus emigrated from South America into Central America 2.5–2.9 Ma, reaching as far north as southern Mexico (Venegas-Anaya et al., Citation2008). Further evidence reveals that the split between C. crocodilus and C. yacare likely occurred prior to 5.7–6.7 Ma within South America.
We should note that the presence of other caimanines during the Eocene in southern Argentina (Rusconi, Citation1937; Simpson, Citation1937) may or may not affect the biogeographic conclusions drawn here. We presume based on temporal and geographic proximity that these taxa represent forms closely related to the better-known Eocaiman. Likewise, we presume Caiman tremembensis (Chiappe, Citation1988) closely resembles in morphology other members of Caiman, and thus its dispersal would be similar. Once more material is known and studied within a rigorous phylogenetic context, more can be said regarding the phylogeny and biogeography of these poorly known taxa.
Temperature Tolerance
Prior to this discovery, both the common ancestor of Alligatoridae and the oldest caimanine fossils came from warm temperate high latitudes (; Otto-Bliesner and Upchurch, Citation1997). This implied a high-latitude, warm-temperate origin for Caimaninae, regardless of which location was considered to be the center of origin. However, given the lack of fossil crocodylians from the New World Tropics in the Paleogene, this may be due to undersampling. Regardless, the presence of the basal-most caimanine, or its predecessor, in the New World Tropics indicates a likely origin of Caimaninae within a tropical ecosystem.
Extant caimans have a tropical distribution, unlike extant alligators, which have a much more temperate distribution (Ross, Citation1998). Alligators today are capable of surviving hard freezes in temperate environments by establishing breathing holes as the water surface freezes. The relative cold tolerance of Alligator and Caiman was tested by Brandt and Mazzotti (Citation1990) who placed both in the same enclosure in South Carolina over winter to test temperature tolerance between the two groups. The Caiman suffered much higher fatality rates, particularly after the first freeze event, which combined with the recorded increased basking time of the Caiman suggests that they are either behaviorally or physiologically less well adapted to environments with seasonally cold temperatures (Brandt and Mazzotti, Citation1990). This observation may explain the lack of high-latitude caimanines as climate cooled significantly across the Neogene (Zachos et al., Citation2008).
Tropical habitat preference for extant caimans appears to be a plesiomorphic trait for all Caimaninae. The presence of at least three warm temperate caimanines in the Paleocene and Eocene suggests that independent adaptations for more temperate habitats evolved from a primitive stock of caimanines. These caimanines inhabited higher latitude at a time when temperatures were more similar to modern subtropics and thus may have experienced milder winters than modern temperate-adapted alligators (Roehler, Citation1993). Regardless, these higher latitude caimanines were unable to persist in these areas past the warm Eocene epoch.
Salinity Tolerance
Dispersal of caimanines between continents likely occurred via land connections or closely spaced islands due to the lack of saltwater tolerance in all extant alligatorids (Taplin and Grigg, Citation1989). Extant Alligator have populated offshore islands in Florida and Georgia (Jacobsen, Citation1983; Tamarack, Citation1989) and extant Caiman have populated the island of Tobago (Ross, Citation1998); however, whether or not these represent marine dispersals or relict populations from once-continuous landmasses remains to be determined. Populations of Caiman in Cuba, Puerto Rico, and Florida were introduced historically (Ross, Citation1998). A study conducted on the salinity tolerance of wild Caiman latirostris in Brazil revealed that the species will not drink saltwater and that the species becomes more rapidly dehydrated in saltwater than Crocodylus porosus (Grigg et al., Citation1998). The study group appeared to survive by making short forays into brackish water to find prey, and then returned to a nearby freshwater habitat (Grigg et al., Citation1998).
At least one dispersal to South America from North America must have occurred, and the timing of the fossil record suggests that it may have occurred along the Greater Antillean Arc (). Due to the possibility of a relict basal caimanine population in Central America, the presence of Culebrasuchus mesoamericanus in Panama does not necessitate a saltwater dispersal. However, the presence of Centenariosuchus gilmorei and possible occurrence of Purussaurus does imply a saltwater dispersal of a caimanine. Brochu (Citation2010) suggested at least two dispersals from South America to North America during or prior to the Eocene to explain the occurrence of Tsoabichi and Orthogenysuchus, both known from the Eocene of Wyoming, but which are not sister taxa. These Eocene dispersals are more problematic from the salinity standpoint, because the Greater Antillean Arc had migrated away from North and South America by this time (Coney, Citation1982). This back dispersal likely occurred in the late Paleocene or early Eocene when there was a short-lived volcanic arc between North and South America (Bayona et al., 2010; Cardona et al., Citation2010). The presence of a basal caimanine in Central America suggests that rather than a population making the long immigration from southern Argentina to Wyoming during the Paleocene, instead two dispersals, one north and one south, of basal caimanines likely occurred (). Later, in the Miocene when Panama was above sea level, Culebrasuchus mesoamericanus or its ancestor would have immigrated to Panama. An alternative to this would be the presence of the Culebrasuchus ancestor being present in northern South America, without a fossil record, then a dispersal by the Miocene across the approximately 200 km gap to Panama (CitationMontes et al., 2012). Marine dispersal would have been required to account for the presence of Centenariosuchus gilmorei and cf. Purussaurus given their closer relationships to South American taxa. The presence of Caiman in Central America today has been shown to have likely occurred after the formation of the Panamanian isthmus, via non-marine dispersal (Venegas-Anaya et al., Citation2008).
Longirostrine Significance
Gavialosuchus was originally named on the basis of a fossil skull from the Miocene of Austria (Toula and Kail, Citation1885) and given the species name Gavialosuchus eggenburgensis. A fossil from the Miocene of Florida was originally named Tomistoma americana (Sellards, Citation1915), but this species was then reassigned to Gavialosuchus (Mook, Citation1921). Erickson and Sawyer (Citation1996) named a new species of Gavialosuchus, G. carolinensis, for late Oligocene fossils from South Carolina. Myrick (Citation2001) synonymized Gavialosuchus americanus with Thecachampsa antiqua; however, T. antiqua (Leidy, Citation1852) was described on the basis of a single tooth whereas Gavialosuchus is based on a nearly complete skull, and is thus a better-defined taxon. Many fossils of Gavialosuchus americanus have been identified from early–late Miocene deposits in Florida (Hulbert, Citation2001). Its presence in other regions depends on interpretations of taxonomy, which range from occurring only in Florida and Costa Rica, to also occurring in many mid-Atlantic sites up to New Jersey, as well as California, Baja in Mexico, Bolivia, and Portugal (Carrasco, Citation1986; Myrick, Citation2001; Riff et al., Citation2010). We abide by Sellards (Citation1915) and Mook (Citation1921) in their identification of a species independent of T. antiqua.
Likewise, the longirostrine Charactosuchus from Colombia has been identified from North American localities based on dubious records. Webb and Tessman (Citation1967) published a faunal list from the Pliocene-aged Manatee County dam site in Florida, including cf. Charactosuchus. Charactosuchus was named by Langston (Citation1965) for two partial mandibles from the Miocene-aged La Venta locality, Colombia. A Florida occurrence would imply that this small crocodylid had emigrated from South America. Webb and Tessman (Citation1968) provided a more thorough description in which they noted that their earlier identification was based on a single tooth (UF 11938). They made the assignment to cf. Charactosuchus based on comparisons with a partial maxilla and tooth that were described, but not identified, by Auffenberg (Citation1957) from South Carolina (ChM. 41.188.45) that had a similarly ribbed tooth. Steel (Citation1973) agreed with Webb and Tessman (Citation1968) and reported Charactosuchus as present in North America. However, Langston and Gasparini (Citation1997) refuted the identifications of both the Florida tooth and the South Carolina maxilla. Hulbert (Citation2001) cited Langston and Gasparini (Citation1997) as the authority for the lack of Charactosuchus in his faunal list of Florida fossil vertebrates. At the time of submission of this article, the Florida tooth (UF 11938) appears to have been lost (R. Hulbert, pers. comm.). Regardless, it appears there is no reliable record of Charactosuchus in North and Central America. Thus, the only longirostrine taxon described for this time period in North and Central America remains the tomistomine Gavialosuchus americanus. Due to similar alveolar spacing and orientation, it is likely that the Panamanian longirostrine taxon represents G. americanus as well, but more diagnostic fossils will be required to be certain.
Mixed Fauna
Miocene crocodylians from North America have similar ecomorphology, but different phylogenetic affinities as compared with the crocodylians of South America. In both regions, fossil crocodylians have both short- and long-snouted forms co-occurring. In southern North America, Miocene crocodylians are typified by fossil species of the short-snouted extant genus Alligator and the extinct long-snouted tomistomine Gavialosuchus americanus (Hulbert, Citation2001). In northern South America, Miocene crocodylians filling the short-snouted niche are various taxa of caimanines (e.g., Paleosuchus, Caiman, Melanosuchus) and the long-snouted niche is occupied by various gharials (e.g., Gryposuchus, Hesperogavialis, and Ikanogavialis) and the crocodylid Charactosuchus (Langston, Citation1965; Riff et al., Citation2010). Both North and South American alligatorids are thought to be incapable of dispersing across large oceanic distances, and thus are expected to have remained isolated on their respective continents if the separation between North and South America was large. In contrast, the presence of caimans with South American affinities in Panama during a time when it was not connected to South America might provide additional support for a closer proximity of the two landmasses, as recently suggested by CitationMontes et al. (2012). Conversely, a gharial with South American affinity was found in Puerto Rico (Vélez-Juarbe et al., Citation2007) and thus at least one gharial was therefore capable of dispersing across large spans of saltwater.
FIGURE 13 Summary diagram of each of the formations from the Panama Canal Zone from which crocodylian fossils have been recovered thus far.
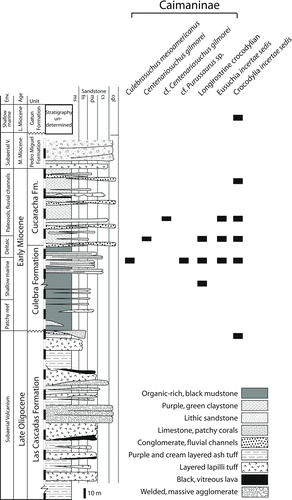
The progression of crocodylians preserved in the Panama Canal Zone indicates a mixed fauna of North and South American taxa (). The crocodylian fossils from the Las Cascadas Formation (late Oligocene/early Miocene) are the oldest fossil record of crocodylians in Central America. Their fossil record thus far is the most incomplete of the Panamanian formations, but indicates that both small- and large-bodied individuals were present. The lithology and other fauna present (e.g., camels) indicate a terrestrial, fluvial setting (Rincon et al., Citation2012). The preserved teeth (Fig. S7) and vertebra (Fig. S5) are entirely consistent with referral to Caimaninae and none of the teeth reflect the elongate, strongly recurved teeth () or wide, triangular teeth () that are typical of Gavialosuchus. Due to the large size of the vertebra and small size of most of the teeth, it seems most likely that both large- and small-bodied caimanines were present. An alternative would be only that a large-bodied caimanine was present as well as its young. Caimanines are known in North America from the Eocene and in South America from Paleocene–Recent. Whether these earliest crocodylians were most similar to the North American or South American taxa will require more identifiable fossils to be recovered.
The early Miocene Culebra Formation is dominantly marine in lithology, but toward the upper part of this formation, it grades into more terrestrial deposits (Kirby et al., Citation2008; CitationMontes et al., 2012). Thus far no crocodylian fossils have been recovered from the marine section, but identifiable fossils have been recovered from the more littoral upper section. Thus far the upper Culebra Section has had the most diverse array of crocodylian fossils, with excellent remains of a small-bodied caimanine (Culebrasuchus mesoamericanus), a large-bodied caimanine (cf. Purussaurus sp.), and a small- to medium-sized longirostrine crocodylian (). These three crocodylian body types would have been occupying separate roles within the aquatic ecosystem during the early Miocene. The longirostrine taxon would have been dominantly piscivorous (Rayfield et al., Citation2007), the Purussaurus-like caimanine would have specialized in large, tough-bodied prey, such as the turtles also found at the site. Only the cf. Purussaurus sp. would necessarily indicate that a marine dispersal had occurred by this time (). The less certain presence of a large Purussaurus in the earlier Las Cascadas Formation may indicate that this dispersal had occurred earlier, but more identifiable fossils are needed in order to be certain. The presence of a tomistomine (e.g., Gavialosuchus) and not a gavialid would be surprising due to the close proximity of several genera of gharials in South America at the time (Riff et al., Citation2010), especially considering their presence in the Oligocene of Puerto Rico (Vélez-Juarbe et al., Citation2007).
The early or middle Miocene Cucaracha Formation has well-preserved remains of both small-bodied caimanines and longirostrine crocodylians. The possible presence of the North American Gavialosuchus americanus and the South American immigrant Centenariosuchus would indicate the persistence of a mixed crocodylian fauna in the Miocene of Panama. With the exception of the South American River Turtle (Podocnemididae), no other tetrapod taxon from the Miocene of Panama has South American affinities. Freshwater-adapted crocodylians and turtles seem to have been able to disperse from South America across the seaway to Panama more readily than the terrestrial mammals.
The late Miocene Gatun Formation possesses strongly marine indicators in both lithology and fossil fauna (Collins et al., Citation1996), and the water depth may have been shallow enough to support a shark nursery (Pimiento et al., Citation2010). The few crocodylian remains thus far discovered indicate that a large-bodied crocodylian was present () and the smaller teeth (Fig. S8) also indicate the presence of a likely smaller or younger piscivore. Given the possible presence of Gavialosuchus americanus based on symphyseal dentary sections in earlier formations, the continued presence of this taxon in the Gatun Formation seems also possible, especially considering the taxon's presence in the marine faunas of Florida (Hulbert, Citation2001). Without new fossils from the Gatun or other formations, the persistence of crocodylians of mixed continental origins cannot be discerned for this stratigraphic horizon. The modern presence of just two crocodylian species in central Panama is likely the result of terrestrial dispersal enabled by the land connection of North and South America, allowing for the presence of the American Crocodile, Crocodylus acutus, and Spectacled Caiman, Caiman crocodilus (Ross, Citation1998; Venegas-Anaya et al., Citation2008).
Hastings et al Supplementary Data
Download Zip (4 MB)ACKNOWLEDGMENTS
We would like to thank the Smithsonian Tropical Research Institute (STRI) and Florida Museum of Natural History (FLMNH) for support and assistance with the research project and for collection of the fossil described here. The accomplishment of this project was possible thanks to the support of the U.S. National Science Foundation Partnerships in International Research and Education grant 0966884 (OISE, EAR, DRL), EAR 0824299, and EAR 0418042; the Autoridad del Canal de Panama; the Mark Tupper Fellowship; Ricardo Perez SA; and the Smithsonian Institute. Thanks to U. Denetclaw for collection of the holotype of Centenariosuchus gilmorei and L. Grawe-Desantis for collection of one of the longirostrine dentary sections (UF 259873). Thanks to J. Bourque for excellent preparation of the holotype and paratype specimens and to K. Rowe and J. Nestler for research assistance. Thanks to M. Brett-Surman (USNM), R. Hulbert (FLMNH), and D. Winkler (SMU) for access to, and assistance with, collections.
Handling editor: You Hailu
Related Research Data
LITERATURE CITED
- Aguilera , O. A. , Riff , D. and Bocquentin-Villanueva , J. 2006 . A new giant Purussaurus (Crocodyliformes, Alligatoridae) from the upper Miocene Urumaco Formation, Venezuela . Journal of Systematic Palaeontology , 4 : 221 – 232 .
- Aoki , R. 1992 . Fossil crocodilians from the late Tertiary strata in the Sinda Basin, eastern Zaire . African Study Monographs , 17 : 67 – 85 .
- Auffenberg , W. 1957 . Notes on fossil crocodilians from southeastern United States . Journal of the Florida Academy of Sciences , 20 : 107 – 113 .
- Barbosa-Rodrigues , B. 1892 . Les Reptiles fossiles de la vallée de l’Amazone . Vellosia , 2 : 41 – 46 .
- Bayona , G. , Montes , C. , Cardona , A. , Jaramillo , C. , Ojeda , G. , Valencia , V. and Ayala-Calvo , C. 2011 . Intraplate subsidence and basin filling adjacent to an oceanic arc-continent collision: a case from the southern Caribbean-South America plate margin . Basin Research , 23 : 403 – 422 .
- Benton , M. J. , Clark , J. M. and Benton , M. J. 1988 . “ Archosaur phylogeny and the relationships of the Crocodylia ” . In The Phylogeny and Classification of the Tetrapods , Vol. 1 , 295 – 338 . Oxford : Clarendon Press .
- Bocquentin-Villanueva , J. and Buffetaut , E. 1981 . Hesperogavialis cruxenti n. gen. n. sp., nouveau gavialidae (Crocodylia, Eusuchia) du Miocéne supérieur (Huayquerian) d’Urumaco (Venezuela) . Géobios , 14 : 415 – 419 .
- Bona , P. 2007 . Una nueva especie de Eocaiman Simpson (Crocodylia, Alligatoridae) del Paleoceno Inferior de Patagonia . Ameghiniana , 44 : 435 – 445 .
- Brandt , L. A. and Mazzotti , F. J. 1990 . The behavior of juvenile Alligator mississippiensis and Caiman crocodilus exposed to low temperature . Copeia , 1990 : 867 – 871 .
- Brochu , C. A. 1996 . Closure of neurocentral sutures during crocodilian ontogeny: implications for maturity assessment in fossil archosaurs . Journal of Vertebrate Paleontology , 16 : 49 – 62 .
- Brochu , C. A. 1999 . Phylogeny, systematics, and historical biogeography of Alligatoroidea . Memoir of the Society of Vertebrate Paleontology , 6 : 9 – 100 .
- Brochu , C. A. 2003 . Phylogenetic approaches toward crocodylian history . Annual Review of Earth and Planetary Sciences , 31 : 357 – 397 .
- Brochu , C. A. 2010 . A new alligatorid from the lower Eocene Green River Formation of Wyoming and the origin of caimans . Journal of Vertebrate Paleontology , 30 : 1109 – 1126 .
- Buscalioni , A. D. , Sanz , J. L. and Casanovas , M. L. 1992 . A new species of the eusuchian crocodile Diplocynodon from the Eocene of Spain . Neues Jahrbuch für Geologie und Paläontologie, Abandlungen , 187 : 1 – 29 .
- Cantino , P. D. and de Queiroz , K. 2010 . International Code of Phylogenetic Nomenclature, Version 4c . http://www.ohio.edu/phylocode/. Accessed May 2012
- Cardona , A. , Ruiz , J. , Valencia , V. , Bayona , G. , Duque , J. , Jaramillo , C. , Montes , C. and Ojeda , G. 2010 . Turonian to Eocene accretion and subduction in the Santa Marta massif and Rancheria Basin, Colombia: implications for Northern Andean orogeny . Terranova , 23 : 26 – 34 .
- Carrasco , C. P. 1986 . Notas preliminares sobrela geologia del extremo noroccidental del Departamento de Pando. VII Congreso Geologico de Bolivia, Cochabamba, Septiembre 1984 . Actas , 1 : 175 – 180 .
- Chiappe , L. M. 1988 . Un nuevo caiman (Crocodylia, Alligatoridae) de la Formación Tremembé (Oligoceno), Estado de São Paulo, Brasil, y su significado paleoclimatico . Paula-Coutiana , 3 : 49 – 66 .
- Cisneros , J. C. 2005 . New Pleistocene vertebrate fauna from El Salvador . Revista Brasileria de Paleontologica , 8 : 239 – 255 .
- Clark , J. M. 1994 . “ Patterns of evolution in Mesozoic crocodyliforms ” . In In the Shadow of the Dinosaurs , Edited by: Fraser , N. C. and Sues , H.-D. 84 – 97 . New York : Cambridge University Press .
- Coates , A. G. , Collins , L. S. , Aubry , M.-P. and Berggren , W. A. 2004 . The geology of the Darien, Panama, and the late Miocene-Pliocene collision of the Panama arc with northwestern South America . Geological Society of America Bulletin , 116 : 1327 – 1344 .
- Collins , L. S. , Coates , A. G. , Berggren , W. A. , Aubry , M. P. and Zhang , J. 1996 . The late Miocene Panama isthmian strait . Geology , 24 : 687 – 690 .
- Coney , P. J. 1982 . Plate tectonic constraints on the biogeography of Middle America and the Caribbean region . Annals of the Missouri Botanical Garden , 69 : 432 – 443 .
- Erickson , B. R. and Sawyer , G. T. 1996 . The estuarine crocodile Gavialosuchus carolinensis n. sp. (Crocodylia: Eusuchia) from the late Oligocene of South Carolina, North America . Monograph of the Science Museum of Minnesota , 3 : 1 – 47 .
- Farlow , J. O. , Hurlburt , G. R. , Elsey , R. M. , Britton , A. R. C. and Langston , W. Jr. 2005 . Femoral dimensions and body size of Alligator mississippiensis: estimating the size of extinct mesoeucrocodylians . Journal of Vertebrate Paleontology , 25 : 354 – 369 .
- Farris , D. W. , Jaramillo , C. , Bayona , G. , Restrepo-Moreno , S. , Montes , C. , Cardona , A. , Mora , A. , Speakman , R. J. , Glasscock , M. D. , Reiners , P. and Valencia , V. 2011 . Fracturing of the Panamanian Isthmus during initial collision with South America . Geology , 39 : 1007 – 1010 .
- Ferrusquía-Villafranca , I. 1977 . Distribution of Cenozoic vertebrate faunas in middle America and problems of migration between North and South America . Boletín Universidad Nacional Autonoma de Mexico, Instituto de Geología , 101 : 193 – 321 .
- Fisher , D. C. 1981 . Crocodilian scatology, microvertebrate concentrations, and enamel-less teeth . Paleobiology , 7 : 262 – 275 .
- Gillette , D. D. 1984 . A marine ichthyofauna from the Miocene of Panama, and the Tertiary Caribbean faunal province . Journal of Vertebrate Paleontology , 4 : 172 – 186 .
- Gilmore , C. W. 1912 . Fossil crocodiles from the Canal Zone . Proceedings of the Biological Society of Washington , 25 : 94
- Gmelin , J. F. 1789 . “ Caroli a Linné, Systema Naturae ” . Edited by: Beer , G. E. Leipzig , , Germany Tome I, Pars III., 1057 pp.
- Gray , J. E. 1844 . Catalogue of Tortoises, Crocodilians, and Amphisbaenians in the Collection of the British Museum , 88 London : British Museum (Natural History) .
- Grigg , G. C. , Beard , L. A. , Moulton , T. , Melo , M. T. Q. and Taplin , L. E. 1998 . Osmoregulation by the broad-snouted caiman, Caiman latirostris, in estuarine habitat in southern Brazil . Journal of Comparative Physiology, B , 168 : 445 – 452 .
- Hastings , A. , MacFadden , B. , Rincon , A. , Montes , C. and Jaramillo , C. 2009 . Filling in the Central American crocodile gap: evidence from the Middle Cenozoic of Panama . Journal of Vertebrate Paleontology, Program and Abstracts , 2009 : 111A – 112A .
- Hauff , F. , Hoernle , K. , van den Bogaard , P. , Alvarado , G. and Garbe-Schönberg , D. 2000 . Age and geochemistry of basaltic complexes in western Costa Rica: contributions to the geotectonic evolution of Central America . Geochemistry, Geophysics, Geosystems , 1 : 1999GC000020 doi: 10.1029/1999GC000020
- Heinicke , M. P. , Duellman , W. E. and Hedges , S. B. 2007 . Major Caribbean and Central American frog faunas originated by ancient oceanic dispersal . Proceedings of the National Academy of Sciences of the United States of America , 104 : 10092 – 10097 .
- Herrera , F. , Manchester , S. R. and Jaramillo , C. 2012 . Permineralized fruits from the late Eocene of Panama give clues of the composition of forests established early in the uplift of Central America . Review of Palaeobotany and Palynology , 175 : 10 – 24 .
- Hulbert , R. C. Jr. 2001 . The Fossil Vertebrates of Florida , 350 Gainesville , Florida : University Press of Florida .
- Huxley , T. H. 1875 . On Stagonolepis roberstoni, and on the evolution of the Crocodilia . Quarterly Journal of the Geological Society of London , 31 : 423 – 438 .
- Iordansky , N. N. 1973 . “ The skull of the Crocodilia ” . In Biology of the Reptilia , Edited by: Gans , C. and Parsons , T. Vol. 4 , 201 – 260 . London : Academic Press .
- Iturralde-Vinent , M. A. 2006 . Meso-Cenozoic Caribbean paleogeography: implications for the historical biogeography of the region . International Geology Review , 48 : 791 – 827 .
- Jacobsen , T. 1983 . Crocodilians and islands: status of the American alligator and the American crocodile in the Lower Florida Keys . Florida Field Naturalist , 11 : 1 – 24 .
- Kirby , M. X. , Jones , D. S. and MacFadden , B. J. 2008 . Lower Miocene stratigraphy along the Panama Canal and its bearing on the Central American peninsula . PLoS ONE , 3 : e2791
- Kodandaramaiah , U. 2010 . Use of dispersal–vicariance analysis in biogeography—a critique . Journal of Biogeography , 37 : 3 – 11 .
- Langston , W. Jr. 1965 . Fossil crocodilians from Colombia and the Cenozoic history of the Crocodilia in South America . University of California Publications in Geological Sciences , 52 : 1 – 157 .
- Langston , W. and Gasparini , Z. 1997 . “ Crocodilians, Gryposuchus, and the South American gavials ” . In Vertebrate Paleontology in the Neotropics: The Miocene Fauna of La Venta, Colombia , Edited by: Kay , R. F. , Madden , R. H. , Cifelli , R. L. and Flynn , J. J. 113 – 54 . Washington , D.C. : Smithsonian Institution .
- Laurito , C. A. and Valerio , A. L. 2008 . The first record of Gavialosuchus americanus Sellards (1915)† (Eusuchia: Crocodylidae, Tomistominae) for the Late Tertiary of Costa Rica and Central America . Revista Geológica de América Central , 39 : 107 – 115 .
- Leidy , J. L. 1852 . Description of a new species of crocodile from the Miocene of Virginia . Journal of the Academy of Natural Sciences of Philadelphia , 2 : 135 – 138 .
- MacFadden , B. J. 2006 . North American Miocene land mammals from Panama . Journal of Vertebrate Paleontology , 26 : 720 – 734 .
- Mead , J. I. , Cubero , R. , Zamora , A. L. V. , Swift , S. L. , Laurito , C. and Gómez , L. D. 2006 . Plio-Pleistocene Crocodylus (Crocodylia) from southwestern Costa Rica . Studies on Neotropical Fauna and Environment , 41 : 1 – 7 .
- Meers , M. B. 2003 . Crocodylian forelimb musculature and its relevance to archosauria . The Anatomical Record, Part A , 274A : 891 – 916 .
- Montes , C. , Cardona , A. , McFadden , R. R. , Moron , S. , Silva , C. A. , Restrepo-Moreno , S. , Ramirez , D. , Hoyos , N. , Wilson , J. , Farris , D. W. , Bayona , G. , Jaramillo , C. , Valencia , V. , Bryan , J. and Flores , J.-A. 2012 . Evidence for middle Eocene and younger emergence in Central Panama: implications for Isthmus closure . Geological Society of America Bulletin , 124 : 780 – 799 . doi: 10.1130/B30528.1
- Mook , C. C. 1921 . Skull characters and affinities of the extinct Florida gavial, Gavialosuchus americanus (Sellards) . Bulletin of the American Museum of Natural History , 44 : 33 – 41 .
- Mook , C. C. 1959 . A new Pleistocene crocodilian from Guatemala . American Museum Novitates , 1975 : 1 – 6 .
- Myrick , A. C. Jr. 2001 . Thecachampsa antiqua (Leidy, 1852) (Crocodylidae: Thoracosaurinae) from the fossil marine deposits at Lee Creek Mine, Aurora, North Carolina, USA . Smithsonian Contributions to Paleobiology , 90 : 219 – 225 .
- Norell , M. A. 1988 . Cladistic approaches to paleobiology as applied to the phylogeny of alligatorids , 272 New Haven , Connecticut : Yale University . Ph.D. dissertation
- Norell , M. A. 1989 . The higher level relationships of the extant Crocodylia . Journal of Herpetology , 23 : 325 – 335 .
- Norell , M. A. and Clark , J. M. 1990 . A reanalysis of Bernissartia fagesii, with comments on its phylogenetic position and its bearing on the origin and diagnosis of the Eusuchia . Bulletin de l’Institut Royal des Sciences Naturelles de Belgique , 60 : 115 – 128 .
- Otto-Bliesner , B. L. and Upchurch , G. R. Jr. 1997 . Vegetation-induced warming of high-latitude regions during the Late Cretaceous period . Nature , 385 : 804 – 807 .
- Pimiento , C. , Ehret , D. J. , MacFadden , B. J. and Hubbell , G. 2010 . Ancient nursery area for the extinct giant shark Megalodon from the Miocene of Panama . PLoS ONE , 5 : e10552
- Poe , S. 1996 . Data set incongruence and the phylogeny of crocodilians . Systematic Biology , 45 : 393 – 414 .
- Proenza , J. A. , Díaz-Martínez , R. , Iriondo , A. , Marchesi , A. , Melgarejo , J. C. , Gervilla , F. , Garrido , C. J. , Rodríguez-Vegar , A. , Lozano-Santacruz , R. and Blanco-Moreno , J. A. 2006 . Primitive cretaceous island-arc volcanic rocks in eastern Cuba: the Téneme Formation . Geologica Acta , 4 : 103 – 121 .
- Rage , J. C. 1986 . South American/North American terrestrial interchanges in the latest Cretaceous: short comments on Brett-Surman and Paul (1985), with additional data . Journal of Vertebrate Paleontology , 6 : 382 – 383 .
- Rayfield , E. J. , Milner , A. C. , Xuan , V. B. and Young , P. G. 2007 . Functional morphology of spinosaur ‘crocodile-mimic’ dinosaurs . Journal of Vertebrate Paleontology , 27 : 892 – 901 .
- Riff , D. , Romano , P. S. R. , Oliveira , G. R. and Aguilera , O. A. 2010 . “ Neogene crocodile and turtle fauna in northern South America ” . In Amazonia, Landscape and Species Evolution: A Look into the Past , Edited by: Hoorn , C. and Wesselingh , E. P. 259 – 280 . Oxford : Wiley-Blackwell .
- Rincon , A. F , Bloch , J. I. , Suarez , C. , MacFadden , B. J. and Jaramillo , C. A. 2012 . New floridatragulines (Mammalia, Camelidae) from the early Miocene Las Cascadas Formation, Panama . Journal of Vertebrate Paleontology , 32 : 456 – 475 .
- Roehler , H. W. 1993 . Eocene climates, depositional environments, and geography, greater Green River Basin, Wyoming, Utah, and Colorado . U.S. Geological Survey Professional Paper 1506-F , : 1 – 74 .
- Ross , J. P. 1998 . Crocodiles, Status Survey and Conservation Action Plan, , second edition , 97 Cambridge , , U.K : IUCN/SSC Crocodile Specialist Group .
- Rusconi , C. 1937 . Nuevo aligatorino del Paleoceno Argentino . Bulletin Paleontologico de Buenos Aires , 8 : 1 – 5 .
- Salas-Gismondi , R. , Antoine , P.-O. , Baby , P. , Brusset , S. , Benammi , M. , Espurt , N. , de Franceschi , D. , Pujos , F. , Tejada , J. and Urbina , M. 2007 . Middle Miocene crocodiles from the Fitzcarrald Arch, Amazonian Peru . Cuadernos del Museo Geominero , 8 : 355 – 360 .
- Scotese , C. R. 2001 . Digital Paleogeographic Map Archive , University of Texas at Arlington , Texas : PALEOMAP Project .
- Sellards , E. H. 1915 . A new gavial from the Late Tertiary of Florida . American Journal of Science, Series 4 , 40 : 135 – 138 .
- Simpson , G. G. 1937 . An ancient eusuchian crocodile from Patagonia . American Museum Novitates , 965 : 1 – 20 .
- Steel , R. 1973 . Crocodylia. Encyclopedia of Paleoherpetology , Vol. 16 , 116 Stuttgart : Gustav Fischer Verlag .
- Swofford , D. L. 2004 . PAUP: Phylogenetic Analysis Using Parsimony, Version 4.0b10 , London : MacMillan .
- Tamarack , J. L. 1989 . Georgia's coastal island alligators, variations in habitat and prey availability . Proceedings of the International Union for Conservation of Nature, Quito, Ecuador, 13–18 October 1986. Crocodile Specialist Group 8 , 1986 : 105 – 118 .
- Taplin , L. E. and Grigg , G. C. 1989 . Historical zoogeography of the eusuchian crocodilians: a physiological perspective . American Zoologist , 29 : 885 – 901 .
- Toula , F. and Kail , J. A. 1885 . Über einen Krokodil−Schädel aus den Tertiärablagerungen von Eggenburg in Niederösterreich: eine paläontologische Studie . Denkschriften der Keiserlichen Akademie der Wissenschaften von Wien, Mathematisch-naturwissenschaftliche Klasse , 50 : 299 – 355 .
- Vélez-Juarbe , J. , Brochu , C. A. and Santos , H. 2007 . A gharial from the Oligocene of Puerto Rico: transoceanic dispersal in the history of a non-marine reptile . Proceedings of the Royal Society B , 274 : 1245 – 1254 .
- Venegas-Anaya , M. , Crawford , A. J. , Escobedo Galván , A. H. , Sanjur , O. I. , Densmore , L. D. III and Bermingham , E. 2008 . Mitochondrial DNA phylogeography of Caiman crocodilus in Mesoamerica and South America . Journal of Experimental Zoology , 309A : 614 – 627 .
- Webb , S. D. and Tessman , N. 1967 . Vertebrate evidence of a low sea level in the Middle Pliocene . Science , 156 : 379
- Webb , S. D. and Tessman , N. 1968 . A Pliocene vertebrate fauna from low elevation in Manatee County, Florida . American Journal of Science , 265 : 777 – 811 .
- Whitmore , F. C. Jr. and Stewart , R. H. 1965 . Miocene mammals and Central American seaways . Science , 148 : 180 – 185 .
- Willis , P. M. A. 1993 . Trilophosuchus rackhami gen. et sp. nov., a new crocodilian from the early Miocene limestones of Riversleigh, northwestern Queensland . Journal of Vertebrate Paleontology , 13 : 90 – 98 .
- Witmer , L. M. 1997 . The evolution of the antorbital cavity of archosaurs: a study in soft-tissue reconstruction in the fossil record with an analysis of the function of pneumaticity . Society of Vertebrate Paleontology Memoir 3, 17 (supplement to number 1) , : 1 – 73 .
- Yu , Y. , Harris , A. J. and He , X.-J. 2010a . S-DIVA (Statistical Dispersal-Vicariance Analysis) 1.5 . http://mnh.scu.edu.cn/soft/blog/sdiva. Accessed September 2, 2011
- Yu , Y. , Harris , A. J. and He , X.-J. 2010b . S-DIVA (statistical dispersal-vicariance analysis): a tool for inferring biogeographic histories . Molecular Phylogenetics and Evolution , 56 : 848 – 850 .
- Zachos , J. C. , Dickens , G. R. and Zeebe , R. E. 2008 . An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics . Nature , 451 : 279 – 283 .
APPENDIX 1. The cladistic matrix of Brochu (Citation1999) was modified by eliminating parsimoniously uninformative characters and including only members of Alligatoridae and three outgroup members (Stangerochampsa mccabei, Brachychampsa montana, and B. rachychampsa sealyi). The matrix is now 75 characters from the original 164. For each character, the source(s) from which it was modified is given as well as the original character number.
| 1. | Hypapophyseal keels extend to: eleventh vertebra behind atlas (0); twelfth vertebra behind atlas (1); tenth vertebra behind atlas (2). From Brochu (Citation1999:char. 7). | ||||
| 2. | Proatlas: with prominent anterior process (0); lacks anterior process (1). From Brochu (Citation1999:char. 10). | ||||
| 3. | Anterior half of axis neural spine: oriented horizontally (0); slopes anteriorly (1). From Brochu (Citation1999:char. 11). | ||||
| 4. | Axis neural spine: crested (0); not crested (1). From Brochu (Citation1999:char. 12). | ||||
| 5. | Dorsal margin of atlantal rib: generally smooth with modest dorsal process (0); with prominent process (1). From Brochu (Citation1999:char. 14). | ||||
| 6. | Large articular facets at anterior ends of atlantal ribs: absent (0); present (1). From Brochu (Citation1999:char. 15). | ||||
| 7. | Very thin medial laminae at anterior end of atlantal ribs: absent (0); present (1). From Brochu (Citation1999:char. 16). | ||||
| 8. | Proatlas: with tall dorsal keel (0); lacks tall dorsal keel, dorsal side smooth (1). From Brochu (Citation1999:char. 17). | ||||
| 9. | Deep fork of axial hypapophysis: present (0); absent (1). From Brochu (Citation1999:char. 19). | ||||
| 10. | Scapular blade: flares dorsally at maturity (0); sides subparallel, minimal dorsal flare at maturity (1). From Brochu (Citation1999:char. 22); adapted from study by Benton and Clark (Citation1988). | ||||
| 11. | Deltoid crest of scapula: very thin at maturity, with sharp margin (0); very wide at maturity, with broad margin (1). From Brochu (Citation1999:char. 23). | ||||
| 12. | Scapulocoracoid synchondrosis closes: very late in ontogeny (0); relatively early in ontogeny (1). From Brochu (Citation1999:char. 24). | ||||
| 13. | Scapulocoracoid facet anterior to glenoid fossa: uniformly narrow (0); broad immediately anterior to glenoid fossa, and tapering anteriorly (1). From Brochu (Citation1999:char. 25). | ||||
| 14. | Dorsal margin of iliac blade: rounded with smooth border (0); rounded, with modest dorsal indentation (1); rounded, with strong dorsal indentation, ‘wasp-waisted’ (2); narrow, with dorsal indentation (3); rounded with smooth border, posterior tip of blade very deep (4). From Brochu (Citation1999:char. 28). | ||||
| 15. | Interclavicle: flat along length, without dorsoventral flexure (0); with moderate dorsoventral flexure (1); with severe dorsoventral flexure (2). From Brochu (Citation1999:char. 30). | ||||
| 16. | Anterior end of interclavicle: flat (0); rod-like (1). From Brochu (Citation1999:char. 31). | ||||
| 17. | Dorsal midline osteoderms: rectangular (0); square or equant (1). From Brochu (Citation1999:char. 36); adapted from Norell and Clark (Citation1990:char. 16) and Clark (Citation1994:char. 95). | ||||
| 18. | Contiguous dorsal osteoderms per row at maturity: four (0); six (1); eight (2); or ten 10 (3). From Brochu (Citation1999:char. 37); adapted from Norell and Clark (Citation1990:char. 12) and Clark (Citation1994:char. 97). | ||||
| 19. | Nuchal shield: grades continuously into dorsal shield (0); differentiated from dorsal shield, four nuchal osteoderms (1); differentiated from dorsal shield, six nuchal osteoderms with four central and two lateral (2); differentiated from dorsal shield, eight nuchal osteoderms in two parallel rows (3). From Brochu (Citation1999:char. 38). | ||||
| 20. | Ventral armor: absent (0); present, osteoderms single (1); present, osteoderms consist of paired ossifications that suture together (2). From Brochu (Citation1999:char. 39); adapted from Buscalioni et al. (Citation1992:char. 21). | ||||
| 21. | Splenial: with anterior perforation for mandibular ramus of cranial nerve V (0); lacks anterior perforation for mandibular ramus of cranial nerve V (1). From Brochu (Citation1999:char. 41); adapted from Norell (Citation1988:char. 15) and Norell (Citation1989:char. 8). | ||||
| 22. | Mandibular ramus of cranial nerve V exits splenial: anteriorly only (0); splenial has singular perforation for mandibular ramus of cranial nerve V posteriorly (1); splenial has double perforation for mandibular ramus of cranial nerve V posteriorly (2). From Brochu (Citation1999:char. 42); adapted from Norell (Citation1988:char. 15) and Norell (Citation1989:char. 8). | ||||
| 23. | Splenial: participates in mandibular symphysis, splenial symphysis adjacent to no more than five dentary alveoli (0); splenial excluded from mandibular symphysis, anterior tip of splenial passes ventral to Meckelian groove (1); splenial excluded from mandibular symphysis, anterior tip of splenial passes dorsal to Meckelian groove (2); deep splenial symphysis, longer than five dentary alveoli, splenial forms wide ‘V’ within symphysis (3); deep splenial symphysis, longer than five dentary alveoli, splenial constricted within symphysis and forms narrow ‘V’ (4). From Brochu (Citation1999:char. 43); adapted from Clark (Citation1994:char. 77). | ||||
| 24. | Articular-surangular suture: simple (0); with anterior process dorsal to lingual foramen (‘crocodyline process’ of Aoki, 1994) (1); with anterior process ventral to lingual foramen (2). From Brochu (Citation1999:char. 44). | ||||
| 25. | Lingual foramen for articular artery and alveolar nerve: on surangular entirely (0); on surangular-angular suture (1); on angular entirely (2). From Brochu (Citation1999:char. 45). | ||||
| 26. | Coronoid: bounds posterior half of foramen intermandibularis medius (0); completely surrounds foramen intermandibularis medius at maturity (1); obliterates foramen intermandibularis medius at maturity (2). From Brochu (Citation1999:char. 46); adapted from Norell (Citation1988:char. 12). | ||||
| 27. | Angular-surangular suture: contacts external mandibular fenestra at posterior angle at maturity (0); passes broadly along ventral margin of external mandibular fenestra late in ontogeny (1). From Brochu (Citation1999:char. 47); adapted from Norell (Citation1988:char. 40). | ||||
| 28. | Surangular: extends to posterior end of retroarticular process (0); pinched off anterior to tip of retroarticular process (1). From Brochu (Citation1999:char. 51); adapted from Norell (Citation1988:char. 42). | ||||
| 29. | Superior edge of coronoid: slopes strongly anteriorly (0); almost horizontal (1). From Brochu (Citation1999:char. 54). | ||||
| 30. | Inferior process of coronoid: laps strongly over inner surface of Meckelian fossa (0); remains largely on medial surface of mandible (1). From Brochu (Citation1999:char. 55). | ||||
| 31. | Coronoid: imperforate (0); with perforation posterior to foramen intermandibularis medius (1). From Brochu (Citation1999:char. 56). | ||||
| 32. | Dorsal projection of hyoid cornu: narrow, with parallel sides (0); flared (1). From Brochu (Citation1999:char. 58). | ||||
| 33. | Surangular spur bordering the dentary toothrow lingually for at least one alveolus length: present (0); absent (1). From Brochu (Citation1999:char. 61). | ||||
| 34. | External mandibular fenestra: small, foramen intermandibularis caudalis not visible laterally (0); large, foramen intermandibularis caudalis visible laterally (1). From Brochu (Citation1999:char. 64); adapted from Norell (Citation1988:char. 14). | ||||
| 35. | Surangular-dentary suture intersects external mandibular fenestra: anterior to posterodorsal corner (0); at posterodorsal corner (1). From Brochu (Citation1999:char. 65). | ||||
| 36. | Angular: extends dorsally toward or beyond anterior end of foramen intermandibularis caudalis, anterior tip acute (0); does not extend dorsally beyond anterior end of foramen intermandibularis caudalis, anterior tip very blunt (1). From Brochu (Citation1999:char. 66). | ||||
| 37. | Surangular-angular suture lingually meets articular: at ventral tip (0); dorsal to ventral tip (1). From Brochu (Citation1999:char. 67). | ||||
| 38. | Dentary between fourth and tenth alveoli: gently curved (0; deeply curved (1); linear (2). From Brochu (Citation1999:char. 68). | ||||
| 39. | Dentary teeth: all occlude lingual to maxillary teeth (0); occlusion pit present between seventh and eighth maxillary teeth, all other dentary teeth occlude lingually (1); occlude in line with maxillary toothrow (2). From Brochu (Citation1999:char. 78); adapted from Norel (Citation1988:char. 5) and Willis (Citation1993:char. 1). | ||||
| 40. | Naris projects: anterodorsally (0); dorsally (1). From Brochu (Citation1999:char. 79). | ||||
| 41. | Quadratojugal: extends to superior angle of infratemporal fenestra (0); does not extend to superior angle of infratemporal fenestra, quadrate participates in fenestra (1). From Brochu (Citation1999:char. 80); adapted from Buscalioni et al. (Citation1992:char. 6). | ||||
| 42. | Frontoparietal suture: deeply within supratemporal fenestra, frontal prevents broad contact between postorbital and parietal (0); with modest entry into supratemporal fenestra at maturity, postorbital and parietal in broad contact (1); suture on skull table entirely (2). From Brochu (Citation1999:char. 81). | ||||
| 43. | Supraoccipital exposure on dorsal skull table: small (0); absent (1); large (2); or large such that parietal is excluded from posterior edge of table (3). From Brochu (Citation1999:char. 82); adapted from Norell (Citation1988:char. 11). | ||||
| 44. | Frontoparietal suture: concavoconvex (0); linear (1). From Brochu (Citation1999:char. 86). | ||||
| 45. | Supratemporal fenestra: with fossa, dermal bones of skull roof do not overhang rim at maturity (0); with dermal bones of skull roof overhanging rim, near maturity (1); closes during ontogeny (2). From Brochu (Citation1999:char. 87); adapted from Norell (Citation1988:char. 9). | ||||
| 46. | Posterior notch of suborbital fenestra: absent (0); present (1). From Brochu (Citation1999:char. 88). | ||||
| 47. | Largest maxillary alveolus: m3 (0); m5 (1); m4 (2); m4 and m5 same size (3); maxillary teeth homodont (4). From Brochu (Citation1999:char. 89); adapted from Norell (Citation1988: char. 1). | ||||
| 48. | Lacrimal: makes broad contact with nasal, no posterior process of maxilla (0); maxilla sends posterior process within lacrimal (1); maxilla sends posterior process between lacrimal and prefrontal (2). From Brochu (Citation1999:char. 93). | ||||
| 49. | Lateral edges of palatines: smooth anteriorly (0); with lateral process projecting from palatines into suborbital fenestra (1). From Brochu (Citation1999:char. 94). | ||||
| 50. | External naris: bisected by nasals (0); nasals contact external naris, but do not bisect it (1); nasals excluded, at least externally, from naris, nasals and premaxillae still in contact (2); nasals and premaxillae not in contact (3). From Brochu (Citation1999:char. 95); adapted from Norell (Citation1988:char. 3) and Clark (Citation1994:chars. 13 and 14). | ||||
| 51. | Palpebral forms from: single ossification (0); multiple ossifications (1). From Brochu (Citation1999:char. 96); adapted from Norell (Citation1988:char. 8) and Clark (Citation1994:char. 65). | ||||
| 52. | Premaxilla early in posthatching ontogeny has: five teeth (0); four teeth (1). From Brochu (Citation1999:char. 97); adapted from Norell (Citation1988:char. 17). | ||||
| 53. | Prefrontal pillar: solid (0); with large pneumatic sinus (prefrontal recess of Witmer, Citation1997) (1). From Brochu (Citation1999:char. 99). | ||||
| 54. | Prefrontals: separated by frontals and nasals (0); meet medially (1). From Brochu (Citation1999:char. 100); adapted from Norell (Citation1988:char. 27). | ||||
| 55. | Margin of orbit: flush with skull surface (0); dorsal edges of orbits upturned (1); orbital margin telescoped (2). From Brochu (Citation1999:char. 103). | ||||
| 56. | Medial parietal wall of supratemporal fenestra: imperforate (0); bearing foramina (1). From Brochu (Citation1999:char. 104) and Norell (Citation1988:char. 51). | ||||
| 57. | Lateral edge of suborbital fenestra: straight (0); bowed medially (1). From Brochu (Citation1999:char. 105). | ||||
| 58. | Surangular: continues to dorsal tip of lateral wall of glenoid fossa (0); truncated and not continuing dorsally (1). From Brochu (Citation1999:char. 106). | ||||
| 59. | Posterior rim of internal choanae: not deeply notched (0); deeply notched (1). From Brochu (Citation1999:char. 107). | ||||
| 60. | Anterior face of palatine process: rounded or pointed anteriorly (0); invaginate (1). From Brochu (Citation1999: char. 108). | ||||
| 61. | Laterosphenoid bridge: composed entirely of laterosphenoid (0); with ascending process of pterygoid (1). From Brochu (Citation1999:char. 115). | ||||
| 62. | Ectopterygoid-pterygoid flexure: disappears during ontogeny (0); remains throughout ontogeny (1). From Brochu (Citation1999:char. 116). | ||||
| 63. | Lacrimal: longer than prefrontal (0); shorter than prefrontal (1); same length as prefrontal and both are elongate (2). From Brochu (Citation1999:char. 117); modified from Norell (Citation1988:char. 7). | ||||
| 64. | Incisive foramen: small, less than half the greatest width of premaxilla (0); large, more than half the greatest width of premaxilla (1); large, and intersects premaxillary-maxillary suture (2). From Brochu (Citation1999:char. 124). | ||||
| 65. | Vomer: entirely obscured by premaxilla and maxilla (0); exposed on palate at premaxillary-maxillary suture (1). From Brochu (Citation1999:char. 125); adapted from Norell (Citation1988:char. 22). | ||||
| 66. | Premaxillary surface lateral to naris: smooth (0); with deep notch lateral to naris (1). From Brochu (Citation1999:char. 142). | ||||
| 67. | Canthi rostralii at maturity: absent or very modest (0); very prominent (1). From Brochu (Citation1999:char. 143); adapted from Norell (Citation1988:char. 34). | ||||
| 68. | Dorsal premaxillary processes: short, not extending beyond third maxillary alveolus (0); long, extending beyond third maxillary alveolus (1). From Brochu (Citation1999:char. 145). | ||||
| 69. | Exoccipital: terminates dorsal to basioccipital tubera (0); sends robust process ventrally and participates in basioccipital tubera (1); sends slender process ventrally to basioccipital tubera (2). Modified from Brochu (Citation1999:char. 151); adapted from Norell (Citation1988:char. 20) and Clark (Citation1994:chars. 57 and 60). | ||||
| 70. | Internal choanae: not septate (0); with septum that remains recessed within choana (1); with septum that projects out of choana (2). From Brochu (Citation1999:char. 152). | ||||
| 71. | Incisive foramen: completely situated far from premaxillary toothrow, at the level of the second or third alveolus (0); abuts premaxillary toothrow (1); projects between first premaxillary teeth (2). From Brochu (Citation1999:char. 153). | ||||
| 72. | Ventral collar scales: not enlarged relative to other ventral scales (0); in a single enlarged row (1); in two parallel enlarged rows (2). From Brochu (Citation1999:char. 156); adapted from Poe (Citation1996:char. 115). | ||||
| 73. | Median pelvic keel scales: form two parallel rows along most of tail length (0); form single row along tail (1); merge with lateral keel scales to form ‘Y’-shaped keel (2). From Brochu (Citation1999:char. 157); adapted from Poe (Citation1996:char. 116). | ||||
| 74. | Naris: circular or keyhole-shaped (0); wider than long (1). From Brochu (Citation1999:char. 161). | ||||
| 75. | Postorbital-squamosal suture ventral to skull table: oriented ventrally (0); passes medially (1). From Brochu (Citation1999:char. 163). | ||||