ABSTRACT
Fishers are elusive carnivorans, with few occurrences in the fossil record. The origin and early evolution of fishers is unclear, but they likely originated in Asia. A new record of Pekania from the Rattlesnake Formation of Oregon represents the earliest known occurrence of a fisher, more than 5 million years earlier than other records in North America. This specimen has an unambiguous derived trait shared with other members of the genus, an external median rootlet on the upper carnassial. The age of this new find is inferred to be between 7.05 and 7.3 Ma, through radiometric and magnetostratigraphic dating. This age is supported by the presence of specimens of a shrew, Sorex edwardsi, and a rhinoceros, cf. Teleoceras, found at the same locality, as well as a rabbit, Hypolagus cf. vetus, and a tapir nearby, all of which are well known from early Hemphillian deposits. This find indicates that fishers were in both North America and Asia in the late Miocene, around the time of their divergence from other members of the clade as estimated from genetic data. Although it is over 7 million years old, this species shows remarkable similarity to extant P. pennanti, highlighting the highly conservative nature of gulonine mustelids. The Rattlesnake specimen is more robust than other fisher species, possibly representing something close to the ancestry of all fishers.
INTRODUCTION
The extant fisher, Pekania pennanti, is an elusive medium-sized mustelid endemic to North America. Fishers are generalized predators and good climbers, favoring forest habitats with continuous canopy (Powell, Citation1981). The subfamily Guloninae, which includes the fisher, as well as martens, the wolverine, and the tayra, is distributed over much of North America and Eurasia today. This subfamily was formerly referred to as the Martinae, but we follow Sato et al. (Citation2009) in using the name Guloninae, which has priority according to the International Code of Zoological Nomenclature. The gulonine clade of mustelids is known for having conservative morphology (apart from Gulo), making morphological studies of their relationships particularly difficult (Anderson, Citation1970, Citation1994). Recent genetic studies (Hosoda et al., Citation2000; Stone and Cook, 2002; Koepfli et al., Citation2008; Sato et al., Citation2009; Wolsan and Sato, Citation2010; Nyakatura and Bininda-Emonds, Citation2012; Sato et al., Citation2012) have shown that the fisher (formerly Martes pennanti) is more closely related to the tayra (Eira barbara) and wolverine (Gulo gulo) than to other species of the genus Martes, and was thus reclassified in the genus Pekania (Koepfli et al., Citation2008; Sato et al., Citation2012).
Gulonine mustelids are known from the fossil records of North America and Eurasia, but primarily from fragmentary remains that have been described as local species (including Schlosser, Citation1924; Zdansky, Citation1924; Gidley, 1927; Colbert, Citation1935; Helbing, Citation1936; Orlov, Citation1941; Teilhard and Leroy, Citation1945; Dehm, Citation1950; Viret, Citation1951; Ginsburg, Citation1961; Wilson, 1968; Mein and Ginsburgh, Citation1997; Stevens and Stevens, 2003; Nagel et al., Citation2009; Koufos, Citation2011). Anderson (Citation1994) suggested that the earliest records of this clade include specimens referred to the genus Martes, from the early Miocene of Germany (Dehm, Citation1950). However, Sato et al. (Citation2003) indicated that this ‘Martes’ laevidens is not a member of that genus. Members of the Guloninae spread across Eurasia, and may have immigrated to North America from Asia in the late Miocene (Sato et al., Citation2012). The earliest records of marten-like mustelids in North America include Dinogale siouxensis from the late Hemingfordian–aged Sheep Creek Formation of Nebraska (Cook and Macdonald, Citation1962), ‘Martes’ glareae from the early Barstovian–aged Olcott Formation of Nebraska (Sinclair, 1915), ‘Martes’ gazini from the early Barstovian Butte Creek Volcanic Sandstone Formation of Oregon (Hall, 1931), and ‘Martes’ stirtoni from the Clarendonian-aged Ogallala Formation of Kansas (Wilson, 1968). However, some authors have synonymized these taxa with Plionictis or other ischyrictines, considering them outside of the Guloninae (Sutton 1977; Anderson, Citation1994; Baskin, Citation1998). Anderson (Citation1994) believed many fossil taxa referred to ‘Martes’ were not martens and not closely related to the ancestry of extant taxa. She considered the extant American marten, Martes americana, to be a late Pleistocene immigrant to North America (Anderson, Citation1994). Sato et al. (Citation2003) suggested that the oldest true Martes known is Martes wenzensis from the Pliocene of Poland (Wolsan, Citation1989).
Fisher fossils are particularly rare. Two species have been described from the late Miocene and Pliocene of Asia, Martes palaeosinensis and M. anderssoni (Schlosser, Citation1924; Zdanzky, Citation1924; Anderson, Citation1970). Anderson (Citation1994) indicated that both probably represented a single species, although CitationWang et al. (2012) question whether M. anderssoni is truly related to the fisher clade. The earliest fishers known from North America are M. diluviana (Cope, 1899) and M. parapennanti (Gidley and Gazin, 1933) from the Irvingtonian of the eastern United States, which were later synonymized as M. diluviana (Hall, 1936). Both M. palaeosinensis and M. diluviana were referred by Anderson (Citation1970, Citation1994) to the subgenus Pekania. Given the morphological distinctness of these species from martens (Martes), and following the recommendations Koepfli et al. (Citation2008) and Sato et al. (Citation2012) made based upon their genetic divergence, we refer them to the genus Pekania as P. palaeosinensis and P. diluviana, as did CitationWang et al. (2012).
The specimen described here, from the late early Hemphillian– aged Rattlesnake Formation of Oregon, represents the earliest dated record of a fisher and predates other known fishers in North America by over 5 million years. This species likely represents an early immigrant from Asia that may be close to the ancestor of other fossil and extant fishers. Given the rarity of fishers in modern ecosystems and in the fossil record, this find greatly improves our understanding of the evolution and biogeography of fishers.
METHODS
Measurements of the teeth, to the nearest 0.1 mm, were made using Mitutoyo Absolute digital calipers. Measurements were taken at the enamel dentine junction surface of the teeth and included anteroposterior length and transverse breadth; descriptions of measurements are provided in Appendix 1. Comparisons were made with specimens of extant Pekania and Martes species from the United States National Museum of Natural History, Natural History Museum of Los Angeles County, and Donald R. Dickey Collection of the University of California, as well as photographs and measurements in several publications (including Zdansky, Citation1924; Gidley and Gazin, 1933; Anderson, Citation1970; CitationWang et al., 2012).
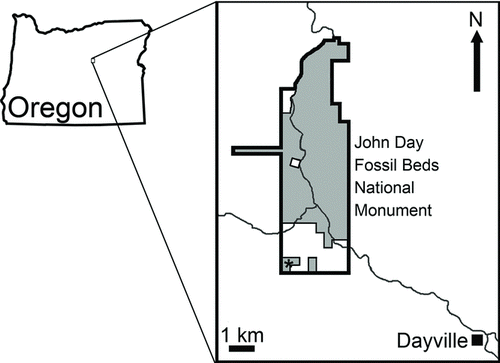
FIGURE 1 Map showing the Sheep Rock unit of John Day Fossil Beds National Monument, Grant County, Oregon. The area where the specimen described here was found is indicated by an asterisk.
Specimen Repositories
JODA, John Day Fossil Beds National Monument, Kimberly, Oregon; LACM, Natural History Museum of Los Angeles County, Los Angeles, California; UCLA, Donald R. Dickey Collection of the University of California, Los Angeles, Los Angeles, California; USNM, United States National Museum of Natural History, Washington, D.C.
GEOLOGICAL SETTING
John Day Fossil Beds National Monument (JODA) was created to help protect key fossil deposits within the John Day Basin of eastern Oregon. The John Day Basin contains hundreds of fossil localities spanning the late Eocene through the end of the Miocene; the stratigraphically highest unit exposed in the monument is the Rattlesnake Formation. The area south of Picture Gorge and west of Dayville, Oregon () was first examined by Merriam (Citation1901), and made the type area of the Rattlesnake Formation by Merriam et al. (Citation1925). This area was later studied by Enlows (Citation1976) and Martin (Citation1983, Citation1996). When the Hemphillian Land Mammal Age was originally designated, the Rattlesnake Formation was chosen as a reference fauna (Wood et al., Citation1941). The Hemphillian age of the Rattlesnake fauna has been confirmed by several studies (Martin, Citation1983, Citation1996; Fremd et al., Citation1994; Tedford et al., Citation2004; Samuels and Zancanella, Citation2011). In the area west of Dayville, the Rattlesnake Formation unconformably overlies the Mascall Formation, which is of Barstovian (middle Miocene) age.
The Rattlesnake Formation type area geology consists mainly of fluvial deposits, including siltstones, sandstones, and conglomerates (Enlows, Citation1976; Martin, Citation1996; Martin and Fremd, Citation2001). Specimens described here were found as surface float on a small exposure of reddish-brown siltstone, on the east face of a gently sloping northeast-trending ridge. The Rattlesnake Formation has many small isolated exposures with little exposed stratigraphy, making it difficult to correlate their precise position within a stratigraphic section. The prominent Rattlesnake Ash Flow Tuff (RAFT), which is widely distributed across central Oregon, has been Ar40/Ar39 dated to 7.05 ± 0.01 Ma (Streck and Grunder, Citation1995). Although the RAFT is not exposed in the Rattlesnake type area, regional correlation confirms this site lies well below that unit. The site lies just above a second marker bed within the lower Rattlesnake Formation, an undated white vitric tuff (‘Ted's Tuff’). This tuff, which lies approximately 16 m above the base of the section (Retallack, Citation1999; Retallack et al., Citation2002), is exposed on a small exposure 70 m southeast of where the specimens were found. Prothero et al. (Citation2006) did a paleomagnetic stratigraphy study of the type Rattlesnake section, correlating it with magnetic Chrons C3Bn to C3Br2n (6.9–7.3 Ma). Multiple lines of evidence thus confirm a late early Hemphillian age for the entire formation (Martin, Citation1983, Citation1996; Prothero et al., Citation2006).
Based upon Merriam's description of the type Rattlesnake area, Enlows (Citation1976) divided the Formation into three members: a lower fanglomerate member, the Rattlesnake Ash Flow Tuff (called Rattlesnake Ignimbrite), and an upper fanglomerate member. Walker (Citation1979, Citation1990) redefined the Rattlesnake Formation as only including the Rattlesnake Ash Flow Tuff, effectively orphaning the upper and lower fanglomerate members, and the biostratigraphically important associated fauna (Wood et al., Citation1941). More recent authors have suggested treatment of the Rattlesnake Formation as including three members (Martin, Citation1996) or elevating it to group status with three formations (Martin and Fremd, Citation2001). Like a variety of other recent studies (e.g., Retallack et al., Citation2002; Prothero et al., Citation2006; Samuels and Zancanella, Citation2011), we use the broader definition of Rattlesnake Formation suggested by Martin (Citation1996).
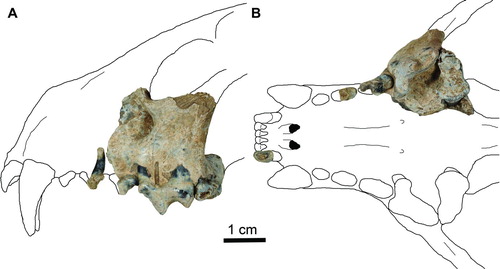
FIGURE 2 Holotype of Pekania occulta (JODA 15214) from the Rattlesnake Formation, Oregon. Specimen consists of a left maxilla with partial P3, P4, and M1, and partial left P2 and right I3. A, left lateral view; B, occlusal view. Scale bar equals 1 cm
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, Citation1758
Order CARNIVORA Bowditch, Citation1821
Family MUSTELIDAE Fischer von Waldheim, Citation1817
Subfamily GULONINAE Gray, 1825
Genus PEKANIA Gray, Citation1865
Type Species
Pekania pennanti Erxleben, 1777.
Included Species
P. occulta, new species; P. diluviana (Cope, 1899); P. palaeosinensis (Zdansky, Citation1924); P. pennanti Erxleben, 1777.
Emended Diagnosis
A synapomorphy that unites species of Pekania is a P4 with a prominent exposed external median rootlet (Anderson, Citation1970).
Distribution
Early Hemphillian (late Miocene) of Oregon; late Miocene and Pliocene of China and Mongolia; Irvingtonian of Arkansas, Colorado, Pennsylvania, Maryland, and West Virginia; Rancholabrean (late Pleistocene) of eastern United States; historic range included northern forests of North America, extending south in the Appalachian mountains and Pacific Coast range, Cascades, and Rocky Mountains.
PEKANIA OCCULTA, sp. nov.
(, )
Holotype
JODA 15214 (, ): left maxilla with partial P3, P4, and M1, isolated partial left P2 and right I3. The maxilla was collected by J. Cavin on 8 December 2011; the M1 and a portion of the P3, which had broken off of the maxilla, were collected on 13 January 2012 by J. Samuels and C. Schierup.
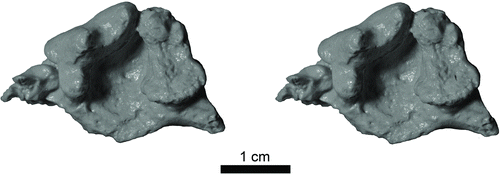
FIGURE 3 Stereophotograph of grayscale cast of Pekania occulta holotype (JODA 15214). Scale bar equals 1 cm.
Type Locality
Lower fanglomerate member of the Rattlesnake Formation, west of Dayville, Grant County, Oregon; approximately 793 m above mean sea level. This area lies within the type area of Merriam, on land administered by the National Park Service within the Sheep Rock Unit of John Day Fossil Beds National Monument (). Precise locality information, including GPS (global positioning satellite) data, is on file at JODA.
Referred Specimens
Known from holotype only.
Diagnosis
Pekania occulta has several features that distinguish it from other members of the genus: more robust dentition; enlarged P4 protocone, anteriorly positioned with weak inflection between protocone and parastyle; reduced M1 metacone and posteroexternal cingulum; distinct M1 shape characterized by narrow external lobe and broad internal lobe.
Distribution
Type locality only, early Hemphillian of Oregon.
Etymology
From the Latin, occulta, meaning ‘hidden’ or ‘secret,’ in reference to the long hidden origin of extant fishers and their elusive nature.
Description and Comparisons
The specimens consist of a left maxilla (, ), which has an incomplete P3 and complete P4 and M1, as well as a partial left P2 and right I3 found nearby (). The teeth show little wear, suggesting that this was a young adult individual; however, there is some substantial erosion to the tooth enamel, particularly in the M1. The maxilla is similar to that of the extant Pekania pennanti, with a round infraorbital foramen 5.6 mm in diameter (P. pennanti mean = 5.09 mm). The base of the orbit is preserved, lying directly above the infraorbital foramen and anterior root of the P4; the orbit is slightly elevated compared with P. pennanti and P. palaeosinensis. The maxillary portion of the zygomatic arch rises sharply above the M1, suggesting strongly curved arch more like P. pennanti than P. palaeosinensis.
The isolated right I3 has a broken surface, precluding detailed description of its morphology. The isolated left P2 is missing the anterior portion of the tooth, but clearly double-rooted. There is a weak cingulum at the posterior margin of the P2, along with a small but distinct ridge running down the midline of the tooth. Although incomplete, the posterior portion of the P3 in the maxilla is intact enough to observe that it has two roots and a single primary cusp. Like the P2, there is a cingulum at the posterior margin of the P3 and a distinct ridge running down the midline of the tooth. The preserved portions of the P2 and P3 in Pekania occulta are relatively robust, noticeably more robust than the narrow premolars of P. palaeosinensis ().
TABLE 1 Dental measurements (in mm) of Pekania and Martes species.
The P4 is very robust, with a prominent exposed external median rootlet, as is characteristic of the genus Pekania (Anderson, Citation1970) (, ). This rootlet is mostly missing, but the broken base and exposed alveolus clearly indicate its presence. The P4 protocone is large compared with other species of Pekania, and anteriorly positioned relative to P. pennanti, extending as far anteriorly as the parastyle. The shape of the anterior portion of the tooth is similar to P. palaeosinensis and the Pleistocene P. diluviana, which also have an anteriorly positioned protocone. However, both of those species have a deeper inflection of the anterior portion of the tooth between the parastyle and protocone; the inflection is weak in P. occulta and the posterior portion of the P3 essentially occupies this space. This position of the P3 between the P4 parastyle and protocone is also seen in individuals of P. palaeosinensis (Zdansky, Citation1924), as well as Martes martes, M. flavigula, and Gulo gulo. In P. pennanti, P. diluviana, and Martes Americana, the posterior portion of the P3 is adjacent to the P4 parastyle. The P4 proportions of the Rattlesnake specimen are more robust than is seen in other Pekania species, with a particularly large protocone and wide paracone and metacone (). There is a slightly worn shear facet between the P4 paracone and metacone, but this is the only real evidence of tooth wear in this individual.
The M1 shape is similar to other gulonines, with the expanded inner lobe characteristic of Martes and Pekania (, ). The M1 external roots are distinctly separated, with the posterior root located medially relative to the anterior root; this is more similar to the characteristics of P. palaeosinensis and P. pennanti than P. diluviana. The M1 of P. occulta is anteriorly constricted between the inner and outer lobes, and the inner lobe is rounded as in P. pennanti, rather than having a hypocone with a square shape as is seen in P. palaeosinensis. The surface of the M1 is eroded and pitted, but the paracone and metacone are small; there is an expanded cingular shelf labial to both cusps, larger anteriorly. Compared with P. pennanti, the metacone is reduced, as is the posteroexternal cingulum. The protocone ridge is crescentic in shape and much narrower than the rounded and expanded hypoconal cingulum. The orientation of the M1 in P. occulta also differs slightly from P. pennanti and P. palaeosinensis, with its occlusal surface parallel to that of the other maxillary teeth. In the later taxa, the posterior portion of the M1 is elevated, resulting in a more acute angle between the occlusal surfaces of the M1 and P4. The M1 proportions of P. occulta differ from other Pekania species, with a particularly narrow external lobe and wide internal lobe (). The ratio of internal to external lobe length (M1Lint/M1Lext) for P. occulta is 0.65, whereas all other Pekania sampled have values between 0.76 and 0.91.
DISCUSSION
The fisher from the Rattlesnake Formation is only represented by a single specimen, but is readily differentiable from known specimens of extant and extinct species of Pekania. The morphologies of the teeth in P. occulta are distinct, most notably the robust P4 protocone and shape of the M1. Additionally, the proportions of the teeth in P. occulta fall outside the range of variation for extant P. pennanti for nearly all measurements. P. occulta shows particularly robust teeth when compared with extant Pekania and Martes, potentially reflecting similarity to a shared ancestor with the wolverine clade (Gulo).
This find represents the earliest known and dated occurrence of Pekania in the world, predating the former earliest record of the genus in North America by more than 5 million years. Because many of the early Eurasian species previously referred to ‘Martes’ likely do not belong to the Guloninae (Anderson, Citation1994; Sato et al., Citation2003), Pekania occulta also represents the earliest known well-dated gulonine. The position of this locality within the Rattlesnake Formation, relative to a dated ash bed and magnetostratigraphy, indicates a late early Hemphillian age, between 7.3 and 7.05 Ma. The presence of a shrew dentary, Sorex edwardsi, and rhinocerotid tooth fragments, cf. Teleoceras, at the site supports a Hemphillian age. Additionally, several other taxa well known from Hemphillian-aged deposits were found on exposures nearby, including a nearly complete rabbit dentary, Hypolagus cf. vetus, and a partial tapir tooth. Teleoceras and Hypolagus are both commonly found within Rattlesnake Formation deposits (Merriam et al., Citation1925; Martin, Citation1983, Citation1996). Although none of these taxa are restricted to the Hemphillian, the occurrence of all of these taxa within the same horizon as P. occulta supports the inferred early Hemphillian age of the site.
This record from Oregon indicates that fishers were in North America by at least 7.05 Ma, either evolving here or emigrating from Asia. Pekania occulta is morphologically distinct from other fisher species in North America and Asia, with both P. palaeosinensis and P. diluviana more similar to the extant P. pennanti (Anderson, Citation1970, Citation1994; CitationWang et al., 2012). This suggests that P. occulta did not directly lead to P. pennanti in North America, but probably displays features close to the ancestor of all fishers. Ancestral-state reconstructions based on molecular data (Koepfli et al., Citation2008; Sato et al., Citation2012) suggest an Old World origin for this clade. Sato et al. (Citation2012) inferred Asia as the ancestral area for the clade through parsimony and Bayesian inference, whereas maximum likelihood favored Asia and North America. Given the presence of other marten-like mustelid fossils earlier in Europe and Asia (Dehm, Citation1950; Sotnikova, Citation1995; Qiu, Citation2003; Nagel, Citation2009; CitationWang et al., 2012), it is likely that Pekania evolved somewhere in Eurasia and P. occulta represents an early immigrant to North America. Most records of P. palaeosinensis from Asia are not well dated, but a new record from the Baodean-aged Baogeda Ula Fauna of Mongolia is likely similar in age to P. occulta from the Rattlesnake Formation (CitationWang et al., 2012). Although that fauna has not been directly dated, that region of Mongolia has a number of dated basalt layers and one that may cap the Baogeda Ula Fauna may be 7.11 ± 0.48 Ma (CitationWang et al., 2012). This is roughly the same age (or slightly younger) than the Rattlesnake Formation, clearly indicating that Pekania was present on both continents in the late Miocene.
In the late early Hemphillian, when Pekania may have dispersed to North America, several other carnivoran species immigrated to North America from Asia (Tedford et al., Citation1987; Qiu, Citation2003; Tedford et al., Citation2004; Woodburne, Citation2004). Another mustelid (Lutravus), as well as a bear (Indarctos) and an ailurid (Simocyon), have all been found in the Rattlesnake Formation (Merriam et al., Citation1916, 1925; Thorpe, Citation1921), and are known from similar-aged (Baodean) deposits in Asia (Zdansky, Citation1924; Qiu, Citation2003). The Baogeda Ula Fauna from Mongolia has Hipparion and Dipoides co-occurring with Pekania (CitationWang et al., 2012); both of these taxa are common in the Rattlesnake Formation (Merriam et al., Citation1925). The beaver, Castor, was also recently found in the Rattlesnake Formation, illustrating its arrival on this continent at the same time (Samuels and Zancanella, Citation2011). Oregon has some of the Hemphillian-aged sites located nearest to the land bridge between North America and Asia, therefore, similarity to Asian faunas and early records of immigrant taxa from Oregon should not be surprising (Shotwell, 1956).
Several recent molecular studies have estimated the divergence dates of the Guloninae from other mustelids. In a study using an extensive sampling of 22 gene segments, Koepfli et al. (2008) estimated that this clade diverged from other mustelids around 11.0 Ma (95% confidence interval [CI]: 9.4–12.5 Ma) and Pekania diverged around 6.4 Ma (95% CI: 4.7–8.0 Ma). Yu et al. (Citation2011) used 17 nuclear gene segments and mitochondrial genome data, yielding estimates for the origin of the Guloninae of 5.97 Ma (95% CI: 3.39–8.90 Ma) and 10.91 Ma (95% CI: 7.82–14.37 Ma), respectively. Sato et al. (Citation2012) examined nine nuclear genes and one mitochondrial gene, estimating divergence of the Guloninae from other mustelids at 12.65 Ma (95% CI: 10.83–14.72 Ma) and divergence of Pekania at 7.90 Ma (95% CI: 6.41–9.64 Ma) using Multidivtime analysis. The age of Pekania occulta is very close to these estimates based on molecular data, and the inclusion of P. occulta as a calibration point in future studies would help yield better estimates of the timing of splits within the Mustelidae. Late Miocene records of Pekania correspond well with the timing of diversification within multiple groups of carnivorans based upon molecular estimates (Koepfli et al., Citation2008; Eizirik et al., Citation2010; Sato et al., Citation2012); 7.3 Ma was actually estimated to be a peak in net diversification rate for carnivorans by Nyakatura and Bininda-Emonds (Citation2012).
Today, fishers (Pekania pennanti) occur across the northern United States and southern Canada. Fishers strongly prefer dense forests with continuous canopy (Powell, Citation1981). Oregon in the Hemphillian has been interpreted as representing tall grassland and shrubland with faunas typical of open habitats (Fremd et al., Citation1994). Retallack et al. (Citation2002) studied paleosols in the Rattlesnake Formation, interpreting the base of the section as riparian woodlands and meadows, and, following aridification through the section, a shift to tall grassland and semiarid wooded shrubland environments. Commonly found cursorial grazers (equids: Pliohippus and Hipparion; antilocaprids: Sphenophalos) and burrowing mammals (Spermophilus and Scapanus) (Merriam et al., Citation1925; Martin, Citation1996) support reconstruction of the Rattlesnake Formation with open paleoenvironments. However, boreal organisms such as Pekania and rare petrified wood fragments (Quercus sp.) suggest the persistence of a heterogenous environment, with some extensive forested areas. These may have existed in riparian areas, which were common, as indicated by paleosol evidence and large root traces (Retallack et al., Citation2002) and the abundance of the beaver Dipoides stirtoni in Rattlesnake Formation deposits.
ACKNOWLEDGMENTS
The following curators and collection managers kindly allowed access to specimens in their care: J. Dines (National History Museum of Los Angeles County), K. Molina (Donald R. Dickey Collection of the University of California, Los Angeles), L. Gordon (National Museum of Natural History), and C. Conroy (Museum of Vertebrate Zoology, University of California, Berkeley). K. Maguire kindly assisted us by providing measurements of specimens at the MVZ. Discussion with and comments by X. Wang greatly improved this paper; he graciously shared information on Pekania from Asia, including photos and a cast, and he offered very helpful suggestions. C. Schierup (John Day Fossil Beds National Monument) and J. Zancanella (Bureau of Land Management) assisted with field work. We thank G. Rougier and J. Harris for editorial assistance, as well as M. Wolsan and two anonymous reviewers for their helpful comments and suggestions.
Handling editor: Guillermo Rougier
LITERATURE CITED
- Anderson , E. 1970 . Quaternary evolution of the genus Martes (Carnivora, Mustelidae) . Acta Zoologica Fennica , 130 : 1 – 132 .
- Anderson , E. 1994 . “ Evolution, prehistoric distribution and systematics of Martes ” . In Martens, Sables and Fishers: Biology and Conservation , Edited by: Buskirk , S. W. , Harestad , A. S. , Raphael , M. G. and Powell , R. A. 13 – 25 . Ithaca , New York : Cornell University Press .
- Baskin , J. A. 1998 . “ Mustelidae; pp ” . In Evolution of Tertiary Mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates, and Ungulate-like Mammals , Edited by: Janis , C. M. , Scott , K. M. and Jacobs , L. L. 152 – 173 . Cambridge , , U.K : Cambridge University Press .
- Bowditch , T. E. 1821 . An Analysis of the Natural Classifications of Mammalia for the Use of Students and Travelers , 151 Paris , , France : J. Smith .
- Burmeister , G. 1850 . Verzeichnis der im Zoologischen Museum der Universität Halle-Wittenberg aufgestellten Säugethiere, Vögel und Amphibien , 84 Halle, Friedrichs- Universität .
- Colbert , E. H. 1935 . Siwalik mammals in the American Museum of Natural History . Transactions of the American Philosophical Society, Philadelphia , 26 : 1 – 401 . New Series
- Cook , H. J. and Macdonald , J. R. 1962 . New Carnivora from the Miocene and early Pliocene of western Nebraska . Journal of Paleontology , 36 : 560 – 567 .
- Cope , E. D. 1899 . Vertebrate remains from Port Kennedy bone deposit . Journal Academy of Natural Sciences of Philadelphia , 11 : 193 – 267 .
- Dehm , R. 1950 . Die Raubtiere aus dem Mittel-Miocän (Burdigalium) von Winstershof-West bei Eichstätt in Bayern , Vol. 58 , 1 – 141 . Neue Folge : Abhandlungen der Bayerischen Akademi der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse .
- Enlows , H. E. 1976 . Petrography of the Rattlesnake Formation at the type area, central Oregon . Department of Geology and Mineral Industries, State of Oregon, Oil and Gas Investigations , 25 : 1 – 34 .
- Eizirik , E. , Murphy , W. J. , Koepfli , K. P. , Johnson , W. E. , Dragoo , J. W. , Wayne , R. K. and O’Brien , S. J. 2010 . Pattern and timing of diversification of the mammalian order Carnivora inferred from multiple nuclear gene sequences . Molecular Phylogenetics and Evolution , 56 : 49 – 63 .
- Erxleben , J. C. P. 1777 . “ Systema Regni Amimalis, per Classes, Ordines, Genera, Species, Varietates, cum Synonymia et Historia Animalium. Classis I, Mammalia ” . 636 Weygand : Leipzig .
- Fischer von Waldheim , G. 1817 . Adversaria zoologica . Mémoires de la Société Imperiale des Naturalistes de Moscou , 5 : 357 – 472 .
- Fremd , T. J. , Bestland , E. A. and Retallack , G. J. 1994 . John Day Basin paleontology field trip guide and road log. Society of Vertebrate Paleontology, 1994 Annual Meeting. Seattle, Northwest Interpretive Association . : 80
- Gidley , J. W. 1927 . A true marten from the Madison Valley (Miocene) of Montana . Journal of Mammalogy , 8 : 239 – 242 .
- Gidley , J. W. and Gazin , C. L. 1933 . New Mammalia in the Pleistocene Fauna from Cumberland Cave . Journal of Mammalogy , 14 : 343 – 357 .
- Ginsburg , L. 1961 . La faune des carnivores Miocènes de Sansan (Gers) . Mémoires du Muséum National d’Histoire Naturelle Série C Geologie , 9 : 1 – 187 .
- Gray , J. E. 1821 . On the natural arrangement of vertebrose animals . The London Medical Repository Monthly Journal and Review , 15 : 296 – 310 .
- Gray , J. E. 1865 . Revision of the genera and species of Mustelidae contained in the British Museum . Proceedings of the Zoological Society of London , 1865 : 100 – 154 .
- Helbing , H. 1936 . Die carnivoren des Steinheimer beckens, A. Mustelidae . Palaeontographica Beiträge zur Naturgeschichte der Vorzeit , 5 : 1 – 56 .
- Hemprich , W. 1820 . Grundriss der Naturgeschichte für höhere Lehranstalten , 432 Berlin : August Rucker .
- Hosoda , T. , Suzuki , H. , Harada , M. , Tsuchiya , K. , Han , S.-H. , Zhang , Y.-p. , Kryukov , A. P. and Lin , L.-K. 2000 . Evolutionary trends of the mitochondrial lineage differentiation in species of genera Martes and Mustela . Genes & Genetics Systems , 75 : 259 – 267 .
- Koepfli , K.-P. , Deere , K. A. , Slater , G. J. , Begg , C. , Begg , K. , Grassman , L. , Lucherini , M. , Veron , G. and Wayne , R. K. 2008 . Multigene phylogeny of the Mustelidae: resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation . BMC Biology , 6 : 10
- Koufos , G. D. 2011 . The Miocene carnivore assemblage of Greece . Estudios Geologicos , 67 : 291 – 320 .
- Kurtén , B. 1970 . The Neogene wolverine Plesiogulo and the origin of Gulo (Carnivora, Mammalia) . Acta Zoologica Fennica , 131 : 1 – 22 .
- Kurtén , B. and Anderson , E. 1980 . Pleistocene Mammals of North America , 442 New York : Columbia University Press .
- Linnaeus , C. 1758 . Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, 1758 , Stockholm : Societatis Zoologicae Germanicae .
- Martin , J. E. 1983 . Additions to the early Hemphillian (Miocene) Rattlesnake Fauna from central Oregon . Proceedings of the South Dakota Academy of Science , 62 : 23 – 33 .
- Martin , J. E. 1996 . Investigation of the late Miocene (Hemphillian) Rattlesnake Formation on lands administered by the Bureau of Land Management, Picture Gorge area, central Oregon . : 22 Unpublished report, Bureau of Land Management, Prineville District Office
- Martin , J. E. and Fremd , T. J. 2001 . Revision of the lithostratigraphy of the Hemphillian Rattlesnake units of central Oregon . PaleoBios , 21 : 89
- McKenna , M. C. and Bell , S. K. 1997 . Classification of Mammals above the Species Level , 631 New York : Columbia University Press .
- Mein , P. and Ginsburg , L. 1997 . Les mammifères du gisement miocène inférieur de Li Mae Long, Thaïlande: systématique, biostratigraphie et paléoenvironnement . Geodiversitas , 19 ( 4 ) : 783 – 844 .
- Merriam , J. C. 1901 . A contribution to the geology of the John Day Basin . University of California, Publication of Department of Geological Sciences , 2 ( 9 ) : 269 – 314 .
- Merriam , J. C. , Stock , C. and Moody , C. L. 1916 . An American Pliocene bear (Rattlesnake beds, John Day region, Oregon) . University of California Publications, Bulletin of the Department of Geology , 10 : 87 – 109 .
- Merriam , J. C. , Stock , C. and Moody , C. L. 1925 . The Pliocene Rattlesnake Formation and fauna of eastern Oregon, with notes on the geology of the Rattlesnake and Mascall deposits . Carnegie Institution of Washington, Contributions to Palaeontology , 347 : 43 – 92 .
- Nagel , D. 2009 . The early Vallesian vertebrates of Atzelsdorf (Late Miocene, Austria). 10. Carnivora . Annalen des Naturhistorischen Museums in Wien , 111A : 605 – 618 .
- Nagel , D. , Stefen , C. and Morlo , M. 2009 . The carnivoran community from the Miocene of Sandelzhausen (Germany) . Paläontologische Zeitschrift , 83 : 151 – 174 .
- Nyakatura , K. and Bininda-Emonds , O. R. P. 2012 . Updating the evolutionary history of Carnivora (Mammalia): a new species-level supertree complete with divergence time estimates . BMC Biology , 10 : 12
- Orlov , J. A. 1941 . “ Tertiary Carnivora of West Siberia, III: Mustelidae; pp ” . In Tertiary Mammalia and Localities of Their Remains , Edited by: Orlov , J. A. 30 – 39 . Travaux de l’Institut Paleontologique, Academie des Sciences de l’Union des Republicques Sovietiques Socialistes . 8
- Powell , R. 1981 . Martes pennanti . Mammalian Species , 156 : 1 – 6 .
- Prothero , D. R. , Hoffman , J. M. and Foss , S. E. 2006 . Magnetic stratigraphy of the upper Miocene (Hemphillian) Rattlesnake Formation, central Oregon . PaleoBios , 26 : 37 – 42 .
- Qiu , Z.-X. 2003 . “ Dispersals of Neogene carnivorans between Asia and North America; pp ” . In Vertebrate Fossils and Their Context: Contributions in Honor of Richard H. Tedford , Edited by: Flynn , L. J. 18 – 31 . Bulletin of the American Museum of Natural History 279 .
- Retallack , G. J. 1999 . Paleosols and Paleoenvironments of the Rattlesnake Formation (late Miocene) near Dayville, Oregon , 61 Final report, National Park Service, John Day Fossil Beds National Monument . no. 1443-PX9325-99-005
- Retallack , G. J. , Tanaka , S. and Tate , T. 2002 . Late Miocene advent of tall grassland paleosols in Oregon . Palaeogeography, Palaeoclimatology, Palaeoecology , 183 : 329 – 354 .
- Samuels , J. X. and Zancanella , J. 2011 . An early Hemphillian occurrence of Castor (Castoridae) from the Rattlesnake Formation of Oregon . Journal of Paleontology , 85 : 930 – 935 .
- Sato , J. J. , Hosoda , T. , Wolsan , M. , Tsuchiya , K. , Yamamoto , M. and Suzuki , H. 2003 . Phylogenetic relationships and divergence times among mustelids (Mammalia:Carnivora) based on nucleotide sequences of the nuclear interphotoreceptor retinoid binding protein and mitochondrial cytochrome b genes . Zoological Science , 20 : 243 – 264 .
- Sato , J. J. , Wolsan , M. , Minami , S. , Hosoda , T. , Sinaga , M. H. , Hiyami , K. , Yamaguchi , Y. and Suzuki , H. 2009 . Deciphering and dating the red panda's ancestry and early adaptive radiation of Musteloidea . Molecular Phylogenetics and Evolution , 53 : 907 – 922 .
- Sato , J. J. , Wolsan , M. , Prevosti , F. J. , D’Elía , G. , Begg , C. , Begg , K. , Hosoda , T. , Campbell , K. L. and Suzuki , H. 2012 . Evolutionary and biogeographic history of weasel-like carnivorans (Musteloidea) . Molecular Phylogenetics and Evolution , 63 : 745 – 757 .
- Schlosser , M. 1924 . Tertiary vertebrates from Mongolia . Palaeontologia Sinica, Series C , 1 : 1 – 132 .
- Sotnikova , M. V. 1995 . Extinct wolverine from the former USSR: review of the genus Plesiogulo (Carnivora: Mustelidae) . Lutreola , 6 : 1 – 8 .
- Streck , M. J. and Grunder , A. L. 1995 . Crystallization and welding variations in a widespread ignimbrite sheet; the Rattlesnake Tuff, eastern Oregon, U.S.A. . Bulletin of Volcanology , 57 : 151 – 169 .
- Tedford , R. H. , Skinner , M. S. , Fields , R. S. , Rensberger , J. M. , Whistler , D. P. , Galusha , T. , Taylor , B. E. , Macdonald , J. R. and Webb , S. D. 1987 . “ Faunal succession and biochronology of the Arikareean through Hemphillian (late Oligocene through earliest Pliocene Epochs) in North America; pp ” . In Cenozoic Mammals of North America , Edited by: Woodburne , M. O. 153 – 210 . Berkeley , , California : University of California Press .
- Tedford , R. H. , Albright , L. B. III , Barnosky , A. D. , Ferrusquia Villafranca , I. , Hunt , R. M. Jr. , Storer , J. E. , Swisher , C. C. III , Voorhies , M. R. , Webb , S. D. and Whistler , D. P. 2004 . “ Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene Epochs) ” . In Late Cretaceous and Cenozoic Mammals of North America , Edited by: Woodburne , M. O. 169 – 231 . New York : Columbia University Press .
- Teilhard de Chardin , P. and Leroy , P. 1945 . Les Mustélidés de Chine . Publications de l’Institut de Géobiologie , 12 : 1 – 56 .
- Thorpe , M. R. 1921 . Two new fossil Carnivora . American Journal of Science , 1 : 477 – 483 .
- Viret , J. 1951 . Catalogue critique de la faune des mammifères miocènes de la Grive Saint-Alban . Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon , 3 : 1 – 104 .
- Wagner , A. 1858 . Geschichte der Urwelt, mit besonderer Berücksichtigung der Menschenrassen und des mosaischen Schöpfungsberichtes (Second Edition) , 528 Leipzig : Leopold Voss .
- Walker , G. W. 1979 . Revisions to the Cenozoic stratigraphy of Harney Basin, southeastern Oregon . Bulletin U.S. Geological Survey , 1475 : 1 – 35 .
- Walker , G. W. 1990 . “ Miocene and younger rocks of the Blue Mountains region, exclusive of the Columbia River Basalt Group and associated mafic lava flows ” . In Geology of the Blue Mountain Region of Oregon, Idaho and Washington: Cenozoic Geology of the Blue Mountains Region , Edited by: Walker , G. W. 101 – 118 . U.S. Geological Survey . Professional Paper 1437
- Wang , X. , Tseng , Z. J. and Takeuchi , G. T. 2012 . Zoogeography, Molecular Divergence, and the Fossil Record—the Case of an Extinct Fisher, Pekania palaeosinensis (Mustelidae, Mammalia), from the Late Miocene Baogeda Ula Formation, Inner Mongolia . Vertebrata PalAsiatica , 50 : 293 – 307 .
- Wolsan , M. 1989 . “ Drapieżne–Carnivora; pp ” . In Historia I ewolucja lą dowej fauny Polski , Edited by: Kowalski , K. 177 – 196 . Folia Quatern . 59, 60
- Wolsan , M. and Sato , J. J. 2010 . Effects of data incompleteness on the relative performance of parsimony and Bayesian approaches in a supermatrix phylogenetic reconstruction of Mustelidae and Procyonidae (Carnivora) . Cladistics , 26 : 168 – 194 .
- Wood , H. E. Jr. , Chaney , R. W. , Clark , J. , Colbert , E. H. , Jepsen , G. L. , Reeside , J. B. Jr. and Stock , C. 1941 . Nomenclature and correlation of the North American continental Tertiary . Bulletin Geological Society of America , 52 : 1 – 48 .
- Woodburne , M. O. 2004 . “ Global events and the North American mammalian biochronology; pp ” . In Late Cretaceous and Cenozoic Mammals of North America , Edited by: Woodburne , M. O. 315 – 343 . New York : Columbia University Press .
- Yu , L. , Peng , D. , Liu , J. , Luan , P. , Liang , L. , Lee , H. , Lee , M. , Ryder , O. A. and Zhang , Y. 2011 . On the phylogeny of Mustelidae subfamilies: analysis of seventeen nuclear non-coding loci and mitochondrial complete genomes . BMC Evolutionary Biology , 11 : 92
- Zdansky , O. 1924 . Jungtertiäre Carnivoren Chinas . Palaeontologia Sinica, Series C , 2 ( 1 ) : 1 – 149 .