
ABSTRACT
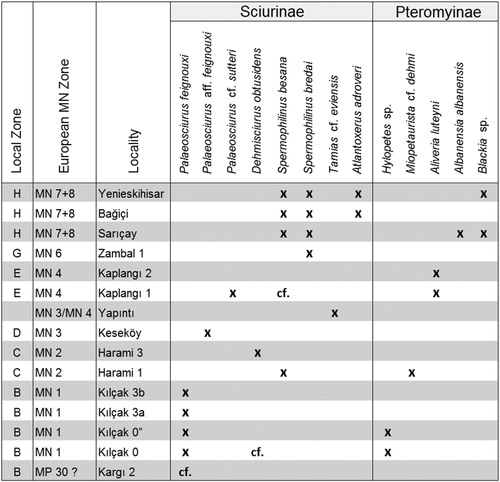
Isolated cheek teeth of Sciuridae (Mammalia, Rodentia) from 15 early and middle Miocene localities in Anatolia (Turkey) are described. The localities range in age from local zone B to local zone H, where zones are correlated to European MN zones 1 to 7 + 8. The material represents the two subfamilies: Sciurinae (ground and tree squirrels) and Pteromyinae (flying squirrels). The number of different species found at a single locality ranges from one to four. The Sciurinae were found to belong to the genera Palaeosciurus, Dehmisciurus (formerly called ?Ratufa), Spermophilinus, Tamias, and Atlantoxerus. Members of the Pteromyinae are Hylopetes, Miopetaurista, Aliveria, Albanensia, and Blackia. The MN 2 locality Harami 1 has yielded the oldest Spermophilinus and Miopetaurista known so far. Atlantoxerus adroveri from Bağiçi and Yenieskihisar (MN 7 + 8) is the first member of the Xerini tribe in Anatolia. The squirrel from Keseköy (MN 3), described as Palaeosciurus aff. feignouxi, shows close morphological resemblance to early Miocene Protospermophilus kelloggi from North America. This may suggest that European Palaeosciurus and American Protospermophilus are closely related genera. Three MN 7 + 8 localities (Sarıçay, Bağiçi, and Yenieskihisar) yielded two Spermophilinus species, which shows that Spermophilinus developed, at least locally, along two parallel lines. Similarity between Anatolian and European squirrel assemblages confirms that Anatolia was connected to the European part of the Eurasian continent in early and middle Miocene times. The lacustrine character of the sediments and the presence of both ground and/or tree and flying squirrels indicate that most of the localities represent a wet, but forested environment.
INTRODUCTION
Squirrels (Sciuridae) are usually scarce in the small mammal fossil faunas of Anatolia, Turkey, making their first appearance in the latest Oligocene (this study). Although remaining rare, the family is diverse throughout the Miocene. In a preceding paper (Bosma et al., Citation2013), we described isolated cheek teeth of Sciuridae from late Miocene localities in central Anatolia. The present paper deals with similar material from the early and middle Miocene.
The teeth to be described were collected within the framework of a collaborative project between the Maden Tetkik ve Arama Enstitüsü (MTA), Ankara (Turkey), and the Bundesanstalt für Bodenforschung, Hannover (Germany), during the 1960s (Sickenberg et al., Citation1975), and within an informal agreement between the MTA and the Department of Earth Sciences of Utrecht University, The Netherlands, during the last two decades of the 20th century (Ünay et al., Citation2003; De Bruijn et al., Citation2013b). Both projects involved wet screening of large amounts of fossiliferous matrix in the field (the Turkish-Dutch team washed over 25 tons from the localities that yielded sciurid remains). Large assemblages of early and middle Miocene rodents were recovered, in which the Muridae dominate (Ünay et al., Citation2003) but the Sciuridae are poorly represented.
Our material includes teeth from 15 localities: Kargı 2, Kılçak 0, 0”, 3a, and 3b, Harami 1 and 3, Keseköy, Yapıntı, Kaplangı 1 and 2, Zambal 1, Sarıçay, Bağiçi, and Yenieskihisar. All these localities are situated in lacustrine deposits. Their geographical positions are indicated on the map in . Global Positioning System (GPS) coordinates and short descriptions of the lithology are given in . presents the estimated biochronological ages of the localities. We follow Ünay et al. (Citation2003), who constructed an informal local biozonation of the Anatolian continental Neogene based on the stage of evolution of the Muridae and Dipodidae, and De Bruijn et al. (Citation2013b), who, on the same basis, suggested correlations to the European MN (= Mammal Neogene zonation) age system. The ages of the localities range from MP 30 (latest Oligocene) (MP = Mammal Paleogene zonation) to MN 7 + 8 (late middle Miocene).
FIGURE 1. Early and middle Miocene sciurid rodent tooth localities in Anatolia, Turkey: Kargı 2; Kılçak 0, 0”, 3a, and 3b; Harami 1 and 3; Keseköy; Yapıntı; Kaplangı 1 and 2; Zambal 1; Sarıçay; Bağiçi; and Yenieskihisar. All these localities are situated in lacustrine deposits.
TABLE 1. List of localities with geographical and lithological data.
TABLE 2. List of localities with proposed biochronological ages.
The squirrels studied are members of the two subfamilies of Sciuridae: Sciurinae (ground and tree squirrels) and Pteromyinae (flying squirrels). Genera belonging to the Sciurinae are Palaeosciurus, Dehmisciurus (formerly called ?Ratufa), Spermophilinus, Tamias, and Atlantoxerus. Genera belonging to the Pteromyinae are Hylopetes, Miopetaurista, Aliveria, Albanensia, and Blackia. As pointed out by Thorington et al. (Citation2005), unequivocal determination of fossil material as belonging to a flying squirrel requires data on the postcranial skeleton. Because our material consists of isolated cheek teeth only, we follow current views when considering a genus to be pteromyine.
Discussions will be presented on the affinities of Miocene squirrels from Anatolia to those from southeastern Europe and other regions, and their bearing on paleoenvironmental reconstruction.
Institutional Abbreviations—CM, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania, U.S.A.; MTA, Maden Tetkik ve Arama Enstitüsü, Ankara, Turkey; UU, Department of Earth Sciences of Utrecht University, Utrecht, The Netherlands.
Locality Abbreviations—Bag, Bağiçi; Har1, Harami 1; Har3, Harami 3; Kap1, Kaplangı 1; Kap2, Kaplangı 2; Karg2, Kargı 2; Kes, Keseköy; Ki0, Kılçak 0; Ki0”, Kılçak 0”; Ki3a, Kılçak 3a; Ki3b, Kılçak 3b; SC, Sarıçay; Yap, Yapıntı; YE, Yenieskihisar; Zam1, Zambal 1.
MATERIALS AND METHODS
The material described here was collected by wet screening of sediment, using a set of screens, the finest of which had a 0.5 mm mesh. The length and width of the teeth were measured using a Leitz Orthoplan measuring microscope equipped with mechanical stage and measuring clocks. All measurements (length × width) are given in mm, and an en dash (–) indicates that a measurement was not possible. The nomenclature of Qiu (Citation1996) is followed to describe the occlusal patterns of the teeth. All teeth are depicted as left teeth. The orientation of the upper teeth is with the lingual side downward and the anterior side to the left, and the orientation of the lower teeth is with the buccal side downward and the anterior side to the left. All new material is from the MTA collections. Protospermophilus kelloggi Black, Citation1963, teeth from the Split Rock Formation (Fremont County, Wyoming, U.S.A.) and reposited in the CM are figured for comparison (A–G, I–K); one P. kelloggi tooth is in the UU collection (H). Photographs were taken using a Jeol JCM-6000 scanning electron microscope.
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, Citation1758
Order RODENTIA Bowdich, Citation1821
Family SCIURIDAE Fischer de Waldheim, Citation1817
Subfamily SCIURINAE Fischer de Waldheim, Citation1817
Genus PALAEOSCIURUS Pomel, Citation1853
Type Species—Sciurus (Palaeosciurus) feignouxi Pomel, Citation1853.
Species Recognized—Palaeosciurus feignouxi Pomel, Citation1853 (MN 2, early Miocene); Sciurus fissurae Dehm, Citation1950 (MN 3, early Miocene); Palaeosciurus goti Vianey-Liaud, Citation1974 (MP 22, early Oligocene); Palaeosciurus sutteri Ziegler and Fahlbusch, Citation1986 (MN 4b, early Miocene); Palaeosciurus ultimus Mein and Ginsburg, Citation2002 (MN 8 or 7, middle Miocene).
Remarks—The holotype of Palaeosciurus feignouxi from Langy, France, is lost. In accordance with Werner (Citation1994), we use the assemblage of P. feignouxi from Montaigu-le-Blin (France) as reference assemblage (cf. Freudenberg, Citation1941; Stehlin and Schaub, Citation1951; Mein and Ginsburg, Citation2002).
We identify teeth from the following localities as Palaeosciurus teeth: Kargı 2, Kılçak 0, Kılçak 0”, Kılçak 3a, Kılçak 3b, Keseköy, and Kaplangı 1. The material from the Kılçak localities can be assigned in straightforward fashion to P. feignouxi, because of its close similarity to the material from Montaigu-le-Blin present in the collections of UU. We will describe the material from Kargı 2, a locality probably older than the Kılçak localities, as P. cf. feignouxi, and not as P. feignouxi, because only four teeth are available and none of these is complete. The material from Kaplangı 1 also enables straightforward determination. It will be described as P. cf. sutteri (not as P. sutteri), because it consists only of two lower teeth, an m1 and an m3. Morphological and size differences between P. feignouxi, P. fissurae, and P. sutteri are small. They are discussed in detail by Ziegler and Fahlbusch (Citation1986) and by Werner (Citation1994). It is unclear whether these three species belong to the same lineage.
The identification of the assemblage from Keseköy is much more complex. The teeth correspond in size with those of Palaeosciurus feignouxi. The majority of the material (a D4, three M1/2s, two p4s, an m1, and an m3) also agrees morphologically with P. feignouxi. Two P4s and two M3s, however, show details of the occlusal structure not frequently observed in P. feignouxi or in other Palaeosciurus species. We will describe the material from Keseköy as P. aff. feignouxi and discuss its close resemblance to Protospermophilus Gazin, Citation1930, a genus known from the Oligocene and Miocene of North America.
PALAEOSCIURUS FEIGNOUXI Pomel, Citation1853
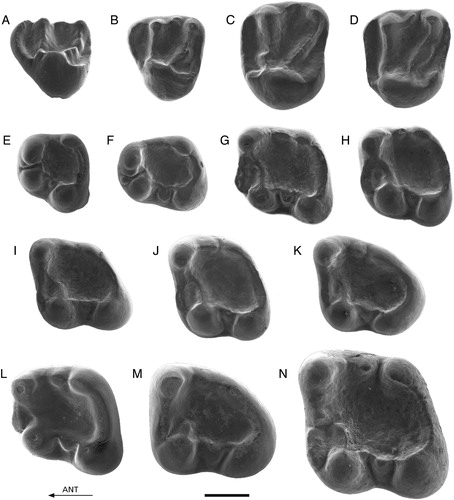
FIGURE 2. A–K, Palaeosciurus feignouxi from Kılçak 0, 0”, and 3a. A, Ki0"-1, D4 (reversed); B, Ki0"-2, P4; C, Ki3a-5, M1/2; D, Ki0-1, M1/2 (reversed); E, Ki3a-11, p4; F, Ki3a-13, p4 (reversed); G, Ki3a-19, m1; H, Ki3a-16, m1; I, Ki0"-7, m2 (reversed); J, Ki3a-22, m2; K, Ki3a-25, m3 (reversed). L–M, Palaeosciurus cf. sutteri from Kaplangı 1. L, Kap1-101, m1; M, Kap1-103, m3. N, Dehmisciurus obtusidens from Harami 3. Har3-1, m2 (reversed). Scale bar equals 1 mm.
Localities—Kılçak 0 (Ki0), Kılçak 0” (Ki0”), Kılçak 3a (Ki3a), and Kılçak 3b (Ki3b).
Level—Local rodent zone B (tentatively correlated to MN 1) (De Bruijn et al., Citation2013b).
Material—Thirty-eight teeth from four localities. Kılçak 0: Ki0-1, an M1/2; Ki0-6, an m2; Ki0-7, an m3 (Ki0-6 and Ki0-7 probably from same individual). Kılçak 0": Ki0"-1, a D4; Ki0"-2, -3, two P4s; Ki0"-4, an M1/2; Ki0"-6, a d4; Ki0"-7, an m2; Ki0"-8, -9, fragments of two m1s or m2s; Ki0"-10, an m3. Kılçak 3a: Ki3a-2, a D4; Ki3a-1, -3, two P4s; Ki3a-4, -5, -6, -7, -8, five M1/2s; Ki3a-9, -10, two M3s; Ki3a-11, -12, -13, -14, four p4s; Ki3a-15, -16, -17, -18, -19, five m1s; Ki3a-21, -22, -23, three m2s; Ki3a-25, -26, two m3s. Kılçak 3b: Ki3b-1, a D4; Ki3b-3, an M1/2.
Measurements—See .
TABLE 3. Measurements (in mm) of teeth of Palaeosciurus feignouxi from Kılçak 0, 0", 3a, and 3b, Palaeosciurus cf. feignouxi from Kargı 2, and Palaeosciurus aff. feignouxi from Keseköy.
Description
Because P. feignouxi is a well-known and well-described species, only particular features of the material are given.
D4—The D4s from Kılçak 3a and Kılçak 0” are well preserved. Specimen Ki3a-2 has a prominent anterior portion with a distinct parastyle. The metaloph carries a small but distinct metaconule. There is no identifiable hypocone. Two very small cuspules are present between the paracone and the metacone. Specimen Ki0"-1 has a less distinct metaconule. The tooth possesses a mesostyle, which has the shape of a low crest. The mesostyle is closely connected to the metacone.
P4—One P4 from Kılçak 3a and one P4 from Kılçak 0” are well preserved. The anterior portion is prominent. There are a well-developed parastyle, an indistinct hypocone, an indistinct metaconule, and a distinct, isolated mesostyle.
M1/2—All M1/2s have a small hypocone. Some possess a weak metaconule. The mesostyle is mostly present as a cuspule or low and narrow crest that is closely connected to the paracone. The mesostyle is an isolated cuspule in Ki0"-4.
M3—The two M3s (Kılçak 3a), both worn, show a high and pointed paracone.
d4—Specimen Ki0"-6 has a short and low anterolophid, which comes from the top of the metaconid and ends low against the anterior wall of the protoconid. There is a distinct entoconid.
p4—Two of the three complete p4s (Kılçak 3a), both slightly worn, show an entoconid. In the third, unworn tooth, the entoconid is indistinct, being part of the posterolophid. A small anteroconid is situated against the anterior tooth wall, between the protoconid and the metaconid. The teeth possess a mesoconid of variable size. There is either no mesostylid or a mesostylid in the form of a cuspule or low crest.
m1 and m2—In most specimens, the entoconid can be recognized as a separate cusp. All teeth carry a mesoconid. Only some teeth show a distinct mesostylid.
m3—The entoconid is more or less identifiable as a separate cusp. All teeth have a mesoconid. One of the two m3s from Kılçak 3a (both little worn) has a metalophid and a small mesostylid.
PALAEOSCIURUS cf. FEIGNOUXI Pomel, Citation1853
Locality—Kargı 2 (Karg2).
Level—Local rodent zone A. This zone is correlated with uncertainty to MP 30 (late Oligocene) (De Bruijn et al., Citation2013b).
Material—Karg2-1, a P4; Karg2-3, -4, two M1/2s; Karg2-6, an M3.
Measurements—See .
Description
The material from Kargı 2 very closely resembles the Kılçak material. A difference to be observed is the smaller anterior portion (parastyle included) of Karg2-1, a P4, in comparison with the anterior portions of the P4s from Kılçak 0” and Kılçak 3a.
Discussion
Palaeosciurus feignouxi has been collected from several localities in western (Spain and France) and central (Germany and Switzerland) Europe (Fortelius, Citation2017). Most of these localities have an MN 2 or MN 1 age, which agrees with the age postulated for Kılçak 0, 0”, 3a, and 3b (MN 1; De Bruijn et al., Citation2013b). In addition to the MN 1 and MN 2 localities, P. feignouxi has been reported from the late Oligocene (MP 30) locality Bergasa in Spain (Cuenca et al., Citation1992) and from the MN 3 locality Frankfurt-Nordbassin in Germany (Stephan-Hartl, Citation1972). The age of Kargı 2 is possibly also MP 30 (De Bruijn et al., Citation2013b). Joniak et al. (Citation2017) recently described P. feignouxi from the Anatolian locality Gökler 4A (local zone C, MN 2).
PALAEOSCIURUS aff. FEIGNOUXI Pomel, Citation1853
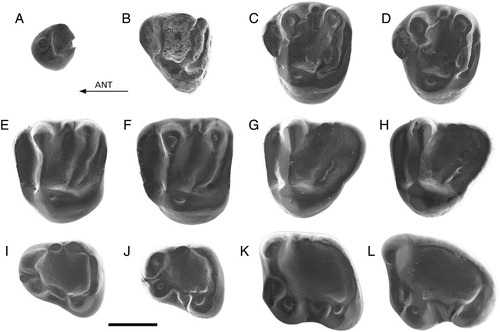
FIGURE 3. Palaeosciurus aff. feignouxi from Keseköy. A, Kes-101, P3 (reversed); B, Kes-102, D4 (reversed); C, Kes-103, P4; D, Kes-104, P4 (reversed); E, Kes-107, M1/2 (reversed); F, Kes-105, M1/2; G, Kes-109, M3 (reversed); H, Kes-108, M3; I, Kes-112, p4 (reversed); J, Kes-111, p4; K, Kes-114, m1; L, Kes-116, m3. Scale bar equals 1 mm.
Locality—Keseköy (Kes).
Level—Local rodent zone D (tentatively correlated to MN 3) (De Bruijn et al., Citation2013b).
Material—Kes-101, a P3; Kes-102, a D4; Kes-103, -104, two P4s; Kes-105, -106, -107, three M1/2s; Kes-108, -109, two M3s; Kes-111, -112, two p4s; Kes-114, a m1; Kes-116, a m3.
Measurements—See .
Description
P3—Specimen Kes-101 most probably belongs to P. aff. feignouxi. The tooth is complete, with the tip of its long, straight root still positioned in a small piece of bone. The diameter of the root is hardly smaller than the diameter of the crown. The worn occlusal surface is flat.
D4—Specimen Kes-102 is slightly damaged on the buccal side. The tooth has a large parastyle, which gives it a triangular outline. It has a protocone, a paracone, a metacone, a protoloph, a metaloph, and a posteroloph. The posteroloph is much lower than the metaloph and is connected to the protocone. The lingual half of the protoloph extends along the anterolingual tooth border.
P4—The two P4s (Kes-103 and Kes-104) may come from the same individual. They possess a well-developed parastyle, which is situated relatively lingually. The protocone, the paracone, and the metacone are large and prominent. The protoloph and the metaloph are high and broad. The lingual half of the protoloph is situated close to the anterolingual border of the tooth. The metaloph carries a large metaconule. The posteroloph is much lower and narrower than the protoloph and the metaloph. The posteroloph is connected to the protocone. The two teeth show a small but distinct mesostyle, which is isolated from the paracone and the metacone. The surface of the central valley is slightly irregular.
M1/2—The M1/2s have a regular occlusal pattern. The protocone, the paracone, the metacone, the protoloph, and the metaloph are high and prominent. The anteroloph and the posteroloph are well developed, but lower. A small hypocone is present at the lingual end of the posteroloph. The hypocone is lower than the protocone. It is connected to the protocone by a high ridge. The protoloph and the metaloph form a ‘V,’ which is directed anteriorly. The metaloph is narrow adjacent to the protocone. There are no protoconule or metaconule. The three teeth each show a small but distinct mesostyle, which is isolated from the paracone and the metacone.
M3—The two M3s (Kes-108 and Kes-109) may come from the same individual. Their outline is triangular. The most prominent elements of the occlusal pattern are the protocone, the paracone, the protoloph, and the anteroloph. The anteroloph is lower than the protoloph. The posteroloph, including a vague mesostyle, borders the tooth posterobuccally, posteriorly, and posterolingually. The posteroloph is connected to the protocone. Both teeth carry a large portion of metaloph. ‘Missing’ parts of the metaloph are those adjacent to the protocone and to the posterior tooth corner.
p4—The two p4s represent slightly different wear stages. Both show the protoconid, the metaconid, and the hypoconid as prominent, rounded cusps. The entoconid is small and elongated and is closely connected to the posterolophid. The protoconid and the metaconid are connected by a narrow metalophid in the less worn tooth. Both teeth have an anteroconid at the anterior tooth wall between the protoconid and the metaconid. In the more worn tooth, the anteroconid is extremely small. In the less worn tooth, the anteroconid is connected to the protoconid. Both teeth bear a mesoconid. In the less worn specimen, the mesoconid is a crest between the protoconid and the hypoconid rather than a cusp. In the more worn p4, the mesoconid forms part of a low and narrow ectolophid. Both teeth possess a distinct, elongated mesostylid, which is isolated from the metaconid and the entoconid. The surface of the central valley is slightly uneven.
m1—The occlusal pattern of the single m1, Kes-114, is very regular. The protoconid, metaconid, and hypoconid are large and prominent. The metaconid is pointed and by far the highest cusp. The entoconid is small and elongated and is incorporated in the posterolophid. The protoconid and the metaconid are connected by the metalophid. The tooth possesses a well-developed anterolophid. The low and narrow ectolophid bears a distinct mesoconid. There is a small mesostylid, which is isolated from the metaconid and from the entoconid.
m3—The occlusal pattern of the single m3, Kes-116, is very regular. The protoconid, metaconid, anterolophid, hypoconid, ectolophid, and mesoconid are as in the m1. The metalophid curves anteriorly in the middle of the tooth. The tooth is bordered at the lingual and posterior sides by a high continuous wall, which is formed by the indistinct mesostylid, the indistinct entoconid, the posterolophid, and the small hypoconid. The surface of the central valley is slightly uneven.
Discussion
As mentioned in the introductory remarks on Palaeosciurus, the single D4, three M1/2s, two p4s, single m1, and single m3 from Keseköy are in size as well as morphology close to P. feignouxi teeth. The remaining teeth in the assemblage are a left and a right P4 (C, D) and a left and a right M3 (G, H), which may, two by two, or all four, have belonged to the same animal. Conspicuous features of the two P4s are the lingually positioned parastyle, the large metaconule, and the irregular surface of the central valley. The two M3s are exceptional in their posterior outline (the anterior and buccal tooth sides are nearly at right angles) and in the presence of a distinct, although incomplete, metaloph.
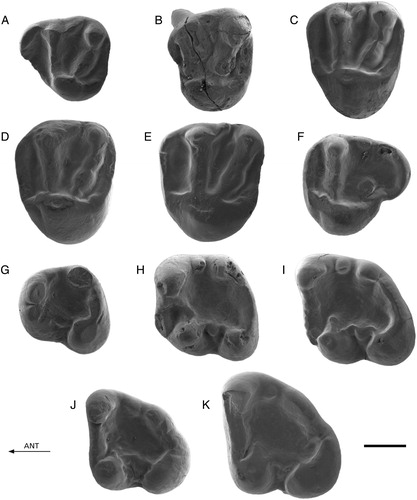
The material from Keseköy resembles in a striking way a set of isolated cheek teeth of Protospermophilus kelloggi Black, Citation1963, from the type locality (Split Rock Formation), which we received on loan from the CM. We illustrate this resemblance by figuring in the present paper a number of these teeth (A–K). The main difference between the species from Keseköy and Pr. kelloggi is the smaller size of the former. The material from Split Rock is of early Miocene (Hemingfordian 2) age. Other species belonging to Protospermophilus Gazin, Citation1930, a genus restricted to North America, are Pr. vortmani (Cope, Citation1879) (Arikareean 1–2, early Oligocene–early Miocene); Pr. angusticeps (Matthew and Mook, Citation1933) (Arikareean 3, early Miocene); Pr. malheurensis (Gazin, Citation1932) (Barstovian 1, middle Miocene); Pr. oregonensis (Downs, Citation1956) (Barstovian 1, middle Miocene); and Pr. quatalensis Gazin, Citation1930 (Clarendonian 2, middle–late Miocene) (age assignments as in Fortelius, Citation2017). Excellent illustrations of material belonging to these species were published by Black (Citation1963). Interestingly, the M3 in the type specimen of Pr. malheurensis, an incomplete skull, is shown to carry a prominent metaconule or incomplete metaloph (Black, Citation1963:pl. 13, fig. 3). It can be questioned whether or not the squirrel from Keseköy should be classified as a new Protospermophilus species. We have chosen allocation to the European genus Palaeosciurus, because of the general similarity to P. feignouxi, and to avoid premature speculation on faunal exchange between North America and Eurasia.
PALAEOSCIURUS cf. SUTTERI Ziegler and Fahlbusch, Citation1986
FIGURE 4. Protospermophilus kelloggi from Split Rock, U.S.A. Comparative material. A, CM 14883, D4; B, CM 15924, P4 (reversed); C, CM 15924, M1/2 (reversed); D, CM 15924, M1/2 (reversed); E, CM 14295, M1/2; F, CM 13521, M3; G, CM14661, p4 (reversed); H, UU 475, m1; I, CM 15855, m2 (reversed); J, CM 15855, m3 (reversed); K, CM 13524, m3. Scale bar equals 1 mm.
Locality—Kaplangı 1 (Kap1).
Level—Local rodent zone E (tentatively correlated to MN 4) (De Bruijn et al., Citation2013b).
Material—Kap1-101, an m1; Kap1-103, an m3.
Measurements—Kap1-101: 2.45 × 2.44 mm; Kap1-103: 2.85 × 2.54 mm.
Description
m1—The tooth is narrower anteriorly than posteriorly. It is slightly worn. The protoconid, metaconid, and hypoconid are prominent, robust cusps. The entoconid is smaller and forms the lingual end of the posterolophid. The metalophid has a deep depression in the middle. The anterolophid ends buccally in a small anteroconid. The anteroconid is lower than the protoconid and is separated from the latter cusp by an incision. The mesoconid, although lower than the protoconid and the hypoconid, is large and forms a prominent structure in the buccal valley. It is connected to the protoconid and the hypoconid by low crests. The tooth bears a distinct mesostylid. The mesostylid is separated from the metaconid by a shallow depression.
m3—Specimen Kap1-103 is little worn. Its posterior portion is relatively broad. The protoconid, metaconid, and hypoconid are robust, rounded cusps. The posterolophid is a continuous crest, which ends in a small mesostylid, without showing a separate entoconid. The metalophid ends halfway along the central valley. The anterolophid connects the top of the metaconid to the anterolingual wall of the protoconid. There is no anteroconid. The mesoconid has the same structure as in the m1. The surface of the central valley carries indistinct crenulations.
Discussion
Palaeosciurus sutteri has so far been known from localities in central (Germany, Switzerland, Czech Republic, Hungary, and Austria) and southeastern (Greece) Europe (Fortelius, Citation2017). Its type locality is the MN 4b locality Forsthart in Germany. Localities in Greece are Aliveri (De Bruijn et al., Citation1980; Ziegler and Fahlbusch, Citation1986) and Antonios (Vasileiadou and Koufos, Citation2005). The species from Aliveri was originally described as P. aff. fissurae.
The two teeth from Kaplangı 1 are relatively large. Their measurements surpass those of m1 and m3 in the type assemblage (Ziegler and Fahlbusch, Citation1986) and in the assemblage from Aliveri (MN 4). In size, they agree more with the m1s and m3s in the assemblages from Langenmoosen and Puttenhausen (Germany; MN 5; Ziegler and Fahlbusch, Citation1986) and Obergänserndorf (Austria; MN 5; Daxner-Höck, Citation1998). Straightforward comparison with the material from Antonios (MN 4–MN 5 boundary) is not possible, because only one tooth, an m2, is available from this locality (Vasileiadou and Koufos, Citation2005). The relatively large size of our two teeth might indicate that Kaplangı 1 is somewhat younger than has been thought so far (De Bruijn et al., Citation2013b), and that its age is MN 5 rather than MN 4.
Genus DEHMISCIURUS Marković, De Bruijn, and Wessels, 2016
Type Species—?Ratufa obtusidens Dehm, Citation1950.
Species Recognized—?Ratufa obtusidens Dehm, Citation1950 (MN 3, early Miocene).
Remarks—The tentative assignment by Dehm (Citation1950) of the obtusidens material from Wintershof-West (Germany) to Ratufa, a genus of extant giant tree squirrels in southeastern Asia, has in recent years become more and more unsatisfactory. Mein and Ginsburg (Citation1997) already suggested that ?Ratufa obtusidens should be classified into a new genus. As a temporary solution, Aldana Carrasco (Citation1992) and Costeur et al. (Citation2012) proposed ascribing ?R. obtusidens to the genus Palaeosciurus Pomel, 1853. Recently, Marković et al. (2016:81) created the genus Dehmisciurus for ?R. obtusidens, with the following diagnosis (for genus and species): “Large sciurid. Mandibular ramus deep with short diastema and masseter scar ending below the p4. Cheek teeth robust with strong blunt cones, shallow basins with slightly irregular enamel surface in unworn specimens. Shape of upper molars rounded, protoloph and metaloph low and sometimes incomplete. m1 and m2 with complete metalophid. All lower molars with a mesoconid and isolated mesostylid.” Marković et al. (Citation2016) consider Dehmisciurus to be an arboreal squirrel. We use the name Dehmisciurus in this paper.
DEHMISCIURUS cf. OBTUSIDENS (Dehm, Citation1950)
Locality—Kılçak 0 (Ki0).
Level—Local rodent zone B (tentatively correlated to MN 1) (De Bruijn et al., Citation2013b).
Material—Ki0-21, an incomplete M1/2.
Description
M1/2—Specimen Ki0-21 is low-crowned and very worn, and 3.37 mm wide (length indeterminate). Its posterobuccal part is missing. The broad protocone, the much smaller hypocone, and parts of the protoloph and metaloph are recognizable from remnants of their dentine. The metacone is slightly higher than the protocone. The anteroloph and the protoloph are complete. The lingual part of the protoloph is broader than the buccal part. The metaloph appears to be absent adjacent to the protocone. The tooth has large wear facets anteriorly and posteriorly.
DEHMISCIURUS OBTUSIDENS (Dehm, Citation1950)
Locality—Harami 3 (Har3).
Level—Local rodent zone C (tentatively correlated to MN 2) (Ünay et al., Citation2003; De Bruijn et al., Citation2013b). Krijgsman et al. (Citation1996) correlated Harami 3 to chron C6Bn.1r, which indicates that the absolute age of the locality is between 22.0 and 22.5 Ma (Hilgen et al., Citation2012).
Material—Har3-1, an m2.
Measurement—3.08 × 3.20 mm.
Description
m2—Specimen Har3-1 is only very slightly worn. It has a low crown. The main cusps (protoconid, metaconid, hypoconid, entoconid) are large and prominent. The metaconid is by far the highest cusp. There is a small mesoconid, which extends into the buccal valley, and is connected to the protoconid and the hypoconid by very low and narrow ridges. The tooth bears a distinct mesostylid, which is separated from the metaconid and the entoconid by depressions. The depression between the mesostylid and the entoconid is very deep, which gives the entoconid an isolated aspect. The metalophid is somewhat irregular and shows a depression in the middle. The posterolophid is strongly developed and is only little lower than the hypoconid and the entoconid. The anterolophid is short and occupies the buccal half of the anterior tooth side only. The anterolophid and protoconid are separated by an incision. The central valley carries a fine network of crenulations.
Discussion
The very close similarity between the m2 from Harami 3 and the first and second lower molars in the type material of Dehmisciurus from Wintershof-West (Germany) (Dehm, Citation1950) excludes any uncertainty with regard to the identity of the tooth. We therefore refer to this tooth as D. obtusidens (and not D. cf. obtusidens).
So far, the total record of Dehmisciurus obtusidens is small. The type assemblage as described by Dehm (Citation1950) includes two incomplete lower jaws, one with incisor and m1–m3 and one with m2, and a number of isolated teeth. Other materials are two incomplete lower jaws with p4–m3 and four isolated teeth from the MN 4 locality El Canyet, Spain (Aldana Carrasco, Citation1992), one M1/2 from the MN 2a locality Ulm-Westtangente, Germany (Werner, Citation1994; Costeur et al., Citation2012), one M3 and possibly a P3 from the late Oligocene locality Banovići, Bosnia and Herzegovina (De Bruijn et al., Citation2013a), and nine isolated teeth from the MN 4 locality Sibnica, Serbia (Marković et al., Citation2016). The type locality Wintershof-West is the reference locality for MN 3.
The ages of Kılçak 0 and Harami 3 are considered to be MN 1 and MN 2, respectively. The occurrences of Dehmisciurus (cf.) obtusidens in Anatolia thus fall within the time range of D. obtusidens in the various European regions.
Genus SPERMOPHILINUS De Bruijn and Mein, Citation1968
Type Species—Sciurus bredai Von Meyer, Citation1848.
Species Recognized—Spermophilinus besana Cuenca Bescós, Citation1988 (MN 4, early Miocene); Sciurus bredai Von Meyer, Citation1848 (MN 6/MN 7 + 8, middle Miocene); Spermophilinus giganteus De Bruijn, Dawson, and Mein, Citation1970 (MN 14, early Pliocene); Spermophilinus turolensis De Bruijn and Mein, Citation1968 (MN 12, late Miocene).
SPERMOPHILINUS BESANA Cuenca Bescós, Citation1988
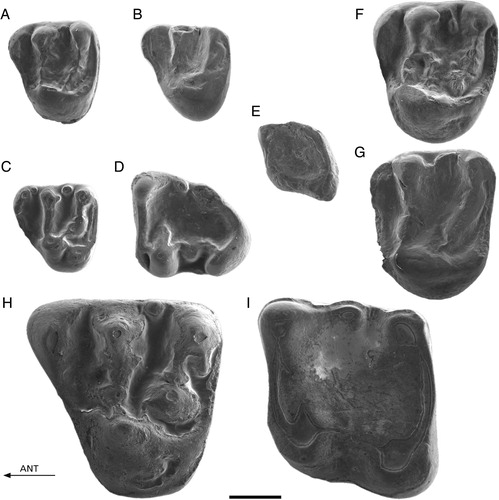
FIGURE 5. A–H, Spermophilinus besana from Sarıçay and Yenieskihisar. A, YE-110, D4; B, SC-103, P4; C, YE-126, M1/2; D, YE-123, M1/2; E, YE-135, M3; F, YE-141, d4; G, YE-151, m1; H, YE-168, m2. I–U, Spermophilinus bredai from Zambal 1, Sarıçay, Bağiçi, and Yenieskihisar. I, YE-105, D4; J, YE-103, D4; K, SC-101, P4; L, Zam1-101, M1/2; M, Zam1-102, M1/2; N, Bag-861, M1/2; O, YE-113, M1/2; P, YE-112, M1/2; Q, YE-131, M3; R, YE-146, p4; S, YE-154, m1; T, YE-164, m2; U, YE-173, m3. All images reversed except E, J, L, and Q. Scale bar equals 1 mm.
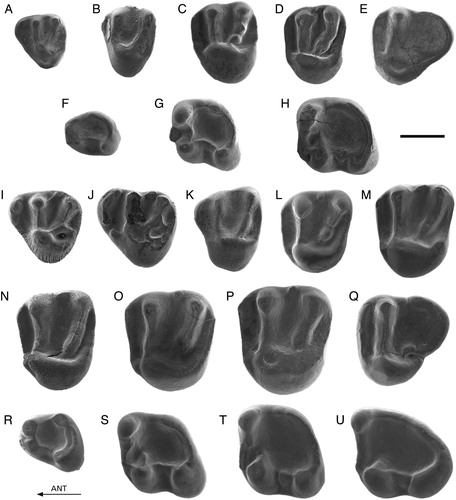
FIGURE 6. A–G, Tamias cf. eviensis from Yapıntı. A, Yap-101, D4; B, Yap-104, M1/2 (reversed); C, Yap-103, M1/2; D, Yap-106, M3; E, Yap-108, m2; F, Yap-110, m2 (reversed); G, Yap-109, m2. H–L, Atlantoxerus adroveri from Bağiçi and Yenieskihisar. H, YE-193, P4; I, Bag-888, M1/2 (reversed); J, Bag-895, m1; K, YE-201, p4; L, Bag-890, m1 (reversed). Scale bar equals 1 mm.
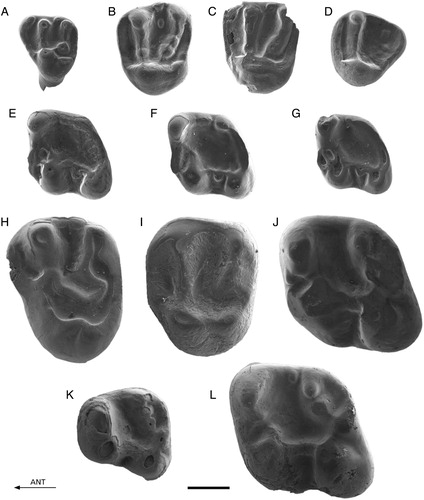
FIGURE 7. A–B, Hylopetes sp. from Kılçak 0 and 0”. A, Ki0"-11, P4; B, Ki0-11, P4. C–D, Aliveria luteyni from Kaplangı 1. C, Kap1-122, D4; D, Kap1-127, m3. E, Blackia sp. from Sarıçay. SC-131, m1. F–G, Miopetaurista cf. dehmi from Harami 1. F, Har1-1732, P4; G, Har1-1736, M1/2. H–I, Albanensia albanensis from Sarıçay. H, SC-141, P4; I, SC-147, m1. All images reversed except B. Scale bar equals 1 mm.
Localities—Harami 1 (Har1), Sarıçay (SC), Bağiçi (Bag), and Yenieskihisar (YE).
Levels—Harami 1: Local rodent zone C (tentatively correlated to MN 2) (De Bruijn et al., Citation2013b). Krijgsman et al. (Citation1996) correlated Harami 1 to chron C6Bn.2n, which indicates that the estimated absolute age of the locality is between 22.0 and 22.5 Ma (Hilgen et al., Citation2012). Sarıçay, Bağiçi, and Yenieskihisar: Local rodent zone H (tentatively correlated to MN 7 + 8) (Ünay et al., Citation2003; De Bruijn et al., Citation2013b).
Material—Fifty-eight teeth from four localities. Harami 1 (20 teeth): Har1-1741, a D4; Har1-1743, -1744, -1745, -1746, -1753, -1754, -1755, seven M1/2s; Har1-1756, -1757, two M3s; Har1-1771, a d4; Har1-1772, a p4; Har1-1763, -1773, two m1s; Har1-1761, -1764, -1774, -1776, four m2s; Har1-1778, -1779, two m3s. Sarıçay (four teeth): SC-103, a P4; SC-115, a p4; SC-118, -119, two m1s. Bağiçi (12 teeth): Bag-863, a D4; Bag-865, -868, two M1/2s; Bag-871, -878, two d4s; Bag-872, -883, two m1s; Bag-873, -874, -881, -882, four m2s; Bag-885, an m3. Yenieskihisar (22 teeth): YE-101, a P3; YE-108, -110, two D4s; YE-117, -118, -119, -120, -121, -122, -123, -124, -125, -126, 10 M1/2s; YE-133, -135, -136, -138, four M3s; YE-141, a d4; YE-151, -152, two m1s; YE-168, an m2; YE-176, an m3 (n.b.: small specimens of Spermophilinus bredai from Sarıçay, Bağiçi, or Yenieskihisar may have been misidentified as S. besana).
Measurements—See .
TABLE 4. Measurements (in mm) of teeth of Spermophilinus besana from Harami 1, Kaplangı 1 (S. cf. besana), Sarıçay, Bağiçi, and Yenieskihisar.
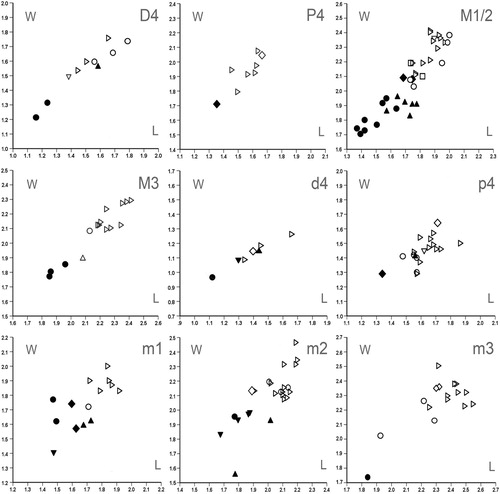
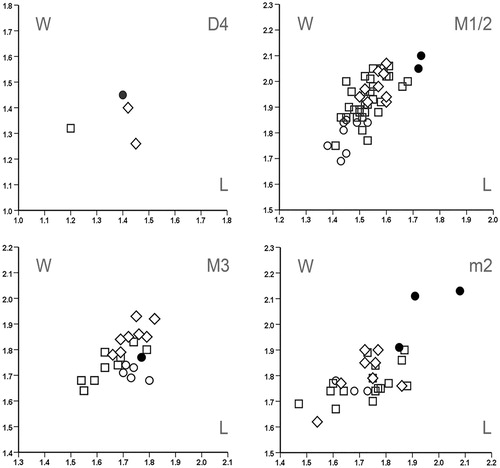
Length/Width Scatter Plots—See , .
FIGURE 8. Length/width scatter plots for upper (D4, P4, M1/2, and M3) and lower (d4, p4, m1, m2, and m3) tooth elements of Spermophilinus besana from Harami 1 (▴), Sarıçay (♦), Bağiçi (▾), and Yenieskihisar (●), and Spermophilinus bredai from Zambal 1 (□), Sarıçay (◊), Bağiçi (▿), and Yenieskihisar (○). Material from Paşalar (▹) is included for comparison.
Description
The morphology of Spermophilinus besana teeth is very similar to that of teeth of (the well-known) S. bredai. For this reason, only particular features of the material are presented.
P3—The crown of the single, unicuspidate P3, YE-101, presumably belonging to S. besana, is small and has the shape of a low, truncated cone.
D4—Specimen Har1-1741 is much larger than the other D4s. It has a small hypocone and a mesostyle. The mesostyle is a narrow ridge.
P4—The single P4, SC-103, is slightly rounded because of wear. Its anterior portion and posteroloph are very low.
M1/2—The M1/2s carry a small or indistinct hypocone. One tooth from Harami 1 has a small metaconule. Most M1/2s from Harami 1 possess a mesostyle in the form of a very small cuspule that is connected to the paracone rather than to the metacone. The majority of the M1/2s from Yenieskihisar are devoid of a mesostyle. One specimen from Harami 1 shows a short and steep ridge that comes down from the top of the protocone anterior to the protoloph.
M3—The M3s have the expanded posterior portion that is characteristic of Spermophilinus.
d4—The d4 from Harami 1 possesses an anteroconid in the form of a small and very flat cuspule in front of the protoconid. The d4s do not show a distinct mesoconid.
p4—The entoconid is recognizable as an indistinct cusp. There is no anteroconid. The (little worn) p4 from Harami 1 has a small mesoconid.
m1 and m2—In most teeth, the entoconid is recognizable as a small cusp. The m1s and m2s from Harami 1, in particular, show a prominent anteroconid at the end of the anterolophid, and a large mesoconid. These teeth often possess a small mesostylid. The surface of the central basin of only slightly worn specimens shows fine crenulations.
m3—The anterolophid is complete, but without an anteroconid. The metalophid is mostly incomplete. The entoconid and the mesostylid are hardly recognizable, being part of the posterolophid. The mesoconid is small. The occlusal surface of little worn specimens is finely crenulated.
SPERMOPHILINUS cf. BESANA Cuenca Bescós, Citation1988
Locality—Kaplangı 1 (Kap1).
Level—Local rodent zone E (tentatively correlated to MN 4) (De Bruijn et al., Citation2013b).
Material—Kap1-111, a D4.
Measurement—See .
Remarks—The tooth is described as Spermophilinus cf. besana (not as S. besana), because it is the only specimen available from this locality.
Description
D4—The tooth is subtriangular in shape. It has a regular occlusal pattern, with complete protoloph and metaloph, which form a ‘V.’ The cusps present are the protocone, the paracone, the metacone, the very low and indistinct hypocone, and the indistinct mesostyle.
SPERMOPHILINUS BREDAI (Von Meyer, Citation1848)
Localities—Zambal 1 (Zam1), Sarıçay (SC), Bağiçi (Bag), and Yenieskihisar (YE).
Levels—Zambal 1: Local rodent zone G (tentatively correlated to MN 6) (De Bruijn et al., Citation2013b). Sarıçay, Bağiçi, and Yenieskihisar: Local rodent zone H (tentatively correlated to MN 7 + 8) (Ünay et al., Citation2003; De Bruijn et al., Citation2013b).
Material—Thirty-four teeth from four localities. Zambal 1 (two teeth): Zam1-101, -102, two M1/2s. Sarıçay (eight teeth): SC-101, a P4; SC-105, -106, two M1/2s; SC-111, a d4; SC-113, a p4; SC-121, an m2; SC-123, -124, two m3s. Bağiçi (three teeth): Bag-861, an M1/2; Bag-876, -877, two p4s. Yenieskihisar (21 teeth): YE-103, -105, -106, three D4s; YE-111, -112, -113, -114, -115, five M1/2s; YE-131, -132, two M3s; YE-143, -144, -145, -146, four p4s; YE-154, an m1; YE-161, -162, -164, three m2s; YE-171, -173, -174, three m3s (n.b.: large specimens of Spermophilinus besana from Sarıçay, Bağiçi, or Yenieskihisar may have been misidentified as S. bredai).
Measurements—See .
TABLE 5. Measurements (in mm) of teeth of Spermophilinus bredai from Zambal 1, Sarıçay, Bağiçi, and Yenieskihisar.
Length/Width Scatter Plots— (n.b.: this figure also includes teeth of S. bredai from the Anatolian locality Paşalar, which we measured to enable direct comparison).
Description
Because the morphology of Spermophilinus bredai teeth is well known, only specimens with less common features are described.
Specimen Bag-861, the sole M1/2 from Bağiçi (N), possesses a deviating occlusal pattern. The protoloph and metaloph are very regular, relatively narrow crests, which run in parallel on the lingual half of the tooth. The protoloph ends in the very large protocone. The metaloph does not reach the protocone. The paracone and metacone are much smaller than the protocone and are pointed. The anteroloph is very low. The posteroloph forms a continuous crest with the protocone. There is a very small mesostyle, which is situated close to the metacone. In spite of its abnormal morphology, we consider this tooth to belong to S. bredai. The three D4s from Yenieskihisar show considerable variation in the size and shape of the parastyle. The parastyle is a strong crest in the largest tooth (J), whereas it is rather a pointed cusp in the two smaller teeth. The metaloph and the posteroloph are connected in one of the latter teeth (I). One M3 from Yenieskihisar possesses, adjacent to the protocone, a very low, short crest, which can be considered a remnant of the metaloph.
Discussion of Spermophilinus
The genus Spermophilinus includes four morphologically very similar species, which differ primarily in size (ordered from small to large): S. besana, S. bredai, S. turolensis, and S. giganteus. The two smaller species, in particular S. bredai, are relatively common; the two larger species are much more rare. In Europe, S. turolensis has been reported from late Miocene localities in Spain, France, Greece, and the Republic of Moldova (De Bruijn, Citation1995; Nicoara, Citation2011; Fortelius, Citation2017). Outside Europe, the species has been described from an MN 9 karst fissure fill in Anatolia (Bosma et al., Citation2013). Spermophilinus giganteus is known only from its type locality Maritsa, Greece.
Spermophilinus besana has been collected from localities in western (Spain and France), central (Germany, Czech Republic, Austria, and Hungary), and southeastern (Greece) Europe (Fortelius, Citation2017). The majority of these localities have an MN 4 or MN 5 age. The only locality with S. besana outside Europe has so far been the MN 9 locality Tuğlu in Anatolia (Joniak and De Bruijn, Citation2015). Spermophilinus besana from Tuğlu represents the youngest occurrence of the species. Until now, the oldest S. besana were from the MN 3 locality Ste. Catherine 2 in France (Aguilar, Citation2002) and the MN 3 localities Merkur-North and Tuchořice in the Czech Republic (Fejfar et al., Citation2003). The species from Harami 1 (MN 2) is now the oldest record of S. besana, and the oldest Spermophilinus known.
Spermophilinus bredai is a frequently occurring member of European Miocene faunas (Fortelius, Citation2017). Its type locality is Öhningen, Germany (MN 6/MN 7 + 8). Bosma et al. (Citation2013) described S. bredai from late Miocene localities in Anatolia. The ages of the localities with S. bredai range from MN 4 to MN 13.
It has generally been assumed among specialists that the four Spermophilinus species belong to one lineage, which is characterized by gradual size increase (De Bruijn, Citation1995). The presence of two species of Spermophilinus in the faunas from Sarıçay, Bağiçi, and Yenieskihisar contradicts this assumption. We need to observe here that suggestions have been made by earlier authors that Spermophilinus might develop along more than one line, and that two species of Spermophilinus might coexist (Aguilar et al., Citation1995b; Nicoara, Citation2011). Also, it was concluded by De Bruijn (Citation1995) that the size of S. bredai from the Anatolian locality Düzyayla does not fit a regular pattern of increase in size through time.
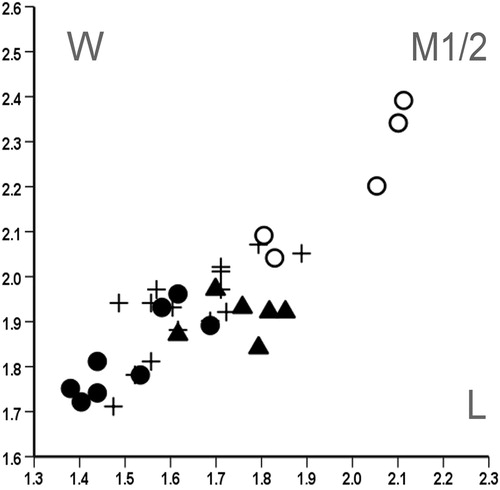
The length/width scatter plots in show that the Spermophilinus besana teeth from Harami 1 are distinctly larger than those from Sarıçay, Bağiçi, and Yenieskihisar. The original description of S. besana from the type locality Vargas 1A, Spain (MN 4), is based on a very small number of teeth. When establishing S. besana, Cuenca Bescós (Citation1988) indicated that Spermophilinus from Vieux Collonges (France; MN 4 and MN 5; Mein, Citation1958) belongs to this species. In , the M1/2s from Harami 1 and Yenieskihisar are compared with 15 M1/2s taken randomly from a collection of S. besana teeth from Vieux Collonges. This collection was kindly made available to us by P. Mein (Lyon, France). shows that the M1/2s from Harami 1 fall within the size range of the M1/2s from Vieux Collonges. Judging from the data given by Cuenca Bescós (Citation1988), the teeth from Harami 1 and Vieux Collonges are slightly larger than those from Vargas 1A.
FIGURE 9. Length/width scatter plot for M1/2 of Spermophilinus besana from Harami 1 (▴) and Yenieskihisar (●) and Spermophilinus bredai from Yenieskihisar (○). The M1/2s from Harami 1 fall within the size range of 15 randomly selected M1/2s of Spermophilinus besana from Vieux Collonges, France (+). See text for details.
The occurrences of the Spermophilinus species in the region of southeastern Europe and Anatolia are summarized in . compares the lengths of the M1/2s in the various assemblages. Length measurements, mean values, and specimen numbers are partly as presented in this paper and partly as given in the literature (see ). is supplemented with measurements of M1/2s of S. besana from Vargas 1A and Vieux Collonges and measurements of M1/2 of S. turolensis from the type locality Los Mansuetos, Spain. The type material of S. bredai consists of a lower jaw with p4–m2 only (Schlosser, Citation1884). In order to provide data for this species, measurements of M1/2s of S. bredai from La Grive L7 and M (France) are included (cf. Major, Citation1899; Black, Citation1966). These measurements were shared with us by P. Mein (Lyon, France).
FIGURE 10. Ranges and mean values of the length of M1/2 in assemblages of Spermophilinus (cf.) besana, S. (cf.) bredai, and S. (cf.) turolensis from southeastern Europe and Anatolia. Data from the literature (see for references) and this study. The numbers of teeth on which values are based are indicated next to the horizontal lines and dots. Kaplangı 1 and Kastellios K1 did not yield M1/2s. No measurements are available for the species from Silata. For Paşalar, two sets of data are given: those provided by Peláez-Campomanes and Daams (Citation2002) (Paşalar B) and those from this study (Paşalar A). For comparison, this figure includes measurements of M1/2 in the type collections of S. besana (Vargas 1A, Spain; MN 4) (Cuenca Bescós, Citation1988), and S. turolensis (Los Mansuetos, Spain; MN 12) (De Bruijn and Mein, Citation1968). In addition, values for M1/2 of S. besana from Vieux Collonges (France; MN 4 and MN 5) are included (cf. ). For S. bredai, measurements of M1/2s from La Grive L7 and M (France; MN 7) are used.
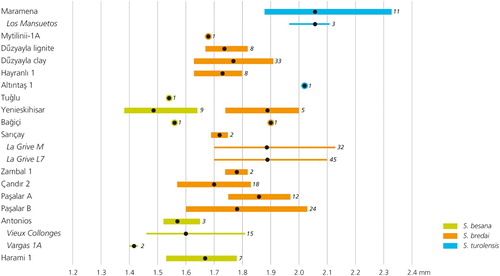
TABLE 6. Localities with Spermophilinus in southeastern Europe and Anatolia.
The evolutionary history of Spermophilinus is more complex than has been assumed previously. The oldest locality with Spermophilinus known at present is Harami 1 (MN 2). It is conceivable that Spermophilinus arrived in western Eurasia from the east. In western Europe, Spermophilinus may have developed further along one line, which is characterized by gradual size increase and is represented by the species S. besana, S. bredai, and S. turolensis, respectively. In the region of southeastern Europe and Anatolia, two lineages of Spermophilinus coexisted during the middle Miocene (). It can only be speculated whether S. besana from Harami 1, Kaplangı 1, and Antonios developed into the smaller (S. besana) or into the larger species (S. bredai) from Sarıçay, Bağiçi, and Yenieskihisar, or into both. Development into the smaller species would imply size decrease in the course of the early, middle, and early late Miocene. Bosma et al. (Citation2013) suggested that S. turolensis from Altıntaş 1 might be an immigrant into the area. It now appears more probable that S. (cf.) bredai from Hayranlı 1, Düzyayla clay and lignite (Bosma et al., Citation2013), and Mytilinii (Vasileiadou and Sylvestrou, Citation2009) are a continuation of the middle Miocene S. besana lineage, and that S. turolensis from Altıntaş 1 (Bosma et al., Citation2013) and Maramena (De Bruijn, Citation1995) have their roots in middle Miocene S. bredai. Spermophilinus giganteus may have originated from S. turolensis.
Our studies show that the genus Spermophilinus is not very useful for detailed biostratigraphic analysis. This in spite of the fact that remains of its species are frequently found in Miocene deposits. Much more work is needed to reconstruct the major steps in its evolution and its distribution in space and in time.
Genus TAMIAS Illiger, Citation1811
Remark—We consider Eutamias Trouessart, Citation1880, and Neotamias Howell, Citation1929, as subgenera of the genus Tamias (cf. Nowak, Citation1999; Wilson and Reeder, Citation2005).
Type Species—Sciurus striatus Linnaeus, Citation1758.
Fossil Species Recognized from the Old World—Tamias allobrogensis Mein and Ginsburg, Citation2002 (MN 7, middle Miocene); Tamias anatoliensis Bosma, De Bruijn, and Wessels, Citation2013 (MN 9, late Miocene); Tamias atsali De Bruijn, Citation1995 (MN 13, late Miocene); Eutamias ertemtensis Qiu, Citation1991 (MN 13?, late Miocene); Tamias eviensis De Bruijn, Van der Meulen, and Katsikatsos, Citation1980 (MN 4, early Miocene); Eutamias lishanensis Qiu, Zheng, and Zhang, Citation2008 (MN 10?, late Miocene); Eutamias orlovi Sulimski, Citation1964 (MN 15, early Pliocene); Eutamias sihongensis Qiu and Liu, Citation1986 (MN 4?, early Miocene); Eutamias urialis Munthe, Citation1980 (MN 9?, late Miocene).
TAMIAS cf. EVIENSIS De Bruijn, Van der Meulen, and Katsikatsos, Citation1980
Locality—Yapıntı (Yap).
Level—Early Miocene, MN 3 to early MN 4 (Ünay et al., Citation2001).
Material—Yap-101, a D4; Yap-103, -104, two M1/2s; Yap-106, an M3; Yap-108, -109, -110, three m2s.
Measurements—Yap-101: 1.40 × 1.45 mm; Yap-103: 1.72 × 2.05 mm; Yap-104: 1.73 × 2.10 mm; Yap-106: 1.77 × 1.77 mm; Yap-108: 2.08 × 2.13 mm; Yap-109: 1.85 × 1.91 mm; Yap-110: 1.91 × 2.11 mm.
Length/Width Scatter Plots—See .
FIGURE 11. Length/width scatter plots for D4, M1/2, M3, and m2 of Tamias species from Greece and Anatolia. Tamias cf. eviensis from Yapıntı, Anatolia (●) (this study), T. eviensis from the type locality, Aliveri, Greece (○) (De Bruijn et al., Citation1980), T. anatoliensis from the type locality, Altıntaş 1, Anatolia (□) (Bosma et al., Citation2013), and T. atsali from the type locality, Maramena, Greece (◊) (De Bruijn, Citation1995). Teeth from Aliveri and Maramena were remeasured to ensure precise comparison.
Description
D4—Specimen Yap-101 has a triangular shape because of the large and elongated, low parastyle. The parastyle forms part of the very low anteroloph. The tooth possesses a large hypocone, which is situated adjacent to the protocone, forming part of the low posteroloph. The lingual tooth wall shows a shallow valley between the protocone and the hypocone. The protoloph and the metaloph form a sharp ‘V,’ which is directed slightly anteriorly. The metaloph bears a large metaconule. There is a distinct, isolated mesostyle, which is situated close to the metacone.
M1/2—Of the two M1/2s, one is complete and unworn. The other is moderately worn and is slightly damaged: parts of the anterolingual, anterobuccal, and posterobuccal corners are missing. The small hypocone forms part of the posteroloph and is closely connected to the protocone. The protoloph and the metaloph are complete. They form a ‘V’ that points slightly in an anterior direction. The protoloph carries an indistinct protoconule. The metaloph bears two indistinct metaconules. The posteroloph is low, the anteroloph very low. Both teeth possess a small, elongated mesostyle. In the unworn specimen, a cuspule is present at the bottom of the valley between the protoloph and the metaloph, close to the mesostyle.
M3—The M3 is triangular in shape. It has a short posterior portion. The protocone and the paracone are well developed. The protoloph is relatively low and narrow, without a protoconule. The low anteroloph is connected to the protocone. The tooth is bordered on the posterior side by a low and narrow posteroloph. The posteroloph extends between the protocone and the paracone and incorporates an indistinct mesostyle. The surface of the valley between the protoloph and the posteroloph is rugose.
m2—The metaconid is by far the most prominent and highest cusp. The entoconid is low and elongated and forms part of the posterolophid. The very low metalophid ends at the buccal side of the metaconid, against the anterolophid. In two teeth, the anterolophid broadens at its buccal end into an anteroconid. All three teeth possess a distinct mesoconid, which forms part of a narrow ectolophid. One tooth carries a small, elongated mesostylid. The surface of the central valley shows irregularities.
Discussion
The teeth from Yapıntı are morphologically very close to those of Tamias eviensis from the type locality Aliveri, Greece (De Bruijn et al., Citation1980), but they are, with the exception of the single M3, which is of about the same size as the M3s from Aliveri, about 15–20% larger (). Because of this size difference, and because the material available is very limited, we describe Tamias from Yapıntı as T. cf. eviensis, and not as T. eviensis, or as a new species. Ünay et al. (Citation2001) proposed that the age of the fauna from Yapıntı is MN 3 to early MN 4. The Yapıntı fauna might thus be slightly older than the fauna from Aliveri (MN 4; De Bruijn, Citation1995; Theocharopoulos, Citation2000). If T. cf. eviensis from Yapıntı and T. eviensis from Aliveri are directly related forms, this implies that size reduction took place during the early Miocene.
In our study of late Miocene Sciuridae from Anatolia (Bosma et al., Citation2013), we have proposed the presence in the region of at least three lineages of Tamias: (1) a T. anatoliensis–T. atsali lineage, represented by T. anatoliensis from Altıntaş 1 and 2 (MN 9; Anatolia) and by T. (aff.) atsali from Maramena and Ano Metochi 3 (MN 13; Greece) and Kangal 1 and Sűleymanlı 2 (MN 13; Anatolia); (2) a T. eviensis lineage, represented by T. (cf.) eviensis from Aliveri (MN 4; Greece) and Karaőzű and Hayranlı 1 (MN 10–11; Anatolia); and (3) a lineage represented by Tamias sp. from Kangal 1 (MN 13; Anatolia). The two teeth from Kangal 1, described as Tamias sp., are a P4 (length: 1.78, width: 1.98) and an M1/2 (length: 1.74, width: 2.14). Strikingly, the D4 from Yapıntı and the P4 from Kangal 1 both show a large metaconule. The M1/2s from Yapıntı and Kangal 1 are of a similar size. As an alternative hypothesis, we carefully speculate that T. cf. eviensis from Yapıntı and Tamias sp. from Kangal 1 belong to the same, third lineage, covering a time span between MN 3/4 and MN 13.
Genus ATLANTOXERUS Major, Citation1893
Type Species—Sciurus getulus Linnaeus, Citation1758.
Formally Named Species—Getuloxerus adroveri De Bruijn and Mein, Citation1968 (MN 12, late Miocene); Getuloxerus blacki De Bruijn, Citation1967 (MN 4, early Miocene); Atlantoxerus cuencae Aguilar, Calvet, and Michaux, 1995 (late Miocene–early Pliocene); Sciurus getulus Linnaeus, Citation1758 (Recent); Atlantoxerus giganteus Wu, Citation1988 (MN 7–8?, middle Miocene); Atlantoxerus humboldti Kordikova, Heizmann, and De Bruijn, Citation2004 (MN 6–7, middle Miocene); Atlantoxerus huvelini Jaeger, Citation1977 (Pleistocene); Atlantoxerus junggarensis Wu, Citation1988 (MN 7–8?, middle Miocene); Atlantoxerus margaritae Adrover, Mein, and Moissenet, Citation1993 (MN 14, early Pliocene); Atlantoxerus martini Aguilar, Citation2002 (MN 3, early Miocene); Atlantoxerus orientalis Qiu, Citation1996 (MN 7–8?, middle Miocene); Atlantoxerus rhodius De Bruijn, Dawson, and Mein, Citation1970 (MN 14, early Pliocene); Getuloxerus tadlae Lavocat, Citation1961 (MN 6, middle Miocene); Atlantoxerus xiyuensis Wei, Citation2010 (middle Miocene).
ATLANTOXERUS ADROVERI (De Bruijn and Mein, Citation1968)
Localities—Bağiçi (Bag) and Yenieskihisar (YE).
Level—Local rodent zone H (tentatively correlated to MN 7 + 8) (Ünay et al., Citation2003; De Bruijn et al., Citation2013b).
Material—Twenty teeth from two localities. Bağiçi (seven teeth): Bag-888, an M1/2; Bag-898, a p4; Bag-890, -891, -893, -895, four m1s; Bag-900, an m2. Yenieskihisar (13 teeth): YE-191, a P3; YE-193, a P4; YE-195, -196, two M1/2s; YE-198, an M3; YE-201, -202, -203, three p4s; YE-208, an m1; YE-205, -206, -209, three m2s; YE-210, an m3.
Measurements—See .
TABLE 7. Measurements (in mm) of teeth of Atlantoxerus adroveri from Bağiçi and Yenieskihisar.
Description
P3—The occlusal surface of YE-191 has the shape of a rounded triangle. The tooth has a large interdental facet. An anterior crest connects two cusps, one high (paracone?) and one lower (protocone?).
P4—Specimen YE-193 is rather worn. It has a small and low anterior portion, with a short anteroloph. There is no posterior valley. The lingual tooth wall has a shallow, but sharp, depression between the protocone and the hypocone. A small, crest-like structure, which resembles a mesostyle, is situated close to the metacone.
M1/2—Specimen Bag-888 has a prominent hypocone, which is situated at the posterolingual tooth corner. The lingual tooth wall has shallow depressions between the protocone and the hypocone, and between the protocone and the anteroloph. A large metaconule is connected to the metacone by a strong crest and to the posteroloph by a very short and indistinct crest. The posteroloph is very low. There is hardly any posterior valley. The two M1/2s from Yenieskihisar, both from the left side, are very worn and may have belonged to the same individual. The posterior valley has disappeared. The lingual tooth wall has shallow depressions between the protocone and the hypocone, and between the protocone and the anteroloph. A low, crest-like elevation, comparable to a mesostyle, is present between the paracone and the metacone. An M1/2 in the type material (UU) has a similar structure. De Bruijn and Mein (Citation1968) incorrectly used the absence of a mesostyle in M1/2 as a diagnostic character for Atlantoxerus adroveri.
M3—The occlusal surface of the very worn YE-198 has the shape of a triangle. It shows the remnants of a large metaconule and protoconule.
p4—The protoconid and the metaconid are connected by a short metalophid, which is low in the middle. The entolophid, connecting the entoconid and the posterolophid, has an irregular appearance. In one p4, the entolophid does not reach the posterolophid. In the same tooth, the posterolophid shows a low and narrow inward curve between the hypoconulid and the hypoconid. The anteroconid is small and of variable size. The ectolophid is low and narrow. There is no mesostylid.
m1 and m2—The metalophid is incomplete in all m1s and m2s. The anterolophid is short. The entolophid is a strong, often irregular crest. The posterolophid and the posterior tooth wall show a rather deep inward curve/depression between the hypoconid and the point where the entolophid and the posterolophid meet. The ectolophid is low and narrow. None of the teeth bears a distinct mesostylid.
m3—Specimen YE-210 is very worn and is broken. The occlusal surface shows the remnants of the (long) anterolophid, the incomplete metalophid, the irregular entolophid, and the narrow ectolophid.
Discussion
The unexpected identification of Atlantoxerus adroveri in the faunas from Bağiçi and Yenieskihisar is based on the very close resemblance, both in size and in morphology, between our material and the type material of the species in UU. The type locality of A. adroveri, Los Mansuetos (Spain), is the reference locality for MN 12. So far, A. adroveri is known from MN 9– MN 14 localities in Spain only (Fortelius, Citation2017). It appears that the species had a much wider geographical and temporal range than was hitherto assumed. Relationships between A. adroveri and other Atlantoxerus species were discussed by Peláez-Campomanes (Citation2001).
Similar species are Atlantoxerus orientalis and A. humboldti. Qiu (Citation1996) described A. orientalis on the basis of nine isolated cheek teeth, some of them broken, from Tunggur-Moergen II (China). The age of the Moergen II fauna is supposed to be MN 7 + 8. Atlantoxerus orientalis has since been reported from other localities in China, with ages of MN 7 + 8 and MN 6 (Fortelius, Citation2017). Atlantoxerus humboldti was described by Kordikova et al. (Citation2004) on the basis of a fragment of an upper jaw with D4–M2, a fragment of a lower jaw with m1–m2, and 40 isolated cheek teeth from Mynsualmas (western Kazakhstan). The proposed age of the fauna from Mynsualmas is MN 6 to MN 7 + 8. Future studies may reveal that Atlantoxerus adroveri, A. orientalis, and A. humboldti are very closely related or even identical species. The two isolated cheek teeth, an M3 and a p4, reported from the MN 7 + 8 locality Bayraktepe I in Anatolia as Atlantoxerus sp. I by Ünay (Citation1981), may belong to the same (group of) species.
Subfamily PTEROMYINAE Brandt, Citation1855
Genus HYLOPETES Thomas, Citation1908
Type Species—Sciuropterus everetti Thomas, Citation1908.
Synonyms—Neopetes Daxner-Höck, Citation2004 (see Bosma et al., Citation2013); Pliopetes Kretzoi, Citation1959 (see Bouwens and De Bruijn, Citation1986).
Remarks on the Contents of Hylopetes—Following Bouwens and De Bruijn (Citation1986), we classify all fossil teeth with a Hylopetes or Petinomys morphology as Hylopetes (see also Bosma et al., Citation2013).
Fossil Species Recognized—Petinomys auctor Qiu, Citation1991 (MN 13?, late Miocene; see discussion in Bosma et al., Citation2013); Hylopetes debruijni Mein and Ginsburg, Citation2002 (MN 7, middle Miocene); Hylopetes hoeckarum De Bruijn, Citation1998 (MN 4, early Miocene); Pliopetes hungaricus Kretzoi, Citation1959 (MN 15, early Pliocene); Hylopetes macedoniensis Bouwens and De Bruijn, Citation1986 (MN 13, late Miocene); Hylopetes magistri Reumer and Van den Hoek Ostende, Citation2003 (early Pleistocene).
HYLOPETES sp.
Localities—Kılçak 0 (Ki0) and Kılçak 0” (Ki0”).
Level—Kılçak 0 and 0": Local rodent zone B (tentatively correlated to MN 1) (De Bruijn et al., Citation2013b).
Material—Ki0-11, a P4; Ki0”-11, a P4.
Measurements—Ki0-11: 1.85 × 1.90 mm; Ki0”-11: 1.89 × 2.06 mm.
Description
P4—Specimen Ki0-11 is notably worn. Its shape is triangular because of the large, elongated parastyle. The tooth has a protocone, a (damaged) paracone, a metacone, a protoloph, and a metaloph. The protoloph, the protocone, and the metaloph form a ‘U.’ The protoloph and the metaloph have an irregular appearance. The posteroloph is practically worn away. Wear facets are present at the posterior and anterior sides of the tooth. The anterior wear facet is situated lingually to the parastyle. Specimen Ki0”-11 is only slightly worn. The protocone is slightly higher than the paracone and the metacone, the elongated parastyle is somewhat lower. There is a deep and sharp depression between the parastyle and the paracone. The slightly irregular protoloph and metaloph are situated partly in parallel. The protoloph connects to the anterior side of the protocone, the metaloph to the posterior side. The protoloph has an incision in the middle. The metaloph bears a small metaconule. The posteroloph is continuous with the protocone, without showing a distinct hypocone. The anteroloph is much lower than the other crests. A small mesoloph is present between the paracone and the metacone. The central valley and the valley between the parastyle and the protoloph show finely wrinkled enamel.
Discussion
The material available is too limited to assign the two teeth to a particular Hylopetes species, either known or new. Possibly, they belong to H. hoeckarum, the oldest fossil Hylopetes described so far. Hylopetes hoeckarum is known from a number of localities in central Europe (Austria, Hungary, and Switzerland; Daxner-Höck, Citation2004) that vary in age from MN 4 to MN 11. The occlusal patterns of our teeth closely resemble that of the P4 of H. hoeckarum from Richardhof-Wald (MN 10) figured by Daxner-Höck (2004:pl. 2, fig. 6). Our specimens are slightly larger than the two P4s from Richardhof-Wald and the one P4 from Schernham (MN 10) in Daxner-Höck’s paper (Citation2004:table 2).
Genus MIOPETAURISTA Kretzoi, Citation1962
Type Species—Sciurus göriachensis Hofmann, Citation1893 (syn. of Sciurus gibberosus Hofmann, Citation1893; see Daxner-Höck, Citation2004).
Species Recognized—Miopetaurista asiatica Qiu, Citation2002 (MN 11 or 12?, late Miocene); Cryptopterus crusafonti Mein, Citation1970 (MN 9, late Miocene); Miopetaurista dehmi De Bruijn et al., Citation1980 (MN 3, early Miocene); Cryptopterus gaillardi Mein, Citation1970 (MN 7 or 8, middle Miocene); Sciurus göriachensis Hofmann, Citation1893 (MN 5, middle Miocene); Sciuropterus lappi Mein, Citation1958 (MN 4 or 5, early or middle Miocene); Cryptopterus neogrivensis Mein, Citation1970 (MN 8 or 7, middle Miocene); Cryptopterus thaleri Mein, Citation1970 (MN 14, early Pliocene).
MIOPETAURISTA cf. DEHMI De Bruijn, Van der Meulen, and Katsikatsos, Citation1980
Locality—Harami 1 (Har1).
Level—Local rodent zone C (tentatively correlated to MN 2) (De Bruijn et al., Citation2013b). Krijgsman et al. (Citation1996) correlated Harami 1 to chron C6Bn.2n, which indicates that the estimated absolute age of the locality is between 22.0 and 22.5 Ma (Hilgen et al., Citation2012).
Material—Har1-1731, -1732, -1734, three P4s; Har1-1733, -1735, -1736, three M1/2s.
Measurements—Har1-1731: 2.40 × 2.70 mm; Har1-1732: 2.14 × 2.48 mm; Har1-1734: – × –; Har1-1733: 2.49 × 3.02 mm; Har1-1735: – × –; Har1-1736: 2.48 × 3.05 mm.
Description
P4—One P4 is complete and little worn, one is rolled, and one is a fragment, representing the lingual tooth half. The protoloph, the elongated protocone, and the metaloph more or less form a ‘U.’ The paracone and the metacone are prominent. The hypocone is indistinct and is part of the posteroloph. The elongated parastyle forms part of the anteroloph. The anteroloph and the posteroloph are low. Two short additional crests are present: a low and narrow mesoloph and a low and narrow crest that extends from the protocone into the central valley. The protoloph, the metaloph, and two additional crests are irregular structures in the little worn specimen. This tooth has a mesoloph connecting buccally to the paracone, and the crest coming from the protocone connects to the middle of the protoloph.
M1/2—Two M1/2s are more or less complete; one is a fragment, representing the posterolingual tooth half. The protocone, paracone, and metacone are prominent. The protoloph and the metaloph form an obtuse ‘V,’ which is directed anteriorly. The anteroloph and the posteroloph are low. The small hypocone is incorporated in the posteroloph. A mesoloph and a second additional crest, as described for P4, are present as tiny structures in the central valley.
Discussion
The identification at the species level of the Miopetaurista material from Harami 1 is complicated for a number of reasons. Little is known about the intraspecific variation of the various Miopetaurista species, because the amount of material collected has always been small (Ziegler and Fahlbusch, Citation1986; De Bruijn, Citation1998). The sample from Harami 1 includes only P4s and M1/2s. Of the three P4s, one is rolled and one is a fragment. Also, one of the three M1/2s is a fragment, whereas the occlusal surface of one of the complete M1/2s is rather worn. However, the morphological details observable, in combination with the size of the teeth, indicate that the material belongs to M. dehmi. Most convincing are the more or less parallel protoloph and metaloph in P4, and the presence of a mesoloph and of an extra loph between the protoloph and the metaloph in M1/2 as well as P4. The type material from Wintershof-West (Germany; reference locality for MN 3), described and figured by De Bruijn et al. (Citation1980), consists of six isolated teeth: a P4, an M1/2, an M3, a p4, an m1 (holotype), and an m3. The P4s from Harami 1 are slightly smaller than the P4 from Wintershof-West. The M1/2s from Harami 1 have about the same size as the M1/2 in the type material. We present the teeth from Harami 1 as M. cf. dehmi, and not as M. dehmi, because of the uncertainties mentioned above.
Miopetaurista dehmi is the oldest and smallest Miopetaurista species described so far. It is known from several localities in central (France, Germany, Poland, Czech Republic, Hungary, and Austria) and southeastern (Serbia and Greece) Europe, ranging in age from MN 3 to MN 6 (Fortelius, Citation2017). Marković et al. (Citation2016) reported M. cf. dehmi from the Serbian locality Sibnica (MN 4). The Greek locality with M. dehmi is the MN 4 locality Aliveri (De Bruijn et al., Citation1980). The presence of the species in Harami 1 (MN 2) indicates that M. dehmi existed earlier than was previously supposed.
Genus ALIVERIA De Bruijn, Van der Meulen, and Katsikatsos, Citation1980
Type Species—Aliveria brinkerinki De Bruijn, Van der Meulen, and Katsikatsos, Citation1980.
Species Recognized—Aliveria brinkerinki De Bruijn et al., Citation1980 (MN 4, early Miocene); Aliveria luteyni De Bruijn et al., Citation1980 (MN 4, early Miocene).
ALIVERIA LUTEYNI De Bruijn, Van der Meulen, and Katsikatsos, Citation1980
Localities—Kaplangı 1 (Kap1) and Kaplangı 2 (Kap2).
Level—Kaplangı 1 and 2: Local rodent zone E (tentatively correlated to MN 4) (De Bruijn et al., Citation2013b).
Material—Six teeth from two localities. Kaplangı 1 (five teeth): Kap1-121, -122, two D4s; Kap1-124, an M1/2; Kap1-127, -128, two m3s; Kaplangı 2 (one tooth): Kap2-101, an m3.
Measurements—Kap1-121: 1.64 × 1.91 mm; Kap1-122: 1.70 × 1.80 mm; Kap1-124: – × – ; Kap1-127: 2.41 × 2.03 mm; Kap1-128: 2.35 × 2.05 mm; Kap2-101: 2.37 × 2.01 mm.
Description
D4—The shape of the teeth is triangular, because of the large parastyle. The protocone, paracone, metacone, protoloph, and metaloph are regular cusps and crests. The protoloph and the metaloph form a ‘V,’ which is directed slightly anteriorly. The protoloph is straight, and without a protoconule. The metaloph carries an elongated metaconule. The hypocone is prominent. The lingual tooth wall has a depression between the protocone and the hypocone. Both teeth possess a small, but distinct and isolated mesostyle. The posteroloph is low.
M1/2—Kap1-124 is somewhat damaged. Observable are the protocone, paracone, metacone, protoloph, metaloph, hypocone, and part of the anteroloph. The metaloph is broad in the middle, which is reminiscent of an elongated metaconule. The protoloph and the metaloph form a sharp ‘V.’
m3—The teeth show the protoconid, metaconid, hypoconid, and entoconid. The pointed metaconid is by far the highest cusp. The entoconid is elongated; it forms the lingual end of the posterolophid. The metalophid is short; it reaches maximally to halfway along the metaconid. The anterolophid extends from the metaconid to the buccal tooth side. Its buccal end is clearly separated from the protoconid. The prominent mesoconid extends into the buccal valley. The teeth possess a small mesostylid. The central valley of the two little worn specimens bears irregular small crests and cuspules.
Discussion
Aliveria luteyni is the smaller of the two Aliveria species described. The material from Kaplangı 1 and 2 is very limited, and D4 and m3 are the only undamaged elements. We nevertheless identify it as A. luteyni, and not as A. cf. luteyni, because of the very characteristic morphology of m3 and the correspondence in size between the teeth from Kaplangı and those in the type assemblage of the species.
Aliveria luteyni was so far known from the type locality Aliveri (Greece; MN 4) only. There is agreement between the age of the fauna from Aliveri and the proposed age of the faunas from Kaplangı 1 and 2, which is also MN 4 (De Bruijn et al., Citation2013b). Teeth from the Serbian MN 4 locality Sibnica have recently been described by Marković et al. (Citation2016) as Aliveria aff. luteyni.
Genus ALBANENSIA Daxner-Höck and Mein, Citation1975
Type Species—Sciuropterus albanensis Major, Citation1893.
Species Recognized—Sciuropterus albanensis Major, Citation1893 (MN 7 + 8, middle Miocene); Sciuropterus albanensis quiricensis Villalta, Citation1950 (MN 7 + 8, middle Miocene); Sciuropterus grimmi Black, Citation1966 (MN 9, late Miocene); Sciurus sansaniensis Lartet, Citation1851 (MN 6, middle Miocene).
ALBANENSIA ALBANENSIS (Major, Citation1893)
Locality—Sarıçay (SC).
Level—Local rodent zone H (tentatively correlated to MN 7 + 8) (Ünay et al., Citation2003).
Material—SC-141, a right P4; SC-143, a left M1/2; SC-144, a right M1/2; SC-146, a left m1; SC-147, a right m1; SC-149, a right m2; SC-150, a right m3.
Measurements—SC-141: 3.64 × 3.50 mm; SC-143: 3.11 × 3.84 mm; SC-144: 2.96 × 3.68 mm; SC-146: 2.98 × 3.44 mm; SC-147: 3.06 × 3.44 mm; SC-149: 3.16 × 3.67 mm; SC-150: 3.56 × 3.09 mm.
Description
P4—The tooth has the shape of a trapezoid. It has a very large parastyle, which shows, apart from the large central wear facet, two small facets, as described by Mein (1970:17) for P4 in the (type) material from La Grive, France. The protocone, the paracone, and the metacone are high and pointed cusps. The protoloph and the metaloph are rather irregular crests, which form a ‘V.’ The protoloph is directed transversely, the metaloph slightly anteriorly in the lingual direction. The metaloph carries a large and rounded metaconule. There is no hypocone. A small and isolated mesostyle, which is situated nearer to the paracone than to the metacone, is present at the buccal tooth border. The degree of complexity of the occlusal surface is very similar to that of the M2 from La Grive figured by Engesser (1972:167, fig. 58-1).
M1/2—Both teeth show (the remnants of) the protocone, paracone, metacone, hypocone, protoloph, metaloph, and anteroloph. The protoloph and the metaloph form a ‘V,’ which points slightly anteriorly. One tooth has traces of a double metaconule. The small and low mesostyle is rather connected to the paracone than to the metacone.
m1 and m2—The teeth show (the remnants of) the protoconid, the high metaconid, the hypoconid, the entoconid, a low and (probably) incomplete metalophid, an anterolophid (m1) or anteroconid (m2), and the posterolophid. The m2 possesses a small mesoconid. All three teeth have a mesostylid, which is connected to the metaconid by a high crest and separated from the entoconid by a depression.
m3—The posterior half of the m3 is very broad. The occlusal surface shows (the remnants of) the protoconid, the high metaconid, the hypoconid, the small and low entoconid, a very small anteroconid, and the posterolophid. A distinct mesoconid can be observed at the buccal tooth border. The tooth carries a mesostylid as in m1 and m2.
Discussion
Albanensia albanensis takes, both in size and in age, a position between A. sansaniensis (smaller and older) and A. grimmi (larger and younger). The identification of our material as Albanensia albanensis is mainly based on the morphology of the single, but very characteristic, P4, in combination with the size of the teeth. With the exception of the P4, all teeth are very worn and may have belonged to the same individual.
Localities with Albanensia albanensis were until now all in western and central Europe: La Grive M (type locality) and L7, France (Mein, Citation1970); Anwil, Switzerland (Engesser, Citation1972); and St. Stefan bei Gratkorn, Austria (Daxner-Höck, Citation2010). The age of these localities is MN 7 + 8 (Mein and Ginsburg, Citation2002, consider La Grive M and L7 to be MN 7). According to Mein (Citation1970), A. albanensis has also been found at the locality Opole, Poland (MN 6/MN 7 + 8). The presence of A. albanensis in the fauna from Sarıçay is in agreement with its proposed age (MN 7 + 8).
Another locality in Anatolia, Çandir 2, has yielded two M1/2s of Albanensia sansaniensis (De Bruijn et al., Citation2003). The age of the Çandir 2 fauna is currently considered to be MN 5 (De Bruijn et al., Citation2003; Ünay et al., Citation2003). Like A. albanensis, A. sansaniensis outside of Anatolia is known only from localities in western and central Europe with, in this case, ages of MN 5 and MN 6 (Fortelius, Citation2017). It is conceivable that the two species represent the same evolutionary line.
Genus BLACKIA Mein, Citation1970
Type Species—Blackia miocaenica Mein, Citation1970.
Formally Named Species—Blackia miocaenica Mein, Citation1970 (MN 7 + 8, middle Miocene); Blackia polonica Black and Kowalski, Citation1974 (MN 14, early Pliocene); Blackia ulmensis Werner, Citation1994 (MN 2, early Miocene); Blackia woelfersheimensis Mein, Citation1970 (MN 15, early Pliocene).
BLACKIA sp.
Localities—Sarıçay (SC) and Yenieskihisar (YE).
Level—Sarıçay and Yenieskihisar: Local rodent zone H (tentatively correlated to MN 7 + 8) (Ünay et al., Citation2003).
Material—SC-131, an m1; YE-181, an m2.
Measurements—SC-131: 1.34 × 1.66 mm; YE-181: 1.51 × 1.80 mm.
Description
m1 and m2—The m1 from Sarıçay is slightly rolled, and the m2 from Yenieskihisar is very worn. The two teeth are small, low-crowned, have the shape of a trapezoid, and have a simple occlusal pattern. Crenulation of the enamel surface is vaguely visible in the tooth from Sarıçay.
Discussion
The two teeth unmistakably belong to Blackia, because of their small size and typical morphology. Although their measurements fall within the range of Blackia miocaenica, we present them as Blackia sp., the material available being too scanty for specific determination.
Blackia miocaenica is known from a large number of localities in western, central, eastern, and southeastern Europe, including Sibnica, Serbia (MN 4; Marković et al., Citation2016), and Aliveri, Greece (MN 4; De Bruijn et al., Citation1980). The species has a very long time range, from the late Oligocene (MP 30) until the late Miocene (MN 11) (Fortelius, Citation2017).
CONCLUSIONS
Collecting of micromammal remains in early and middle Miocene deposits in Anatolia has led to the recovery of several species of Sciuridae. These species represent both subfamilies: Sciurinae (ground and tree squirrels) and Pteromyinae (flying squirrels). Their occurrences at the various localities are summarized in .
FIGURE 12. Occurrences of Sciuridae at 15 early and middle Miocene localities in Anatolia, Turkey.
The oldest localities, Kargı 2 (possibly latest Oligocene) and Kılçak 0, 0”, 3a, and 3b (MN 1), have yielded a ground squirrel, Palaeosciurus (cf.) feignouxi, a tree squirrel, Dehmisciurus cf. obtusidens, and a flying squirrel, Hylopetes sp. Palaeosciurus feignouxi is well known from western and central Europe (MP 30 to MN 3). Dehmisciurus obtusidens is rare, and in the region of southeastern Europe and Anatolia it is known from Bosnia and Herzegovina (latest Oligocene) and Serbia (MN 4) only. Other localities with Dehmisciurus obtusidens are in Germany (MN 2a and MN 3) and in Spain (MN 4). The Hylopetes species is possibly Hylopetes hoeckarum. Bosma et al. (Citation2013) described Hylopetes macedoniensis from the late Miocene (MN 12) localities Düzyayla clay and lignite in Anatolia. Hylopetes from Kılcak being larger, it appears unlikely that this species is a direct ancestor of Hylopetes macedoniensis from Düzyayla.
At the MN 2 localities Harami 1 and 3, another ground squirrel, Spermophilinus besana, and another flying squirrel, Miopetaurista cf. dehmi, have been found, next to Dehmisciurus obtusidens. As at Kılçak, Dehmisciurus obtusidens is represented by one tooth only. Spermophilinus besana is well known from localities in western, central, and southeastern Europe, the majority having an MN 4 or MN 5 age. Spermophilinus besana from Harami 1 is presently the oldest Spermophilinus described. This may suggest that the genus originated in Asia, rather than in Europe. Localities with Miopetaurista dehmi are in central and southeastern Europe. Their ages range from MN 3 to MN 6. Miopetaurista cf. dehmi from Harami 1 is the oldest record of Miopetaurista. Here also, an Asian origin of the genus is indicated.
From the MN 3 locality Keseköy, a ground squirrel has been recovered that shows considerable resemblance to Palaeosciurus feignouxi. The species is presented here as Palaeosciurus aff. feignouxi. However, although smaller, its teeth show close similarity to teeth of Protospermophilus kelloggi, a ground squirrel collected from the early Miocene Split Rock Formation in Wyoming, U.S.A., as well. The Oligocene–Miocene genus Protospermophilus presently comprises six different species and is confined to North America. It may be questioned whether the squirrel from Keseköy should be described as a new Protospermophilus species. We have chosen to assign it to the European genus Palaeosciurus to avoid questions, not to be answered with our present knowledge, regarding migration of this group of rodents between North America and the region of Anatolia, in or before the early Miocene.
Yapıntı (MN 3/MN 4) is the only locality with a chipmunk, Tamias cf. eviensis. We carefully speculate that this species is a predecessor of Tamias sp. from the late Miocene (MN 13) locality Kangal 1 in Anatolia (Bosma et al., Citation2013).
At the Kaplangı localities (MN 4), the ground squirrel Palaeosciurus cf. sutteri and the flying squirrel Aliveria luteyni have been found, next to Spermophilinus cf. besana. Palaeosciurus sutteri is known from localities in central and southeastern Europe, with ages of MN 3 to MN 6. The only locality with Aliveria luteyni was before now its type locality (MN 4) in Greece.
The Sciuridae are most diverse in the assemblages from the middle Miocene (MN 7 + 8) localities Sarıçay, Bağiçi, and Yenieskihisar. Unexpectedly, we could identify two Spermophilinus species: Spermophilinus besana and Spermophilinus bredai. This indicates that, at least in Anatolia, two lineages of Spermophilinus developed in parallel. Bosma et al. (Citation2013) described Spermophilinus turolensis from the late Miocene (MN 9) locality Altıntaş 1 in Anatolia and Spermophilinus bredai from the Anatolian MN 10–12 localities Hayranlı 1, Düzyayla clay, and Düzyayla lignite. In our present paper, we propose that late Miocene Spermophilinus bredai and Spermophilinus turolensis are descendants of middle Miocene Spermophilinus besana and Spermophilinus bredai. Another middle Miocene locality with Spermophilinus bredai is Zambal 1 (MN 6).
Bağiçi and Yenieskihisar are the only localities in Anatolia with a member of the ground squirrel tribe Xerini Murray, Citation1866: Atlantoxerus adroveri. So far, this species was restricted to Spain. Atlantoxerus adroveri is possibly very closely related to, or even identical to, species described from the middle Miocene of China and Kazakhstan: Atlantoxerus orientalis and Atlantoxerus humboldti.
Two MN 7 + 8 localities yielded flying squirrels: Albanensia albanensis was found at Sarıçay, Blackia sp. (most probably Blackia miocaenica) at Sarıçay and Yenieskihisar. Outside Anatolia, Albanensia albanensis is known from MN 7 + 8 localities in western and central Europe. The presence in Anatolia of Albanensia albanensis during MN 7 + 8, and of Albanensia sansaniensis during MN 5 (De Bruijn et al., Citation2003), strengthens the assumption that Albanensia sansaniensis is a direct ancestor of Albanensia albanensis. Blackia miocaenica is a very common species in Europe, collected from localities ranging in age from latest Oligocene to late Miocene.
Practically all sciurid taxa presented in this paper are known from regions in Europe only. This indicates that during the early and middle Miocene, Anatolia was connected to the European part of the Eurasian continent, with ample possibilities for faunal exchange.
Of the 15 localities dealt with in this paper, seven have yielded merely ground squirrels, one only a tree squirrel, and one only a flying squirrel. From six localities, ground squirrels (in one case together with a tree squirrel) as well as flying squirrels were recovered. Possibilities to draw conclusions with respect to the paleoecology represented are very limited, because the amounts of sediment processed to obtain the material differs greatly between localities, as do the numbers of teeth allocated to the various species. All the material described comes from lacustrine deposits. This, together with the frequent occurrence of flying squirrels, indicates that most localities represent a wet, but forested environment.
ACKNOWLEDGMENTS
We are grateful to the General Directorate of the MTA, Ankara, Turkey, for supporting, during many years, the informal collaboration with members of the Department of Earth Sciences of Utrecht University. The project from which this paper is a result would have been impossible without the local knowledge and dedication of E. Ünay and G. Saraç. We owe them many thanks. We thank P. Mein (Lyon, France) for the loan of material of Spermophilinus besana from Vieux Collonges (France) and for sharing with us measurements of M1/2 of Spermophilinus bredai from La Grive L7 and M (France). We acknowledge the CM for the loan of Protospermophilus kelloggi material from Split Rock (Wyoming, U.S.A.). M. Stoete (Communication and Marketing, Faculty of Geosciences, Utrecht University) assisted with the preparation of the figures. We thank S. Sen (Paris, France) and an anonymous reviewer of this paper for their constructive and helpful comments.
ORCID
Wilma Wessels https://orcid.org/0000-0001-9027-9698
Related Research Data
LITERATURE CITED
- Adrover, R., P. Mein, and E. Moissenet. 1993. Los Sciuridae del Turoliense superior, Rusciniense y Villanyense de la región de Teruel. Paleontologia i Evolució 26-27:85–106.
- Aguilar, J.-P. 2002. Les sciuridés des gisements karstiques du Miocène inférieur à moyen du sud de la France: nouvelles espèces, phylogénie, paléoenvironnement. Geobios 35:375–394. doi: 10.1016/S0016-6995(02)00033-5
- Aguilar, J.-P., M. Calvet, and J. Michaux. 1995a. Les rongeurs du gisement karstique Miocène supérieur de Castelnou 1 (Pyrénées-Orientales, France). Geobios 28:501–510. doi: 10.1016/S0016-6995(95)80027-1
- Aguilar, J.-P., L. de Bonis, C. Denys, and G. D. Koufos. 1995b. Découverte d’un Spermophilinus (Rodentia, Mammalia) dans le gisement Vallésien du ‘Ravin de la Pluie’ (Macédoine, Grèce). Annales géologiques des Pays Helléniques 36:645–648.
- Aldana Carrasco, E. J. 1992. Los Sciurinae (Rodentia, Mammalia) del Mioceno de la Cuenca del Vallès-Penedès (Cataluña, España). Treballs del Museu de Geologia de Barcelona 2:69–97.
- Black, C. C. 1963. A review of the North American Tertiary Sciuridae. Bulletin of the Museum of Comparative Zoology, Harvard University 130:109–248.
- Black, C. C. 1966. Tertiary Sciuridae (Mammalia: Rodentia) from Bavaria. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 6:51–63.
- Black, C. C., and K. Kowalski. 1974. The Pliocene and Pleistocene Sciuridae (Mammalia, Rodentia) from Poland. Acta Zoologica Cracoviensia 19:461–486.
- Bosma, A. A., H. de Bruijn, and W. Wessels. 2013. Late Miocene Sciuridae (Mammalia, Rodentia) from Anatolia, Turkey. Journal of Vertebrate Paleontology 33:924–942. doi: 10.1080/02724634.2013.755990
- Bouwens, P., and H. de Bruijn. 1986. The flying squirrels Hylopetes and Petinomys and their fossil record. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 89:113–123.
- Bowdich, T. E. 1821. An Analysis of the Natural Classifications of Mammalia, for the Use of Students and Travellers. J. Smith, Paris, France, 115 pp.
- Brandt, J. F. 1855. Beiträge zur nähern Kenntniss der Säugethiere Russlands. Mémoires mathématiques, physiques et naturelles 7:1–365. Kaiserliche Akademie der Wissenschaften, St. Petersburg, Russia.
- Cope, E. D. 1879. Second contribution to a knowledge of the Miocene fauna of Oregon. Proceedings of the American Philosophical Society 18:370–376.
- Costeur, L., O. Maridet, S. Peigné, and E. P. J. Heizmann. 2012. Palaeoecology and palaeoenvironment of the Aquitanian locality Ulm-Westtangente (MN2, Lower Freshwater Molasse, Germany). Swiss Journal of Palaeontology 131:183–199. doi: 10.1007/s13358-011-0034-3
- Cuenca, G., J. I. Canudo, C. Laplana, and J. A. Andres. 1992. Bio y cronoestratigrafia con mamíferos en la Cuenca Terciaria del Ebro: ensayo de síntesis. Acta Geológica Hispánica 27:127–143.
- Cuenca Bescós, G. 1988. Revisión de los Sciuridae del Aragoniense y del Rambliense en la fosa de Calatayud-Montalbán. Scripta Geologica 87:1–116.
- Daxner-Höck, G. 1998. Säugetiere (Mammalia) aus dem Karpat des Korneuburger Beckens. 3. Rodentia und Carnivora. Beiträge zur Paläontologie (Wien) 23:367–407.
- Daxner-Höck, G. 2004. Flying squirrels (Pteromyinae, Mammalia) from the Upper Miocene of Austria. Annalen des Naturhistorischen Museums in Wien 106A:387–423.
- Daxner-Höck, G. 2010. Sciuridae, Gliridae and Eomyidae (Rodentia, Mammalia) from the Middle Miocene of St. Stefan in the Gratkorn Basin (Styria, Austria). Annalen des Naturhistorischen Museums in Wien 112A:507–536.
- Daxner-Höck, G., and P. Mein. 1975. Taxonomische Probleme um das Genus Miopetaurista Kretzoi, 1962 (Fam. Sciuridae). Paläontologische Zeitschrift 49:75–77. doi: 10.1007/BF02988067
- De Bruijn, H. 1967. Gliridae, Sciuridae y Eomyidae (Rodentia, Mammalia) miocenos de Calatayud (provincia de Zaragoza, España) y su relación con la bioestratigrafía del área. Boletín del Instituto Geológico y Minero de España 78:187–373.
- De Bruijn, H. 1995. The vertebrate locality Maramena (Macedonia, Greece) at the Turolian-Ruscinian boundary (Neogene). 8. Sciuridae, Petauristidae and Eomyidae (Rodentia, Mammalia). Münchner Geowissenschaftliche Abhandlungen A 28:87–102.
- De Bruijn, H. 1998. Vertebrates from the Early Miocene lignite deposits of the opencast mine Oberdorf (Western Styrian Basin, Austria): 6. Rodentia 1 (Mammalia). Annalen des Naturhistorischen Museums in Wien 99A:99–137.
- De Bruijn, H., and P. Mein. 1968. On the mammalian fauna of the Hipparion-beds in the Calatayud-Teruel Basin (Prov. Zaragoza, Spain). Part V. The Sciurinae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 71:73–90.
- De Bruijn, H., M. R. Dawson, and P. Mein. 1970. Upper Pliocene Rodentia, Lagomorpha and Insectivora (Mammalia) from the isle of Rhodes (Greece). III. The Rodentia, Lagomorpha and Insectivora. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 73:568–584.
- De Bruijn, H., Z. Marković, and W. Wessels. 2013a. Late Oligocene rodents from Banovići (Bosnia and Herzegovina). Palaeodiversity 6:63–105.
- De Bruijn, H., P. Y. Sondaar, and W. J. Zachariasse. 1971. Mammalia and Foraminifera from the Neogene of Kastellios hill, (Crete), a correlation of continental and marine biozones. I. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 74:1–13.
- De Bruijn, H., E. Ünay, and K. Hordijk. 2013b. A review of the Neogene succession of the Muridae and Dipodidae from Anatolia, with special reference to taxa known from Asia and/or Europe; pp. 564–580 in X. Wang, L. J. Flynn, and M. Fortelius (eds.), Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. Columbia University Press, New York.
- De Bruijn, H., A. J. van der Meulen, and G. Katsikatsos. 1980. The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 1. The Sciuridae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 83:241–261.
- De Bruijn, H., L. van den Hoek Ostende, E. Kristkoiz-Boon, M. Rummel, C. Theocharopoulos, and E. Ünay. 2003. Rodents, lagomorphs and insectivores, from the middle Miocene hominoid locality of Çandır (Turkey). Courier Forschungsinstitut Senckenberg 240:51–87.
- Dehm, R. 1950. Die Nagetiere aus dem Mittel-Miocän (Burdigalium) von Wintershof-West bei Eichstätt in Bayern. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Abhandlungen B 91:321–428.
- Downs, T. 1956. The Mascall fauna from the Miocene of Oregon. University of California Publications in Geological Sciences 31:199–354.
- Engesser, B. 1972. Die obermiozäne Säugetierfauna von Anwil (Baselland). Tätigkeitsberichte der Naturforschenden Gesellschaft Baselland 28:37–363.
- Fejfar, O., Z. Dvorák, and E. Kadlecová. 2003. New record of Early Miocene (MN3a) mammals in the open brown coal pit Merkur, North Bohemia, Czech Republic. Deinsea 10:163–182.
- Fischer de Waldheim, G. 1817. Adversaria zoologica. Mémoires de la Société Impériale des Naturalistes de Moscou 5:357–428.
- Fortelius, M. (coordinator). 2017. New and Old Worlds Database of Fossil Mammals (NOW). University of Helsinki. Available at www.helsinki.fi/science/now. Accessed December 15, 2017.
- Freudenberg, H. 1941. Die oberoligocänen Nager von Gaimersheim bei Ingolstadt und ihre Verwandten. Palaeontographica, Abteilung A 92:99–164.
- Gazin, C. L. 1930. A Tertiary vertebrate fauna from the upper Cuyama drainage basin, California. Carnegie Institution of Washington Publication 404:55–76.
- Gazin, C. L. 1932. A Miocene mammalian fauna from southeastern Oregon. Carnegie Institution of Washington Publication 418:37–86.
- Hilgen, F. J., L. J. Lourens, and J. A. van Dam. 2012. The Neogene Period; pp. 923–978 in F. M. Gradstein, J. G. Ogg, M. Schmitz, and G. Ogg (eds.), The Geologic Time Scale 2012. Elsevier, Dordrecht, The Netherlands.
- Hofmann, A. 1893. Die Fauna von Göriach. Abhandlungen der Kaiserlich-königlichen geologischen Reichsanstalt 15:1–87.
- Howell, A. H. 1929. Revision of the American chipmunks (genera Tamias and Eutamias). United States Department of Agriculture. Bureau of Biological Survey. North American Fauna 52:1–157. doi: 10.3996/nafa.52.0001
- Illiger, C. 1811. Prodromus Systematis Mammalium et Avium. C. Salfeld, Berlin, 302 pp.
- Jaeger, J. J. 1977. Les rongeurs du Miocène moyen et supérieur du Maghreb. Palaeovertebrata 8:1–166.
- Joniak, P., and H. de Bruijn. 2015. Rodents from the Upper Miocene Tuğlu Formation (Çankırı Basin, Central Anatolia, Turkey). Paläontologische Zeitschrift 89:1039–1056. doi: 10.1007/s12542-015-0257-5
- Joniak, P., P. Peláez-Campomanes, L. W. van den Hoek Ostende, and B. Rojay. 2017. Early Miocene rodents of Gökler (Kazan Basin, Central Anatolia, Turkey). Historical Biology. doi: 10.1080/08912963.2017.1414211.
- Kordikova, E. G., E. P. J. Heizmann, and H. de Bruijn. 2004. Early–Middle Miocene vertebrate faunas from Western Kazakhstan. Part 1. Rodentia, Insectivora, Chiroptera, and Lagomorpha. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 231:219–276. doi: 10.1127/njgpa/231/2004/219
- Kretzoi, M. 1959. Insectivoren, Nagetiere und Lagomorphen der jüngstpliozänen Fauna von Csarnóta im Villányer Gebirge (Südungarn). Vertebrata Hungarica 1:237–244.
- Kretzoi, M. 1962. Fauna und Faunenhorizont von Csarnóta. A Magyar Állami Földtani Intézet Évi Jelentése 1959. Annual Report of the Hungarian Geological Institute of 1959:344–395.
- Krijgsman, W., C. E. Duermeijer, C. G. Langereis, H. de Bruijn, G. Saraç, and P. A. M. Andriessen. 1996. Magnetic polarity stratigraphy of late Oligocene to middle Miocene mammal-bearing continental deposits in Central Anatolia (Turkey). Newsletters on Stratigraphy 34:13–29. doi: 10.1127/nos/34/1996/13
- Lartet, É. A. I. H. 1851. Notice sur la colline de Sansan. Annuaire du Département du Gers, Auch, France 1851:1–47.
- Lavocat, R. 1961. Le gisement de vertébrés miocènes de Beni Mellal (Maroc). Étude systématique de la faune de mammifères, et conclusions générales. Notes et Mémoires du service géologique du Maroc 155:29–144.
- Linnaeus, C. 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Laurentii Salvii, Stockholm, 824 pp.
- Major, C. I. F. 1893. On some Miocene squirrels, with remarks on the dentition and classification of the Sciurinae. Proceedings of the Zoological Society of London 1893:179–215.
- Major, C. I. F. 1899. Some rodents from the Middle Miocene lacustrine deposits of Oeningen, Southern Germany. Geological Magazine, new series, Decade IV 6:363–373. doi: 10.1017/S0016756800142414
- Marković, Z., H. de Bruijn, and W. Wessels. 2016. A revision of the new rodent collections from the Early Miocene of Sibnica, Serbia; pp. 63–117 in Z. Marković and M. Milivojević (eds.), Life on the Shore: Geological and Paleontological Research in the Neogene of Sibnica and Vicinity (Levač basin, Central Serbia). Part 1. Special Issue of the Natural History Museum in Belgrade. Natural History Museum, Belgrade, Serbia.
- Matthew, W. D., and C. C. Mook. 1933. New fossil mammals from the Deep River beds of Montana. American Museum Novitates 601:1–7.
- Mein, P. 1958. Les mammifères de la faune sidérolithique de Vieux-Collonges. Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon 5:1–122.
- Mein, P. 1970. Les sciuroptères (Mammalia, Rodentia) néogènes d’Europe occidentale. Geobios 3:7–77. doi: 10.1016/S0016-6995(70)80001-8
- Mein, P., and L. Ginsburg. 1997. Les mammifères du gisement miocène inférieur de Li Mae Long, Thaïlande: systématique, biostratigraphie et paléoenvironnement. Geodiversitas 19:783–844.
- Mein, P., and L. Ginsburg. 2002. Sur l’âge relatif des différents dépôts karstiques Miocènes de La Grive-Saint-Alban (Isère). Cahiers scientifiques Muséum d’Histoire naturelle, Lyon, fasc. 2/2002:7–47.
- Munthe, J. 1980. Rodents of the Miocene Daud Khel local fauna, Mianwali district, Pakistan. Part 1. Sciuridae, Gliridae, Ctenodactylidae, and Rhizomyidae. Milwaukee Public Museum, Contributions in Biology and Geology 34:1–36.
- Murray, A. 1866. The Geographical Distribution of Mammals. Day and Son, London , 419 pp.
- Nicoara, I. 2011. Upper Turolian Sciuroidea (Rodentia, Mammalia) from the Republic of Moldova. Acta Palaeontologica Romaniae 7:257–265.
- Nowak, R. M. 1999. Walker’s Mammals of the World, Volumes 1 and 2, sixth edition. Johns Hopkins University Press, Baltimore, Maryland, 1936 pp.
- Peláez-Campomanes, P. 2001. Revision of the Aragonian (Miocene) Atlantoxerus (Sciuridae). Journal of Paleontology 75:418–426. doi: 10.1017/S0022336000018217
- Peláez-Campomanes, P., and R. Daams. 2002. Middle Miocene rodents from Paşalar, Anatolia, Turkey. Acta Palaeontologica Polonica 47:125–132.
- Pomel, A. 1853. Catalogue méthodique et descriptif des vertébrés fossiles découverts dans le bassin hydrographique supérieur de la Loire, et surtout dans la vallée de son affluent principal, l’Allier. J.-B. Baillière, Paris, France, 193 pp.
- Qiu, Z. 1991. The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 8. Sciuridae (Rodentia). Senckenbergiana lethaea 71:223–255.
- Qiu, Z. 1996. Middle Miocene Micromammalian Fauna from Tunggur, Nei Mongol. Science Press, Beijing, China, 216 pp. [Chinese 1–165, 214–216; English 166–213]
- Qiu, Z., and Y. Liu. 1986. The Aragonian vertebrate fauna of Xiacaowan, Jiangsu. 5. Sciuridae (Rodentia, Mammalia). Vertebrata PalAsiatica 24:195–209. [Chinese 195–205; English 206–209]
- Qiu, Z.-D. 2002. Sciurids from the late Miocene Lufeng hominoid locality, Yunnan. Vertebrata PalAsiatica 40:177–193. [Chinese 177–179; English 179–193]
- Qiu, Z.-D., S.-H. Zheng, and Z.-Q. Zhang. 2008. Sciurids and zapodids from the Late Miocene Bahe Formation, Lantian, Shaanxi. Vertebrata PalAsiatica 46:111–123.
- Reumer, J. W. F., and L. W. van den Hoek Ostende. 2003. Petauristidae and Sciuridae (Mammalia, Rodentia) from Tegelen, Zuurland, and the Maasvlakte (The Netherlands). Deinsea 10:455–467.
- Schlosser, M. 1884. Die Nager des europäischen Tertiärs, nebst Betrachtungen über die Organisation und die geschichtliche Entwicklung der Nager überhaupt. Palaeontographica, Abteilung A 31:9–162.
- Sickenberg, O., J. D. Becker-Platen, L. Benda, D. Berg, B. Engesser, W. Gaziry, K. Heissig, K. A. Hünermann, P. Y. Sondaar, N. Schmidt-Kittler, K. Staesche, U. Staesche, P. Steffens, and H. Tobien. 1975. Die Gliederung des höheren Jungtertiärs und Altquartärs in der Türkei nach Vertebraten und ihre Bedeutung für die internationale Neogen-Stratigraphie. Geologisches Jahrbuch Reihe B 15:1–167.
- Stehlin, H. G., and S. Schaub. 1951. Die Trigonodontie der simplicidentaten Nager. Schweizerische Palaeontologische Abhandlungen 67:1–385.
- Stephan-Hartl, R. 1972. Die altmiozäne Säugetierfauna des Nordbassin und der Niederräder Schleusenkammer (Frankfurt/M., Hessen) und ihre stratigraphische Stellung. Abhandlungen des Hessischen Landesambtes für Bodenforschung 64:1–97.
- Sulimski, A. 1964. Pliocene Lagomorpha and Rodentia from Weze 1 (Poland): study on the Tertiary bone breccia fauna from Weze near Dzialoszyn in Poland. Part XIX. Acta Palaeontologica Polonica 9:149–261.
- Theocharopoulos, C. D. 2000. Late Oligocene-Middle Miocene Democricetodon, Spanocricetodon and Karydomys n. gen. from the eastern Mediterranean area. Gaia 8:1–92.
- Thomas, O. 1908. The genera and subgenera of the Sciuropterus group, with descriptions of three new species. Annals and Magazine of Natural History Series 8 1:1–8.
- Thorington, R. W., Jr., C. E. Schennum, L. A. Pappas, and D. Pitassy. 2005. The difficulties of identifying flying squirrels (Sciuridae: Pteromyini) in the fossil record. Journal of Vertebrate Paleontology 25:950–961. doi: 10.1671/0272-4634(2005)025[0950:TDOIFS]2.0.CO;2
- Trouessart, E.-L. 1880. Catalogue des mammifères vivants et fossiles. Bulletin de la Société d’Études Scientifiques d’Angers 10:86. Angers, France.
- Ünay, E. 1981. Middle and Upper Miocene rodents from the Bayraktepe section (Çanakkale, Turkey). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 84:217–238.
- Ünay, E., E. Atabey, and G. Saraç. 2001. Small mammals and Foraminifera from the Anatolian (Central Taurus) Early Miocene. Annals of Carnegie Museum 70:247–256.
- Ünay, E., H. de Bruijn, and G. Saraç. 2003. A preliminary zonation of the continental Neogene of Anatolia based on rodents. Deinsea 10:539–547.
- Vasileiadou, K., and G. D. Koufos. 2005. The micromammals from the Early/Middle Miocene locality of Antonios, Chalkidiki, Greece. Annales de Paléontologie 91:197–225. doi: 10.1016/j.annpal.2005.06.002
- Vasileiadou, K., and I. A. Sylvestrou. 2009. The Late Miocene mammal faunas of the Mytilinii Basin, Samos Island, Greece: new collection. 4. Micromammals. Beiträge zur Paläontologie, Wien 31:37–55.
- Vasileiadou, K. V., G. D. Koufos, and G. E. Syrides. 2003. Silata, a new locality with micromammals from the Miocene/Pliocene boundary of the Chalkidiki peninsula, Macedonia, Greece. Deinsea 10:549–562.
- Vianey-Liaud, M. 1974. Palaeosciurus goti nov. sp., écureuil terrestre de l’Oligocène moyen du Quercy. Données nouvelles sur l’apparition des sciuridés en Europe. Annales de Paléontologie, Vertébrés 60:103–122.
- Villalta, J. F. 1950. Sobre un esciuroptéro del Vindoboniense del Vallés-Penedés. Boletín de la Real Sociedad Española de Historia Natural, Sección Geológica 48:53–59.
- Von Meyer, M. 1848. Mittheilungen an Professor Bronn gerichtet. Frankfurt am Main, 21. April 1848. Neues Jahrbuch für Mineralogie, Geognosie, Geologie, und Petrefaktenkunde 1848:465–473.
- Wei, Y.-P. 2010. Atlantoxerus from the middle Miocene of northern Junggar Basin and their environmental significance. Vertebrata PalAsiatica 48:220–234. [ Chinese 220–231; English 231–234]
- Werner, J. 1994. Beiträge zur Biostratigraphie der Unteren Süsswasser-Molasse Süddeutschlands. Rodentia und Lagomorpha (Mammalia) aus den Fundstellen der Ulmer Gegend. Stuttgarter Beiträge zur Naturkunde Serie B 200:1–263.
- Wilson, D. E., and D. M. Reeder. 2005. Mammal Species of the World: A Taxonomic and Geographic Reference, Volumes 1 and 2, third edition. Johns Hopkins University Press, Baltimore, Maryland, 2142 pp.
- Wu, W. 1988. The first discovery of middle Miocene rodents from the Northern Junggar Basin, China. Vertebrata PalAsiatica 26:250–264. [ Chinese 250–259; English 259–264]
- Ziegler, R., and V. Fahlbusch. 1986. Kleinsäuger-Faunen aus der basalen Oberen Süsswasser-Molasse Niederbayerns. Zitteliana, Abhandlungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 14:3–80.