
ABSTRACT
We describe an assemblage of isolated hybodontiform shark teeth from the nonmarine Upper Triassic (Carnian) Momonoki Formation in the town of Ominecho in Mine, Yamaguchi, Japan. These teeth were first reported in 1996 and tentatively referred to the genus Hybodus, but their taxonomy was not rigorously evaluated in an apomorphy-based context, and they have since been mostly overlooked. Herein, we assign them to a new species, Parvodus ominechonensis, based on a combination of synapomorphies of the lonchidiid genus Parvodus and autapomorphic character states. A diversity of Triassic hybodontiforms is known from Japan, but of these, Parvodus ominechonensis sp. nov. is the only taxon known from nonmarine deposits and the first reported occurrence of Lonchidiidae. Parvodus ominechonensis fills a gap in the stratigraphic record of Parvodus between occurrences in Middle Triassic (Anisian) and Middle Jurassic (Bajocian) strata. The global Triassic record of Parvodus comprises Parvodus huizodus from the Olenekian of China, Parvodus sp. from the Anisian of China, and Parvodus ominechonensis from the Carnian of Japan, of which Parvodus ominechonensis and Parvodus huizodus are known from nonmarine deposits. After the Triassic, Parvodus is known from marine and nonmarine strata across Laurasia and in South America until its extinction during the Early Cretaceous. This suggests that Parvodus may have originated in freshwater habitats in the South China region of Pangea following the end-Permian mass extinction and diversified throughout the Triassic in present day East Asia before attaining a global distribution later in the Mesozoic.
http://zoobank.org/urn:lsid:zoobank.org:pub:42B4D1C1-F4BE-4296-896B-01B048BDE25F
INTRODUCTION
Hybodontiformes is an extinct clade of sharks generally considered to be the sister clade to Neoselachii (i.e., skates, rays, and modern sharks) within the chondrichthyan clade Euselachia (Compagno, Citation1977). The earliest known unambiguous hybodontiform body fossils are from the Mississippian (Dick, Citation1978), but isolated teeth extend the earliest records of the group to the Late Devonian (Ginter et al., Citation2002). Hybodontiform fossils are known primarily from coastal and lagoonal strata throughout the late Paleozoic, suggesting that the ancestral paleoecology of the clade was shallow marine; however, some hybodontiforms apparently became euryhaline early in the evolution of the clade, and from the Mississippian onward, hybodontiforms repeatedly invaded freshwater environments (Fischer, Citation2007; Maisey, Citation1989). Hybodontiforms were the most abundant sharks in both marine and nonmarine vertebrate assemblages throughout the early Mesozoic, dwindling in diversity after the Early Jurassic until the extinction of the clade at or near the end of the Cretaceous (Kriwet & Benton, Citation2004; Stumpf et al., Citation2021).
In Japan, the oldest known hybodontiform sharks are isolated teeth from the marine Olenekian (Lower Triassic) Osawa and Taho formations in Miyagi (Kato et al., Citation1995) and Ehime (Goto et al., Citation2010) prefectures, respectively. Isolated hybodontiform teeth remain relatively common in Japanese vertebrate fossil assemblages throughout the rest of the Mesozoic (Goto, Citation1972, Citation1994a, Citation1994b; Goto et al., Citation1991, Citation1996, Citation2010; Kato et al., Citation1995; Kitaura et al., Citation1974; Yabumoto & Uyeno, Citation1994; Yamagishi, Citation2004), with nearly all occurrences reported from marine strata. To date, the only nonmarine hybodontiform materials reported from Japan are two isolated teeth from the freshwater fluvial deposits of the Upper Triassic Momonoki Formation (Mine Group) in Mine, Yamaguchi in southwestern Honshu, the largest of the islands of the Japanese archipelago. These teeth were included in an inventory of fossil fish from the Mine Group in 1996 and tentatively referred to the genus Hybodus (Uyeno et al., Citation1996); however, the anatomy of these specimens was not described, and they have remained overlooked since then. Herein, we describe these and two additional hybodontiform teeth from the Momonoki Formation, evaluate their taxonomy and anatomy in an apomorphy-based context for the first time, and discuss their implications for hybodontiform paleobiogeography.
MATERIAL AND METHODS
Geological Setting
The Upper Triassic Mine Group is a sequence of fossiliferous nonmarine and shallow marine strata exposed between the cities of Mine and Sanyo-Onoda (San’yō-Onoda) in western Yamaguchi Prefecture, Japan (; hereafter, simply ‘Yamaguchi’). These strata are characterized by rich paleobotanical (Imada et al., Citation2022; Maeda & Oyama, Citation2019; Naito, Citation2000; Oishi, Citation1932; Yokoyama, Citation1890, Citation1891, Citation1905) and paleoentomological records (Aizawa, Citation1991; Fujiyama, Citation1973, Citation1991; Khramov et al., Citation2023; Oyama & Maeda, Citation2020; Oyama et al., Citation2023; Oyama et al., Citation2020), but mollusc (Amano, Citation1955; Tokuyama, Citation1958, Citation1959a, Citation1959b, Citation1960a, Citation1960b, Citation1960c) and rare vertebrate fossils are also preserved. In addition to hybodontiform shark teeth (Uyeno et al., Citation1996), the vertebrate fossil assemblage of the Mine Group comprises actinopterygian fish (Uyeno et al., Citation1996, Citation2003), isolated reptile teeth (Hasegawa & Ota, Citation1975), and the partial skull of a dicynodont synapsid (Jinnouchi et al., Citation2018).
FIGURE 1. Geographic positions of Yamaguchi Prefecture and Ominecho within Japan and satellite image of Ominecho with relevant localities labeled.
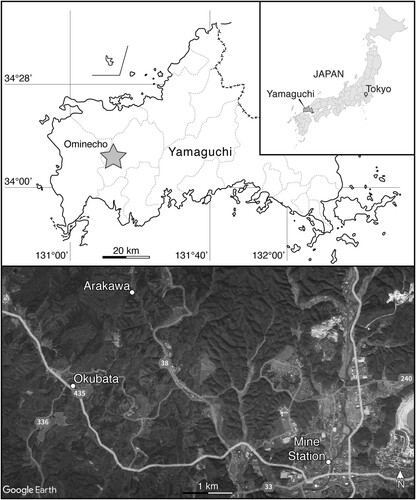
The hybodontiform teeth described herein are from the town of Omine-chō (hereafter romanized Ominecho; sometimes alternatively romanized Ominé (Fujiyama, Citation1991; Oyama et al., Citation2023) or Omine (Imada et al., Citation2022)) in western Mine, Yamaguchi (). In this region, the Mine Group comprises the shallow marine Hirabara Formation, nonmarine Momonoki Formation, and marine Aso Formation (). The Mine Group was deposited along the active continental margin of the South China Block (Kametaka, Citation1999; Taira, Citation2001; Wakita et al., Citation2021), which is reflected by the presence of volcanic rock fragments and detrital zircons in all three of its formations (Kametaka, Citation1999). Volcanic lithic fragments are more abundant in the Momonoki Formation than the underlying Hirabara Formation (Kametaka, Citation1999), which helps discern between the two in isolated outcrops, as both formations are characterized by superficially similar lithologies.
FIGURE 2. A, geological map of the Mine Group in Ominecho. Redrawn and modified from Kimura et al. (Citation1983). B, historical photograph of the outcrop exposed at the Okubata locality between 1987 and 1990 before it was subsequently covered, date unknown. Courtesy of the Mine City Museum of History and Folklore. C, approximate stratigraphic position of hybodontiform teeth at Okubata locality. Stratigraphy of the Momonoki Formation measured near ground-level at outcrop pictured in B. Redrawn and modified from Takahashi et al. (Citation1992). D, historical photograph of the portal to the abandoned Arakawa coal mine, date unknown. Courtesy of the Mine City Museum of History and Folklore. E, approximate stratigraphic position of hybodontiform tooth at Arakawa locality relative to major coal seams within the Momonoki Formation. Geological cross section of the Arakawa coal mine redrawn and modified from Omine Coalfield Memoirs Editorial Committee (Citation2000).
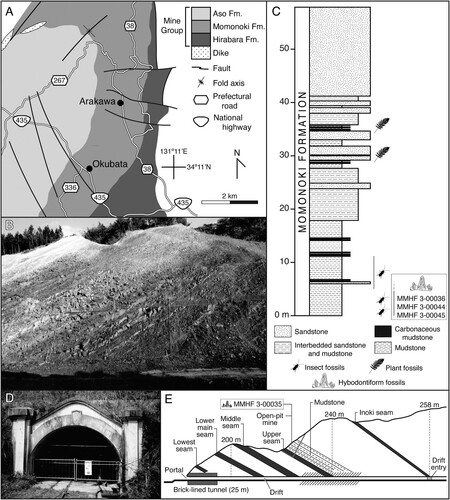
The hybodontiform teeth were collected from the Momonoki Formation, which was initially constrained to the Carnian (Late Triassic) based on bivalve biostratigraphy (Amano, Citation1955; Hase, Citation1951). More recently, U-Pb LA-ICP-MS ages of detrital zircons supporting the Late Triassic age of the formation were reported (Wakita et al., Citation2021), with some grains possibly as young as the Rhaetian; however, the uncertainty of those ages was relatively high, and additional work is needed to date the Mine Group strata more precisely, and we tentatively follow the biostratigraphic correlation to the Carnian. The Momonoki Formation is approximately 1000 m at its thickest and is composed of fluvial facies deposited in a meandering river system (Yukawa et al., Citation2014). It is characterized by point bar, floodplain, overbank, and crevasse splay deposits comprising conglomerates, sandstones, mudstones, and anthracite coal seams along with abundant nonmarine fossils, including in situ tree stumps (Amano, Citation1955; Maeda & Oyama, Citation2019; Yukawa et al., Citation2014). Fossils are most abundant from the mudstones (Maeda & Oyama, Citation2019; Naito, Citation2000; Uyeno et al., Citation1996), and the hybodontiform teeth described herein are from an interval of fossiliferous nonmarine mudstone in the upper part of the formation that has also yielded abundant insect and plant fossils (Fujiyama, Citation1991; Imada et al., Citation2022; Khramov et al., Citation2023; Naito, Citation2000; Oyama & Maeda, Citation2020; Takahashi et al., Citation1992; Ueda, Citation1991; Uyeno et al., Citation1996).
The hybodontiform teeth were collected at two localities within Ominecho. The first of these is in Okubata (alternatively romanized Okuhata [Ueda, Citation1991]; , ), where a broad outcrop of the upper Momonoki Formation was temporarily exposed along the northeast side of Japan National Highway 435 by extensive road construction between October 1987 and February 1990. It was anticipated that this construction would expose the Momonoki Formation and potentially yield new fossil materials, so geological and paleontological surveys were conducted throughout that time by MMHF and the Mine Natural History Association (Mine Shizenshi Tomo no Kai), with the recovery of nearly 7000 fossil specimens (primarily insects and plants) occurring in two phases. Smaller specimens were collected during the first phase from October 1987 to March 1988, whereas the second phase from August 1989 to February 1990 focused mainly on larger and oversized specimens (Takahashi et al., Citation1992). Three hybodontiform teeth (MMHF 3-00036, MMHF 3-00044, MMHF 3-00045) were collected by Takumitsu Utamura during that time, of which only MMHF 3-00036 was reported and figured by Uyeno et al. (Citation1996). The outcrop was subsequently covered upon completion of the construction and is no longer accessible; however, photographs of the outcrop were taken (), and multiple stratigraphic columns were measured while it was exposed (Aizawa, Citation1991; Takahashi et al., Citation1992). The Momonoki Formation at Okubata strikes at 031° and dips at 29° to the west such that the lowest strata were exposed to the southeast side of the outcrop, and it is from these lowest beds that the fossil teeth were collected ().
An additional hybodontiform tooth (MMHF 3-00035) was collected at the site of a now abandoned anthracite coal mine in Arakawa, which lies approximately 2 kilometers north-northeast of Okubata in Ominecho (, ). The mine is one of many within the Omine Coalfield (Omine Tanden), the largest anthracite-producing region in Japan. The Omine Coalfield was discovered during the 1870s, and commercial coal mining in Ominecho began in earnest in 1897 (Kojima, Citation2022a). The coalfield was acquired by the Navy Ministry of the Empire of Japan in 1904 to produce anthracite for use as fuel for Navy vessels, and the Arakawa mine was developed that same year (Omine Coalfield Memoirs Editorial Committee, Citation2000). The Arakawa mine was primarily a drift mine, with a 25-meter brick-lined tunnel connecting the portal to the main entry drift (). The Navy Ministry subsequently sold the mine into the private sector in 1923 in response to the rapid shift from coal to oil as a fuel for warships in the wake of the First World War, and it changed hands several times before being closed in 1970 along with the rest of the Omine Coalfield (Kojima, Citation2022a, Citation2022b; Omine Coalfield Memoirs Editorial Committee, Citation2000). The Arakawa mine was reopened in 1980 by Kibe Mining Co., Ltd (Kibe Kōgyō Kabushiki Kaisha), who renovated the entrance and temporarily resumed coal production until operations were ceased again in 1991. The hybodontiform tooth was collected in 1981 by one of the authors (HT) from an open pit mine on the surface above the drift mine approximately 100 m south-southwest of the portal. No detailed stratigraphic data were taken at the time of collection, but the locality was correlated to the same approximate range of the upper Momonoki Formation that was temporarily exposed at the Okubata locality based on their similar assemblages of nonmarine plant and insect fossils (Uyeno et al., Citation1996) and the open pit mine’s stratigraphic position relative to major coal seams within the Momonoki Formation (). Although there were once intentions to preserve and reopen the first 100 meters of the entry drift of the Arakawa mine as a tourist attraction, those plans were suspended as of 2000, and the mine remains abandoned (Omine Coalfield Memoirs Editorial Committee, Citation2000). The Arakawa mine is now a restricted area (Kojima, Citation2022a), and the current condition of the outcrop at the open pit mine is uncertain.
Imaging and Measurements
All images were acquired at the National Museum of Nature and Science (NMNS) in Tsukuba, Ibaraki, Japan. Digital photographs were taken using a Leica DVM6 digital microscope. Scanning electron microscope (SEM) images of all teeth were acquired using a Jeol JSM-6510 at an acceleration voltage of 3 kV. Each tooth was scanned using an inspeXio SMX-225CT FPD HR microfocus X-ray CT system (Shimadzu Corporation, Kyoto, Japan) at a voltage of 225 kV and 70 uA. CT scan data were processed, segmented, and rendered using VGSTUDIO MAX 3.2 (Volume Graphics Company). A surface mesh of each tooth in PLY (.ply) format is provided in the Supplementary Online Materials. Linear measurements were made using ImageJ 1.54f.
Romanization of Japanese Names and Terms
Multiple systems for romanizing the Japanese language exist, and there is no universal standard, so many Japanese words and names used herein have not been consistently romanized by different authors in the past. To ensure consistency throughout the text of this study, we generally follow the style guidance of the third edition of the Japan Style Sheet (Society of Writers Editors and Translators, Citation2018) and the Monumenta Nipponica Style Sheet (Sophia University, Citation2018), and Japanese names and terms were transcribed following the modified Hepburn system of romanization rather than simply repeating previous transliterations from the literature; however, alternative transliterations are provided when appropriate for clarity. The specific style decisions made are summarized in the Supplemental Materials.
Institutional Abbreviations—KPM, Kanagawa Prefectural Museum of Natural History (Kanagawa Kenritsu Seimei no Hoshi Chikyū Hakubutsukan), Odawara, Kanagawa, Japan; KUJC, Department of Geology and Mineralogy, Graduate School of Science, Kyoto University, Kyoto, Japan; MMHF, Mine City Museum of History and Folklore (Mine-shi Rekishi Minzoku Shiryōkan), Mine, Yamaguchi, Japan; TMNH, Toyohashi Museum of Natural History (Toyohashi-shi Shizenshi Hakubutsukan), Toyohashi, Aichi, Japan; UMUT, The University Museum, The University of Tokyo (Tōkyō Daigaku Sōgō Kenkyū Hakubutsukan), Bunkyo-ku, Tokyo, Japan.
SYSTEMATIC PALEONTOLOGY
CHONDRICHTHYES Huxley, Citation1880
ELASMOBRANCHII Bonaparte, Citation1838
HYBODONTIFORMES Patterson, Citation1966
LONCHIDIIDAE Herman, Citation1977 sensu Rees, Citation2008
PARVODUS Rees and Underwood, Citation2002
Type Species—Parvodus rugianus (Ansorge, Citation1990), Lower Cretaceous (Berriasian–Valanginian) of Rügen, Germany.
PARVODUS OMINECHONENSIS, sp. nov.
(, )
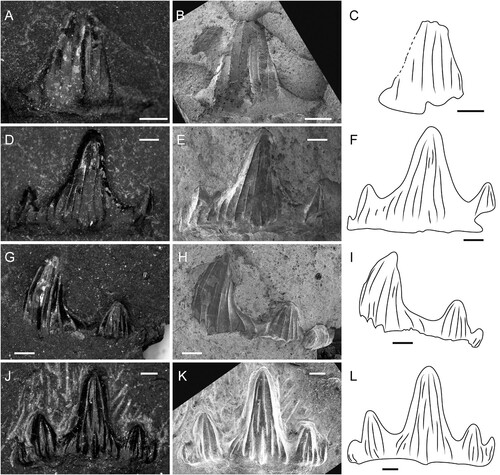
FIGURE 3. Parvodus ominechonensis, DVM, A, D, G, J, and SEM, B, E, H, K, images and interpretative line drawings, C, F, I, L of teeth. A–C, MMHF 3-00044 (holotype) in labial view. D–F, MMHF 3-00036 (paratype) in labial view. G–I, MMHF 3-00045 (paratype) in labial view. J–L, MMHF 3-00035 (paratype) in labial view. Scale bars equal 0.5 mm.
Hybodus Uyeno et al., Citation1996:pl. 11
Etymology—The specific epithet acknowledges the town of Ominecho (pronounced oh-mee-neh-cho), the geographic provenance of the holotype.
Holotype Specimen—MMHF 3-00044, an isolated incomplete anterior tooth crown missing its root.
Holotype Locality—Okubata, Ominecho, Mine, Yamaguchi, Japan.
Paratypes Specimens—MMHF 3-00035, MMHF 3-00036, MMHF 3-00045 (all isolated incomplete anterior tooth crowns).
Paratype Localities—Okubata (MMHF 3-00036, MMHF 3-00045) and Arakawa (MMHF 3-00035), Ominecho, Mine, Yamaguchi, Japan.
Horizon and Age—Upper strata of the Momonoki Formation, Mine Group. Late Triassic, Carnian (Hase, Citation1951; Wakita et al., Citation2021).
Diagnosis—Species of Parvodus distinguished by the following combination of character states (synapomorphies of Parvodus denoted by a cross; autapomorphic character states denoted by an asterisk): gracile crown with moderately high cusp and cusplets in anterior teeth†; moderately blunt main cusp; cutting dentition with strong occlusal crest; labial protuberance present; labial protuberance moderate and rounded†; ornamentation with few nearly parallel vertical folds†; some labial vertical folds bifurcate from the apex to the base; anterior teeth with prominent central vertical fold extending from the apex nearly to the base on the lingual surface of main cusp giving it a subtriangular cross-section*; low crown shoulder†. For a detailed differential diagnosis and further discussion of taxonomy, see Comments.
Description—All specimens of Parvodus ominechonensis are incomplete anterior tooth crowns missing their roots. Each remains embedded in matrix with only its labial surface partially exposed () and is best visualized via CT scan data (). The holotype (MMHF 3-00044; , ) is less complete but better preserved than all paratype specimens, and it best typifies the unique combination of autapomorphic and synapomorphic character states diagnostic of the new species. The crown fragment measures 2.3 mm apicobasally, 3.0 mm mesiodistally, and 0.8 mm labiolingually. The main cusp of the crown is moderately tall and moderately slender and blunt, and there is one complete well-developed lateral cusplet on the distal side of the main cusp and the base of one partial lateral cusplet on the mesial side. The main cusp is slightly recurved distally and two times the height of the complete lateral cusplet. In occlusal view, the crown is slightly concave lingually. The occlusal crest is prominent and runs along the entire occlusal edge of the crown (), and the teeth are characterized by cutting-crushing (as opposed to crushing or grasping-crushing) dentition. The surfaces of the main cusp and lateral cusplet are ornamented with few vertical folds. The folds on the labial surface are relatively weak, and a small, rounded labial protuberance lies at the base of the most prominent fold (). Some of the vertical folds on the labial surface bifurcate about halfway between the tip and the base such that not every fold extends from the base to the tip of the crown. Five stronger non-bifurcating vertical folds mark the lingual surface of the main cusp (), the middle of which is particularly prominent and gives the cusp a subtriangular cross section with the apex pointing lingually and the other two points formed by the occlusal crest (). There are four vertical folds on the labial surface of the cusplet and three on the lingual surface. The surface of the crown is relatively smooth between the vertical folds, and there are no striae at the base of the crown. The region of the crown between the cusp and cusplets and the root (i.e., the crown shoulder) is low, and it appears that the root would have been lingually inclined based on the broken surface of the base of the crown.
FIGURE 4. Parvodus ominechonensis, CT volume renderings of teeth with matrix digitally removed. A–E, holotype tooth (MMHF 3-00044) in labial (A), lingual (B), basal (C), and apical (D) views and cutaway transverse section of the main cusp in apical view (E). The dashed line in B indicates the plane of the transverse section shown in E. F–G, paratype tooth (MMHF 3-00036) in labial (F) and lingual (G) views. H–I, paratype tooth (MMHF 3-00045) in labial (H) and lingual (I) views. J–K, paratype tooth (MMHF 3-00035) in labial (J) and lingual (K) views. Abbreviations: dcl, distal cusplet; lab.pr, labial protuberance; mc, main cusp; mcl, mesial cusplet; oc, occlusal crest; plif, prominent lingual fold.
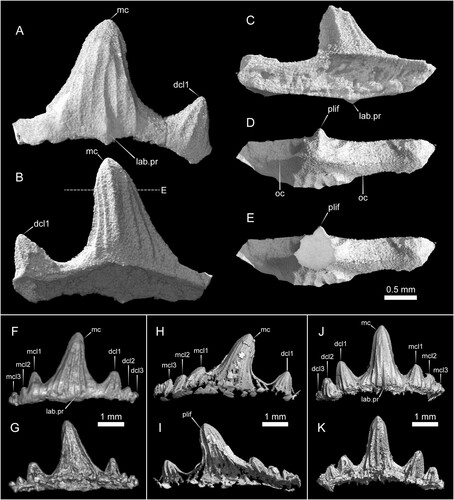
The paratype teeth are generally similar in shape to the holotype with some variation. MMHF 3-00036 (, ) has a tall and slender main cusp, one pair of well-developed cusplets, and two pairs of incipient cusplets that decrease in size both mesially and distally. The main cusp is 2.8 mm tall and is approximately 2.3 times the height of the innermost lateral cusplets. Of the well-developed innermost cusplets, the distal cusplet is more robust than the mesial cusplet. The surfaces of the innermost cusplets are marked by vertical folds like the cusplet of the holotype, whereas the incipient cusplets do not bear any clear vertical folds. A small, rounded labial protuberance is present (), but the lingual surface of the tooth is poorly preserved, and it is unclear whether a prominent vertical fold is present to characterize the cross-sectional shape of the main cusp.
MMHF 3-00045 (, ) has a slender blunt main cusp, one well developed lateral cusplet and traces of the edge of another on the distal side, and at least three incipient lateral cusplets on the mesial side. The main cusp is more distally inclined than any of the other preserved teeth of Parvodus ominechonensis, and the incipient cusplets are similarly distally inclined, reflecting some degree of heterodonty. The well-developed lateral cusplet is blunt, straight, and robust like that of the holotype. The height of the main cusp is 2.8 mm, and it is about twice the height of the innermost lateral cusplet on each side. The surface of the crown is poorly preserved, with only traces of enamel remaining. The main cusp, robust lateral cusplet, and at least the first lateral incipient all bear vertical folds on both the labial and lingual surfaces. The folds are strongest on the lingual surfaces of the main cusp and robust lateral cusplet, and the main cusp is characterized by a prominent lingual fold. The lingual side of the crown is especially poorly preserved; however, the cross-sectional shape of the main cusp appears to be somewhat less triangular than that of the holotype. The base of the crown is incompletely preserved, and it is unclear whether a labial protuberance is present or absent.
MMHF 3-00035 (, ) has a slender main cusp that is taller and somewhat straighter than any other tooth, but its shape is otherwise like the holotype and MMHF 3-00045. The main cusp is 2.5 mm tall and has vertical folds on the labial surface that are generally shorter than those of the holotype, with only two folds on the outer margins of the cusp spanning the distance from the tip to the base of the crown. One fold that begins about halfway up at the base of the crown terminates near the base with a small, rounded labial protuberance (). The lingual surface of the crown is more poorly preserved than the labial surface. The folds on the lingual surface of the main cusp are stronger than those on the labial surface, as in the holotype, but the cusp does not seem to have a triangular cross section. There are two relatively tall well-developed lateral cusplets and one incipient cusplet on the distal side of the main cusp and one shorter well-developed cusplet and at least two incipient cusplets on the mesial side. The two innermost lateral cusplets on each side of the main cusp are marked by vertical folds on both the labial and lingual surfaces. The main cusp is approximately twice the height of the larger innermost cusplet and about 2.5 times the height of the smaller innermost cusplet.
Comments—Although Uyeno et al. (Citation1996) tentatively referred MMHF 3-00036 to Hybodus and MMHF 3-00035 to either Hybodus or Hybodontoidea, depending on how the original Japanese is interpreted, the genus Hybodus is generally considered a problematic wastebasket taxon under which dozens of species have been erected despite a lack of apomorphies shared with the type species Hybodus reticulatus Agassiz, Citation1837 (Maisch & Matzke, Citation2016; Rees, Citation2008). Rees (Citation2008) recommended that only the type species and Hybodus hauffianus Fraas, Citation1895 be considered valid species of Hybodus until a thorough revision of the genus could be conducted, as both species are known not only from isolated teeth but morphologically similar skeletons, and that sentiment has been echoed by subsequent authors (Maisch & Matzke, Citation2016; Stumpf & Kriwet, Citation2019).
An apomorphy-based approach using Rees (Citation2008) and Rees and Underwood (Citation2002) as primary references instead supports a referral of the teeth of Parvodus ominechonensis to the hybodontiform clade Lonchidiidae and the genus Parvodus, respectively. Synapomorphies of Hybodus recognized by Rees (Citation2008) based on Hybodus reticulatus and Hybodus hauffianus are stout cusplets and strong ornamentation, neither of which are present in Parvodus ominechonensis (MMHF 3-00035; MMHF 3-00036; MMHF 3-00044; MMHF 3-00045). Furthermore, the clade of Hybodontinae (Hybodus + Egertonodus) is diagnosed by tall cusps with a nearly circular cross section, in contrast to the moderately high main cusp and lateral cusplets (MMHF 3-00035; MMHF 3-00036; MMHF 3-00044; MMHF 3-00045) and main cusp with a subtriangular cross section (MMHF 3-00044; MMHF 3-00045) in Parvodus ominechonensis. A synapomorphy of the clade Lonchidiidae (comprising at least Lonchidion, Parvodus, Vectiselachos, and Hylaeobatis) recognized by Rees (Citation2008) is the labial protuberance or labial ‘peg,’ which is shared by Parvodus ominechonensis (MMHF 3-00035; MMHF 3-00036; MMHF 3-00044).
Although some apomorphies of the genera within Lonchidiidae were given by Rees (Citation2008), Rees and Underwood (Citation2002) compiled a more detailed list of character states differentiating the genera considered lonchidiids at that time (Lissodus, Lonchidion, Vectiselachos, Hylaeobatis, Parvodus). Of these, Parvodus ominechonensis possesses a combination of character states that is diagnostic of Parvodus, including a cutting-crushing dentition; gracile crown shape; moderate-high central cusp; a strong occlusal crest; a moderate rounded labial protuberance; three pairs of lateral cusplets; a weak crown shoulder; and ornamentation comprising few, strong folds. However, Parvodus ominechonensis is distinguished from other species of Parvodus in the presence of a prominent central vertical fold on the lingual surface of the main cusp of some anterior teeth that extends from the apex nearly to the base and gives the main cusp a subtriangular cross section. The crown also lacks the short horizontal ridge or small crenulae between the main cusp and lateral cusplets that are diagnostic of the Early Triassic species Parvodus huizodus from Yunnan, China (Wen et al., Citation2022). Using an apomorphy-based approach, a referral of the teeth from the Momonoki Formation to a new species of the genus Parvodus is therefore justified.
DISCUSSION
The Triassic Record of Hybodontiformes in Japan
Shark fossils were unknown from the Triassic System in Japan prior to 1975, at which point dermal denticles were reported from the marine Triassic (Ladinian to Carnian) Adoyama Formation in Karasawa, Tochigi (; Goto, Citation1975). In 1991, a single marine shark tooth each was reported from both the Middle Triassic (Anisian) Waruishi Formation, Yakuno Group and the Upper Triassic (Carnian) Heki Formation, Nabae Group in Yakunocho, Kyoto, which were referred to Hybodus sp. (KUJC 91101) and Acrodus sp. (TMNH-00502), respectively (Goto et al., Citation1991). Those specimens represented the first recognized Triassic occurrences of Hybodontiformes in Japan.
FIGURE 5. Geographic distribution of Hybodontiformes from Triassic strata in Japan.
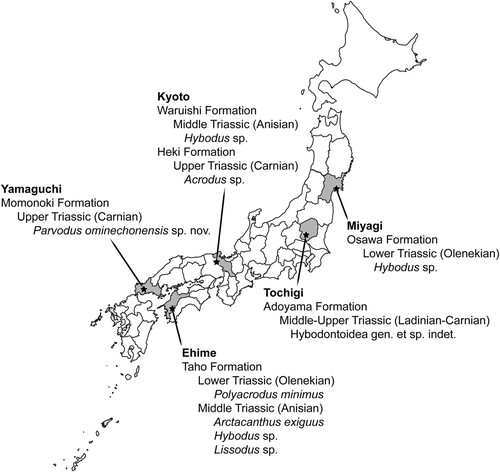
Additional Triassic hybodontiform remains were subsequently reported from both marine and freshwater nonmarine strata across the main Japanese islands of Honshu and Shikoku (). In 1994, a tooth and dermal denticles tentatively referred to Polyacrodus minimus (reposited at Tsurumi University in Yokohama, but no catalog number was provided) were reported from the Lower Triassic (Olenekian) interval of the marine Taho Formation in Shirokawacho, Ehime (Goto, Citation1994a, Citation1994b). In 1994, the elasmobranch material reported from the Adoyama Formation in 1975 was referred to Hybodontidae indet. (Goto, Citation1994a, Citation1994b); however, these were subsequently reclassified more broadly as elasmobranch placoid scales in 1996 (Goto et al., Citation1996), and no justification was given for either classification. In 1995, another tooth referred to the hybodontine hybodontid Hybodus sp. (no repository or catalog number given) was reported from the marine Lower Triassic (Olenekian) Osawa Formation, Inai Group in Utatsu, Minamisanrikucho, Miyagi (Kato et al., Citation1995). That same locality has yielded abundant remains of ammonoids and the ichthyopterygian reptile Utatsusaurus hataii (Kato et al., Citation1995; Motani et al., Citation1998; Shikama et al., Citation1978). Two teeth of the lonchidiid Parvodus ominechonensis (MMHF 3-00035; MMHF 3-00036) from the Upper Triassic (Carnian) Momonoki Formation in Ominecho described herein were first reported and tentatively assigned to Hybodus by Uyeno et al. (Citation1996). The teeth from the Momonoki Formation remain the only nonmarine Triassic hybodontiform teeth known from Japan to date.
Yamagishi (Citation2004) reported an assemblage of hybodontiform teeth and cephalic spines from the Middle Triassic (Anisian) interval of the marine Taho Formation, including the acrodontine hybodontid Acrodus sp. (UMUT MV 28425; UMUT MV 28454; UMUT MV 28457; UMUT MV 28459; UMUT MV 28463; UMUT MV 28467), Hybodus sp. (UMUT MV 28452; UMUT MV 28461; UMUT MV 28465), Polyacrodus sp. (UMUT MV 28449; UMUT MV 28453; UMUT MV 28462; UMUT MV 28466), and the possible hybodontiform Arctacanthus exiguus (UMUT MV 28460-1 [holotype]; UMUT MV 28451; UMUT MV 28460; UMUT MV 28464). That was followed by the report of a tooth referred to the possible lonchidiid Lissodus sp. (KPM-NNV59) from the Olenekian interval of the Taho Formation in Ehime (Goto et al., Citation2010), which is the most recent report of a hybodontiform from Triassic strata in Japan.
Taxonomy and the Global Biostratigraphy of Parvodus
Wen et al. (Citation2022) recently reviewed the fossil record of Parvodus, concluding that the genus comprises at least 11 species ranging from the Lower Triassic to the Lower Cretaceous (see fig. 5 of that study). That study described the new freshwater species Parvodus huizodus Wen et al., Citation2022 from the Lower Triassic (Olenekian) Dongchuan Formation in Yunnan, China, which represents the earliest known occurrence of the genus. Hybodontiform specimens from the Upper Triassic (Carnian) Tiki Formation in Madhya Pradesh, India were initially referred to the genus as the species Parvodus tikiensis (Prasad et al., Citation2008), however, those specimens were subsequently synonymized with specimens of Lissodus duffini from the same formation and together assigned to the new lonchidiid genus Pristrisodus by Bhat et al. (Citation2018). Parvodus ominechonensis therefore fills an Upper Triassic gap in the biostratigraphic record of Parvodus between the next oldest occurrences from the Middle Triassic (Anisian) Yangliujing Formation of China (Chen et al., Citation2007) and the next youngest occurrences from the Middle Jurassic (Bajocian) Xietan Formation of China (Shang et al., Citation2008).
Hybodontiformes is generally considered ancestrally shallow marine, and the clade was again exclusively marine by the Late Cretaceous prior to its extinction, but multiple hybodontiform lineages transitioned to freshwater nonmarine habitats throughout the Mesozoic (Cuny, Citation2012), with freshwater taxa such as Parvodus huizodus and other indeterminate hybodontiforms from the Dongchuan Formation in China (Wen et al., Citation2022); Lissodus tumidoclavus and Polyacrodus sp. from the Burgersdrop Formation in South Africa (Bender & Hancox, Citation2004; Duffin et al., Citation2023); and Lissodus sp. and Hybodus sp. from the Lipovskaya Formation in Russia (Novikov, Citation2010) demonstrating that many freshwater hybodontiforms proliferated in the aftermath of the end-Permian Mass Extinction, and freshwater hybodontiforms remained relatively common across Pangea throughout the Triassic (Benton & Spenser, Citation1995; Heckert, Citation2004; Heckert et al., Citation2007; Heckert & Lucas, Citation2006; Murry, Citation1981; Murry & Kirby, Citation2002; Parker, Citation2005; Peecook et al., Citation2017).
The Triassic record of Parvodus is exclusive to the taxa Parvodus huizodus from the Olenekian (Lower Triassic) of China (Wen et al., Citation2022), Parvodus sp. from the Anisian (Middle Triassic) of China (Chen et al., Citation2007), and Parvodus ominechonensis from the Carnian (Upper Triassic) of Japan. Parvodus ominechonensis is known only from the Momonoki Formation, which is interpreted to have been deposited along the eastern margin of the South China Block (Wakita et al., Citation2021), so the Triassic paleobiogeography of the genus is restricted to the continental South China region of the Pangean landmass, and it may have originated there. Of these three taxa, Parvodus huizodus and Parvodus ominechonensis are known from nonmarine strata, which suggests that the genus Parvodus may have been ancestrally adapted to freshwater habitats.
CONCLUSIONS
Parvodus ominechonensis from the Upper Triassic Momonoki Formation in Mine, Yamaguchi is the first lonchidiid hybodontiform recognized in Japan. The isolated teeth from the Carnian fill a gap in the stratigraphic record of the genus Parvodus between the Middle Triassic and Middle Jurassic. Occurrences of Parvodus are restricted to Mesozoic strata globally, and the genus is known mostly from freshwater nonmarine deposits. The earliest records of Parvodus are from the Triassic of East Asia, including Parvodus huizodus from the Olenekian (Early Triassic) of China, and Parvodus sp. from the Anisian (Middle Triassic) of China, and Parvodus ominechonensis from the Carnian (Late Triassic) of Japan, of which Parvodus huizodus and Parvodus ominechonensis are known from freshwater nonmarine deposits. This suggests that the genus may have originated in the South China region of Pangea and be ancestrally nonmarine.
AUTHOR CONTRIBUTIONS
BTB designed the project, imaged the specimens, rendered the CT data, prepared the figures, and made style decisions for the transcription of all Japanese names and terms into the Latin alphabet for integration into the English language. BTB and NO wrote the manuscript with input from all other authors.
Supplemental Material
Download Zip (3.3 MB)DISCLOSURE STATEMENT
No potential conflict of interest was reported by the author(s).
ACKNOWLEDGMENTS
Thanks to the Mine Public Works Office, Daiwa Construction Co., Inc., and Kibe Mining Co., Inc. for facilitating the paleontological and geological research that produced not only the fossil specimens described herein but countless others; T. Utamura for discovering the fossil teeth at Okubata; S. Nomura, T. Kutsuna, Y. Shigeta, and G. Shinohara for providing access to the X-ray CT scanning facility at NMNS; Y. Kimura and M. Saito-Kato for assisting with DVM and SEM imaging; K. Kawano for providing the vector map of Yamaguchi used in ; A. Marsh for useful comments that greatly improved an early draft of this manuscript; M. Kojima and Y. Yabumoto for useful discussions; and two anonymous reviewers for their constructive comments. This material is based upon work supported by the National Science Foundation under Award No. 2204588 (BTB) and the Japan Society for the Promotion of Science under Postdoctoral Fellowship for Research Abroad No. 202360114 (NO).
DATA AVAILABILITY STATEMENT
The raw data that support the findings of this study are openly available at MorphoSource via https://www.morphosource.org/projects/000612459 and are also reposited in the Mine City Museum of History and Folklore.
SUPPLEMENTARY FILE(S)
mmhf3-00044_holotype.ply: Surface mesh of the holotype tooth of Parvodus ominechonensis (MMHF 3-00044).
mmhf3-00035_paratype.ply: Surface mesh of the paratype tooth of Parvodus ominechonensis (MMHF 3-00035).
mmhf3-00036_paratype.ply: Surface mesh of the paratype tooth of Parvodus ominechonensis (MMHF 3-00036).
mmhf3-00045_paratype.ply: Surface mesh of the paratype tooth of Parvodus ominechonensis (MMHF 3-00045).
Supplementary Online File.docx: Strategies for romanization of Japanese names and terms
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
LITERATURE CITED
- Agassiz, L. (1833–1844). Recherches sur les poissons fossiles. Imprimerie de Petitpierre. (French)
- Aizawa, J. (1991). Fossil insect-bearing strata of the Triassic Mine Group, Yamaguchi Prefecture. Bulletin of Kitakyushu Museum of Natural History, 10, 91–98. (Japanese)
- Amano, M. (1955). Occurrence of a new species of Pleuronectites in the Triassic of Japan. Transactions and proceedings of the Paleontological Society of Japan. New series, 17, 23–28.
- Ansorge, J. (1990). Fischreste (Selachii, Actinopterygii) aus der Wealdentonscholle von Lobber Ort (Mönchgut/Rügen/DDR) [Fossil fish remains (Selachii, Actinopterygii) from an erratic Wealden day deposit of Lobber Ort (Isle of Rügen/DDR)]. Paläontologische Zeitschrift, 64, 133–144. https://doi.org/10.1007/BF02985927 (German)
- Bender, P. A., & Hancox, P. J. (2004). Newly discovered fish faunas from the Early Triassic, Karoo Basin, South Africa, and their correlative implications. Gondwana Research, 7, 185–192. https://doi.org/10.1016/S1342-937X(05)70317-8
- Benton, M. J., & Spenser, P. S. (1995). Fossil reptiles of Great Britain. 386. https://doi.org/10.1007/978-94-011-0519-4
- Bhat, M. S., Ray, S., & Datta, P. M. (2018). A new hybodont shark (Chondrichthyes, Elasmobranchii) from the Upper Triassic Tiki Formation of India with remarks on its dental histology and biostratigraphy. Journal of Paleontology, 92, 221–239. https://doi.org/10.1017/jpa.2017.63
- Bonaparte, C. L. (1838). Synopsis vertebratorum systematis. Nuovi Annali delle Scienze Naturali, 2, 105–133. (Latin)
- Chen, L., Cuny, G., & Wang, X. (2007). The chondrichthyan fauna from the Middle–Late Triassic of Guanling (Guizhou province, SW China). Historical Biology, 19, 291–300. https://doi.org/10.1080/08912960701248234
- Compagno, L. J. V. (1977). Phyletic Relationships of Living Sharks and Rays. American Zoologist, 17(2), 303–322. http://www.jstor.org/stable/3882097
- Cuny, G. (2012). Freshwater hybodont sharks in Early Cretaceous ecosystems: A review. Bernissart dinosaurs and Early Cretaceous terrestrial ecosystems, 519–529. https://www.jstor.org/stable/j.ctt16gzfhx.34
- Dick, J. R. F. (1978). On the Carboniferous shark Tristychius arcuatus Agassiz from Scotland. Transactions of the Royal Society of Edinburgh, 70(4), 63–108. https://doi.org/10.1017/s0080456800012898
- Duffin, Christopher J., Heckert, Andrew B., & Hancox, P. J. (2023). A new low diversity lacustrine elasmobranch fauna from the Lower Triassic Burgersdorp Formation of South Africa with descriptions of Lissodus tumidoclavus n. sp. (Chondrichthyes: Hybodontoidea). Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 308(2), 151–169. https://doi.org/10.1127/njgpa/2023/1134
- Fischer, J. (2007). Palaeobiogeography of the hybodont shark Lissodus during the Carboniferous and Permian of Central Europe.
- Fraas, E. (1895). Ein Fund von Skelettresten von Hybodus (Hybodus hauffianus E. Fraas). Jahresberichte und Mitteilungen des oberrheinischen geologischen Vereins, 28, 24–26. (German)
- Fujiyama, I. (1973). Mesozoic insect fauna of East Asia Part I. Introduction and Upper Triassic faunas. Bulletin of National Science Museum, 16(2), 331–386.
- Fujiyama, I. (1991). Late Triassic Insects from Mine, Yamaguchi, Japan, Part 1. Odonata. Bulletin of the National Science Museum Series C (Geology & Paleontology), 17(2), 49–56.
- Ginter, M., Hairapetian, V., & Klug, C. (2002). Famennian chondrichthyans from the shelves of North Gondwana. Acta Geologica Polonica, 52, 169–215.
- Goto, M. (1972). Nihon-san no kaseki nankotsugyorui ni tsuite ichi sōkatsu [Fossil Chondrichthyes of Japan]. Chishitsugaku zasshi [Journal of the Geological Society of Japan], 78, 585–600. https://doi.org/10.5575/geosoc.78.585 (Japanese)
- Goto, M. (1975). New find of the Permian and Triassic fish fossils in Japan : On the dermal denticles and teeth of fishes discovered from Karasawa area, southeast Asio Mountains. Earth Science, 29(2), 72–74a.
- Goto, M. (1994a). On the Paleozoic and Mesozoic fish remains from the Japanese islands. Monograph of the Association for the Geological Collaboration, 43, 1–16. (Japanese)
- Goto, M. (1994b). Palaeozoic and early Mesozoic fish faunas of the Japanese Islands. The Island Arc, 3, 247–254. https://doi.org/10.1111/j.1440-1738.1994.tb00114.x
- Goto, M., Kuga, N., & Hachiya, K. (1991). On the hybodont elasmobranch teeth of three genera from the Mesozoic of Japan. The Journal of the Geological Society of Japan, 97, 743–750. https://doi.org/10.5575/geosoc.97.743 (Japanese)
- Goto, M., Tanaka, T., & Utsunomiya, S. (2010). On a tooth remain of Lissodus (Elasmobranchii) from the Taho Formation (Lower Triassic) in Seiyo City, Ehime Prefecture, southwest Japan. Chikyū Kagaku [Earth Science], 64, 111–116. https://doi.org/10.15080/agcjchikyukagaku.64.3_111 (Japanese)
- Goto, M., Uyeno, T., & Yabumoto, Y. (1996). Summary of Mesozoic elasmobranch remains from Japan. Mesozoic Fishes-Systematics and Paleoecology, 1, 73–82.
- Hase, A. (1951). Yamaguchi-ken no Sanjō-ki [The Triassic of Yamaguchi Prefecture]. Chishitsu Chōsajo Hōkoku Tokubetsugō [Geological Survery of Japan Report Special Number], 72–89. https://doi.org/10.11501/1371747 (Japanese)
- Hasegawa, Y., & Ota, M. (1975). The Triassic reptilian tooth from the Hirabara Formation, Mine Group, Yamaguchi Prefecture, Japan. Bulletin of the Kitakyushu Museum of Natural History, 11, 25–28. (Japanese)
- Heckert, A. B. (2004). Late Triassic microvertebrates from the lower Chinle Group (Otischalkian–Adamanian: Carnian), southwestern U.S.A. New Mexico Museum of Natural History and Science Bulletin, 27, 1–170.
- Heckert, A. B., Ivanov, A., & Lucas, S. G. (2007). Dental morphology of the hybodontoid shark Lonchidion humblei Murry from the Upper Triassic Chinle Group, USA. New Mexico Museum of Natural History and Science Bulletin, 41, 45–48.
- Heckert, A. B., & Lucas, S. G. (2006). Micro-and small vertebrate biostratigraphy and biochronology of the Upper Triassic Chinle Group, southwestern USA. New Mexico Museum of Natural History and Science Bulletin, 37, 94–104.
- Herman, J. (1977). Les Sélaciens des terrains néocrétacés et paléocènes de Belgique et des contrées limitrophes. Eléments d'une biostratigraphie intercontinentale. Mémoires pour servir à l'explication des Cartes géologiques et minières de la Belgique, 15, 1–401.
- Huxley, T. H. (1880). On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Scientific Meetings of the Zoological Society of London, 43, 649–662.
- Imada, Y., Oyama, N., Shinoda, K., Takahashi, H., & Yukawa, H. (2022). Oldest leaf mine trace fossil from East Asia provides insight into ancient nutritional flow in a plant–herbivore interaction. Scientific Reports, 12, 1–12. https://doi.org/10.1038/s41598-022-09262-1
- Jinnouchi, K., Kusuhashi, N., Liu, J., Takahashi, F., Shinoda, K., & Hasegawa, Y. (2018). Yamaguchi-ken no Jōbu Sanjō-kei Mine-sōgun Momonoki-sō kara sanshutsu shita Dikinodon-rui kaseki [Dicynodont fossils from the Upper Triassic Momonoki Formation, Mine Group, Yamaguchi, Japan]. The Palaeontological Society of Japan 167th Regular Meeting, Abstracts with Programs, 42. (Japanese)
- Kametaka, M. (1999). Jōbu Sanjō-kei Mine-sōgun no kōhaichi [Provenance of the Upper Triassic Mine Group, Southwest Japan]. The Journal of the Geological Society of Japan, 105, 651–667. https://doi.org/10.5575/geosoc.105.651 (Japanese)
- Kato, T., Hasegawa, K., & Ishibashi, T. (1995). Nanbu Kitakami sanchi Ōsawa-sō (Kabu Sanjō-kei) yori Hibodusu-zoku Bansairui no hakken [Discovery of Early Triassic hybodontoid shark tooth from the southern Kitakami Massif]. The Journal of the Geological Society of Japan, 101, 466–469. https://doi.org/10.5575/geosoc.101.466 (Japanese)
- Khramov, A. V., Oyama, N., Kenji, S., & Takahashi, H. (2023). Late Triassic lacewings (Insecta: Neuroptera) from Japan. Historical Biology. https://doi.org/10.1080/08912963.2023.2244519
- Kimura, T., Naito, G., & Ohana, T. (1983). Baiera cf. furcata (Lindley and Hutton) BRAUN from the Carnic Momonoki Formation, Japan. Bulletin of the National Science Museum Tokyo, Series C, 9, 91–114.
- Kitaura, T., Yoshida, K., & Ohe, F. (1974). Gifu-ken Mitarai no same no ha (Jura-ki) ni tsuite [On the some small teeth of Jurassic shark of Mitarai (Lower Tedori Group), Gifu Prefecture, Japan]. Kaseki no Tomo, 11, 2–3. (Japanese)
- Kojima, M. (2022a). Japanese coal transition reexamined: the remains of coal [Unpublished PhD Dissertation, Tokyo University of Foreign Studies].
- Kojima, M. (2022b). Toward a Sustainable Coal Transition: A Case Study of the Omine Coalfield. The International Journal of Social Sustainability in Economic, Social, and Cultural Context, 18(2), 61–76. https://doi.org/10.18848/2325-1115/CGP/v18i02/61-76
- Kriwet, J., & Benton, M. J. (2004). Neoselachian (Chondrichthyes, Elasmobranchii) diversity across the Cretaceous–Tertiary boundary. Palaeogeography, Palaeoclimatology, Palaeoecology, 214(3), 181–194. https://doi.org/10.1016/s0031-0182(04)00420-1
- Maeda, H., & Oyama, N. (2019). Yamaguchi-ken seibu ni bunpu suru Sanjō-kei Mine-sōgunto Jura-kei Toyoura-sōgun no sōjo to kaseki-gun [Stratigraphy and fossil assemblages of the Triassic Mine Group and Jurassic Toyora Group in western Yamaguchi Prefecture]. The Journal of the Geological Society of Japan, 125, 585–594. https://doi.org/10.5575/geosoc.2019.0020 (Japanese)
- Maisch, M. W., & Matzke, A. T. (2016). A new hybodontid shark (Chondrichthyes, Hybodontiformes) from the Lower Jurassic Posidonienschiefer Formation of Dotternhausen, SW Germany. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 280, 241–257. https://doi.org/10.1127/njgpa/2016/0577
- Maisey, J. G. (1989). Hamiltonichthys mapesi, g. & sp. nov. (Chondrichthyes, Elasmobranchii), from the Upper Pennsylvanian of Kansas American Museum of Natural History Novitates, 2931, 1–42. https://www.biodiversitylibrary.org/item/317150
- Motani, R., Minoura, N., & Ando, T. (1998). Ichthyosaurian relationships illuminated by new primitive skeletons from Japan. Nature, 393, 255–257. https://doi.org/10.1038/30473
- Murry, P. A. (1981). A new species of freshwater hybodont from the Dockum Group (Triassic) of Texas. Journal of Paleontology, 55, 603–607.
- Murry, P. A., & Kirby, R. E. (2002). A new hybodont shark from the Chinle and Bull Canyon Formations, Arizona, Utah, and New Mexico. Bulletin of the New Mexico Museum of Natural History and Science, 21, 87–106.
- Naito, G. (2000). Mine-sōgun-san shokubutsu kaseki [Fossil Plants from the Mine Group]. Mine City Board of Education. (Japanese)
- Novikov, I. V. (2010). New data on trematosauroid labyrinthodonts of Eastern Europe: 2. Trematosaurus galae sp. nov.: Cranial morphology. Paleontological Journal, 44, 457–467. https://doi.org/10.1134/S003103011004012X
- Oishi, S. (1932). Rhaetic Plants from Province Nagato Yamaguchi Prefecture, Japan. Journal of the Faculty of Science, Hokkaido Imperial University, 2, 51–68.
- Omine Coalfield Memoirs Editorial Committee. (2000). Ōmine tanden kaikoroku Muentan [Muentan: Omine Coalfield Memoirs]. Mine-shi Kyōiku Iinkai [Mine City Board of Education]. (Japanese)
- Oyama, N., & Maeda, H. (2020). Madygella humioi sp. nov. from the Upper Triassic Mine Group, southwest Japan: the oldest record of a sawfly (Hymenoptera: Symphyta) in East Asia. Paleontological Research, 24, 64–71. https://doi.org/10.2517/2019PR005
- Oyama, N., Shinoda, K., Takahashi, H., Doi, E., & Béthoux, O. (2023). A new species of the Triassic genus Ideliopsina (Grylloblattida: Ideliidae) from the Ominé locality (Momonoki Formation, southwest Japan). Palaeoentomology, 6, 205–213.
- Oyama, N., Yukawa, H., & Maeda, H. (2020). Nihon-san chūseidai konchū kaseki: toku ni Jōbu Sanjō-kei Mine-sōgun-san konchū kaseki no ichizuke ni tsuite [Mesozoic insect fossils of Japan: significance of the Upper Triassic insect fauna of the Mine Group, Yamaguchi Pref]. Bulletin of the Mine City Museum Yamaguchi Prefecture, Japan, 33, 1–13. (Japanese)
- Parker, W. G. (2005). Faunal Review of the Upper Triassic Chinle Formation of Arizona. Mesa Southwest Museum Bulletin, 11, 34–54.
- Patterson, C. (1966). British wealden sharks. Bulletin of the British Museum (Natural History) Geology, 11, 281–350. https://doi.org/10.5962/p.150189
- Peecook, B. R., Steyer, J. S., Tabor, N. J., & Smith, R. M. H. (2017). Updated geology and vertebrate paleontology of the Triassic Ntawere Formation of northeastern Zambia, with special emphasis on the archosauromorphs. Journal of Vertebrate Paleontology, 37, 8–38. https://doi.org/10.1080/02724634.2017.1410484
- Prasad, G. V. R., Singh, K., Parmar, V., Goswami, A., & Sudan, C. S. (2008). Hybodont shark teeth from the continental Upper Triassic deposits of India. Mesozoic Fishes 4, 413–432.
- Rees, J. (2008). Interrelationships of Mesozoic hybodont sharks as indicated by dental morphology-Preliminary results. Acta Geologica Polonica, 58, 217–221.
- Rees, J., & Underwood, C. J. (2002). The status of the shark genus Lissodus Brough, 1935, and the position of nominal Lissodus species within the Hybodontoidea (Selachii). Journal of Vertebrate Paleontology, 22, 471–479. https://doi.org/10.1671/0272-4634(2002)022[0471:TSOTSG]2.0.CO;2
- Shang, Q., Cuny, G., & Chen, L. (2008). Early Middle Jurassic vertebrate microremains from the Three Gorges area, southern China. Historical Biology, 20, 87–99. https://doi.org/10.1080/08912960701616935
- Shikama, T., Kamei, T., & Murata, M. (1978). Early Triassic Ichthyosaurus, Utatsusaurus hataii Gen. et Sp. Nov., from the Kitakami Massif, Northeast Japan. The Science Reports of the Tohoku University. Second series, Geology, 48, 77–97.
- Society of Writers Editors and Translators. (2018). Japan Style Sheet (Third Edition). https://doi.org/10.2307/489221
- Sophia University. (2018). Monumenta Nipponica Style Sheet. Sophia University.
- Stumpf, S., & Kriwet, J. (2019). A new Pliensbachian elasmobranch (Vertebrata, Chondrichthyes) assemblage from Europe, and its contribution to the understanding of late Early Jurassic elasmobranch diversity and distributional patterns. PalZ, 93, 637–658. https://doi.org/10.1007/s12542-019-00451-4
- Stumpf, S., López-Romero, F. A., Kindlimann, R., Lacombat, F., Pohl, B., Kriwet, J., & Cavin, L. (2021). A unique hybodontiform skeleton provides novel insights into Mesozoic chondrichthyan life. Papers in Palaeontology, 7(3), 1479–1505. https://doi.org/10.1002/spp2.1350
- Taira, A. (2001). Tectonic evolution of the Japanese island arc system. Annual Review of Earth and Planetary Sciences, 29, 109–134. https://doi.org/10.1146/annurev.earth.29.1.109
- Takahashi, H., Naito, G., Ishida, H., & Fukudomi, T. (1992). Kokudō 435-gōsen dōro kairyōkōji chōsahōkoku –shokubutsu kaseki ni tsuite– [Survey report on highway improvement along Japan National Highway 435: Plant Fossils]. Bulletin of the Mine City Museum, Yamaguchi Prefecture, Japan, 8, 1–23. (Japanese)
- Tokuyama, A. (1958). Late Triassic Palaeopharus in Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 32, 291–298.
- Tokuyama, A. (1959a). “Bakevellia” and “Edentula” from the Late Triassic Mine series in west Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 35, 147–155.
- Tokuyama, A. (1959b). Late Triassic Pteriacea from the Atsu and Mine series, west Japan. Japanese Journal of Geology and Geography, 30, 1–19.
- Tokuyama, A. (1960a). Late Triassic pelecypod fauna of the Aso formation in west Japan. Japanese Journal of Geology and Geography, 31, 23–38.
- Tokuyama, A. (1960b). On the bio- and litho-facies of the Late Triassic Hirabara Formation in Province of Nagato, west Japan. Japanese Journal of Geology and Geography, 31, 185–200.
- Tokuyama, A. (1960c). On the pelecypod fauna of the Late Triassic Hirabara Formation in west Japan. Japanese Journal of Geology and Geography, 31, 201–217.
- Ueda, K. (1991). A Triassic fossil of scorpion fly from Mine, Japan. Bulletin of Kitakyushu Museum of Natural History, 10, 99–103.
- Uyeno, T., Takahashi, F., Doi, E., & Naito, G. (1996). Yamaguchi-ken Mine-shi narabini San'yō-chō kara no Sanjō-ki Gyorui kaseki-gun no sanshutsu [Triassic fishes collected in Mine City and Sanyocho in Yamaguchi Prefecture]. Bulletin of the Mine City Museum, Yamaguchi Prefecture, Japan, 12, 1–27. (Japanese)
- Uyeno, T., Yabumoto, Y., & Takahashi, F. (2003). Yamaguchi-ken San'yō-chō, Sanjō-ki Mine-sōgun no Pareonisukusu-me Gyorui no shin hyōhon [A new specimen of palaeonisciform fish from Triassic Mine Formation from Sanyo-cho, Yamaguchi Prefecture, Japan]. Bulletin of the Mine City Museum, Yamaguchi Prefecture, Japan, 18, 1–17. (Japanese)
- Wakita, K., Nakagawa, T., Sakata, M., Tanaka, N., & Oyama, N. (2021). Phanerozoic accretionary history of Japan and the western Pacific margin. Geological Magazine, 158, 13–29. https://doi.org/10.1017/S0016756818000742
- Wen, W., Kriwet, J., Zhang, Q., Benton, M. J., Duffin, C. J., Huang, J., Zhou, C., Hu, S., & Ma, Z. (2022). Hybodontiform shark remains (Chondrichthyes, Elasmobranchii) from the Lower Triassic of Yunnan Province, China, with comments on hybodontiform diversity across the PTB. Journal of Vertebrate Paleontology, 42, e2108712. https://doi.org/10.1080/02724634.2022.2108712
- Yabumoto, Y., & Uyeno, T. (1994). Late Mesozoic and Cenozoic fish faunas of Japan. Island Arc, 3, 255–269. https://doi.org/10.1111/j.1440-1738.1994.tb00115.x
- Yamagishi, H. (2004). Elasmobranch remains from the Taho Limestone (Lower-Middle Triassic) of Ehime Prefecture, Southwest Japan. Mesozoic Fishes 3: Systematics, Paleoenvironments and Biodiversity, 565–574.
- Yokoyama, M. (1890). Sanjō-ki shokubutsu no hakken [The discovery of Triassic plants]. Chigaku Zasshi [Journal of Geography], 2, 293–294. https://doi.org/10.5026/jgeography.2.293a (Japanese)
- Yokoyama, M. (1891). On some fossil plants from the Coal-Bearing Series of Nagato. The journal of the College of Science, Imperial University, Japan, 4, 239–247.
- Yokoyama, M. (1905). Mesozoic Plants of Nagato and Bitchu. Journal of the College of Science, Imperial University, Tokyo, Japan, 20, 1–13.
- Yukawa, H., Shinoda, K., & Suzuki, S. (2014). Mine-shi Kaseki Saishū-ba ni okeru Jōbu Sanjō-kei Mine-sōgun Momonoki-sō no taiseki kankyō to shokubutsu kaseki no san-jō [Occurrences of plant fossils and depositional environment of the Upper Triassic Momonoki Formation of the Mine Group at the Mine Fossil Collection Facility, Yamaguchi Prefecture, SW Japan]. Bulletin of the Mine City Museum Yamaguchi Prefecture, Japan, 29, 25–30. (Japanese)