
ABSTRACT
Removal of logging residues causes significant nutrient losses from the harvesting site. Furthermore, collection of residues into piles could lead to small-scale differences in establishment conditions for seedlings. We studied the effects of stem-only (SOH) and aboveground whole-tree harvesting (WTH) on Norway spruce (Picea abies) seedling growth and pine weevil (Hylobius abietis) damage at two sites (SE and W Norway). We also compared two planting environments within the WTH plots (WTH-0: areas with no residues, WTH-1: areas where residue piles had been placed and removed before planting). In practice, one-third of the residues were left on site after WTH. After three growing seasons there were no differences for height or diameter increment between SOH and WTH (WTH-1 and WTH-0 combined) treatments. However, relative diameter increment was largest for WTH-1 seedlings and lowest for WTH-0 seedlings. Few seedlings sustained pine weevil attacks at the W Norway site, with no differences among treatments. At the SE Norway site, the percent of seedlings damaged by pine weevils and average debarked area were significantly higher after WTH (82% and 3.3 cm2) compared to SOH (62% and 1.7 cm2). We conclude that WTH may lead to spatial differences in establishment conditions.
Introduction
In many countries, biomass is increasingly being used as an energy source in order to reduce CO2 emissions from fossil fuels. Logging residues (branches, needles and tree tops) are a potential bioenergy source, especially since it is not utilized for other purposes in the forest industry. In Norway, a large part of the potential logging residues comes from Norway spruce (Picea abies (L.) Karst) stands, as it constitutes 64% of the annual cut volume (Tomter & Dalen Citation2014).
Spruce stands in Norway are usually clear-cut and new stands established through planting, often without preceding site preparation (Granhus et al. Citation2015). Planting density varies with site index, but generally lies within 1800–2500 seedlings ha−1. The removal of logging residues after clear-cutting is a form of aboveground whole-tree harvesting (WTH) and may influence the spruce regeneration in different ways. Firstly, WTH affects nutrient availability, as the amount of nutrients is much higher in needles and branches than in stem wood. According to Mälkönen (Citation1976), the removal of nitrogen (N) is 2–4 times higher after WTH than after conventional stem-only harvesting (SOH). However, in the first years after planting, the nutrient demand of seedlings is modest due to their small size, and nutrient availability is usually high due to increased decomposition of organic material at the site (Kreutzweiser et al. Citation2008). None the less, there are reports indicating that WTH negatively affects the nutrient status of seedlings and saplings (Olsson et al. Citation2000; Sikström Citation2004; Thiffault et al. Citation2006; Saarsalmi et al. Citation2010). There are no specific guidelines in Norway about leaving parts of the logging residues on site during WTH. Under practical conditions, however, some of the branches and needles will be left on site even after WTH. If the residue is left to dry for some time before it is collected, a layer of needles and small twigs will remain on the ground after the residue piles are removed. On the other hand, in between the piles the ground will be more or less without branches and twigs. This could lead to small-scale spatial differences in establishment conditions and soil nutrient availability after logging.
Secondly, biomass harvesting might influence seedling survival and growth through effects on weed competition and microclimate (Thiffault et al. Citation2011), and indirectly through soil perturbation and compaction caused by the machinery used to recover the residues (Ampoorter et al. Citation2011; Cambi et al. Citation2015). As retention of logging residues might suppress the growth of weeds (Proe & Dutch Citation1994), removing it could lead to increased vegetation competition for water and nutrients. This is especially the case in Norway, where mechanical site preparation before planting is uncommon (Granhus et al. Citation2015) and the use of herbicides is infrequent (Fløistad et al. Citation2009). Furthermore, the removal of logging residues could affect water and temperature conditions in the soil, which could in turn affect the root growth and establishment of seedlings (Proe & Dutch Citation1994; Zabowski et al. Citation2000) as well as changing the mineralization rates of N in soil organic matter (Smolander et al. Citation2010, Citation2013). There is still no consensus on how WTH will affect seedling and sapling growth. Some studies have shown small or no effects of WTH (Nilsson & Örlander Citation1995; Sikström Citation2004; Saarsalmi et al. Citation2010; Tamminen & Saarsalmi Citation2013) while for others, effects have been more evident after some years (Proe et al. Citation1996; Egnell & Valinger Citation2003; Walmsley et al. Citation2009; Egnell Citation2011; Wall & Hytönen Citation2011; Jacobson et al. Citation2016). Sometimes the effects are found to be temporal (Egnell Citation2011) or species specific (Smolander et al. Citation2015).
Finally, WTH could also affect the abundance of pine weevils (Hylobius abietis), which are probably the most important pest to conifer seedlings in Southern Scandinavia (Örlander & Nilsson Citation1999). Pine weevils typically occur in great numbers on fresh clear-cuts where they feed on the bark of young seedlings, causing death or injuries that delay growth, mainly in the first three seasons after logging (Nordlander et al. Citation2017). They use fresh conifer stumps and roots as their reproduction substrate, and are attracted to fresh clear-cuts by the smell of resin from stumps and logging residues (Eidmann Citation1974; Solbreck & Gyldberg Citation1979). The relative significance of logging residues and stumps as attractants is not well known. Thus, changes in amounts or distribution of logging residues could influence the distribution of the weevils and the subsequent seedling damage. For instance, Rahman et al. (Citation2015) found that multiple residue piles after stump harvesting increased the number of pine weevils, but decreased seedling damage. If branches and tops are removed immediately after harvest, less resin odor could potentially result in fewer weevils being attracted to the harvested area. However, in Norway logging residues are often left in piles over the summer to be harvested in the autumn. This concentration of residues could potentially attract migrating pine weevils to areas with residue piles. On the other hand, as pine weevils also feed on the bark of freshly cut branches, the presence of logging residues may reduce the feeding pressure on seedlings (Örlander et al. Citation2001).
Also the nutrient status of the seedlings could be affected by WTH, which in turn could impact on the risk of bark feeding as well as the ability of seedlings to withstand such damage. Several studies have shown that the weevils prefer seedlings with a high nutrient level (Selander & Immonen Citation1991; Zas et al. Citation2006, Citation2008), although contradictory results were obtained by Wallertz and Petersson (Citation2011), who found no difference in bark feeding damage following nutrient loading of spruce seedlings prior to planting. The risk of bark feeding leading to lethal damage is highly dependent on seedling vitality and size (Thorsén et al. Citation2001; Zas et al. Citation2014). Thus, a healthy and fast growing seedling would have more energy to withstand and survive damage, and would grow faster into a “safe” size.
There are few studies investigating how WTH affects the level of pine weevil damage. Örlander and Nilsson (Citation1999) found only a minor effect of slash removal on pine weevil damage, with reduced mortality in one out of four years. Jacobson et al. (Citation2016) reported that damage was quite evenly distributed across treatments with and without residue harvesting, but did not register pine weevil damage in detail. Thus, the knowledge of how and whether WTH will affect pine weevil damage and thereby the early survival of seedlings is still insufficient.
The rate of residue removal will influence the effects on seedling growth and survival. In most scientific studies of seedling growth after residue removal at final felling, the removal rate of logging residues is decided in advance, and usually all residues are removed in WTH (Proe et al. Citation1996; Egnell & Leijon Citation1999; Thiffault et al. Citation2006; Walmsley et al. Citation2009; Saarsalmi et al. Citation2010). In ordinary forestry, total residue removal is not practically possible and also not desirable from a biological point of view (Skogsstyrelsen Citation2008). Jacobson et al. (Citation2016) examined residue removal and seedling growth under more practical conditions. The actual proportion harvested in practical forestry in Norway has not yet been studied. Especially, there is a lack of knowledge concerning how much of the needles that fall off during storage in piles.
The main objectives of the present study were to investigate the effects of WTH after clear-cutting on the growth of planted Norway spruce seedlings and subsequent pine weevil damage. Our hypotheses were that WTH will (1) affect seedling growth negatively and (2) increase the pine weevil damage to seedlings. In addition, we studied the residue removal rate and the amount of N lost through biomass removal during practical residue harvesting.
Materials and methods
Experimental sites
The experiment was established in two sites; in 2008 at Gaupen, Ringsaker municipality (60°52′ N, 10°45′ E) in SE Norway (), and in 2010 at Vindberg, Voss municipality (60°35′ N, 6°35′ E) in W Norway (). In the following description, data on bedrock and quaternary deposits are from the Norwegian Geological Survey (http://geo.ngu.no/kart/arealisNGU/) while the climate data are based on 1961–1990 normals from the nearby climate stations Kise (Gaupen) and Reimegrend (Vindberg) (The Norwegian Meteorological Institute, http://www.eklima.no). Temperatures are corrected for altitude (Laaksonen Citation1976).
Figure 1. Map of the experimental sites Gaupen and Vindberg in SE and W Norway.
The site at Gaupen was located at 200 m a.s.l. on a fertile forest type, with site index G20 (H40) (Tveite Citation1977) corresponding to a yield capacity of 9.5 m3 ha−1 year−1. The bedrock consists of Precambrian sandstones that form a gentle W–SW facing slope, covered by a thin layer of moraine deposits. Annual precipitation is 585 mm year−1, and the annual mean temperature is 3.2°C. The forest was planted with Norway spruce in 1946/1947. Thus, the mean age of dominant trees was about 62 years at the time of felling. Stand density before harvesting was 880 trees ha−1 and the standing volume 360 m3 ha−1. The stand was dominated by Norway spruce, with some Scots pine (Pinus sylvestris L.) and scattered downy birch (Betula pubescens Ehrh.). The three species amounted to 87%, 12% and 1% of the standing volume, respectively.
The Vindberg site was a pure spruce stand located on a hillside at around 400 m a.s.l. The site index varied between plots, but on average the yield capacity was 7.0 m3 ha−1 year−1. The Cambro-Silurian bedrock forms a relatively steep N–NE facing slope, covered by thin moraine deposits. The annual precipitation is 1550 mm, and the annual mean temperature is 4.3°C. The forest stand was naturally regenerated, and has probably been selectively cut in the past. The mean age of dominant trees was about 135 years. Stand density before harvesting was 680 trees ha−1 and the standing volume 302 m3 ha−1.
Both stands were cut using a harvester and forwarder, the harvest operations at Gaupen taking place in March 2009, and at Vindberg in January 2011. Before harvest, 12 plots were established at the Gaupen site and 10 at Vindberg. Plot size was 20 × 20 m with an additional buffer zone of 5 m width at Gaupen, and 12 × 12 m with a buffer zone of 4 m at Vindberg. The lower plot number and plot size at Vindberg was due to a smaller stand and more uneven topography and stand characteristics. All measurements took place in the inner squares. The plots were paired in blocks so that of two adjacent plots, one was randomly assigned to SOH while the other was assigned to WTH. On WTH plots, logging residues were gathered in piles during felling, and harvested in September 2009 (Gaupen) and 2011 (Vindberg), respectively, using a forwarder. On SOH plots the residues were manually spread out in order to obtain a relatively uniform layer of logging residues on the ground.
Measurements of stem volume and logging residues
Before harvesting, all trees were measured and volume per plot was calculated using the volume functions of Vestjordet (Citation1967) and Brantseg (Citation1967). The amounts of biomass and N in logging residues including unmerchantable tops (diameter limit 7 cm outside bark) were estimated from sample trees. The trees in each plot were divided into three diameter classes (<20 cm DBH, 20–29.9 cm DBH and ≥30 cm DBH). Sample trees were then picked at random from each group. At Gaupen, a total of 36 trees were chosen as sample trees, while in Vindberg 20 trees were chosen. Two trees from Gaupen were later discarded due to low vitality and breakage during felling, so that the final number of sample trees was 34 and 20, respectively.
The sample trees were felled using a harvester, and then strapped and weighed using a balance (Crane Scale OCS-XZL-3t, accuracy ±1.0 kg) hung from the crane boom. The green part of the crown was divided into three parts of equal length, and two live branches from the middle of each crown part, from opposite sides of the tree, were sampled for measurement of dry weight and nutrient analyses. In addition, the total number of dead branches was counted and samples were collected for analyses. Afterwards, the tree was delimbed and the bole and stem top were strapped and weighed again. The potential fresh weight of logging residues was then calculated as total weight of the tree minus bole weight.
Branch samples were dried at 103°C until stable weight was achieved to give data on dry weight for needles and live and dead branches. Estimates for whole trees were based on ordinary dry weight to fresh weight ratio calculations. It was assumed that the stem tops had the same portion of needles as the branches. Aggregated plot data for the different biomass components were estimated by functions derived from the sample trees with DBH as the independent variable.
N concentrations in different components were analyzed by standard procedures used at the Norwegian Institute of Bioeconomy Research (Ogner et al. Citation1999).
To estimate the amount of branches and needles removed during residue harvesting in September 2009 and 2011, the forwarder was weighed with and without load as it delivered the logging residues from WTH plots to the roadside. These measurements included five of the plots at Gaupen and four of the plots at Vindberg. The weighing was performed on level ground, using a weighing station (Dini Argeo, model WWSE10T) with one weighing platform placed under each of the four trailer wheels. Residue samples were taken from two residue piles in each WTH plot, and later dried at 103°C and weighed in order to compute the dry weight of the residues at the time of harvesting.
During the summer, some of the needles in the residue piles will fall off and be left on the ground. In order to estimate the needle loss, logging residue from six trees at Gaupen was left on fiber cloths measuring 5 × 3 m in March 2009, and correspondingly two trees from Vindberg in January 2011. The amount of needles and branches put on each fiber cloth was calculated using the functions derived from the sample trees. In September 2009 (Gaupen) and September 2011 (Vindberg), the branches were removed and the needles on the cloth was collected, dried at 103°C and weighed. The ratio between the weight of needles left on the cloth and the total needle mass estimated from the derived functions was then calculated for each sample tree.
Seedlings
In May 2010 and 2012, two-year-old Norway spruce seedlings were planted at Gaupen and Vindberg, respectively. The seedlings were grown in 50 cm3 multipot containers with 95 pots per container. For the Gaupen plots the seeds originated from Sanderud seed orchard, while at Vindberg they came from Kaupanger seed orchard. In agreement with local practices, the planting densities were 2000 and 2200 seedlings ha−1, respectively. As part of normal nursery procedure, the seedlings were treated with the insecticide Merit Forest (active ingredient Imidacloprid) prior to planting to protect against pine weevils.
In each plot, 30 seedlings were labeled, and measured by recording height and diameter at ground surface directly after planting. All together 360 seedlings were measured in Gaupen and 300 in Vindberg. In the SOH plots, the 30 seedlings were randomly chosen from 2 transects going diagonally through the plots (Gaupen) or, due to smaller plots, randomly from the whole area (Vindberg). In the WTH plots, 15 seedlings were picked from areas where the residue piles had been removed the previous autumn, while the remaining 15 seedlings were chosen from transects (Gaupen) or at random evenly spread out in the plots (Vindberg), in areas where there had been no residue piles. The residue piles occupied around 30% of the plot area. Thus, the seedlings were planted in three different environments:
Areas with an even distribution of logging residues (the SOH plots),
Areas with a layer of needles, twigs and some larger branches on the ground (WTH plots where the residue piles had been placed and later removed, hereafter called WTH-1) and
Areas with no residues (WTH plots in between the residue piles, hereafter called WTH-0).
In October the first, second and third year, seedling height, leader length and diameter at stem base were measured. Pine weevil damage was recorded measuring the debarked area in cm2, and girdling of the seedlings was registered. Other types of damage (including drowning, drought, browsing and mechanical damage) were also recorded. Vitality was subjectively registered on a four-level scale where 3 = healthy, 2 = some reduced vitality, 1 = severely reduced vitality and 0 = dead. In June, the second and third growing season vitality and damage were registered.
Calculations and statistical analyses
Relative height (RH) increment after three seasons was calculated by the formula(1) where H = height after three growing seasons and He = height at establishment. A corresponding equation was used for finding the relative diameter increment.
Because the two sites were different in terms of plant material, climate and year of initialization, each site was analyzed separately.
All continuous response variables were analyzed based on the linear mixed model(2) where Yijk is the response variable observed for plant k, in block j with treatment i. μ is the overall mean, τi is the fixed effect of treatment i, Bj is the random effect of block j, (τB)ij is the random interaction effect between treatment i and block j and εijk is the residual error. The analyses were performed with the main treatments SOH and WTH as well as with SOH, WTH-1 and WTH-0 as treatments. In the latter case, the sample sizes were different due to fewer measured seedlings in WTH-1 and WTH-0.
All the Bjs, (τB)ijs, and εijks are assumed to be independent and normally distributed, with expected means equal to zero. The model in (2) contain three variances, one for the Bjs, one for the (τB)ijs, and for the εijks. The response variable for debarked area per seedling was square root transformed before the analysis, in order to stabilize the variance. The proportion of seedlings attacked by pine weevils was analyzed using a generalized linear mixed model with the treatments, blocks, and their interactions as in (2). The response variable was assumed to follow a binary distribution and we used the logit link function.
The value 0.05 was used as significance level for all analyses. We used SAS® version 9.4 (SAS® Institute Inc., Cary; NC, USA) for the statistical analyses, applying the Mixed procedure for the linear mixed model and the Glimmix procedure for the generalized linear mixed model. The Satterthwaite approximation was used to calculate the denominator degrees of freedom. The statistical significance of differences between individual treatments was assessed by Tukey’s test.
Results
Biomass and N loss with residue removal
The calculated amounts of biomass and N in the residues, as well as the amounts actually removed, are shown in . Even though the stem volume per hectare was larger at Gaupen than at Vindberg, the amount of logging residues was higher at Vindberg. Due to more uneven stand characteristics, the variation in residue amounts between plots was also larger at Vindberg. According to our harvester load data, about two-thirds of the potential amount of logging residues was actually removed from the stand. In our calculations of the amounts of N removed from the sites we assumed that 20% of the needles fall off during seasoning (see below), and that another 5% was lost during handling and transport. Thus, as a comparably larger amount of needles are left at the site compared to the more nutrient poor branches, the corresponding amount of nutrients removed is smaller, accounting to 57% and 58% for the two sites.
Table 1. The amount of potential and harvested residues and nitrogen at Gaupen and Vindberg.
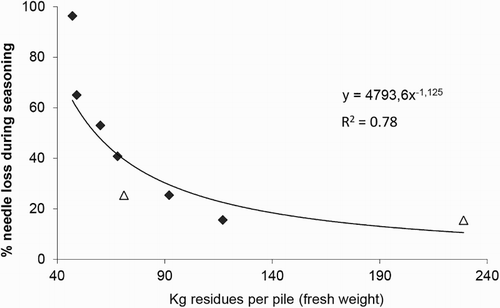
The amount of needle loss from the residue piles put on fiber cloth clearly decreased with the size of the pile (). For the larger piles, which are closest in size to actual piles made during logging, the needle loss was around 20%.
Figure 2. Needle loss (in percent) from logging residues, in relation to the size of the residue piles. The residues were put on fiber cloth and left to dry from the logging operation in winter until September. Filled squares = piles at Gaupen (2009), open triangles = piles at Vindberg (2011).
Seedling growth
After three growing seasons, the average height and diameter were 59 cm and 9.9 mm at Gaupen and 43 cm and 8.0 mm at Vindberg. The RH and diameter increment of the seedlings in the different treatments are shown in . At both sites, relative diameter increment was largest for the WTH-1 seedlings and lowest for the WTH-0 seedlings, and the differences were statistically significant (). The WTH-0 seedlings did not differ significantly from the seedlings growing in the SOH plots. On average, height increment was highest in the WTH-1 plots, but the differences were not statistically significant.
Figure 3. Effects of harvesting treatments on spruce seedling growth after three growing seasons relative to measurements taken directly after planting (height; upper panels, diameter; lower panels). Gaupen at left panels and Vindberg at the right. Different letters above the bars indicate significant differences according to Tukey’s test. SOH = stem-only harvesting, WTH-1 = whole-tree harvesting with residues left in piles for seasoning, WTH-0 = whole-tree harvesting with all branches removed during felling. Ls means ± standard errors.
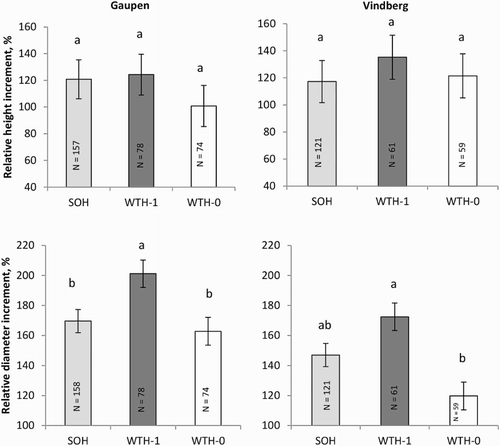
Table 2. Results of the linear mixed model Type III tests for the harvesting treatments.
There were no significant differences for height or diameter increment between the two main treatments SOH and WTH (WTH-1 and WTH-0 combined) after three growing seasons ().
Pine weevil damage
In Vindberg, the pine weevil attacks were generally low. After three seasons in the field, 19% of the seedlings had some kind of damage, and each seedling had on average 0.3 cm2 of bark gnawed. There were no statistically significant differences between the treatments at Vindberg (). At Gaupen, 71% of the seedlings had been attacked after three seasons, with an average area of 2.5 cm2. In the following, only the results for Gaupen are shown.
Table 3. Results of the linear mixed model Type III tests for the harvesting treatments.
The share of seedlings subjected to pine weevil damage is shown in . After the first season there were no statistically significant differences between the treatments. After the second and third season the percentage of seedlings with bark wounds was highest in WTH-1, and significantly higher than in SOH plots. There was also a significant difference between the two main treatments WTH and SOH after the second and third season (). After three seasons, the percentage of attacked seedlings was 62% in the SOH plots, and 82% in the WTH plots.
Figure 4. Effects of harvesting treatments on seedling damage by the pine weevil at Gaupen. (A) Percentage of attacked seedlings and (B) debarked area. Accumulated values in autumn each year. Different letters indicate significant differences according to Tukey’s test, with separate tests for each year. Means ± standard errors. For treatment abbreviations see .
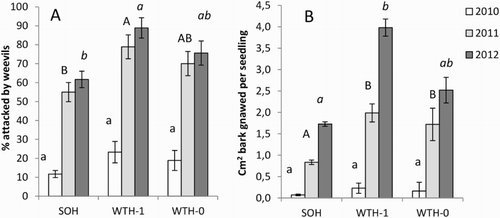
The average area of bark gnawed off on each seedling at Gaupen was small the first season after planting, but increased considerably the second season (). In the autumn this year, the area gnawed was significantly larger in WTH-1 and WTH-0 compared to SOH (). In 2012, after three growing seasons, the debarked area was still significantly larger in WTH-1 than in SOH. When the main treatments (SOH and WTH) were compared, the difference in debarked area was significant for the first and second season, and close to significance for the third season (). The average debarked area after three growing seasons was 1.7 and 3.3 cm2 for SOH and WTH, respectively.
Mortality
At both sites, pine weevils were the most important reason for seedling damage and mortality. At Vindberg, some seedlings had severe bark wounds even though the average damage was low, and 10% of the seedlings died following pine weevil attacks (12% in WTH plots and 9% in SOH). In the WTH plots at Gaupen, 12% of the seedlings had died after three seasons due to pine weevil attacks, while the corresponding number for SOH plots was 6%.
However, some seedlings also died due to mechanical damage to the bark or stem (1% at both sites), drowning (1% at Gaupen) or of unknown reasons (3% at Gaupen and 9% at Vindberg). More seedlings died from these other reasons at SOH plots that at WTH. Altogether, the mortality rates after three seasons were similar for the two main treatments: 13% for SOH and 15% for WTH at Gaupen, while at Vindberg both treatments had a mortality rate of 21%.
Discussion
Biomass and N loss with residue removal
The removal rate of forest residues will vary with operator skill, equipment, stand conditions and season (Andersson et al. Citation2002). Thiffault et al. (Citation2015) found that the average removal rate for a number of case studies from Europe and North-America was 52%. However, for Nordic (Swedish and Finnish) studies the average was 72%, varying from 23% to 89%. Our results were consistent with this average. The fact that about one-third of the residues are actually left on site, may lead to different results concerning seedling growth and development than if all branches and needles are removed during WTH. However, the residue removal still represents a considerable manipulation with nutrients. In the present study, the amount of N removed at the two sites (169 and 134 kg ha−1) is comparable with a standard forest fertilization dose (Nilsen Citation2001).
In Norway, residue harvesting is usually done by piling the residues during timber harvesting and leaving it to dry over summer. One reason for this is to allow the needles to fall off, thus diminishing the nutrient loss from the site. Olsson et al. (Citation2000) suggested that leaving the nutrient-rich needles well distributed on site may counteract negative effects of residue harvesting. Our study showed, however, that a relatively small part of the needles fall off during seasoning. This is in accordance with the findings of Jacobson et al. (Citation2016). Instead of falling down on the ground, most of the needles seemed to stick together in moist lumps in between the branches, being transported out together with the rest of the residues. However, enough needles and small twigs fall off to make a distinct layer on the ground where the residue piles had been removed.
The piles in our study were rather small, each consisting of branches from one tree only. Even though the amount of fallen needles seemed to level off at around 15–20% as pile size increased, further studies are needed to assess needle fall-off in large piles.
The weather conditions will affect needle fall-off, with dry weather expectedly giving a higher percentage. However, the percentages were comparable at Gaupen and Vindberg, even though annual rainfall was 2–3 times higher at Vindberg in 2011 compared to Gaupen in 2009 (data from local climate stations).
Effects on growth
In our study, there were no significant differences in growth between the SOH treatment on one side and the combined WTH treatments (WTH-0 and WTH-1) on the other. However, the present study shows significant differences in diameter growth between WTH-0 and WTH-1 seedlings, leading to larger internal differences within the WTH plots compared to the SOH plots. Whether this effect has been present in earlier studies that have reported no difference between WTH and SOH (e.g. Nilsson and Ôrlander Citation1995; Sikström Citation2004; Tamminen & Saarsalmi Citation2013), is not known. Some studies have shown negative effects of WTH on seedling/sapling growth on a longer time scale (Proe et al. Citation1996; Egnell & Valinger Citation2003; Egnell Citation2011; Wall & Hytönen Citation2011; Jacobson et al. Citation2016). As our study concerns growth and survival the first three seasons after planting, we cannot conclude about the long-time effects.
Even though the logging residues make up only a small part of the total ecosystem organic matter and nutrient pools, removing them means taking away relatively large amounts of important elements like N. Studies show that although the nutrient availability is rather good at a clear-cut site, additional fertilization increases early seedling growth (Johansson et al. Citation2012), as does site preparation treatments that increases nutrient supply (Hallsby Citation1995; Heiskanen & Rikala Citation2006). In a study of the decomposition rate of needles and small twigs at our two sites, Eldhuset et al. (Citation2017) found that after three years, about 25% of the N had been released from the residues. The release was faster from needles than from twigs, which is in accordance with Palviainen et al (Citation2004a). Studies have also shown increased mineralization rates of N in soil organic matter underneath residue layers (Emmett et al. Citation1991; Smolander et al. Citation2010, Citation2013). Smolander et al. (Citation2015) found that seedling growth correlated positively with net N mineralization, even though they did not find any consistent effects of logging residues on soil nutrients 10 years after logging. So, even though soil microbes may temporarily immobilize some of the nutrients (Hyvönen et al. Citation2000; Palviainen et al. Citation2004a, Citation2004b), it is likely that the seedlings planted in the SOH and WTH-1 treatment had better access to nutrients than the seedlings in WTH-0. Furthermore, seedlings planted in layers of needles in WTH-1 may have had even better nutrient supply the first seasons than seedlings from SOH, where needles were much more spread out and a larger part of the residues consisted of coarse debris like branches.
An obvious effect of the residue piles was that competing vegetation was suppressed. This effect was clearly visible in the whole registration period, especially for wavy hair-grass (Deschampsia flexuosa). Økland et al. (Citation2016) showed that three growing seasons after clear-cutting, the mean cover of graminoids for WTH-1, WTH-0 and SOH at Gaupen was 12.6%, 75.1% and 59.6%, respectively. For Vindberg, the corresponding values were 7.1%, 26.5% and 30.0%. Thus, residue harvesting created environments with large variation in vegetation regrowth, with little regrowth in WTH-1 and rapid establishment in WTH-0. Weed competition may suppress seedling growth, through competition for water, light and nutrients (Nilsson & Örlander Citation1995, Citation1999; Örlander et al. Citation1996). Mulch made from slash has been shown to create a favorable growth environment, by reducing competition and improving soil moisture and mineralization (Johansson et al. Citation2006). In the WTH-1 plots, the layer of needles and twigs may be seen as a form of mulching, even though the layer was thinner than as applied in Johansson et al. (Citation2006).
Soil temperature will influence soil respiration and mineralization of nutrients and will also affect root growth (Lyr Citation1996; Alvarez-Uria & Körner Citation2007). At our sites, the soil temperatures at 5 cm depth increased in the WTH plots after residue harvesting, compared to SOH plots (O. J. Kjønaas pers. comm). The difference was, however, only 0.2–0.5°C throughout the three first growing seasons (average of four measurements between May and October at three locations in each treatment and block). There were no differences between WTH-1 and WTH-0.
In conclusion, both potential nutrient supply, level of weed competition and soil temperature were favorable in the WTH-1 treatment, and all may have contributed to the increased growth found here. In light of the small differences in soil temperatures between treatments, it seems likely that less competition and larger nutrient release from residues played a larger role than soil temperatures.
Pine weevil damage
The pine weevil damage were more severe at Gaupen than at Vindberg in terms of debarked area and percentage of attacked seedlings. Gaupen is situated close to lake Mjøsa in Hedmark county, which is an area known to be subjected to pine weevil attacks (Hanssen Citation2012). The relatively high mortality due to pine weevils at Vindberg, despite the low average level of debarking, has no obvious explanation. It may be due to differences in plant material or climate between the two sites.
The heavier attacks from pine weevils in the WTH plots at Gaupen show that residue harvesting, performed by leaving the slash in piles for the summer, can affect the degree of seedling damage. This may be caused by higher attraction to WTH sites, and/or less alternative food sources for the weevils after residue harvesting. As the pine weevils are attracted to the smell of volatile compounds like monoterpenes and ethanol from conifer stumps and logging residue (Tilles et al. Citation1986a, Citation1986b; Nordlander Citation1987, Citation1991), it is likely that a fresh pile of logging residues will attract more weevils than scattered branches. Furthermore, the attraction is reduced as residues dry out (Långström Citation1982), which may happen more slowly in a pile. The first season, the pine weevils might use the logging residues as a food source (Örlander et al. Citation2001) until they retreat to the soil in the early autumn (Långström Citation1982). When they emerge the following spring, the piles have been removed. The dispersal distance for weevils on the ground is limited (Eidmann Citation1997), and the newly planted spruce seedlings stand out as one of few food sources, leading to severe damage in and near the areas where the piles have been removed. In the study of Rahman et al. (Citation2015), the extent of seedling damage was significantly lower in plots with stump residue piles compared to control plots, even though pine weevils were more abundant. In their study the piles were still present when the seedlings were planted, thus probably acting as a food source for the weevils. However, in our study relatively few WTH-1 seedlings died despite the rather heavy debarking. The fact that seedlings grew well in WTH-1 may to some extent have counteracted the effects of the wounds.
At our sites, plots with residue harvesting were placed side by side with plots where the branches were more or less evenly spread. We do not know whether residue harvesting in practice will lead to more pine weevils being attracted to the site, or whether it will just lead to larger small-scale variation in the weevil population within the clear-cut, and consecutive differences in damage level with distance to piles. Also during ordinary stem-only harvesting, branches are to some extent left in piles, which could lead to similar effects as in our WTH plots. Contrary to our study, Jacobson et al. (Citation2016) did not find differences in pine weevil damage between treatments with residues left in small strings, spread out, or removed. Their plots were even smaller than ours. The question of scale should thus be studied further.
Conclusions and practical implications
Our study from two climatically different areas in W and SE Norway showed that in practice, more than one-third of the residues was left at the site during harvesting of forest residues, but that leaving the residues in large piles to dry over summer made only a small part of the needles fall off. Harvesting of forest residues (WTH) did not affect early seedling growth compared to SOH in either of the sites, but it led to differences in diameter growth within the WTH plots. WTH also led to more severe pine weevil damage at the site in SE Norway, and forest practitioners should be aware of possible higher mortality due to pine weevils close to residue piles. Thus, the need for supplementary planting could be larger after residue harvesting. However, our results also show that there is a possibility of better growth for seedlings planted in areas where the residue piles have been placed and later removed. In areas where the population of pine weevils is low, these areas could represent good planting spots.
Acknowledgements
The authors thank the land owners, Leif and Anne Cathrine Blakstad at Gaupen and Lars Istad at Vindberg, for hosting the field experiments, and Hans Nyeggen and Roald Brean at NIBIO for scrupulous field work. Torfinn Torp at NIBIO is thanked for statistical advice.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Alvarez-Uria P, Körner C. 2007. Low temperature limits of root growth in deciduous and evergreen temperate tree species. Funct Ecol. 21:211–218. doi: 10.1111/j.1365-2435.2007.01231.x
- Ampoorter E, De Frenne P, Hermy M, Verheyen K. 2011. Effects of soil compaction on growth and survival of tree saplings: a meta-analysis. Basic Appl Ecol. 12:394–402. doi: 10.1016/j.baae.2011.06.003
- Andersson G, Asikainen A, Björheden R, Hall PW, Hudson JB, Jirjis R, Mead DJ, Nurmi J, Weetman GF. 2002. Production of forest energy. In: Richardson J, Björheden R, Hakkila P, Lowe AT, Smith CT, editors. Bioenergy from sustainable forestry: guiding principles and practice. Dordrecht: Kluwer Academic; p. 49–124.
- Brantseg A. 1967. Furu sønnafjells. Kubering av stående skog. Funksjoner og tabeller. (Volume functions and tables for Scots pine. South Norway). Meddr norske Skogfors Ves. 22:690–739.
- Cambi M, Certini G, Neri F, Marchi E. 2015. The impact of heavy traffic on forest soils: a review. For Ecol Manage. 338:124–138. doi: 10.1016/j.foreco.2014.11.022
- Egnell G. 2011. Is the productivity decline in Norway spruce following whole-tree harvesting in the final felling in boreal Sweden permanent or temporary? For Ecol Manage. 261:148–153. doi: 10.1016/j.foreco.2010.09.045
- Egnell G, Leijon B. 1999. Survival and growth of planted seedlings of Pinus sylvestris and Picea abies after different levels of biomass removal in clear-felling. Scand J For Res. 14:303–311. doi: 10.1080/02827589950152610
- Egnell G, Valinger E. 2003. Survival, growth, and growth allocation of planted Scots pine trees after different levels of biomass removal in clear-felling. For Ecol Manage. 177:65–74. doi: 10.1016/S0378-1127(02)00332-8
- Eidmann HH. 1974. Hylobius Schönh. In: Schwenke W, editor. Die Forstschädlinge Europas. Band 2. Hamburg: Verlag Paul Parey; p. 275–293.
- Eidmann HH. 1997. Assessment of pine weevil numbers on clear-cut and forest sites with shelter boards and pitfall traps. Anz Schadlingskd Pflanzenschutz Umweltschutz. 70:68–72. doi: 10.1007/BF02039130
- Eldhuset TD, Kjønaas OJ, Lange H. 2017. Decomposition rates and nutrient dynamics of Picea abies needles, twigs and fine roots after stem-only harvesting in eastern and western Norway. Plant Soil. (in revision).
- Emmett BA, Anderson JM, Hornung M. 1991. The controls on dissolved nitrogen losses following two intensities of harvesting in a Sitka spruce forest (N. Wales). For Ecol Manage. 41:65–80. doi: 10.1016/0378-1127(91)90119-G
- Fløistad IS, Granhus A, Hanssen KH. 2009. Norway. In Willoughby I, Balandier P, Bentsen NS, McCarthy N, Claridge J, editors. Forest vegetation management in Europe: current practice and future requirements. Brussels: COST Office; p. 91–98.
- Granhus A, Eriksen R, Moum SO. 2015. Resultatkontroll skogbruk/miljø. Rapport 2014. NIBIO Report. Ås: NIBIO; 38 pp.
- Hallsby G. 1995. Field performance of outplanted Norway spruce: effects of organic matter amendments and site preparation. Can J For Res. 25:1356–1367. doi: 10.1139/x95-148
- Hanssen KH. 2012. Snutebilleundersøkelsen 2010 – Hedmark og Oppland [The pine weevil survey 2010 – Hedmark and Oppland counties]. Oppdragsrapport fra Skog og landskap. Norsk institutt for skog og landskap. 10 pp.
- Heiskanen J, Rikala R. 2006. Root growth and nutrient uptake of Norway spruce container seedlings planted in mounded boreal forest soil. For Ecol Manage. 222:410–417. doi: 10.1016/j.foreco.2005.10.047
- Hyvönen R, Olsson BA, Lundkvist H, Staaf H. 2000. Decomposition and nutrient release from Picea abies (L.) Karst. and Pinus sylvestris L. logging residues. For Ecol Manage. 126:97–112. doi: 10.1016/S0378-1127(99)00092-4
- Jacobson S, Högbom L, Ring E, Nohrstedt H-Ö. 2016. The distribution of logging residues and its impact on seedling establishment and early plant growth in two Norway spruce stands. Scand J For Res. 32:134–141. doi: 10.1080/02827581.2016.1194456
- Johansson K, Langvall O, Bergh J. 2012. Optimization of environmental factors affecting initial growth of Norway spruce seedlings. Silva Fenn. 46:27–38. doi: 10.14214/sf.64
- Johansson K, Örlander G, Nilsson U. 2006. Effects of mulching and insecticides on establishment and growth of Norway spruce. Can J For Res. 36:2377–2385. doi: 10.1139/x06-121
- Kreutzweiser DP, Hazlett PW, Gunn JM. 2008. Logging impacts on the biogeochemistry of boreal forest soils and nutrient export to aquatic systems: a review. Environ Rev. 16:157–179. doi: 10.1139/A08-006
- Laaksonen K. 1976. The dependence of mean air temperatures upon latitude and altitude in Fennoscandia (1921–1950). Ann Acad Sci Fenn. 119, Series A:5–19.
- Långström B. 1982. Abundance and seasonal activity of adult Hylobius weevils in reforestation areas during first years following final felling. Comm Inst For Fenn. 106:1–22.
- Lyr H. 1996. Effect of the root temperature on growth parameters of various European tree species. Ann Sci For. 53:317–323. doi: 10.1051/forest:19960214
- Mälkönen E. 1976. Effect of whole-tree harvesting on soil fertility. Silva Fenn. 10:157–164. doi: 10.14214/sf.a14790
- Nilsen P. 2001. Fertilization experiments on forest mineral soils: a review of the Norwegian results. Scand J For Res. 16:541–554. doi: 10.1080/02827580152699376
- Nilsson U, Örlander G. 1995. Effects of regeneration methods on drought damage to newly planted Norway spruce seedlings. Can J For Res. 25:790–802. doi: 10.1139/x95-086
- Nilsson U, Örlander G. 1999. Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden. Can J For Res. 29:1015–1026. doi: 10.1139/x99-071
- Nordlander G. 1987. A method for trapping Hylobius abietis (L.) with a standardized bait and its potential for forecasting seedling damage. Scand J For Res. 2:199–213. doi: 10.1080/02827588709382458
- Nordlander G. 1991. Host finding in the pine weevil Hylobius abietis: effects of conifer volatiles and added limonene. Entomol Exp Appl. 59:229–237. doi: 10.1111/j.1570-7458.1991.tb01507.x
- Nordlander G, Hellqvist C, Hjelm K. 2017. Replanting conifer seedlings after pine weevil emigration in spring decreases feeding damage and seedling mortality. Scand J For Res. 32:60–67. doi: 10.1080/02827581.2016.1186220
- Ogner G, Wickstrøm T, Remedios G, Gjelsvik S, Hensel GR, Jacobsen JE, Olsen M, Skretting E, Sørlie B. 1999. The chemical analysis program of the Norwegian Forest Research Institute 2000. Ås: The Norwegian Forest Research Institute; 23 pp.
- Økland T, Nordbakken J-F, Lange H, Røsberg I, Clarke N. 2016. Short-term effects of whole-tree harvesting on understory plant species diversity and cover in two Norway spruce sites in southern Norway. Scand J For Res. 31:766–776. doi: 10.1080/02827581.2016.1164889
- Olsson BA, Lundkvist H, Staaf H. 2000. Nutrient status in needles of Norway spruce and Scots pine following harvesting of logging residues. Plant Soil. 223:163–175. doi: 10.1023/A:1004892109615
- Örlander G, Nilsson U. 1999. Effect of reforestation methods on pine weevil (Hylobius abietis) damage and seedling survival. Scand J For Res. 14:341–354. doi: 10.1080/02827589950152665
- Örlander G, Nilsson U, Hällgren J-E. 1996. Competition for water and nutrients between ground vegetation and planted Picea abies. New Zeal J For Sci. 26:99–117.
- Örlander G, Nordlander G, Wallertz K. 2001. Extra food supply decreases damage by the pine weevil Hylobius abietis. Scand J For Res. 16:450–454. doi: 10.1080/02827580152632847
- Palviainen M, Finér L, Kurka AM, Mannerkoski H, Piirainen S, Starr M. 2004a. Release of potassium, calcium, iron and aluminium from Norway spruce, Scots pine and silver birch logging residues. Plant Soil. 259:123–136. doi: 10.1023/B:PLSO.0000020938.78215.bd
- Palviainen M, Finér L, Kurka AM, Mannerkoski H, Piirainen S, Starr M. 2004b. Decomposition and nutrient release from logging residues after clear-cutting of mixed boreal forest. Plant Soil. 263:53–67. doi: 10.1023/B:PLSO.0000047718.34805.fb
- Proe MF, Cameron AD, Dutch J, Christodoulou XC. 1996. The effect of whole-tree harvesting on the growth of second rotation Sitka spruce. Forestry. 69:389–401. doi: 10.1093/forestry/69.4.389
- Proe MF, Dutch J. 1994. Impact of whole-tree harvesting on second-rotation growth of Sitka spruce: the first 10 years. For Ecol Manage. 66:39–54. doi: 10.1016/0378-1127(94)90147-3
- Rahman A, Viiri H, Pelkonen P, Khanam T. 2015. Have stump piles any effect on the pine weevil (Hylobius abietis L.) incidence and seedling damage?. Global Ecol Conserv. 3:424–432. doi: 10.1016/j.gecco.2015.01.012
- Saarsalmi A, Tamminen P, Kukkola M, Hautajärvi R. 2010. Whole-tree harvesting at clear-felling: impact on soil chemistry, needle nutrient concentrations and growth of Scots pine. Scand J For Res. 25:148–156. doi: 10.1080/02827581003667314
- Selander J, Immonen A. 1991. Effect of fertilization on the susceptibility of Scots pine seedlings to the large pine weevil, Hylobius abietis (Coleoptera, Curculionidae). Folia Forestalia. 771: 21 pp.
- Sikström U. 2004. Survival, growth and needle element concentrations of Picea abies (L.) Karst. seedlings after brash removal in a previously N fertilized stand. For Ecol Manage. 203:123–134. doi: 10.1016/j.foreco.2004.07.066
- Skogsstyrelsen. 2008. Rekommendationer vid uttag av avverkningsrester och askåterföring. Meddelande 2/2008. Skogsstyrelsen, Jönköping. 33 pp.
- Smolander A, Kitunen V, Kukkola M, Tamminen P. 2013. Response of soil organic layer characteristics to logging residues in three Scots pine thinning stands. Soil Biol Biochem. 66:51–59. doi: 10.1016/j.soilbio.2013.06.017
- Smolander A, Kitunen V, Tamminen P, Kukkola M. 2010. Removal of logging residue in Norway spruce thinning stands: long-term changes in organic layer properties. Soil Biol Biochem. 42:1222–1228. doi: 10.1016/j.soilbio.2010.04.015
- Smolander A, Saarsalmi A, Tamminen P. 2015. Response of soil nutrient content, organic matter characteristics and growth of pine and spruce seedlings to logging residues. For Ecol Manage. 357:117–125. doi: 10.1016/j.foreco.2015.07.019
- Solbreck C, Gyldberg B. 1979. Temporal flight pattern of the large pine weevil, Hylobius abietis L (Coleoptera, Curculionidae), with special reference to the influence of weather. J Appl Entomol. 88:532–536.
- Tamminen P, Saarsalmi A. 2013. Effects of whole-tree harvesting on growth of pine and spruce seedlings in southern Finland. Scand J For Res. 28:559–565. doi: 10.1080/02827581.2013.786124
- Thiffault E, Béchard A, Paré D, Allen D. 2015. Recovery rate of harvest residues for bioenergy in boreal and temperate forests: a review. Wiley Interdiscip Rev Energy Environ. 4:429–451. doi: 10.1002/wene.157
- Thiffault E, Hannam KD, Paré D, Titus BD, Hazlett PW, Maynard DG, Brais S. 2011. Effects of forest biomass harvesting on soil productivity in boreal and temperate forests – a review. Environ Rev. 19:278–309. doi: 10.1139/a11-009
- Thiffault E, Paré D, Bélanger N, Munson A, Marquis F. 2006. Harvesting intensity at clear-felling in the boreal forest: impact on soil and foliar nutrient status. Soil Sci Soc Am J. 70:691–701. doi: 10.2136/sssaj2005.0155
- Thorsén A, Mattsson S, Weslien J. 2001. Influence of stem diameter on the survival and growth of containerized Norway spruce seedlings attacked by pine weevils (Hylobius spp.). Scand J For Res. 16:54–66. doi: 10.1080/028275801300004415
- Tilles DA, Nordlander G, Nordenhem H, Eidmann HH, Wassgren AB, Bergström G. 1986a. Increased release of host volatiles from feeding scars: a major cause of field aggregation in the pine weevil Hylobius abietis (Coleoptera, Curculionidae). Environ Entomol. 15:1050–1054. doi: 10.1093/ee/15.5.1050
- Tilles DA, Sjödin K, Nordlander G, Eidmann HH. 1986b. Synergism between ethanol and conifer host volatiles as attractants for the pine weevil, Hylobius abietis (L.) (Coleoptera: Curculionidae). J Econ Entomol. 79:970–973. doi: 10.1093/jee/79.4.970
- Tomter SM, Dalen LS. 2014. Bærekraftig skogbruk i Norge. Norsk institutt for skog og landskap, Ås. 241 pp.
- Tveite B. 1977. Bonitetskurver for gran. Meddelelser fra Norsk institutt for skogforskning 33:1–84.
- Vestjordet E. 1967. Funksjoner og tabeller for kubering av stående gran [Functions and tables for the volume of standing trees. Norway spruce]. Meddr norske Skogfors Ves. 22:539–574.
- Wall A, Hytönen J. 2011. The long-term effects of logging residue removal on forest floor nutrient capital, foliar chemistry and growth of a Norway spruce stand. Biomass Bioenerg. 35:3328–3334. doi: 10.1016/j.biombioe.2010.08.063
- Wallertz K, Petersson M. 2011. Pine weevil damage to Norway spruce seedlings: effects of nutrient-loading, soil inversion and physical protection during seedling establishment. Agric For Entomol. 13:413–421. doi: 10.1111/j.1461-9563.2011.00536.x
- Walmsley JD, Jones DL, Reynolds B, Price MH, Healey JR. 2009. Whole tree harvesting can reduce second rotation forest productivity. For Ecol Manage. 257:1104–1111. doi: 10.1016/j.foreco.2008.11.015
- Zabowski D, Java B, Scherer G, Everett RL, Ottmar R. 2000. Timber harvesting residue treatment: part 1. Responses of conifer seedlings, soils and microclimate. For Ecol Manage. 126:25–34. doi: 10.1016/S0378-1127(99)00081-X
- Zas R, Björklund N, Nordlander G, Cendán C, Hellqvist C, Sampedro L. 2014. Exploiting jasmonate-induced responses for field protection of conifer seedlings against a major forest pest, Hylobius abietis. For Ecol Manage. 313:212–223. doi: 10.1016/j.foreco.2013.11.014
- Zas R, Sampedro L, Moreira X, Martins P. 2008. Effect of fertilization and genetic variation on susceptibility of Pinus radiata seedlings to Hylobius abietis damage. Can J For Res. 38:63–72. doi: 10.1139/X07-128
- Zas R, Sampedro L, Prada E, Lombardero MJ, Fernández-López J. 2006. Fertilization increases Hylobius abietis L. damage in Pinus pinaster Ait. seedlings. For Ecol Manage. 222:137–144. doi: 10.1016/j.foreco.2005.10.008