To the Editor
Mutations in the highly conserved human DKC1 gene cause the rare genetic disease X-linked recessive dyskeratosis congenita (X-DC) Citation[1]. X-DC patients display features of premature aging, mucosal leukoplakia, nail dystrophy, skin pigmentation, interstitial fibrosis of the lung, bone-marrow failure and increased susceptibility to cancer Citation[2]. Hypomorphic DKC1 mutant mice recapitulate the major features of X-DC, including increased cancer susceptibility. These mice were in fact highly prone to tumors and developed a variety of them, most commonly from lung and mammary gland, indicating that DKC1 may act as an important tumor-suppressor in vivo Citation[3]. DKC1 encodes a nucleolar protein, named dyskerin, which acts as pseudouridine synthase and constitutes one of the four core protein components of the specific H/ACA RNPs involved in RNA pseudouridylation Citation[1]. Dyskerin proved to be essential also for proper rRNA processing, thus playing multiple roles on ribosome biogenesis Citation[3]. Moreover, this protein has been recently identified as an essential component of the catalytically active human telomerase complex, together with the human telomerase reverse trascriptase (TERT) and the RNA component of telomerase (TERC) Citation[4]. A quantitative analysis of dyskerin mRNA expression was recently performed by real-time RT-PCR on a series of breast carcinomas Citation[5]. In this study, the TERC levels and the overall degree of rRNA pseudouridylation were also evaluated. The amount of dyskerin mRNA was found to be variable, but always significantly associated with TERC and rRNA pseudouridylation levels.
Considering that dyskerin and TERT are both constituents of the active human telomerase enzyme complex Citation[4] and that TERT has been indicated as a potential biomarker for colorectal cancer Citation[6], we wished to check whether dyskerin and TERT mRNA levels would vary in parallel in colorectal tumors. According to the multiple role played by dyskerin in ribosome biogenesis, its expression levels were expected to be highly variable in different patients, possibly depending on age, sex and general metabolic conditions. To take in account this aspect, in our experiments we always referred as control to adjacent non-tumor mucosa matched samples.
Normal colorectal mucosa and colorectal cancer tissues were then sampled from 8 patients affected by sporadic colon cancer and assayed by quantitative real-time RT-PCR analysis. Institutional ethical approval and informed consent were given by the patients undergoing surgery for colorectal cancer. Total RNA was extracted purified and reverse-transcribed; quantitative real-time PCR was then performed by using the iQ™ 5 Multicolor Real-Time PCR Detection System (Bio-Rad). All PCR reactions were carried out in a total volume of 15 µl using 5 ng of cDNA, 5 pmol of each primer and 7.5 µl of iQ™ SYBR Green Supermix 2X (Bio-Rad). All quantifications were normalized to the expression of large subunit of RNA pol II (POLR2A) endogenous control. PCR oligoprimers, designed by using the Primer Express software (Applied Biosystems), were: h-TERT Ex-5/6-F tgtactttgtcaaggtggatgtga; h-TERT Ex-6-R ttgatgatgctggcgatga; h-DKC1 Ex-11-F ctcggaagtggggtttaggt; h-DKC1 Ex-13-R gaggtggttgctgaagtggt; h-POLR2A Ex-22-F tgctccgtattcgcatcatg; h-POLR2A Ex-23-R tccatcttgtccaccacctctt. Quantitative PCR analysis was carried out using the 2(-Delta Delta C(T)) method (2−ΔΔCt) Citation[7].
The results obtained, shown in , indicated that TERT exhibited a variable mRNA level, although in most cases it was up-regulated respect to the matched controls (p = 0.066, one-sided t test). This observation essentially confirmed the variability of TERT mRNA levels recently described by Saleh et al. in colorectal tumor tissues respect to adjacent non-tumor mucosa Citation[6]. In contrast to the fluctuation displayed by TERT levels, in our experiments we found that the amounts of dyskerin mRNA were always significantly higher in tumors than in normal tissues (p = 0.0004, one-sided t test), despite the fact that the two proteins are equimolecular constituents of the active human telomerase enzyme complex Citation[4]. This different behaviour is likely due to the additional roles played by dyskerin as structural component of the H/ACA snoRNPs involved in rRNA processing, folding and modification. Indeed, increase of dyskerin mRNA levels is expected to be a general feature marking highly proliferating cells. In this light, the variable levels of DKC1 mRNA observed in breast carcinomas Citation[5] might be explained by the fact that SW48 in vitro cultured cells, instead of adjacent non-tumor tissues, were utilised as calibrator. This way, the fluctuation of dyskerin mRNA level existing among different patients would have been ignored.
Figure 1. Expression level of TERT and DKC1 mRNA in colon tumor tissues compared to each matched normal mucosa. This experiment was performed three times with similar results. A typical experiment is shown.
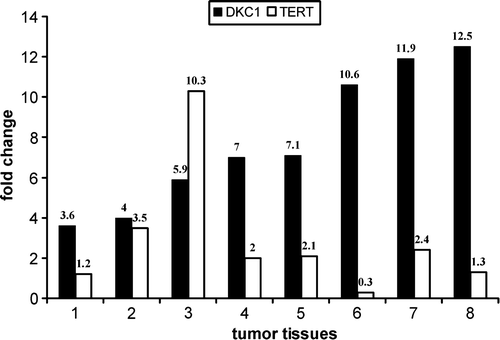
In conclusion, our data support the view that DKC1expression may furnish a less variable and more sensible marker for proliferative colon disorders. Although analysis of a more exhaustive series of tumors is needed, the high concordance of the data obtained led us to suggest that evaluation of DKC1 mRNA level can provide a useful clue in the diagnosis and prognosis of colorectal cancer, and possibly of other proliferative disorders.
References
- Heiss NS, Knight SW, Vulliamy TJ, Klauck SM, Wiemann S, Mason PJ, et al. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolare functions. Nat Genet 1998; 19: 32–8
- Dokal I. Dyskeratosis congenita in all its forms. Br J Haematol 2000; 110: 768–79
- Ruggero D, Grisendi S, Piazza F, Rego E, Mari F, Rao PH, et al. Dyskeratosis congenita and cancer in mice deficient in ribosomal RNA modification. Science 2003; 299: 259–62
- Cohen SB, Graham ME, Lovrecz GO, Bache N, Robinson PJ, Reddel RR. Protein composition of catalytically active human telomerase from immortal cells. Science 2007; 315: 1850–3
- Montanaro L, Brigotti M, Clohessy J, Barbieri S, Ceccarelli C, Santini D, et al. Dyskerin expression influences the level of ribosomal RNA pseudo-uridylation and telomerase RNA component in human breast cancer. J Pathol 2006; 210: 10–8
- Saleh S, Lam AK, Ho YH. Real-time PCR quantification of human telomerase reverse transcriptase (hTERT) in colorectal cancer. Pathology 2008; 40: 25–30
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001; 25: 402–8