
ABSTRACT
Conspecific broadcasts are effective to increase detection of owls. To determine the most appropriate time of the year to survey owls, we played conspecific owl vocalisations monthly in a temperate rainforest of southern Chile. From 12 broadcast points surveyed we recorded detections of Glaucidium nana, Strix rufipes and Tyto alba. Glaucidium nana presented a bimodal detection curve throughout the year and we recorded two regular vocalisations in response to broadcasting: contact pair call and territorial call. Strix rufipes and T. alba both showed a peak of detection between February and May. Strix rufipes presented three vocalisations: territorial call, contact pair call and female contact pair call while T. alba uttered two vocalisations: territorial call and twittering call. We recommend surveys during the end of the breeding season (austral summer–autumn) when detection is higher in most owls. Surveys should also take into consideration the variability of the vocalisations and include covariates in monitoring to evaluate occupancy/detection models.
Introduction
Owls are difficult to detect, especially for species that occur in forested areas, because of their low densities and cryptic nighttime behaviour (Barnes & Belthoff Citation2008). Using broadcasts of conspecifics is one way to increase the detectability of owls to obtain information on their behaviour, distribution, relative abundance, habitat use (Mosher et al. Citation1990), breeding densities and sex ratios (Stewart et al. Citation1996; Navarro et al. Citation2005).
In Chile, studies on owl assemblages are scarce (Jaksic et al. Citation2002; Contreras & Gonzalez Citation2007) and records of most species are incidental (e.g. Figueroa et al. Citation2000; Díaz et al. Citation2002; Jaksic et al. Citation2002, Citation2004; Mella Citation2005; Elgueta et al. Citation2006; Ibarra, Martin, Altamirano, et al. Citation2014). The owls that are easier to detect in the temperate rainforests of southern Chile are: (1) barn owl (Tyto alba), a nocturnal and crepuscular species, which occupies almost all environments, though preferring open spaces with forest fragments. This species nests in holes of old trees mainly along the forest margins during the austral spring–summer (König & Weick Citation2008); (2) rufous-legged owl (Strix rufipes), a common nocturnal and sedentary species, which occupies habitat dominated by mature and regrowth native rainforests, and agroecosystems with patches of native forest and lowland streams. This species nest in holes of old trees and sometimes on the ground, during the austral spring–autumn (Trejo et al. Citation2006; König & Weick Citation2008); and (3) austral pygmy owl (Glaucidium nana) a common and widely distributed species that is crepuscular/diurnal; however, it also shows nocturnal activity (König & Weick Citation2008; Norambuena & Muñoz-Pedreros Citation2012; Ibarra et al. Citation2015). This species occupies many types of habitats with trees and/or bushes, and it nests in tree cavities during the austral spring–summer. Despite this, their vocalisations and behaviours, for all this owls, are poorly known (Muñoz-Pedreros & Norambuena Citation2011; Raimilla et al. Citation2012), which is the basic information needed to generate adequate owl surveys (Fuller & Mosher Citation1981; Barnes & Belthoff Citation2008).
Detectability of owls may be influenced by environmental factors such as weather conditions (e.g. temperature, wind, rain and humidity), type of habitat and prey abundance, and usually shows seasonal variation (Johnson et al. Citation1981; Smith & McKay Citation1984; Clark & Anderson Citation1997; O’Donnell Citation2004; Ibarra et al. Citation2012; Ibarra, Martin, Drever, et al. Citation2014; Ibarra, Martin, Altamirano, et al. Citation2014). In this study we present a detailed quantitative description of vocal repertoires of the barn owl, austral pygmy owl and rufous-legged owl in southern Chile, and also the first evaluation of seasonal variation in their vocalisations. This will allow researchers to select the time of the year in which population censuses and nest searching for owls is most effective. This information will facilitate and improve the methods to monitor the status of these surrogate species for biodiversity conservation (Ibarra & Martin Citation2015). The objectives of this study were: (1) to characterise the vocalisations of each owl species; and (2) to examine the seasonal patterns of response (detectability) of owls during call-broadcast surveys conducted throughout the year.
Materials and methods
Study area
The study was conducted at Cerro Ñielol Natural Monument (CÑNM) (114 ha) (38°43′S, 72°35′W), a publicly protected wildlife area in the central lowland of the Araucanía region in southern Chile by the city of Temuco. Most of CÑNM (76%) is covered by temperate forest, dominated by boldo-roble (Peumus boldus–Nothofagus obliqua), peumo-boldo (Cryptocarya alba–Peumus boldus) and olivillo (Aextoxicon punctatum) stands. Open shrublands occur to a lesser extent and consist of maqui (Aristotelia chilensis), retamilla (Teline monspessulana), blackberry (Rubus ulmifolius) and colonial bentgrass (Agrostis capillaris) (Hauenstein et al. Citation1988). The CÑNM ranges from 115 m above sea level to 322 m at its summit. The climate is temperate humid with dry summers (Koeppen Citation1936). The annual temperature average is 12 °C with a maximum average of 23.5 °C in January and a minimum average of 3.9 °C in July (CIREN Citation1999). Rainfall varies between 1000 and 1500 mm annually, resulting in dry periods of two or more months (February and March; Inzunza Citation2003).
Methods
We established 12 broadcast stations with a random starting point on a road of the CÑNM, with a distance between points of 250–500 m. These covered all potential habitat types of the CÑNM (temperate forest and grasslands). The call-broadcast stations were visited monthly from March 2009 to February 2010, between 20:00 to 01:00 h for a total of 76 h of broadcasting (monthly mean of 6.33 ± 0.43 SD h). We did not perform any calling during rainy and windy nights (winds > 15 km/h; Clark & Anderson Citation1997; O’Donnell Citation2004). We broadcasted from all stations with a digital device (iPod Nano) connected to a megaphone (Power Show ER-66, 25 W), using the territorial vocalisations described in Norambuena & Muñoz-Pedreros (Citation2012) of the three owl species previously reported in the study area: barn owl, austral pygmy owl and rufous-legged owl (Norambuena & Raimilla Citation2009) and two owl species that were habitat-generalists and may hunt and/or nest within the forest (Trejo et al. Citation2006): great horned owl (Bubo virginianus magellanicus) and short-eared owl (Asio flammeus). Each call point started with 1 min of silence and a passive listening period. The broadcast time of each species lasted for 1 min, followed by a 5 min listening period for each species (for the five species we surveyed, this gave a total of 30 min for each call point). The order of the species’ calls was randomised. During the broadcast of vocalisations, the megaphone was directed towards all four cardinal directions (i.e. 15 s in each direction) and was set at shoulder height. Every time we received a response of a rufous-legged owl or a barn owl, these species were omitted from the next call-broadcast point to avoid double counting of individuals (Martínez & Jaksic Citation1996). We estimated the location of each vocally-responsive individual owl through triangulation.
During surveys we recorded each owl's response vocalisations with a Zoom H4n Handy Mobile 4-Track Recorder and a Sennheiser ME66/K6 shotgun condenser microphone. We made standard fine-scale measurements of vocalisations by using Raven Pro 1.4 (Bioacoustics Research Program Citation2011). The variables measured were: (1) call duration; (2) number of notes; (3) minimum frequency (Fmin); (4) maximum frequency (Fmax); (5) frequency of maximum amplitude (FMA) for the entire call; and (6) number of notes per second.
Vocalisation types were identified following Martínez (Citation2005) for the rufous-legged owl, who described three vocalisations: (1) contact pair call coo-coo-coo; (2) female contact pair call miiiiiiiiiooo; and (3) territorial call coo-coo- juaa-juaa. For the barn owl, we follow Marks et al. (Citation1999) who described four vocalisations: (1) territorial call shrrreeeeee; (2) twittering tiiick-tiiick-tiiick; (3) pair call shrriiee shrriiee; and (4) nestlings’ call. For the austral pygmy owl we followed Barros (Citation1949) and Jiménez & Jaksic (Citation1989) who described two vocalisations: (1) contact pair call huj-huj- huj-huj; and (2) territorial call trui-trui-yi-yi.
Nocturnal surveys were used to assess if owls were nesting. We searched our study area to identify sites in which we heard nestlings calling and/or detected pairs vocalising or defending a tree or cavity-tree during the breeding season from September to March (austral spring and summer) (sensu Martínez & Zuberogoitia Citation2002). It should be noted we were unlikely to have detected all nests on our study area and our results should be considered as minimal estimates.
Statistical analyses
We analysed our recordings in relation to the month of the year and the different periods of the reproductive cycle, as defined for the Southern Hemisphere by Marks et al. (Citation1999): non-breeding period (April–July); courtship (August–September); incubation (October–November); nestling period (December–January); and post-fledging period (February–March). To determine the occupancy and detection probabilities of owls during call-broadcast surveys, we analysed presence/absence data by using single-season occupancy models (MacKenzie et al. Citation2003) in PRESENCE 8.3 software (Hines Citation2006). This model includes two basic parameters, Ψ (the probability that a site is occupied by the species) and ρ (the probability of detecting the species, given that it was present; MacKenzie et al. Citation2003, Citation2006). For estimating Ψ we included two site-specific covariates: (1) linear distance of broadcast points to urban population (dis), calculated as a straight line from the broadcast station to the closest point of the urban radius, using a geographic information system; and (2) vegetation or canopy cover (cov), measured as a proportion (%) of the sky covered by the canopy in each broadcast stations; this was calculated in the day by using plots obtaining an average value for each broadcast point. For ρ we also include two sampling-occasion covariates or time-varying covariates: (1) median temperature (T°); and (2) presence of a nest of the target species near (< 100 m) the broadcast station. We also evaluated the potential seasonal effects (season) on our candidate set of single-season occupancy models included a null model Ψ (.), ρ, where occupancy and detection probabilities were constant across space and time (seasons), in addition to 22 alternative models incorporating explanatory covariates singly or combined to predict Ψ or ρ. We ranked models using the information–theoretic approaches (AIC; Burnham & Anderson Citation2002).
To analyse the temporal variation in vocalisations we calculated a monthly detection rate as the number or frequency of detections and vocalisations of each species divided by the total number of broadcast stations for each month (sensu Zembal & Massey Citation1987; Crozier et al. Citation2003). We used the Kruskal–Wallis H-test and Mann–Whitney U-test with α = 0.05 to compare monthly variation in vocalisation rates for each owl species.
Results
Owl detections
We recorded 153 detections of three owl species from 144 events at 12 broadcast points. In four events more than one individual was registered. Austral pygmy owls were the most frequently detected species (63.4% of detections), followed by rufous-legged owls (24.8%) and barn owls (11.8%); great horned owls and short-eared owls were not detected.
We evaluated a total of 22 occupancy models for each owl species. The models with the greatest support for the austral pygmy owl contained either none or one covariate for Ψ and ρ (). Probabilities of occupancy were negatively associated with distances less than 339 ± 183 m to an urban population and positively associated with medium (40%–50%) to high vegetation cover (> 70%; ). The three best models suggest that the probability of detecting an austral pygmy owl increased with the presence of conspecific nesting territories at the broadcasting point (). Detectability varied throughout the season from a minimum of ρ = 0.29 ± 0.07 during the non-breeding period and early courtship, and a maximum of ρ = 0.66 ± 0.07 during the post-fledging period.
Table 1. Model selection statistics based on Akaike's Information Criterion (AIC) for estimating probability of occurrence (Ψ) and detection (ρ) of three owl species in temperate rainforests of southern Chile. Site-specific covariates: linear distance of broadcasts points to urban population (dis), and vegetation or canopy cover (cov), as a proportion (%) of the sky covered by canopy in each broadcast points. Sampling-occasion covariates or time-varying covariates: median temperature (T°) and presence of nest of the target species near (< 100 m) the broadcast point (nest). Only the top five model sets are shown.
The model with highest support for rufous-legged owl contained two covariates for Ψ and ρ (). Probabilities of occupancy were positively associated with distances greater than 412 ± 325 m to an urban population and with vegetation cover greater than 50% (). The best model suggests that the probability of detecting a rufous-legged owl increased with temperature (T°) and the presence of conspecific nesting territories around the broadcasting point (). The detectability of rufous-legged owls increased during the non-breeding period with a peak of ρ = 0.66 ± 0.03.
The models with highest support for barn owl contained either none or one covariate for Ψ (). Probabilities of occupancy were associated with distance to an urban population (); however, the occupancy was equal for every distance to the broadcasting point. One of the models was positively associated with vegetation cover greater than 40% (). The probabilities of occupancy of barn owl were not associated with temperature or with the presence of nesting territories around the broadcasting point (). The detectability of barn owl did not vary seasonally and was low for the top three selected models (0.14 to 0.44).
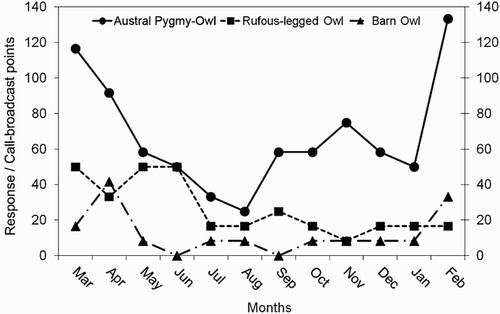
Based on the frequency of detections, austral pygmy owls exhibited a bimodal curve of detection throughout the year, with a peak between September and December (austral spring) during the breeding period, and another peak between February and April (austral autumn) when juveniles joined the vocalisations of adults (). Rufous-legged owls and barn owls showed a peak between February and May (austral summer and autumn) during the fledging period, and then decreased their vocal activity during the incubation and rearing periods ().
Figure 1. Annual detection patterns of the three owl species recorded in Cerro Ñielol Natural Monument, southern Chile (March 2009–February 2010).
Vocalisations
The austral pygmy owl used mainly two vocalisations: contact pair call (44.7%) and territorial call (55.3%), whose frequencies were similar throughout the year (U = 80.0; P = 0.66). The contact pair call was a whistle of one note of low frequency (FMA: 1421.2 ± 86.1 Hz) (; A). This vocalisation showed two peaks, the highest between courtship and nestling period and a lower peak at the end of the post-fledgling period, with a marked decrease during the non-breeding period (A). The territorial call was a sharp trill (FMA: 3674.9 ± 198.9 Hz) with an even progression of ascending notes, similar to the contact pair call, but call duration was shorter (3.1 ± 0.6 sec) (; B). This vocalisation showed a peak during the post-fledging period, with a marked decrease during the non-breeding period and a non-linear increase during the courtship and nestling period. Another four vocalisations, quantitatively undescribed, were recorded less frequently. We describe these here: (3) nestling call or trigigick, reported by König & Weick (Citation2008), is a series of soft metallic chirps (FMA: 6477.2 ± 449.2 Hz) with a fast progression of ascending notes (; D); (4) courtship call given as a short whistle (FMA: 5512.5 Hz) during the courtship period, and described as ti-ri-ri-ri-ri-rit (; E); (5) diuh diuh call (FMA: 3617.6 Hz) (F); and (6) tick tiririck call (FMA: 5857.1 ± 243.6 Hz). Both of the two latter calls were made by both sexes, usually before the territorial call is uttered (; C).
Figure 2. Spectrograms showing the vocalisations recorded from the three owl species. A, Austral pygmy owl contact pair call; B, austral pygmy owl territorial call; C, austral pygmy owl tick tiririck call; D, austral pygmy owl nestling call or trigigick call; E, austral pygmy owl courtship call; F, austral pygmy owl diuh diuh call; G, rufous-legged owl territorial call; H, rufous-legged owl female contact pair call; I, rufous-legged owl contact pair call; J, barn owl territorial call; K, barn owl twittering call.
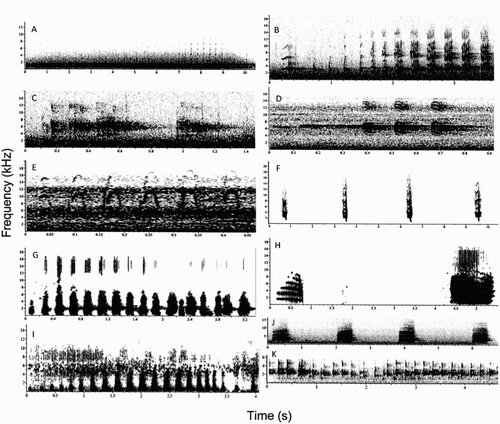
Figure 3. Annual variation in vocalisations in the three owl species in Cerro Ñielol Natural Monument, southern Chile (March 2009–February 2010). A, Austral pygmy owl; B, rufous-legged owl; C, barn owl.
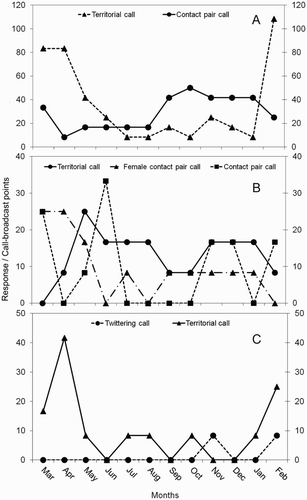
Table 2. Fine-structural measurements of the vocalisations of three owl species. The results are presented as mean ± SD.
The rufous-legged owl used three vocalisations: territorial call (40.4%), contact pair call (29.8%) and female contact pair call (29.8%), whose frequencies were similar throughout the year (H2 = 1.96; P = 0.37). The territorial call is a set of two notes; one of low frequency co-co-co followed by a rapid ascendant jua-jua-jua (FMA: 2583.9 ± 322.3 Hz) (; G). This vocalisation was used more regularly throughout the year, with a peak during the post-fledging period, a decrease during the non-breeding period, and another peak during the incubation and nestling periods (B). The female contact pair call (H) was a high-pitched vocalisation (FMA: 3330.4 ± 99.5 Hz; Fmax: 14644.5 ± 6944.2 Hz) used irregularly, with a peak during the non-breeding period, a non-linear decrease during courtship and beginning of incubation, and another peak during the nestling period (B). The contact pair call is a single, low frequency note (FMA: 172.3 Hz; Fmax: 11.025 Hz) (; I).
The barn owl used two vocalisations: territorial call (88.2%) and twittering (11.8%), whose frequencies were different throughout the year (U = 111.0; P = 0.012). The territorial call was a rasping screech of high frequency (FMA: 2756.2 Hz; Fmax: 10768.6 Hz) (; J), with a peak during the post-fledging period when juveniles joined the vocalisations of adults, not being detected in June, September, November or December. The twittering call is a single low note (FMA: 2239.5 Hz; ; K) only detected twice during the incubation and nestling periods (C).
The response to broadcasting showed a higher specificity in austral pygmy owl, whereas the rufous-legged owl was hetero-specific, responding preferentially to the broadcast of great (Magellanic) horned owl and less frequently to the broadcast of conspecifics. Barn owls responded more frequently to the broadcast of great (Magellanic) horned owl and rufous-legged owl ().
Table 3. Frequency of responses (%) of austral pygmy owl, rufous-legged owl and barn owl to broadcasting of five owl species, in the Cerro Ñielol Natural Monument, southern Chile (March 2009–February 2010).
Discussion
The owls recorded in the CÑNM showed a non-linear detection pattern, which varied depending on the species and the period of the reproductive cycle. The highest peak of detection for the three owl species occurred during the nesting period, when the breeding pairs defend their territories (Ritchison et al. Citation1988; Martínez Citation2005), and the post-fledging period when juveniles and parents frequently call to each other and both age classes readily respond to calls of other owls (Zembal & Massey Citation1987). We found another peak of detection in austral pygmy owls during the courtship and incubation periods, probably associated with increased territoriality and nest-guarding behaviour (Barros Citation1949; Goodall et al. Citation1951; Marks et al. Citation1999). The lowest peak of detection for the three owls occurred during the non-breeding period when the breeding pairs decreased their territorial defence, reducing their response to broadcasts in their nesting sites, despite still being present in their territories. Ibarra et al. (Citation2012) also found a decline of vocal activity in austral pygmy owls and rufous-legged owls in Andean forests during the non-breeding period. However, this decline was attributed to adverse environmental or climatic conditions of the area (i.e. presence of snow) that might force the owls to leave the area (Ibarra et al. Citation2012). For barn owl and rufous-legged owl the lowest detection period continued until the incubation and beginning of the nestling period and could be explained by a decrease in vocal activity of juveniles and non-territorial owls to avoid antagonistic encounters with breeding individuals (McNicholl Citation1978; Zembal & Massey Citation1987) and by a decrease of the vocalisations emitted by females during incubation, to avoid revealing their location to potential predators (Tremain et al. Citation2008). This low vocal activity of rufous-legged owl between non-breeding and nestling periods has also been recorded in temperate rainforests of Argentina (Trejo et al. Citation2011).
The probability of occupancy and detection showed a contrasting pattern between owl species, and was influenced by site-specific and sampling-occasion covariates. Only in the rufous-legged owl did we find an effect on detection probability during the non-breeding period, probably by a behavioural response of parents against the presence of juveniles on their territory after breeding. Austral pygmy owl was favoured by proximity to an urban population, whereas, in contrast, the rufous-legged owl was favoured by increasing distance to urban areas. The detection of the barn owl in the study area followed an irregular pattern (it was detected in all the broadcasting points) that can explain its lack of association with distance to an urban population. Even considering the fact that we modelled some detection covariates, other important covariates were not included in the analysis, such as amount and brightness of moonlight, which might better explain the detectability of owls (see Ibarra, Martin, Altamirano, et al. Citation2014). The presence of nesting individuals near the broadcasting points had a positive effect on detections in two of the three owl species.
Despite being considered a principally crepuscular/diurnal species (Norambuena & Muñoz-Pedreros Citation2012), austral pygmy owls quite often responded to broadcasting during sunrise. The contact pair call was associated with the courtship and incubation period (Barros Citation1949; Goodall et al. Citation1951; Marks et al. Citation1999), whereas territorial calling was higher during the post-fledging period and used irregularly during the incubation and nestling periods. This indicates that the territorial call would not be used during foraging, as indicated by Goodall et al. (Citation1951); rather, it corresponds to an aggressive vocalisation similar to that described for rufous-legged owl (see Martínez Citation2005), apparently using it to repel competitors during the breeding period and to keep juveniles or intruders away from their territories during the non-breeding period. Unlike what has been reported for rufous-legged owl by Martínez (Citation2005), the territorial call was recorded more frequently than the contact pair call, and the female contact pair call was used throughout the entire year, with a higher peak during the post-fledging period (February–March). The nonlinear patterns in the contact pair call and female contact pair call could be related to the alarm and contact functions of these vocalisations (Martínez Citation2005). Territorial defence in rufous-legged owls is highly cooperative, with both members of the pair responding aggressively to the broadcasting (mainly with territorial calls); this cooperative defence corresponds to the most effective strategy to maintain the breeding territory (Ens et al. Citation1993) and also the pair bond (Perrins & McCleery Citation1985; Bradley et al. Citation1990). In austral pygmy owl and barn owl we could not determine if cooperative defence existed because of the difficulty in differentiating the sexes from their vocalisations. Although barn owl has a diverse vocal repertoire (Marks et al. Citation1999), it vocalised infrequently and the territorial call is most frequently used during courtship and territorial defence (König et al. Citation1999). The twittering call tends to be exclusively used by adults during their flights around the nest (König et al. Citation1999) and also during hunting (E. Rodríguez, Universidad de Concepción, Chile 2016, pers. comm.).
The low hetero-specific response in austral pygmy owl suggests that this species defends its territory primarily from conspecifics, but has a reduced response to larger species (i.e. great [Magellanic] horned owl) due to the predation risk. However, Ibarra, Martin, Altamirano, et al. (Citation2014) found that the probability of detecting an austral pygmy owl increased from 0.34 to 0.62 when a rufous-legged owl was detected; the same occurred with the rufous-legged owl when an austral pygmy owl was detected in the same area (Ibarra, Martin, Altamirano, et al. Citation2014). In contrast, barn owls and rufous-legged owls had highly hetero-specific responses in our study, indicating that they likely need to defend their territories more often from other larger species compared to conspecifics. However, further broadcasting experiments are needed to determine if this is a widespread pattern in each species. The results we obtained are a first step for future research in these models and we recommend further work with more detailed information on the presence of the nest and, where possible, with larger sample sizes.
Recommendations
The results of our study suggest that consideration of time of the year is critical when using the broadcasting method to survey for owls. Due to seasonal variation in the detection and vocalisations used by owls it is recommended to conduct surveys at the end of the breeding season between December and April (austral summer–autumn). Additionally, conducting surveys during this period will reduce disturbance during nesting (see Proudfoot & Beasom Citation1996). However, if the objective is to map nesting territories, the sampling surveys should begin at the onset of the breeding period (September–December or austral spring). In addition, two other variables must be considered: (1) favourable weather conditions without rain or extreme temperatures, and wind speeds lower than 15 km/h (see Johnson et al. Citation1981; Smith & McKay Citation1984; Clark & Anderson Citation1997; O’Donnell Citation2004); and (2) type of environment being surveyed as call-broadcasts and detections are affected by vegetation cover (see Schieck Citation1997; Ibarra, Martin, Drever, et al. Citation2014). In forest environments it is recommended to increase the number of call-broadcast points (Schieck Citation1997).
We also recommend that researchers consider three aspects and potential biases of the broadcast method before reaching conclusions: (1) the ‘false absence’ or ‘silent owls’; (2) double counts; and (3) sampling covariates. Non-response (‘silent owls’) cannot be considered as evidence for the non-presence of an owl in the study area (‘false absence)’. If sampling effort (broadcasting) is increased both temporarily (e.g. during the entire high detectability period) and spatially (i.e. more call-broadcast points), then the probability that the owl hears the broadcast and responds to it should increase. It is also possible to include the false detection in occupancy model analysis (see MacKenzie et al. Citation2003). Home range studies for the species being surveyed may shed light on the spacing between broadcast points and used to adjust methodology. Double counts can be controlled by increasing/reducing the distance between call-broadcast points (without exceeding the average home range of the target species). Finally, the inclusion of covariates and evaluation of different occupancy models can provide a better description of the observed data, and observers should consider all the covariates described in Ibarra, Martin, Altamirano, et al. (Citation2014) and in this study.
Acknowledgements
We would like to express our thanks to V. Raimilla, R. Reyes, S. Zamorano, M. Jara, J. Riquelme, J. Norambuena, E. von Jentschyk, J. Vega (CONAF) and to the ranger team of the CÑNM for their support in the field. We also thank B. Guiñez, P. Sánchez and J.E. Jiménez for providing literature and comments on the manuscript and to E. Sandvig and A. Jaramillo for checking the English and commenting on the manuscript. We also thank two anonymous reviewers for comments that greatly improved the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Barnes KP, Belthoff JR. 2008. Probability of detection of flammulated owls using nocturnal broadcast surveys. Journal of Field Ornithology. 79(3):321–328. doi: 10.1111/j.1557-9263.2008.00166.x
- Barros R. 1949. La historia del chucho, Glaucidium nanum (King). Rev Univ (Chile). 35:17–30.
- Bioacoustics Research Program. 2011. Raven Pro: Interactive Sound Analysis Software (version 1.4). Ithaca (NY): The Cornell Lab of Ornithology.
- Bradley JS, Wooler RD, Skira IJ, Serventy DL. 1990. The influence of mate retention and divorce upon reproductive success in short-tailed shearwaters Puffinus tenuirostris. Journal Animal Ecology. 59:487–496. doi: 10.2307/4876
- Burnham KP, Anderson DR. 2002. Model selection and inference: a practical information-theoretic approach. 2nd ed. New York: Springer-Verlag.
- CIREN. 1999. Estudio Agrológico IX Región. Publicación 122. Santiago de Chile: Centro de Información de Recursos Naturales. p. 326.
- Clark KA, Anderson SH. 1997. Temporal, climatic and lunar factors affecting owl vocalizations of western Wyoming. Journal of Raptor Research. 31(4):358–363.
- Contreras P, González MC. 2007. Respuestas en aves rapaces frente al uso de señuelos acústicos en censos diurnos y nocturnos, en el sur de Chile. Gestión Ambiental. 14:77–83.
- Crozier ML, Seamans ME, Gutiérrez RJ. 2003. Forest owls detected in the central Sierra Nevada. Western Birds. 34:149–156.
- Díaz IA, Sarmiento C, Ulloa L, Moreira R, Navia R, Véliz E, Peña C. 2002. Vertebrados terrestres de la Reserva Nacional Río Clarillo, Chile central: representatividad y conservación. Revista Chilena de Historia Natural. 75(2):433–448. doi: 10.4067/S0716-078X2002000200013
- Elgueta E, Reid S, Pliscoff P, Méndez MA, Núñez J, Smith-Ramírez C. 2006. Catastro de vertebrados terrestres y análisis en seis hábitats presentes en la Reserva Nacional Futaleufú, Provincia de Palena, X Región, Chile. Gayana. 70(2):195–205.
- Ens BJ, Safriel UN, Harris MP. 1993. Divorce in the long-lived and monogamous oystercatcher, Haematopus ostralegus: incompatibility or choosing the better option? Animal Behaviour. 45:1199–1217. doi: 10.1006/anbe.1993.1142
- Figueroa RA, Bravo C, Corales ES, López R, Alvarado S. 2000. Avifauna del Santuario de la Naturaleza Los Huemules de Niblinto, Región del Bío Bío, Chile. Boletín Chileno de Ornitología. 7:2–12.
- Fuller MR, Mosher JA. 1981. Estimating the numbers of terrestrial birds. In: Ralph CJ, Scott JM, editors. Methods of detecting and counting raptors: a review. Kansas: Cooper Ornithological Society; p. 235–246. Studies in Avian Biology 6.
- Goodall J, Johnson A, Philippi RA. 1951. Las aves de Chile: su conocimiento y sus costumbres. 2nd ed. Buenos Aires: Platt Establecimientos Gráficos.
- Hauenstein E, Ramírez C, Latsague M. 1988. Evaluación florística y sinecológica del Monumento Natural Cerro Ñielol (IX Región, Chile). Boletín del Museo Regional de la Araucanía. 3:7–32.
- Hines JE. 2006. PRESENCE2 - Software to estimate patch occupancy and related 615 parameters. USGS-PWRC. [accessed 2017 January 23]. http://www.mbr-pwrc.usgs.gov/software/presence.html.
- Ibarra JT, Altamirano TM, Vergara G, Vermehren A, Verga FH, Martin K. 2015. Historia natural y autoecología del chuncho (Glaucidium nana) en el bosque templado andino de la Araucanía, sur de Chile. Boletín Chileno de Ornitología. 21:29–40.
- Ibarra JT, Gálvez N, Gimona A, Altamirano TA, Rojas I, Hester A, Laker J, Bonacic C. 2012. Rufous-legged owl (Strix rufipes) and austral pygmy-owl (Glaucidium nanum) stand use in a gradient of disrupted and old growth Andean temperate forests, Chile. Studies on Neotropical Fauna and Environment. 47(1):33–40. doi: 10.1080/01650521.2012.665632
- Ibarra JT, Martin K. 2015. Beyond species richness: an empirical test of top predators as surrogates for functional diversity and endemism. Ecosphere. 6(8):art142. doi:10.1890/ES15-00207.1.
- Ibarra JT, Martin K, Altamirano TA, Vargas FH, Bonacic C. 2014. Factors associated with the detectability of owls in South American temperate forests: implications for nocturnal raptor monitoring. The Journal of Wildlife Management. 78(6):1078–1086. doi: 10.1002/jwmg.740
- Ibarra JT, Martin K, Drever MC, Vergara G. 2014. Occurrence patterns and niche relationships of sympatric owls in South American temperate forests: a multi-scale approach. Forest Ecology and Management. 331:281–291. doi: 10.1016/j.foreco.2014.08.027
- Inzunza J. 2003. Climas de Chile: Meteorología Descriptiva. Santiago de Chile Capítulo 15:3: Universidad de Concepción.
- Jaksic FM, Iriarte JA, Jiménez JE. 2002. The raptors of Torres del Paine National Park, Chile: biodiversity and conservation. Revista Chilena de Historia Natural. 75(2):449–461. doi: 10.4067/S0716-078X2002000200014
- Jaksic FM, Silva-Aranguiz E, Silva SI. 2004. Historia Natural del Parque Nacional Fray Jorge. La Serena, Chile. In: Squeo FA, Gutiérrez JR, Hernández LR, editors. Fauna del Parque Nacional Bosque Fray Jorge: una revisión bibliográfica. La Serena: Ediciones Universidad de La Serena; p. 93–114.
- Jiménez JE, Jaksic FM. 1989. Biology of the austral pygmy-owl. Wilson Bulletin. 101:377–389.
- Johnson RR, Brown BT, Haight LT, Simpson JM. 1981. Estimating the numbers of terrestrial birds. In: Ralph CJ, Scott JM, editors. Playback recordings as a special avian censusing technique. Kansas: Cooper Ornithological Society; p. 68–75. Studies in Avian Biology 6.
- Koeppen W. 1936. Handbuch der Klimatologie. In: Koeppen W, Geiger R, editors. Das geographsiche system der climate. Berlin: Verlag von Gebrüder, Borntraeger; p. 5–44.
- König C, Weick F. 2008. Owls of the world. 2nd ed. London: Christopher Helm.
- König C, Weick F, Becking JH. 1999. Owls: a guide to the owls of the world. New Haven (CT): Yale University Press.
- MacKenzie DI, Nichols JD, Hines JE, Knutson MG, Franklin AB. 2003. Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology. 84:2200–2207. doi: 10.1890/02-3090
- MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey LA, Hines JE. 2006. Occupancy modeling and estimation. San Diego (CA): Elsevier.
- Marks JS, Cannings RJ, Mikkola H. 1999. Family Strigidae (Typical owls). In: del Hoyo J, Elliott A, Sargatal J, editors. Handbook of the birds of the world. Volume 5. Barn owls to hummingbirds. Barcelona: Lynx Edicions; p. 76–242.
- Martínez DR. 2005. Historia, Biodiversidad y Ecología de los Bosques Costeros de Chile. In: Smith-Ramirez C, Armesto JJ, Valdovinos C, editors. El concón (Strix rufipes) y su hábitat en los bosques templados australes. Santiago de Chile: Editorial Universitaria; p. 477–482.
- Martínez DR, Jaksic FM. 1996. Habitat, relative abudance, and diet of rufous-legged owls (Strix rufipes King) in temperate forest remnants of southern Chile. Ecoscience. 3(3):259–263. doi: 10.1080/11956860.1996.11682340
- Martínez JA, Zuberogoitia I. 2002. Factors affecting the vocal behavior of eagle owls Bubo bubo: effects of sex and territorial status. Ardeola. 49(1):1–9.
- McNicholl MK. 1978. A census of screech owls (Otus asio) using tape-recorded calls. Jack-Pine Warbler. 52:99–101.
- Mella JE. 2005. Cambios estacionales en la avifauna del Monumento Natural El Morado, cordillera de Santiago. Boletín Chileno de Ornitología. 11:2–10.
- Mosher JA, Fuller MR, Kopeny M. 1990. Surveying woodland raptors by broadcast of conespecific vocalizations. Journal of Field Ornithology. 61(4):453–461.
- Muñoz-Pedreros A, Norambuena HV. 2011. Dos siglos de conocimiento sobre las aves rapaces de Chile (1810–2010). Gestión Ambiental. 21:69–93.
- Navarro J, Minguez E, García D, Villacorta C, Botella F, Sánchez-Zapata JA, Carrete M, Giménez A. 2005. Differential effectiveness of playbacks for little owls (Athene noctua) surveys before and after sunset. Journal of Raptor Research. 39(4):454–457.
- Norambuena HV, Muñoz-Pedreros A. 2012. Diurnal activity of the austral pygmy owl (Glaucidium nana) in southern Chile. The Wilson Journal of Ornithology. 124(3):633–635. doi: 10.1676/11-118.1
- Norambuena HV, Raimilla V. 2009. Monumento Natural Cerro Ñielol, refugio de aves a un costado de la ciudad de Temuco. La Chiricoca. 8:50–58.
- O’Donnell RP. 2004. Effects of environmental conditions on owl responses to broadcast calls. Transactions of the Western Section on the Wildlife Society. 40:101–106.
- Perrins CM, McCleery RH. 1985. The effect of age and pair bond on the breeding success of great tits Parus major. Ibis. 127:306–315. doi: 10.1111/j.1474-919X.1985.tb05072.x
- Proudfoot GA, Beasom SL. 1996. Responsiveness of cactus ferruginous pygmy-owls to broadcasted conspecific calls. Wildlife Society Bulletin. 24:294–297.
- Raimilla V, Rau JR, Muñoz-Pedreros A. 2012. Estado del arte del conocimiento de las aves rapaces de Chile: situación actual y proyecciones futuras. Revista Chilena de Historia Natural. 85:469–480. doi: 10.4067/S0716-078X2012000400009
- Ritchison G, Cavanagh PM, Belthoff JR, Sparks EJ. 1988. The singing behavior of eastern screech-owls: seasonal timing and response to playback of conspecific song. Condor. 90:648–652. doi: 10.2307/1368354
- Schieck J. 1997. Biased detection of bird vocalizations affects comparisons of bird abundance among forested habitats. Condor. 99:179–190. doi: 10.2307/1370236
- Smith DG, McKay B. 1984. The effects of weather and effort on Christmas bird counts of owls in Connecticut. American Birds. 38:383–387.
- Stewart AC, Campbell RW, Dickin S. 1996. Use of dawn vocalizations for detecting breeding Cooper’s hawks in an urban environment. Wildlife Society Bulletin. 24:291–293.
- Trejo A, Beaudoin F, Ojeda V. 2011. Response of rufous-legged owls to broadcast of conspecific calls in southern temperate forest of Argentina. Journal of Raptor Research. 45(3):267–271. doi: 10.3356/JRR-10-102.1
- Trejo A, Figueroa RA, Alvarado S. 2006. Forest-specialist raptors of the temperate forest of southern South America: a review. Revista Brasileira de Ornitologia. 14:317–220.
- Tremain SB, Swiston KA, Mennill DJ. 2008. Seasonal variation in acoustic signals of pileated woodpeckers. The Wilson Journal of Ornithology. 120(3):499–504. doi: 10.1676/07-137.1
- Zembal R, Massey BW. 1987. Seasonality of vocalizations by light-footed clapper rails. Journal of Field Ornithology. 58(1):41–48.