Abstract
Archaeozoological data from two New Zealand regions, Greater Hauraki in northern North Island and Otago-Catlins in southern South Island, are examined to document exploitation of the marine environment by Maori prior to European settlement. Data from 107 reliably dated archaeological assemblages are summarized to show the range of shellfish, finfish, marine bird and marine mammal taxa that were harvested and the relative importance of species within each of these classes. Regional differences in faunal spectra are detected and shown to be attributable chiefly to geographic variations in availability of taxa. Changes over time are apparent in the northern region, but are much less marked in the south. Marine mammals and birds disappear or decline in abundance, with human predation the most likely cause. Changes in the composition of shellfish and finfish harvests reflect changes in the location and organization of human settlements, driven largely by expanding demand for land suitable for horticulture.
Introduction
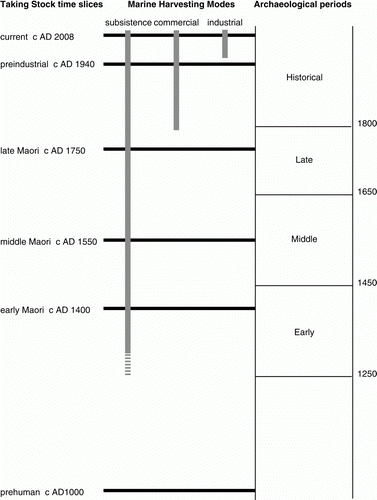
This paper summarizes archaeological data on human use of marine resources in two New Zealand study areas. The purpose is to assist in understanding the nature and causes of changes in coastal and marine shelf ecosystems over the timescale of human occupation. It is part of a broader collaborative project that is attempting to use knowledge of the past to improve future fisheries management. Led by Alison MacDiarmid of NIWA, the scope of this project is summarized by its title—Taking Stock: Long-term effects of climate variation and human impacts on the structure and functioning of New Zealand marine shelf ecosystems. This sets out to build a mass balance model of current coastal and marine shelf ecosystems in two study areas, and then estimate how it might have operated during five earlier time slices: c. 60 years ago (before modern industrial fishing), 250 years ago (before European whaling and sealing,), 450 years ago (about the middle of the Maori period of occupation), 600 years ago (soon after Maori arrival in New Zealand) and 1000 years ago (before human settlement) (). For each of the earlier periods reconstruction depends in part on estimation of the taxonomic composition and biomass of removals from the marine environment through human activities, alongside evidence for environmental changes likely to have altered marine productivity. Archaeological data provide the main source of evidence for estimating what people harvested from the sea prior to the emergence of written records.
Figure 1 Time slices examined in Taking Stock project in relation to marine harvesting modes and archaeological periods.
There is a long history of using archaeological data to infer changes in past ecosystems. Numerous instances of animal extinctions and distributional changes have been detected, dated and examined through archaeological research (e.g. Grayson Citation2001; Rick & Erlandson Citation2008). In New Zealand, as elsewhere in the world, most of these examples concern terrestrial fauna, and in many cases predation by people and their commensals or human-induced habitat modification has been implicated (Anderson Citation1989, Citation1997a; McGlone Citation1989; Holdaway Citation1999; Worthy Citation1999).
For the marine environment, evidence is much more equivocal. New Zealand seals provide one well-explored case of pre-industrial human impacts (Smith Citation1989, Citation2005), and internationally there are others for terrestrial-breeding marine mammals (Bryden et al. Citation1999; Burton et al. Citation2001). Equally, there are cases of apparently stable, long-term exploitative relationships (Etnier Citation2007), and for marine-breeding animals there is little undisputed evidence of dramatic human impact before the emergence of commercial whaling in the eighteenth and nineteenth centuries (Reeves & Smith Citation2006) and more recent industrial-scale fisheries (Pauly et al. Citation1998; Myers & Worm Citation2003). Indeed, the New Zealand data for pre-European shell and fin fisheries show that it is difficult to separate potential effects of human predation and climate without intensive and closely targeted archaeological research (Leach Citation2006).
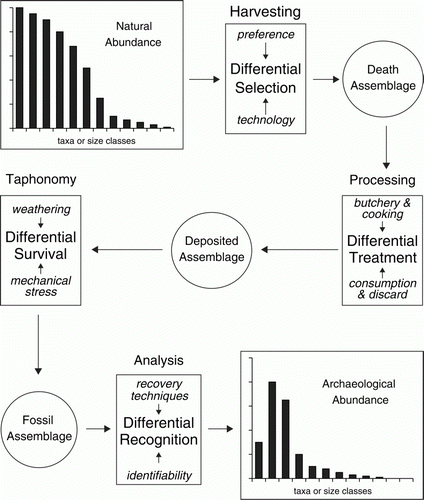
Direct estimation of animal population biomass from archaeological data is not generally possible. The archaeological window into past ecological systems is blurred by transformative processes that influence the creation of the archaeological record. These include harvesting and carcass processing, which are largely determined by cultural patterns, along with natural taphonomic processes of decay, and variations in the accuracy with which different items are amenable to archaeozoological analysis (). Furthermore archaeologists are primarily concerned with determining long-term patterns and regularities in human behaviour, rather than reconstructing past ecosystems, and their data acquisition and analytical methods are designed accordingly. In order to be useful in palaeoecological reconstruction, archaeozoological data must be interpreted with due regard to the cultural and natural formation processes that shaped them, and the archaeological filters through which they have passed.
Figure 2 Transformation processes that potentially modify the archaeological reflection of abundance patterns in the natural world.
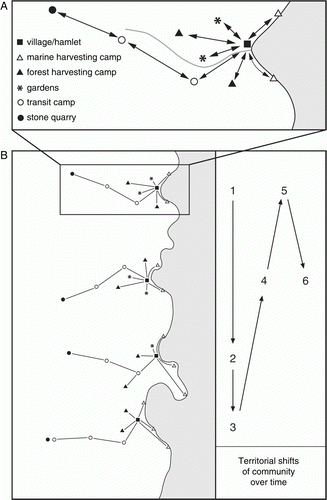
In the case of pre-European New Zealand, one of the key cultural factors that must be accommodated is the mobile nature of human settlement, whereby members of a community are hypothesized (Anderson & Smith Citation1996b; Walter et al. Citation2006) to have made regular intra-annual shifts of residence to facilitate the exploitation of dispersed, seasonally available resources (A), and communities made occasional territorial shifts over time (B). Thus no single site can be considered to provide a complete picture of the pattern of marine resource exploitation by a community, and some sites may represent multiple phases of exploitation with differing return intervals. In these circumstances it is essential to aggregate data at a regional level from a judiciously selected range of sites.
Figure 3 Model of prehistoric settlement patterns in New Zealand. A, Inter-relation of functionally discrete sites utilized by a community. B, Territorial shifts of a community over time.
With these cautions in mind, some inferences about palaeoecology are possible. The presence of physical remains of an animal species in a regional set of archaeological sites can generally be used to infer that this species occurred in the catchment area of those sites at the time of their occupation, and thus provides a basis for reconstructing the distribution of that species in the past. Similarly, where age or sex can be determined from physical remains, the presence of animals of specific age or sex classes allows some inferences to be made about the age composition and breeding status of exploited populations. Potential confounding factors include long-distance transportation of preserved food remains, industrial usage of bones, teeth or shells from distant sources or older archaeological deposits, and disturbance of archaeological deposits introducing taxa from earlier or later time periods. Where recovery and analytical procedures are adequate, problems of this kind can usually be identified and ameliorated. In contrast, the absence of a species, age or sex class in the archaeological record is not so clear cut. Cultural factors such as dietary preference and harvesting technology, or analytical factors such as sample size, may have intervened. These must be accounted for before archaeological absences can be used to infer lacunae in past animal distributions.
Considerable caution is required in making inferences about the abundance of various species in the past from archaeological data. The relative abundance of taxa in archaeozoological assemblages is primarily a record of the frequency with which they were harvested, modified over time by taphonomic decay. Nonetheless it is reasonable to infer that species which are regularly represented in high frequencies in a regional sample of sites were relatively commonly available. Furthermore, where there is a significant decline in the relative abundance of a species over time, without any evidence for changes in harvesting technology, a decline in their availability can be inferred.
Two broad approaches were taken to generate information useful to the Taking Stock project. First, an overview of marine resource utilization was constructed for each study area based upon the presence/absence of marine taxa in archaeozoological assemblages and, where suitable data were available, their relative abundance. These data are summarized in the present paper. The second approach involved estimating the magnitude of marine biomass removals through human exploitation in each study area. This was undertaken using estimates of the size of human populations, their energy requirements and the relative contributions of marine foods to their diet. The latter component of the project will be reported elsewhere.
Study areas, sites and time frames
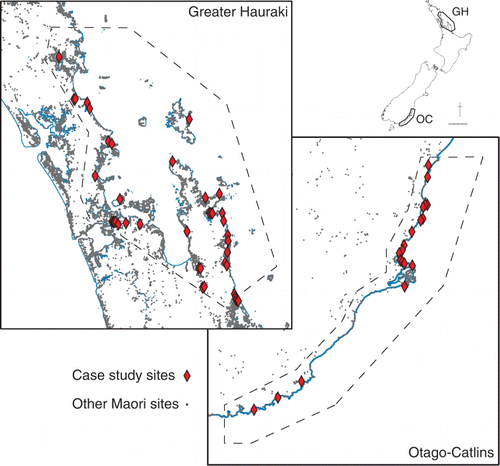
The study areas for the Taking Stock project were selected to best utilize available data on past and present marine shelf ecology, human impacts and climate. The Greater Hauraki study area extends from just south of Whananaki to Waihi Beach on the east coast of the North Island, while the Otago-Catlins study area extends from just north of Oamaru to Slope Point on the east coast of the South Island (). While the marine shelf environments that are the focus of the Taking Stock project extend from mean high water out to a depth of 200 m, it is the archaeological sites on the adjacent coasts that are of concern here. Both study areas have large numbers of recorded pre-European sites, although there are significantly more in the Greater Hauraki area, reflecting the marked concentration of Maori population in the northern third of the country. However, only a small proportion of these sites have been investigated by archaeological excavation.
Figure 4 Greater Hauraki and Otago-Catlins study areas showing location of case study sites and other recorded archaeological sites of presumed pre-European age.
A thorough review was undertaken of information available about excavated sites in each study area, using published literature, theses and dissertations in archaeology from University of Auckland and University of Otago, and the NZ Historic Places Trust's Archaeological Report Digital Library. Sites were selected for detailed analysis on the basis of two criteria: the availability of data on fauna suitable for the methodology described below; and the availability of reliable chronological information (rigorously scrutinized radiocarbon datasets) enabling the sites, or specific assemblages from them, to be placed securely in time. The latter is of particular importance as many previously reported summaries of archaeological fauna relied upon dates that are now regarded as inadmissible (Smith Citation2008:369), and did not have access to more recent, reliable age determinations. In the present context the primary concern was to identify assemblages relevant to Taking Stock's c. AD 1400, c. AD 1550 and c. AD 1750 time slices. With the need to compile data from a broad range of sites in each region, and in view of the uncertainties inherent in radiocarbon dating, data were compiled in terms of three broader period designations: Early (c. AD 1250–1450), Middle (AD 1450–1650), and Late (AD 1650–1800) (). Assemblages were assigned to periods using protocols described in detail elsewhere (Smith Citation2010). In practice the radiocarbon data for almost a third (32%) of the assemblages overlap the period boundaries, and these were classified as either Early/Middle or Middle/Late. Although they do not represent discrete time spans, these designations usefully group assemblages that cluster in age around the arbitrary boundaries, and for the general descriptive purposes of the present paper are used as if they are discrete periods. The abbreviations E, EM, M, ML and L are used for the period names.
For the Greater Hauraki area a total of 75 assemblages from 48 sites were analysed, and 32 assemblages from 19 sites in the Otago-Catlins area. The temporal distribution of the assemblages is summarized in , and the spatial distribution of the sites in . Full details of these sites and their chronology are presented elsewhere (Smith & James-Lee Citation2010:tables 1, 2, appendix 1).
Table 1 Frequency of study assemblages by period.
Method
Data on faunal remains from each of the study assemblages was extracted from published and unpublished sources, and is presented in detail elsewhere (Smith & James-Lee Citation2010:appendices 2–9). Where assemblages had been re-examined (e.g. Millener Citation1981; Leach & Boocock Citation1993; Worthy Citation1998a) revised identifications were incorporated. Reported taxonomic identifications were checked and where necessary updated in the light of revisions of nomenclature, based on the following sources: for shellfish, Spencer et al. (Citation2009); fish, Froese and Pauly (Citation2010); birds, Checklist Committee (OSNZ) (Citation2010); and mammals, King (Citation1995) and Baker et al. (Citation2010). Summaries of data were compiled for four broad classes of fauna: shellfish, finfish, coastal birds and marine mammals. For the purposes of ecosystem modelling in the Taking Stock project, coastal birds include both marine species and shorebirds that are significant feeders in tidal or marine environments during any part of the year.
At their simplest, the summaries of data involved recording the number of assemblages in which each species was represented and, in order to assess changes over time, the proportion of assemblages that they occurred within each time period. For marine birds and seals information was also compiled on the presence of age or sex classes that could indicate the former presence of breeding populations. For most assemblages data were also available on the frequency with which each taxon was represented. While NISP (number of identified specimens) is often preferred for inter-assemblage comparisons (Lyman Citation2008), the only measure available for most of the assemblages was the minimum number of individuals (MNI): the smallest number of individual animals necessary to account for all the remains of a taxon in an archaeological assemblage (Reitz & Wing Citation2008). These data were used to determine which species regularly made up a significant proportion of the animals harvested within each faunal class. However, to avoid the exaggerating effects of small sample sizes (Grayson Citation1978), this procedure was only undertaken with larger data sets (MNI≥10). Where appropriate, two measures of taxonomic composition were also calculated from the larger datasets. Species richness was assessed using Margalef's Index, D Mg (Magurran Citation2004:76–77), which adjusts the number of taxa in an assemblage to control for sample size. The evenness of species representation was evaluated with Simpson's measure, E 1/D (Magurran Citation2004:115–116, 239–40), which yields values between zero and one, with higher values indicating that individuals are distributed more evenly across all taxa present in an assemblage. Correlations between index values and MNI were examined to determine whether observed differences may reflect no more than variations in sample size, and only those with no significant correlation are reported below. In practice this restricted their use to the analysis of shellfish and finfish assemblages.
Results
Shellfish: Greater Hauraki
Shellfish are virtually ubiquitous in New Zealand coastal sites, and they are represented in all 75 study assemblages from the Greater Hauraki region. A total of 147 taxa are represented, 106 identified to species level, and the remaining 41 less precisely (Smith & James-Lee Citation2010:appendix 2). lists 24 species that occur in 10 or more assemblages, and/or make up≥10% of molluscan MNI in any of the 70 assemblages for which quantified data are available.
Table 2 Most common and abundant shellfish in Greater Hauraki assemblages.
Pipis and cockles were the most frequently exploited species. Although the former occur in a slightly greater number of assemblages, the latter is more often the predominant species. These are also the only two species to overwhelmingly dominate any of the assemblages, with cockles making up ≥ 75% molluscan MNI in 20 assemblages and pipis in 12. Few other species make a significant impact in the molluscan assemblages. Tuatua and cats eyes form a second rank of widely and occasionally heavily exploited species. A third group that includes the mudsnail, scallop, speckled whelk, green-lipped mussel, rock oyster and ostrich foot occurs in between a third and half of all assemblages. Amongst these, only the mudsnail and green-lipped mussel ever make up a significant proportion of MNI. The remaining 15 species listed in are best described as only occasionally exploited.
lists 36 taxa that occur in 25% or more of the assemblages in any one time period, showing the proportion of study assemblages within each period in which they occur. These data suggest that over time a narrower range of taxa were represented. Seven species were widely exploited during the E period, occurring in≥70% of assemblages. Over time the number of species that occur this widely narrows to three in the EM period, and then two in the remaining periods. Confirmation of this observation from presence-absence data in the full suite of assemblages can be found when species richness is calculated for the subset of assemblages with MNI data, which shows a significant lowering of species richness over time (A). While most assemblages (78%) have low richness values (≤2), all but four of the 13 assemblages with higher values are from the E or EM periods. Conversely, all but four of the 28 assemblages with very low richness values (≤1) are from the M, ML or L periods. These variations are not driven by sample size, with no significant correlations between richness and MNI for any of the periods ().
Figure 5 Species richness (D MG) plotted against sample size (MNI) for shellfish assemblages from A, Greater Hauraki and B, Otago-Catlins. The progressive lowering of intercepts in the best-fit regression lines for successive periods in the Greater Hauraki series is highly significant (F = 7.10185, P < 0.0001). There is no significant difference between periods in Otago-Catlins.
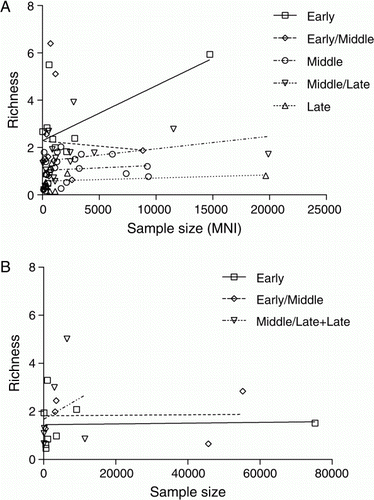
Table 3 Temporal distribution of frequently exploited shellfish in Greater Hauraki assemblages.
Table 4 Correlations of species richness and sample size for shellfish assemblages.
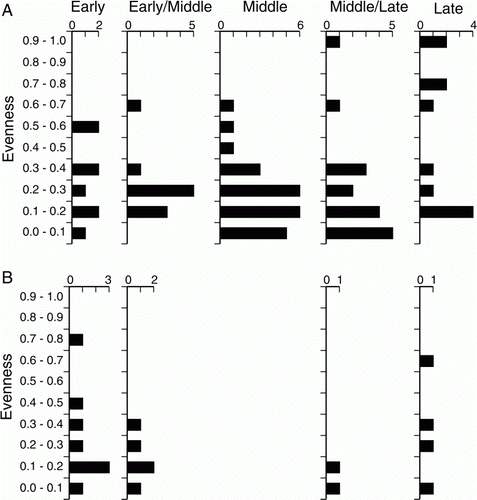
Evenness values are generally low through all periods, reflecting a tendency for assemblages to have one dominant species amongst a range of less abundant taxa (A). Higher values are evident only in the ML and L periods where some assemblages are composed of more or less even numbers of just two taxa, cockle and pipi. One or other of these nearly always predominates in the assemblages with lower evenness values throughout the M, ML and L periods. Prior to that, however, it is taxa from rocky shore habitats that fulfill this role, providing five of the six species that are the most abundant in E period assemblages, and four out of seven in the EM period. This trend is also evident in the presence-absence data () which shows that six of the seven most widely represented species in the E period are rocky shore taxa (cats eye, green-lipped mussel, radiate limpet, black nerita, white rock shell, Cook Strait limpet). In all subsequent periods the predominant species are from estuarine (pipi, cockle) or open sandy beach (tuatua) habitats. While rocky shore species continue to be exploited—they make up between 34% and 45% of all taxa throughout all periods—all but three of the 29 instances in which any one of these makes up≥10% of molluscan MNI are confined to the E and EM periods. Potential explanations for this will be considered below.
Figure 6 Numbers of shellfish assemblages in taxonomic evenness (E 1/D) bands for (A) Greater Hauraki and (B) Otago-Catlins.
Shellfish: Otago-Catlins
As in Greater Hauraki, shellfish are represented in all the study assemblages from the Otago-Catlins region. A total of 90 taxa are represented, 68 identified to species level and the remaining 22 less precisely (Smith & James-Lee Citation2010:appendix 6). However, the majority of these are neither widely represented nor abundant. Only 19 taxa are represented in five or more of the study assemblages, and include all those that make up ≥ 10% of molluscan MNI in the 22 assemblages for which these data are available (). Taxonomic composition indices were calculated for 19 assemblages with MNI≥20, and disclosed no significant correlation between MNI and species richness ().
Table 5 Most common and abundant shellfish in Otago-Catlins assemblages.
Five species stand out as the most common and abundant. Blue mussels, cockles, and pipis were clearly the most frequently and heavily exploited species. Cats eyes were as widely represented, but much less often in high numbers, while mudsnails occurred in slightly fewer assemblages but quite commonly in high numbers. It should also be noted that the predominance of blue mussels may be even greater than these data suggest as the poor survival of their shells in archaeological sites generally leads to underestimation of their abundance in MNI counts.
Paua, Cooks turban, and green-lipped mussels are slightly less widely represented and seldom very common in the molluscan assemblages, suggesting lower rates of harvesting. Again, differences in archaeological survival may be a factor, with Cooks turban and paua particularly difficult to quantify accurately, and therefore probably under-represented. It will also be noted that a moderately large number of assemblages have Haliotis remains that cannot be positively assigned to species. At least one of limpets of the Cellana genus is probably equally widely represented, but in the majority of assemblages it was not possible to identify these to species. The mud oyster, shield limpet, spotted top shell, ribbed mussel, tuatua, a whelk and the rock shell Haustrum lacunosum form a third group of moderately well represented but seldom abundant species.
There is conflicting evidence as to whether there was change over time. Among the assemblages for which it was calculated, species richness values range from low to moderate at all periods, with only one high value, from an ML period assemblage (B). Likewise there is no obvious shift in taxonomic evenness, with a range of low to moderate values for most assemblages from all periods (B). However, a trend is apparent when presence-absence data for the full sample are considered (). For this purpose the M and ML periods, each represented by only two assemblages, are excluded from consideration. The E and EM periods exhibit a broadly consistent pattern with the same six or seven species occurring in more than half of all assemblages in each period. However, in the L period the two most widely represented species occur in only 56% of the assemblages, with another two in 44%. This appears to indicate a shift in shellfishing practice towards a narrower range of species at any one locality, but variation between localities in which species were targeted. With this apparent change in shellfishing strategy it is difficult to assess any changes at the species level, although it can be noted that all of the assemblages in which cockles make up ≥ 25% of molluscan MNI are confined to the E and EM periods, while all but one of those dominated by blue mussels are from the EM, ML or L periods.
Table 6 Temporal distribution of frequently exploited shellfish taxa in Otago-Catlins assemblages.
Finfish: Greater Hauraki
Finfish occur in 56 of the 75 Greater Hauraki study assemblages (Smith & James-Lee Citation2010:appendix 3). They are absent in two from the EM period, seven from the the M period, three from the ML period and four from the L period. A total of 35 taxa have been identified (). It should be noted that some of these are family or higher-level groupings necessitated by the nature of archaeological data: it is virtually impossible to identify the various species of Labridae on the basis of skeletal morphology; Carangidae are often difficult to distinguish; and the poor survival of elasmobranch remains frequently precludes specific identification.
Table 7 Frequency and abundance finfish in Greater Hauraki assemblages.
Snapper were clearly the main focus of fishing activity, occurring in 75% of all assemblages, and dominating finfish MNI counts in more assemblages than any other species. The next most widespread species, kahawai, occurs in less than half as many assemblages and is abundant in only two. Wrasses and barracouta are moderately widely represented, but again only rarely abundant. But for difficulties in identification, one or more species of mackerel may have had similar representation. The only other species of note is the leatherjacket, which, although not widespread in occurrence, made a significant contribution to finfish MNI in more assemblages than any species other than snapper.
The finfish assemblages disclose several indicators of change over time (). The proportion of assemblages in which finfish occur (including those not identified to species) declines from 100% in the E and EM periods to 76% in the M period and 77% in the L period. Reasons for this will be considered in the Discussion. Throughout all periods snapper remain the most frequently represented species, although their predominance is markedly lower during the M and ML periods. Wrasses, kahawai and leatherjacket were all widely represented in the E period, but never again occur so prominently. This decline is particularly marked for leatherjackets, with all the assemblages in which they made up ≥ 10% finfish MNI confined to the E and EM periods. The majority of other species also decline in frequency of representation after the Early period, suggesting that there may have been a narrowing of the taxonomic range in the finfish catch over time. However, this is not supported by the analysis of species richness (, A), although it should be noted that quantified data of suitable sample size were available for less than half of the assemblages, with none from the L period, so some caution must remain.
Figure 7 Species richness (D MG) plotted against sample size (MNI) for finfish assemblages from (A) Greater Hauraki and (B) Otago-Catlins. The progressive lowering of intercepts in the best-fit regression lines for successive periods in the Otago-Catlins series is highly significant (F = 5.81369, P=0.01192). There is no significant difference between periods in Greater-Hauraki.
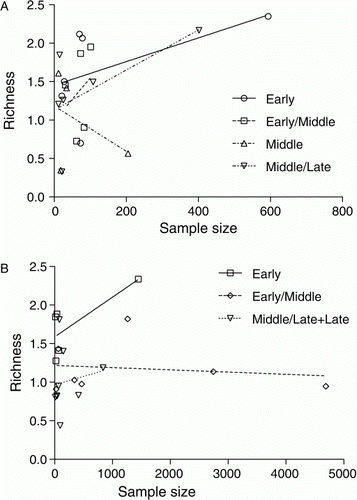
Table 8 Temporal distribution of finfish in Greater Hauraki assemblages.
Table 9 Correlations of species richness and sample size for finfish assemblages.
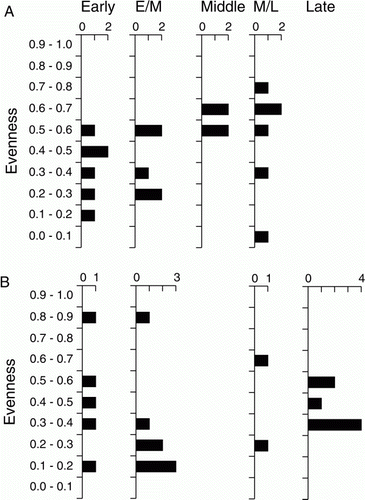
Another potential indicator of change is that eight of the 11 highest evenness values are from the the M and ML periods, indicating that fish catches of these periods were less likely to be dominated by a single species (A). However, this might be a product of identification difficulties, which may have disguised an increase in mackerel fishing during the M and ML periods. Because the bones of jack mackerel and horse mackerel can be difficult to distinguish they are often identified as Trachurus ?sp, and it is likely that most of those identifed as Carangidae ?sp also derive from one of these species. When the presence-absence data for these taxa are summed, they suggest mackerel were represented at 25% of sites in the E period and 33% in the EM period, rising to 67% in the M period and 53% in the ML period, before falling again to 11% in the L period.
Figure 8 Numbers of finfish assemblages in taxonomic evenness (E 1/D) bands for (A) Greater Hauraki and (B) Otago-Catlins.
Finfish: Otago-Catlins
Finfish remains occur almost as frequently as shellfish in Otago-Catlins sites, occuring in all but two of the 32 study assemblages (Smith & James-Lee Citation2010:appendix 7). They are reported as absent from an L period assemblage at Katiki Beach, and from the E period in the lowest layer of the TT1 excavation at Papatowai. In the latter case, at least, this is clearly a product of archaeological sampling, as other E period assemblages from the same site do contain fish remains.
A total of 32 taxa have been identified (), with some only to family or taxa above species for reasons outlined earlier. Six taxa stand out as the main focus of Maori fishing in the Otago-Catlins region. Barracouta are represented in every assemblage, and make up more than 50% on finfish MNI in over half of those with assemblages large enough for meaningful analysis. Red cod are almost as widely represented, frequently second to barracouta in MNI counts, and outnumber them in a small number of assemblages. Ling, hapuku, wrasses and blue cod are the only other taxa to occur in more than half of the study assemblages, and occasionally make up significant proportions of total finfish numbers. Of the less widely represented taxa, only the black cods ever figure strongly in MNI counts. Both the assemblages where this occurs are from the same site (Ross's Rocks, East Otago), suggesting that it represents a localized pattern of either species abundance or fishing practice.
Table 10 Frequency and abundance finfish in Otago-Catlins assemblages.
There are indications of both stability and change over time in the Otago-Catlins fishery (). The two main species, barracouta and red cod, remain overwhelmingly dominant through all time periods. Among the second rank of species there are apparent declines in the frequency of exploitation for blue cod, trumpeter and black cods, especially if the inadequate sampling from M and ML period sites is set to one side. In contrast to this, however, ling and hapuku appear to occur more often in sites after the E period. At a broader level, there are clear signs that the range of fish exploited declined over time. Of the 22 taxa represented in E period assemblages, six are not represented again, and another four occur in only one other time period. Species richness values fall significantly over time (, B), while evenness at the high and low extremes seen in some E and EM period assemblages is no longer evident by the L period (B). Once again, reasons for this will be considered below.
Table 11 Temporal distribution of finfish in Otago-Catlins assemblages.
Coastal birds: Greater Hauraki
Birds are relatively scarce in the Greater Hauraki study assemblages, occurring in only 24 of the 75 under analysis (Smith & James-Lee Citation2010:appendix 4). Moas and smaller birds from terrestrial and wetland habitats occur in 16 of these, and coastal birds in 17. Where quantified data are available, coastal birds make up between 16% and 100% of MNI from small birds (i.e. other than moa). These are predominantly small assemblages—only four have coastal bird MNI ≥ 10, with the largest at 94—which limits the certainty with which patterns of abundance can be inferred and precludes meaningful analysis of taxonomic composition indices.
A total of 22 coastal bird species have been identified (). It should be noted that it is often difficult to achieve positive identifications to species level from archaeological remains of birds, and in the present dataset another five less certain taxonomic categories are represented. The little penguin is the most widely represented species, with the spotted shag the only other to occur in a moderately large number of sites. These species, along with the fluttering shearwater, black-backed gull, sooty shearwater, common diving petrel and pied shag, are the only ones that ever form a large proportion of coastal bird MNI, although as already noted the small size of most assemblages limits confidence in these data. Furthermore shag, shearwater and petrel bones that have not been positively identified to species occur in a moderately large number of assemblages, and sometimes as abundantly as the positively identified taxa, suggesting that some of the latter may be under-represented. Eighteen of the positively recorded species are thought to have been breeding in northern New Zealand at the time of first human arrival (Holdaway et al. Citation2001). For five of these, bones of immature animals have been reported from at least one study assemblage, indicating exploitation of breeding sites. The four remaining species are all recorded seasonal or occasional visitors to coasts of the Greater Hauraki region.
Table 12 Frequency, abundance and breeding status of coastal birds in Greater Hauraki assemblages.
There is strong evidence for change over time. The proportion of assemblages in which coastal birds are represented declines from 100% in the E period to 45% of the EM sites. Coastal birds are totally absent in the M period, and then occur in 11% of ML sites and 15% of L sites (). Further to this, the majority of coastal birds in both ML and L sites are represented by single bones, raising the possibility that they were raw materials collected for artefact manufacture, perhaps from beach wrecks, rather than remains of animals hunted for food. It is also noteworthy that apart from one example of a spotted shag from an ML site, all of the immature bird bones reported for the study sites derive from the E period.
Table 13 Temporal distribution of coastal birds in Greater Hauraki assemblages.
Coastal birds: Otago-Catlins
Birds are present in all but one of the Otago-Catlins assemblages (Smith & James-Lee Citation2010:appendix 8). These include moas in 18 assemblages (along with another five where their bones are present as industrial raw materials), smaller terrestrial and wetland birds in 26 assemblages, and coastal birds in all 31. The latter make up 50% or more of total small bird MNI in two-thirds of the 27 assemblages for which quantified data are available.
A total of 30 coastal bird species have been identified from the Otago-Catlins study assemblages (). All yellow-eyed penguin identifications are listed here as Megadyptes sp. as virtually all were made prior to recent separation of an extinct smaller species (M. waitaha) from the larger form (M. antipodes) now resident in the study area (Bossenkool et al. Citation2009). Not listed in are any of the reported identifications only to taxa above species level. Careful scrutiny of these suggests that in all cases they are likely to derive from species already positively identified within the assemblage concerned. However, it is likely that frequency counts underestimate the abundance of some taxa.
Table 14 Frequency, abundance and breeding status of coastal birds in Otago-Catlins assemblages.
In contrast to what was observed with shellfish, finfish and Greater Hauraki birds, none of the Otago-Catlins coastal bird assemblages is ever dominated by one species that contributes more than 50% of total category MNI, indicating that birding here tended to be a broad-spectrum rather than closely targeted pursuit. Nonetheless the spotted shag stands out as the most widely represented species and the one which most often occurs in moderately large numbers in assemblages. A second group of species that are widely represented, but less often abundant in sites, includes the white-capped albatross, Stewart Island shag, little blue, crested and yellow-eyed penguins, and the common diving petrel. The only other species that ever form a significant part of the study assemblages are two shearwaters (sooty, fluttering), and the pied shag. Nine of these 10 species are thought to have maintained breeding populations in southern New Zealand at the time of human arrival (Holdaway et al. Citation2001). In the case of the tenth, the presence of osteologically immature bones of white-capped albatross at two sites supports the suggestion (Holdaway et al. Citation2001) that this species was also breeding in the region. Remaining species have all been recorded as seasonal or occasional visitors to Otago-Catlins coasts.
There are some fluctuations in the frequency with which species occur in each time period (). Of the seven species represented in 50% or more of E sites, five show evidence for decline. Crested, little blue and yellow-eyed penguins along with sooty shearwaters and fairy prions are all represented in lower proportions of later assemblages (leaving aside the poorly sampled M and ML periods). Indeed if all three penguins are combined as a group they decline from occurrence in 90% of the E period assemblages, through 78% in the EM period to just 33% in the L period. Likewise petrels, prions and shearwaters as a group occur in 80% of E period assemblages, falling to 67% in the EM and 44% in the L period, with both the sooty shearwater and fairy prion absent from the later. In contrast to this, shags as a group, and the spotted and Stewart Island shags as their main representatives, show little change over time, while the albatross/mollymawk group of species occur more frequently in the L period than in earlier times.
Table 15 Temporal distribution of coastal birds in Otago-Catlins assemblages.
Marine mammals: Greater Hauraki
Mammals are represented in 27 of the 75 Greater Hauraki study assemblages (Smith & James-Lee Citation2010:appendix 5), with terrestrial mammals (dog, rat) occurring in all but one of these. Marine mammals are confined to just 17 assemblages, although in four cases their remains are almost certainly raw materials for artefact manufacture and may not represent animals caught at that time or place (). All of the primary remains are from E or EM period sites. The fur seal is the most widely represented species, followed reasonably closely by the sea lion, with all the specimens identified only to the otariid family deriving from one or other of these two species. These data are consistent with previous observations that there were once breeding populations of fur seals and sea lions in the Greater Hauraki region, and that these disappeared by c. 1500 A.D. (Smith Citation1989, Citation2005). Elephant seal remains also occur only in the E period.
Table 16 Frequency and temporal distribution of marine mammals in Greater Hauraki assemblages.
Pilot whales were almost certainly exploited only by scavenging from naturally stranded animals. Notable by their absence in any of the study assemblages are dolphins. Dolphin ?sp bones have been reported from at least one undated site in the Greater Hauraki region and others further north on the east coast of Northland (Smith Citation1985, Citation1989; Furey Citation2002a). In addition, bone harpoon heads thought to have been used in dolphin hunting are also known from the region (Smith Citation1989), making it likely that these animals were at least an occasional part of the marine harvest.
Marine mammals: Otago-Catlins
Mammals are represented in all of the Otago-Catlins study assemblages except for one from the Late period (Smith & James-Lee Citation2010:appendix 9). The remaining 31 assemblages all include terrestrial mammals (dogs, rats), with marine mammals represented in 28. However, each of the assemblages where the latter are missing is from a site that yielded marine mammal remains from one or more other assemblage of the same period, indicating that the apparent absences are sampling errors. Taxonomic identifications have usually been reported for seal remains, although these are not available for two EM period assemblages, but have seldom been reported for cetaceans because of the difficulty of achieving accurate identification.
The fur seal is clearly the most widely represented species and, where marine mammal assemblage sizes are sufficiently large to make comparisons meaningful, they generally make up more than half of marine mammal MNI (). Sea lions and elephant seals occur in about half as many assemblages and seldom make up more than a third of marine mammal MNI. Leopard seals occur rarely, with never more than a single individual represented in an assemblage. Cetacean remains are also relatively uncommon. The only species-level identifications reported are for cranial remains from single individuals of Hectors and common dolphins from one Late period assemblage. On the basis of size, it is suspected that remains from other sites include both dolphins and some larger whales.
Table 17 Frequency and temporal distribution of marine mammals in Otago-Catlins assemblages.
Change over time may be indicated by the decline in occurrence, then disappearance from the study assemblages of sea lions, elephant seals and leopard seals by the end of the ML period. More detailed analysis elsewhere (Smith Citation1985, Citation1989, Citation2005) indicates that there were fur seal breeding populations on the Otago-Catlins coast until about the end of the seventeenth century, with only non-breeders present thereafter.
Discussion
The evidence reviewed above indicates that a broad range of marine resources were exploited by the pre-European Maori occupants of the two study areas. Nonetheless these represent just a small proportion of what was available. For example, the 147 molluscan taxa present in the Greater Hauraki sites represent only about 26% of those reported for waters shallower than 50 metres within the study area (Morley & Hayward Citation2009), the 35 fishfish taxa similarly represent about 20% of those available in the northeastern North Island (Paul Citation2000), and from both study areas only three cetacean species have been positively identified out of some 56 that have been reported from New Zealand waters (Baker et al. Citation2010). Several factors are likely to have contributed to this pattern, and it is pertinent to consider which of these were likely to be in operation with each class of fauna in the two study areas, and whether they exhibit any changes over time.
In the case of shellfish, accessibility was important with deepwater shellfish beyond reach of shoreline harvesting. Two types of shoreline were clearly preferred collecting zones, as most species found archaeologically derive from rocky shore or estuarine settings, and relatively few from open sandy beaches. In each of the quantified study assemblages there is an overwhelming focus on species from just one of these zones (), suggesting that harvesting was localized and presumably focused on what was available in close proximity to the human settlements concerned.
Figure 9 Proportions of rocky shore, estuarine and open beach shellfish MNI per assemblage by time period in A, Greater Hauraki and B, Otago-Catlins.
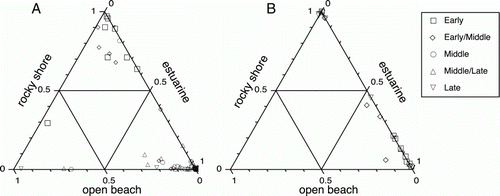
Within each of the two main harvesting zones differing selection processes appear to have been in operation. Relative abundance was significant in estuarine settings, with virtually all harvesting focused on cockles, pipis, and to a lesser extent mudsnails, which often occur in extremely dense beds (Shawcross Citation1967). Size was much more important on the rocky shore, where Anderson (Citation1981) has shown that the optimal collecting strategy involves taking the largest individuals encountered, regardless of species. In the present study six of the nine rocky shore species that occur in 50% or more Greater Hauraki assemblages at any one time period () are from large or moderate size classes (≥ 3 g wet meat weight), as are all the rocky shore taxa that occur with this frequency in Otago-Catlins (). Size may also have been the primary consideration when harvesting on open sandy beaches, as it is only the larger species such as tuatua, scallops and ringed venus that occur with any frequency in the archaeological record.
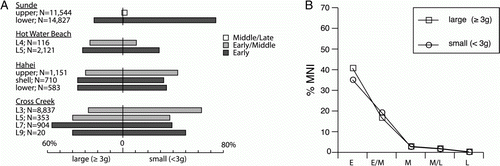
Although shellfish were ubiquitous throughout the prehistoric sequence in both study areas (), there were distinct changes in assemblage composition over time. In Greater Hauraki, E and EM period assemblages are dominated by rocky shore shellfish with estuarine species predominant in later periods, while in Otago-Catlins there appears to have been a transition in the opposite direction (). The former trend has been noted previously, and was suggested to have arisen from harvesting pressure on preferred rocky shore taxa (Green Citation1970). Detailed time-series data available from four Greater Hauraki sites (A) allow some testing of this. Two of these (Cross Creek, Hahei) exhibit the progressive shift in emphasis from larger to smaller species that Anderson's (1981) model predicts as the outcome of sustained harvesting on the rocky shore, but this is not apparent at the two other sites (Hot Water Beach, Sunde), and nor is it evident in summary-level data for the whole region (B). This indicates that while there might have been some localized effects, harvesting pressure cannot explain the changes observed at a regional level. There are no size-frequency data available for estuarine shellfish from the Otago-Catlins sites with which to assess whether there was evidence of harvesting pressure (cf. Swadling Citation1976), but it seems unlikely that this could have been sufficient to lead to cessation of estuarine shellfishing.
Figure 10 Changes over time in proportions of assemblages containing each main class of marine fauna in A, Greater Hauraki and B, Otago-Catlins.
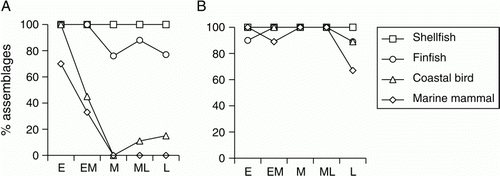
Figure 11 Changes over time in proportions of total shellfish MNI contributed by large and small rocky shore shellfish species in Greater Hauraki. A, Time series from four stratified sites. B, Mean proportions for all assemblages per period.
Given the strongly localized nature of shellfishing identified above, a more likely explanation for the observed changes in each region is that there were shifts in preferred settlement location. For the Greater Hauraki region it can be postulated that growth of the human population in conjunction with the shifting nature of Maori horticulture led to increasing demand for land suitable for horticulture, and it seems likely that there was greater availability of this in the vicinity of estuaries than adjacent to rocky shores. Horticulture was not a factor in Otago-Catlins, as this region was too cold for any of the prehistoric cultigens to grow. Almost all of the ML and L period sites dominated by rocky shore shellfish are on headlands with earthwork fortifications, suggesting that defensibility might have been increasingly important in selecting settlement location. However, with only six quantified assemblages post-dating the M period it is difficult to determine whether the observed patterns are artefacts of sampling rather than real changes in shellfish selection and settlement location.
Maori harvesting of finfish employed both shore-based and boat-based fishing strategies utilizing a technological suite that included a diverse range of hooks, nets and traps along with single- and double-hulled canoes (Leach Citation2006). The archaeological data from Greater Hauraki and Otago-Catlins indicate taxonomic composition of fish catches consistent with patterns identified for broader Northern North Island and Southern South Island data sets in previous reviews (Leach & Boocock Citation1993; Anderson Citation1997b; Leach Citation2006), supporting Anderson's conclusion that the concentration upon a relatively narrow range of medium-sized carnivorous taxa arose from their ready availability in the most easily accessible inshore waters, coupled with reliance upon angling as the predominant harvesting technology. At the same time, the present study suggests that broad regional catch patterns disguise some more localized variation, such as the high relative abundance of leatherjackets in the Coromandel, although it is difficult to determine whether this reflects localized natural abundance or a greater emphasis on fishing with nets (Witter Citation1969; Mann Citation2009).
Both study areas exhibit consistency in the fish that were most commonly caught, with barracouta and red cod predominant throughout the Otago-Catlins sequence, and snapper in Greater Hauraki. Some variations over time are apparent amongst the lesser ranked taxa. In Otago-Catlins there was a significant fall in finfish taxonomic richness, with at least six species not occurring after the E period, and blue cod, trumpeter and black cods caught less often after the EM period. Time series data from strata within the Shag River Mouth site show rapid declines in each of these species, suggesting that they may have been vulnerable to persistent local harvesting (Anderson & Smith Citation1996a). At a region-wide level their place seems to have been taken by hapuku and ling. However, the limited number and size of mid and late sequence assemblages from this study area make it difficult to be confident that these variations in minor taxa are anything other than sampling effects.
In Greater Hauraki we can have greater confidence that apparent changes through time are not the product of sample size, although they may reflect the geographic composition of the data set rather than changes in fishing practice. The mid-sequence rise in prominence of mackerel fishing closely parallels changes in the relative proportions of study assemblages from the Coromandel and Inner Gulf sub-regions. The latter make up only 13% and 27% of assemblages in the E and EM periods, before rising to 76% and 67% in M and ML periods, then falling again to 8% in the L period. This raises the possibility that mackerel were encountered more frequently in the waters of the Inner Gulf than along the eastern shore of the Coromandel, and that this has shaped the overall composition of the regional sample. Reduced representation of Coromandel sites might also have influenced the lower representation of leatherjackets after the EM period. However, this species is absent from all 13 post-EM period Coromandel assemblages with fish identified to species. It is notable that all of these assemblages are dominated by estuarine and/or open sandy beach shellfish, which suggests that it was the movement of settlements away from rocky shore environments that was the main reason for reduced catches of leatherjackets, and perhaps also of wrasses, tarakihi and blue cod.
Changes in settlement pattern may also lie behind the absence of fish from between 12% and 24% of M, ML and L period assemblages in Greater Hauraki (). In some circumstances absence of bones from the archaeological record may be attributed to use of preserved food supplies captured elsewhere, but the drying and/or smoking methods used by Maori to preserve fish did not involve the separation of flesh and bone (Colenso Citation1869; Hammond, cited in Furey Citation1996:74; Anderson Citation1998:120–122), and thus cannot have influenced the pattern observed here. Nor can it have been due to differential survival, as all of the fish-less assemblages contain shellfish which would have neutralized any acidity in the soil likely to compromise bone survival. What is notable is that the fish-less assemblages are composed solely of shellfish, suggesting that they derive from localities occupied for brief periods during which a limited range of activities were undertaken. In several cases the excavators have interpreted these as camps associated with gardening activity (Phillips Citation1997; Furey Citation2002b; Moore Citation2006), although other kinds of specialized activity or short-term occupation may equally be represented. It can be hypothesized that restricted-function sites such as these became a more prominent part of the Greater Hauraki cultural landscape from about the middle of the prehistoric sequence.
There are two ways in which the archaeological record may not accurately reflect past fishing practice. Poor survival of their predominantly cartilaginous skeletons raises the possibility that sharks and rays are under-represented by the small numbers of elements recovered in the study assemblages. This is most likely to be the case in Greater Hauraki, as late eighteenth- and nineteenth-century records indicate that small schooling sharks were an important component of harbour fisheries in Greater Hauraki (Polack Citation1840; Watson Citation1969; Murdoch Citation1996) and some other parts of the North Island (Taylor Citation1855; Colenso Citation1869; Matthews Citation1910). While there are similar records for parts of the South Island (Anderson Citation1997b), these do not include the Otago-Catlins region (Bathgate Citation1992), suggesting that there is likely to have been as much regional variation in this as has been documented for other fisheries. Equally, it would be unwise to presume that the accounts of shark fishing in Greater Hauraki can be projected back in time any further than the Late period without some corroborative evidence.
Eels are also scarcely represented in the archaeological record, despite nineteenth- and twentieth-century accounts of mass capture, preservation and their importance as a food resource (Colenso Citation1891; Beattie Citation1920; Best Citation1929). Although predominantly freshwater fish, Anguillidae are included in the present discussion because part of their life cycle is in the marine environment and it was during their annual downstream migration to sea that mass harvesting took place. It has been argued that their scarcity in New Zealand archaeological sites is due to poor survival of their bones, rather than the absence of large-scale eeling during prehistory (Marshall Citation1987; Paulin Citation2007). In contrast Leach (2006:186–190) has proposed that their scarcity may reflect cultural preference, with eels generally avoided as a food source until the historic era. The abundance of eel bones in sites of considerable antiquity in Britain and Europe (Wheeler & Jones Citation1989:164) and the Pacific (Leach & Ward Citation1981:59; Kirch & Yen Citation1982:292), along with the good preservation conditions for fishbone already noted for the study assemblages, makes it difficult to accept differential survival as an explanation for their scarcity in the dataset under analysis here. What these data indicate is that eels were a minor but consistent part of the fisheries throughout the prehistoric sequence in both study areas, making it unlikely that they were explicitly avoided as a source of food. At the same time there is nothing suggestive of mass harvesting and, following Leach, it is proposed here that this form of exploitation did not emerge until very late in the prehistoric sequence or during the historic era.
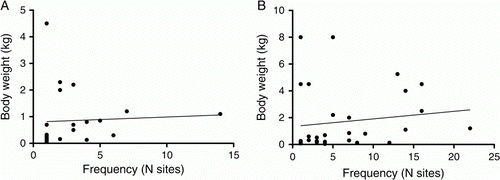
Birds were hunted throughout Polynesia, particularly during the initial colonization phase, using techniques ranging from snaring and spearing to hand harvesting of burrow-nesting and colonial seabirds (Steadman Citation1997). It has been suggested that Maori fowling strategies focused on birds of larger size (e.g. Cassels Citation1984; Holdaway Citation1999) and while this seems to have been the case for terrestrial taxa, the evidence is equivocal for coastal birds, as data from the present study show significant correlation between body size and frequency of occurrence in Greater Hauraki sites, but not in Otago-Catlins (). Worthy's (Worthy Citation1998b) analysis of avifauna from Marfell's Beach has shown that frequencies of culturally deposited taxa closely match those deposited naturally, indicating that, with few exceptions, bird species were hunted as they were encountered, rather than being explicitly targeted. If this observation can be generalized across the country, it would imply that penguins, shags and Procellariidae dominated the coastal bird harvest in Greater Hauraki and Otago-Catlins because they were the most abundant taxa, and perhaps also because their colonial breeding habits made them easily and predictably accessible.
Figure 12 Frequency of occurrence of coastal bird taxa shows significant correlation with mean body weight in A, Greater Hauraki (Spearmans r=0.4306, P=0.0455), but not in B, Otago-Catlins (Spearmans r=0.2213, P=0.2315).
Coastal birds show a dramatic decline by mid-sequence in the Greater Hauraki data set which is not paralleled in Otago-Catlins (). Although not discussed here, terrestrial birds exhibit almost identical trajectories in each region (Smith & James-Lee Citation2010), suggesting that there may have been a rapid and significant reduction in the availability of birds in the north. There have been several nationwide assessments of Holocene avifaunal losses in New Zealand which have disclosed high rates of extinction and extirpation following human arrival, especially amongst flightless terrestrial taxa of large body size (Cassels Citation1984; Holdaway Citation1999; Worthy Citation1999; Worthy & Holdaway Citation2002). Among coastal birds there were only two extinctions—Puffinus spelaeus (Holdaway & Worthy Citation1994) and Megadyptes waitaha (Bossenkool et al. Citation2009)—but many other coastal taxa were extirpated from the main North and South Islands or had their ranges severely reduced (Holdaway Citation1999). While the loss of larger terrestrial taxa has been attributed principally to hunting by Maori, it has been proposed that predation by the Pacific rat (Rattus exulans) played a much larger part in the demise of smaller birds, especially the burrow-nesting petrels, prions and shearwaters (Holdaway Citation1999). Holdaway's analysis did not detect any difference between the North and South Islands in the rate of loss of these birds over the entirety of the prehistoric period. However, it should be noted that his study, along with the other nationwide assessments, did not have the benefit of well-dated assemblages with quantified relative abundance data from within that period. The present data set shows that bird species most likely to be at risk from Pacific rats (body size ≤ 0.5 kg and/or egg length ≤ 60 mm; Holdaway Citation1999) actually increase as a proportion of the coastal avifauna harvested by Maori over time in Greater Hauraki while remaining more or less stable in Otago-Catlins (). This suggests that it is unlikely that the differences between the two regions can be attributed to varying impacts of rat predation, and alternative explanations need to be considered.
Table 18 Proportion of coastal bird harvest vulnerable to rat predation.
Sampling could provide one such alternative, if the later datasets derived only from small excavations yielding restricted faunal assemblages that might have failed to detect comparatively rare taxa. However, the mean size (total MNI) of the 56 M, ML and L assemblages from Greater Hauraki is 2347 (range 4–19,909), which is slightly greater than the 2153 (range 38–15,577) for the 19 E and EM assemblages. If sample size was a contributing factor it would be more likely to have operated in Otago-Catlins, where the 11 later assemblages are much smaller (mean 391; range 11–3194) than the 19 earlier ones (mean 11,176; range 36–77,508). However, as noted, birds are much better represented in later sites in Otago-Catlins than in Greater Hauraki.
Changes in settlement pattern in Greater Hauraki could also have contributed to the observed pattern. Whereas the E and EM period datasets derive from sites that appear to have been hamlets or villages at which a wide range of subsistence activities are represented (Anderson & Smith Citation1996b; Walter et al. Citation2006), the shellfish and finfish data reviewed above suggest that some of the assemblages from subsequent periods derived from functionally specialized short-term camps, each of which would be expected to yield only part of the faunal spectrum harvested by a community. However, the approach taken in this study does not rely upon single sites, instead aggregating data at a regional level, so that there can be reasonable confidence that the full range of subsistence pursuits undertaken during the M, ML and L periods are represented among the large number of Greater Hauraki assemblages from these periods. As noted, birds are consistently poorly represented across all these assemblages.
For reasons outlined in relation to fish, the low representation of birds in later Greater Hauraki sites is most unlikely to be due to poor bone survival. It is more difficult to be sure that it was not influenced by the preservation and transport of bird carcasses. Some nineteenth-century accounts indicate that bones were removed from pigeons and other large birds before they were cooked and potted in fat (Best Citation1909:474), while the method usually described for ‘mutton birds’ left all the bones except those from the head and extremities of the limbs to travel with the carcass to wherever it was consumed (Richdale Citation1948:100–102). Although it might be presumed that the latter is likely to have been the norm for at least the Procellariidae, it is impossible to rule out regional variations in preservation methods as a contributing factor in the paucity of marine birds after the EM period in Greater Hauraki. Even with that caution in mind, the magnitude of difference between the trajectories of bird representation in the two study regions strongly suggests that Maori harvesting had a much more rapid and dramatic impact on coastal bird populations in northern New Zealand.
Marine mammal exploitation in tropical Polynesia included hunting dolphins with harpoons (Leach et al. Citation1984; Anderson et al. Citation1994) and very rare examples of capturing seals (Walter & Smith Citation1998; Smith et al. Citation2001). The strong focus on the later, and especially fur seals, in New Zealand can be attributed to their abundance and the predictability with which they could be captured at breeding colonies and haul-outs (Smith Citation2005). Technological factors such as the size and strength of bone harpoon heads, fibre lashings and rope, and stability of canoes are likely to have contributed to the range of cetaceans exploited being restricted to dolphins and only those larger species that commonly strand.
Marine mammals disappeared from the Greater Hauraki study sample by the end of the EM period (). This is largely a record of declining presence of fur seals, sea lions and elephant seals, and all but the first mentioned of these were no longer represented in Otago-Catlins sites by the beginning of the Late period. Detailed analysis elsewhere (Smith Citation1985, Citation1989, Citation2005) has shown that the southward retreat of fur seal and sea lion breeding ranges cannot be attributed to climatic change, habitat modification or introduced predators. Instead it was due to moderate but sustained human predation, and happened most rapidly in northern New Zealand where human population growth was sustained by horticultural production. There is as yet no evidence that elephant seals maintained breeding colonies in New Zealand, although they were present prehistorically in Tasmania (Bryden et al. Citation1999), but their disappearance from both study regions after the EM period is suggestive of a predation effect. Leopard seals appear to never have been more than occasional visitors to New Zealand, and there is nothing in the archaeological record to suggest any change in that through time. Difficulties of identification preclude any meaningful interpretation of the record for cetaceans, although, as noted above, it is suspected that dolphins were hunted more often than is reflected in the present study samples and that this probably occurred throughout the prehistoric sequence (Smith Citation1989).
Conclusion
Previous analyses of pre-European Maori exploitation of marine resources have generally been either detailed analyses of a single site, national reviews of data for a single class of fauna, or very general overviews. Most were undertaken prior to recent reassessments of New Zealand's prehistoric chronology which relegated to an ‘undated’ status much of the evidence upon which they were based (Smith Citation2008:369). The present study explicitly set out to examine the full range of marine resources evidenced in the archaeological record, using information from as many sites within each of two study regions that met data quality standards and could be securely placed in time. While the broad pattern of marine exploitation described here is consistent with many of those earlier studies, there is now greater clarity about the nature, extent and causes of both regional differences and changes over time.
The evidence from this study indicates that Maori harvested marine resources in a pragmatic and opportunistic way, with those taxa most abundantly and predictably available near at hand featuring most prominently in the archaeological record. Harvesting technology provided some constraints on the range of species that could be taken, and it is possible that others were excluded due to cultural preferences. Regional differences in exploitation largely reflect geographical variations in the availability of marine taxa. There is very little evidence for targeted exploitation of specific taxa.
Changes over time are apparent in two main ways. Some taxa, including seals and coastal birds, were exploited less often and less widely as time progressed, and it is most likely that this reflects the impact of sustained harvesting by Maori. These changes occurred most rapidly and more completely in the northern study region, where human population growth, enabled by horticultural production, is likely to have been more rapid and sustained. Changes over time in the composition of shellfish and finfish harvests are much less likely to be due to harvesting pressure. At least some of these changes in the northern study area have been linked to changes in human settlement patterns which appear to reflect increasing demand for horticultural land, highlighting the importance of viewing Maori use of marine resources within the broader context of their total subsistence and settlement patterns.
Acknowledgements
I am grateful to Alison MacDiarmid for the opportunity to participate in this project, Foss Leach for inspiration and support, and Tiffany James-Lee for assistance with data collection. The text has benefitted from comments by Trevor Worthy and an anonymous referee. The research was supported by funding from Ministry of Fisheries.
References
- Anderson , AJ . 1981 . A model of prehistoric collecting on the rocky shore . Journal of Archaeological Science , 8 : 109 – 121 .
- Anderson , AJ . 1989 . Prodigious birds: Moas and moa-hunting in prehistoric New Zealand , 238 Cambridge : Cambridge University Press .
- Anderson , AJ . 1997a . “ Prehistoric Polynesian impact on the New Zealand environment: Te Whenua Hou ” . In Historical ecology of the Pacific Islands: Prehistoric environmental and landscape change , Edited by: Kirch , PV and Hunt , TL . 271 – 283 . New Haven : Yale University Press .
- Anderson , AJ . 1997b . Uniformity and regional variation in marine fish catches from prehistoric New Zealand . Asian Perspectives , 36 : 1 – 22 .
- Anderson , AJ . 1998 . The welcome of strangers: an ethnohistory of southern Maori A.D. 1650–1850 , Dunedin : University of Otago Press. 249 p .
- Anderson , AJ and Smith , IWG . 1996a . “ Analysis of fish remains ” . In Shag River Mouth: the archaeology of an early southern Maori village , Edited by: Anderson , AJ , Allingham , B and Smith , IWG . 237 – 244 . Canberra : ANH Publications .
- Anderson , AJ and Smith , IWG . 1996b . The transient village in southern New Zealand . World Archaeology , 27 : 359 – 371 .
- Anderson , AJ , Leach , HM , Smith , IWG and Walter , RK . 1994 . Reconsideration of the Marquesan sequence in East Polynesian prehistory, with particular reference to Hane (MUH1) . Archaeology in Oceania , 29 : 29 – 52 .
- Baker , C , Chilvers , B , Constantine , R , DuFresne , S , Mattlin , R , van Helden , A and Hitchmough , R . 2010 . Conservation status of New Zealand marine mammals (suborders Cetacea and Pinnipedia), 2009 . New Zealand Journal of Marine and Freshwater Research , 44 : 101 – 115 .
- Bathgate MA 1992 . Evidence of Dr Murray Alexander Bathgate on the archaeological and early documentary record concerning Maori fishing in the South Island, including reference to two reports by Dr Foss Leach (#S4 and #S5) . Waitangi Tribunal Report, 5WTR: S2 .
- Beattie , JH . 1920 . Nature-lore of the southern Maori . Transactions of the New Zealand Institute , 32 : 53 – 77 .
- Best , E . 1909 . Maori forest lore: Being some account of native forest lore and woodcraft, as also of many myths, rites, customs, and superstitions connected with the flora and fauna of the Tuhoe or Urewera district.—Part III . Transactions of the New Zealand Institute , 42 : 433 – 481 .
- Best , E . 1929 . Fishing methods and devices of the Maori , 230 Wellington : Dominion Museum .
- Bossenkool , S , Austin , JJ , Worthy , TH , Scofield , P , Cooper , A , Seddon , PJ and Waters , JM . 2009 . Relict or colonizer? Extinction and range expansion of penguins in southern New Zealand . Proceedings of the Royal Society B , 276 : 815 – 821 .
- Bryden , MM , O'Connor , S and Jones , R . 1999 . Archaeological evidence for the extinction of a breeding population of elephant seals in Tasmania in prehistoric times . International Journal of Osteoarchaeology , 9 : 430 – 437 .
- Burton , RK , Snodgrass , JJ , Gifford-Gonzalez , D , Guilderson , T , Brown , T and Koch , PL . 2001 . Holocene changes in the ecology of northern furseals: insights from stable isotopes and archaeofauna . Oecologia , 128 : 107 – 115 .
- Cassels , R . 1984 . “ Role of prehistoric man in the faunal extinctions of New Zealand and other Pacific Islands ” . In Quaternary extinctions: A prehistoric revolution , Edited by: Martin , PS and Klein , RG . 741 – 767 . Tucson : University of Arizona Press .
- Checklist Committee OSNZ 2010 . Checklist of the birds of New Zealand, Norfolk and Macquarrie Islands, and the Ross Dependency of Antarctica. , 4th edition Wellington , Ornithological Society of New Zealand and Te Papa Press . 500
- Colenso , W . 1869 . On the Maori races of New Zealand . Transactions of the New Zealand Institute , 1 : 339 – 424 .
- Colenso , W . 1891 . Vestiges: reminiscences: memorabilia of works, deeds and sayings of the ancient Maori . Transactions of the New Zealand Institute , 24 : 445 – 467 .
- Etnier , M . 2007 . Defining and identifying sustainable harvests of resources: archaeological examples of pinniped harvests in the eastern North Pacific . Journal for Nature Conservation , 15 : 196 – 207 .
- Froese R , Pauly D 2010 . FishBase . www.fishbase.org, version 09, 2010 (accessed 21 September 2010) .
- Furey , L . 1996 . Oruarangi: The archaeology and material culture of a Hauraki pa , Auckland : Auckland Institute and Museum. 222 p .
- Furey , L . 2002a . Houhora. A fourteenth century Maori village in Northland , 169 Auckland : Auckland War Memorial Museum .
- Furey , L . 2002b . Archaeological investigations, Whitianga Waterways , 17 Report to New Zealand Historic Places Trust .
- Grayson , DK . 1978 . Minimum numbers and sample size in vertebrate faunal analysis . American Antiquity , 43 : 53 – 65 .
- Grayson , DK . 2001 . The archaeological record of human impacts on animal populations . Journal of World Prehistory , 16 : 1 – 68 .
- Green , RC . 1970 . A Review of the prehistoric sequence in the Auckland province , 2nd edition , 60 Dunedin : University Bookshop .
- Holdaway , RN . 1999 . “ Introduced predators and avifaunal extinction in New Zealand ” . In Extinctions in near time: Causes, contexts and consequences , Edited by: MacPhee , RDE . 189 – 238 . New York : Kluwer Academic .
- Holdaway , RN and Worthy , TH . 1994 . A new fossil species of shearwater Puffinus from the late Quaternary of the South Island, New Zealand, and notes on the the biogeography and evolution of the Puffinus gavia super-species . Emu , 94 : 201 – 215 .
- Holdaway , RN , Worthy , TH and Tennyson , AJD . 2001 . A working list of breeding bird species of the New Zealand region at first human contact . New Zealand Journal of Zoology , 28 : 119 – 187 .
- King , CM . 1995 . The handbook of New Zealand mammals , 600 Auckland : Oxford University Press .
- Kirch , PV and Yen , DE . 1982 . Tikopia: The prehistory and ecology of a Polynesian outlier , Honolulu : Bernice P. Bishop Museum. 396 p .
- Leach , BF . 2006 . Fishing in pre-european New Zealand , 359 Wellington : New Zealand Journal of Archaeology, and Archaeofauna .
- Leach BF , Ward GK 1981 . Archaeology on Kapingamarangi atoll: a Polynesian outlier in the Eastern Caroline Islands . Dunedin, University of Otago Studies in Prehistoric Anthropology 16 . 64
- Leach BF , Boocock AS 1993 . Prehistoric fish catches in New Zealand . Oxford, British Archaeological Reports International Series No. 584 . 38
- Leach , BF , Intoh , M and Smith , IWG . 1984 . Fishing, turtle hunting and mammal exploitation at Fa'ahia, Huahine, French Polynesia . Journal de la Societe des Oceanistes , 79 : 183 – 197 .
- Lyman , RL . 2008 . Quantitative paleozoology , 348 Cambridge : Cambridge University Press .
- Magurran , AE . 2004 . Measuring biological diversity , 256 Oxford : Blackwell Publishing .
- Mann SR 2009 . How to catch a Leatherjacket: Prehistoric fishing strategies at Arthur Black's midden, Opito Bay, New Zealand . Unpublished MA thesis, Anthropology, University of Otago . 136
- Marshall , YM . 1987 . Maori mass capture of freshwater eels: an ethnoarchaeological reconstruction of prehistoric subsistence and social behaviour . New Zealand Journal of Archaeology , 9 : 55 – 79 .
- Matthews , RH . 1910 . Reminiscences of Maori life fifty years ago; 1. Shark fishing . Transactions of the New Zealand Institute , 43 : 598 – 603 .
- McGlone , MS . 1989 . The Polynesian settlement of New Zealand in relation to environmental and biotic changes . New Zealand Journal of Ecology , 12 ( suppl ) : 115 – 129 .
- Millener PR 1981 . The Quaternary avifauna of the North Island, New Zealand . Unpublished PhD thesis, University of Auckland, New Zealand . 897
- Moore PR 2006 . Archaeological Monitoring and Investigation, Site U13/1246, The Crescent, Waihi Beach . Waihi, Peninsula Research Report No. 2006–03 . 20
- Morley , MS and Hayward , BW . 2009 . Marine mollusca of Great Barrier Island, New Zealand . Records of the Auckland Museum , 46 : 15 – 51 .
- Murdoch , GJ . 1996 . A history of the human occupation of the Whakakaiwhara block , 68 Auckland : Auckland Regional Council .
- Myers , RA and Worm , B . 2003 . Rapid worldwide depletion of predatory fish communities . Nature , 423 : 280 – 283 .
- Paul , LJ . 2000 . New Zealand fishes: identification, natural history & fisheries , 253 Auckland : Reed .
- Paulin , CD . 2007 . Perspectives of Maori fishing history and techniques. Nga ahua me nga purukau me nga hangarau ika o te Maori . Tuhinga , 18 : 11 – 47 .
- Pauly , D , Christensen , V , Dalsgaard , J , Froese , R and Torres , FJ . 1998 . Fishing down marine food webs . Science , 279 : 860 – 863 .
- Phillips C 1997 . Archaeological excavation of site (T12/959), Moana Park subdivision, Whanagamata . Report to the New Zealand Historic Places Trust . 46
- Polack , JS . 1840 . Manners and customs of the New Zealanders: With notes corroborative of their habits, usages etc., and remarks to intending emigrants. 2 Volumes , London : James Madden and Co .
- Reeves , RR and Smith , TD . 2006 . “ A taxonomy of world whaling: operations and eras ” . In Whales, whaling, and ocean ecosystems , Edited by: Estes , JA , DeMaster , DP , Doak , DF , Williams , TM and Brownell , RLJ . 82 – 101 . Berkeley : University of California Press .
- Reitz , EJ and Wing , ES . 2008 . Zooarchaeology , 2nd edition , 533 Cambridge : Cambridge University Press .
- Richdale LE 1948 . Maori and muttonbird . Dunedin, Otago Daily Times and Witness , Wildlife Series No. 7 . 15
- Rick TC , Erlandson JM 2008 . Human impacts on ancient marine ecosystems: A global perspective . Berkley , University of California Press . 336
- Shawcross , W . 1967 . An evaluation of the theoretical capacity of a New Zealand harbour to carry a human population . Tane , 13 : 3 – 11 .
- Smith IWG 1985 . Sea mammal hunting and prehistoric subsistence in New Zealand . Unpublished PhD thesis, Anthropology, University of Otago . 576
- Smith IWG 1989 . Maori impact on the marine megafauna: pre-European distributions of New Zealand sea mammals . In : Sutton DG Saying so doesn't make it so: Papers in honour of B. Foss Leach . Dunedin , New Zealand Archaeological Association Monograph 17 . 76 – 108 .
- Smith , IWG . 2005 . “ Retreat and resilience: fur seals and human settlement in New Zealand ” . In The exploitation and cultural importance of marine mammals , Edited by: Monks , GG . 6 – 18 . Oxford : Oxbow Books .
- Smith IWG 2008 . Maori, Pakeha and Kiwi: peoples, cultures and sequence in New Zealand archaeology . In : Clark GR , Leach BF , O'Connor S Islands of inquiry: Colonisation, seafaring and the archaeology of maritime landscapes . Canberra , ANU EPress . 367 – 380 .
- Smith , IWG . 2010 . Protocols for organising radiocarbon dated assemblages from New Zealand archaeological sites for comparative analysis . Journal of Pacific Archaeology , 1 : 184 – 187 .
- Smith IWG , James-Lee T 2010 . Data for an archaeozoological analysis of marine resource use in two New Zealand study areas . Revised edition. Otago Archaeological Laboratory Report, No 7. 85 p. www.otago.ac.nz/anthropology/anth/publications.
- Smith IWG , Clark GR , White P 2001 . Mammalian and reptilian fauna from Emily and Cemetry Bays, Norfolk Island . In : Anderson AJ , White P The prehistoric archaeology of Norfolk Island, southwest Pacific . Sydney , Australian Museum , Records of the Australian Museum, Supplement 27 . 77 – 81 .
- Spencer HG , Willan RC , Marshall B , Murray TJ 2009 . Checklist of the recent mollusca recorded from the New Zealand exclusive economic zone . www.molluscs.otago.ac.nz/index.html (accessed 20 September 2010) .
- Steadman , DW . 1997 . “ Extinctions of Pacific birds: reciprocal impacts of birds and people ” . In Historical ecology in the Pacific islands: Prehistoric environmental and landscape change , Edited by: Kirch , PV and Hunt , TL . 51 – 79 . New Haven : Yale University Press .
- Swadling , PL . 1976 . Changes induced by human exploitation in prehistoric shellfish populations . Mankind , 10 : 156 – 62 .
- Taylor , R . 1855 . Te Ika a Maui , 490 London : Wertheim and MacIntosh .
- Walter , RK and Smith , IWG . 1998 . Identification of a New Zealand fur seal (Arctocephalus forsteri) bone in a Cook Island archaeological site . New Zealand Journal of Marine and Freshwater Research , 32 : 483 – 487 .
- Walter , RK , Smith , IWG and Jacomb , C . 2006 . Sedentism, subsistence and socio-political organization in prehistoric New Zealand . World Archaeology , 38 : 274 – 290 .
- Watson N 1969 . Pre-european tribal trouble, Part 2 . Ohinemuri Regional History Journal 11. www.ohinemuri.org.nz/journal (accessed 12 November 2010) .
- Wheeler , A and Jones , AJG . 1989 . Fishes , 210 Cambridge : Cambridge University Press .
- Witter , D . 1969 . Notes on the significance of the leather jacket fish in middens . New Zealand Archaeological Association Newsletter , 12 : 52
- Worthy , TH . 1998a . Quaternary fossil faunas of Otago, South Island, New Zealand . Journal of the Royal Society of New Zealand , 28 : 421 – 521 .
- Worthy , TH . 1998b . A remarkable fossil and archaeological avifauna from Marfells Beach, Lake Grassmere, South Island, New Zealand . Records of the Canterbury Museum , 12 : 79 – 176 .
- Worthy , TH . 1999 . What was on the menu? Avian extinction in New Zealand . New Zealand Journal of Archaeology , 19 : 125 – 160 .
- Worthy , TH and Holdaway , RN . 2002 . The lost world of the Moa , 718 Christchurch : Canterbury University Press .