Abstract
The extinction date of the giant flightless New Zealand ratite bird, the crested moa (Pachyornis australis), is of considerable interest because the youngest verified remains are dated to the Pleistocene–Holocene transition c. 10,000 yr BP, which was characterised by severe climatic and habitat change, and are considerably earlier than the late Holocene extinctions of the other eight moa species. Analysis of a partial crested moa skeleton (NMNZ S23569) from Castle Keep Entrance, Bulmer Cave System, Mount Owen, South Island, generated a radiocarbon date of 564±26 yr BP (544–508 cal yr BP; 95.4% AD 1396–1442). As a result the Bulmer Cave specimen represents the youngest moa yet found from a natural site in New Zealand. Combined with additional crested moa remains dated to the late Holocene from Cheops Cave (Mount Arthur) and Magnesite Quarry (Cobb Valley), this indicates that crested moa did not go extinct during the Pleistocene–Holocene transition but survived until after Polynesian colonisation in c. AD 1280. The new radiocarbon dates reported here have important implications for the timing of moa extinction and the late survival of moa in alpine areas of New Zealand.
Introduction
Crested moa (Pachyornis australis Oliver, 1949) has the most restricted distribution of the currently recognised nine species of moa (Bunce et al. Citation2009; Gill et al. Citation2010), and was formally found mainly in northwest Nelson, South Island (Worthy & Holdaway Citation2002) () in areas where sub-alpine grassland-shrubland dominated (Worthy Citation1989). The remains of this species are common in caves above 800 m asl (Worthy Citation1989, Citation1997a) and are especially prevalent in Pleistocene deposits from the Honeycomb Hill cave system in the Oparara Valley, north of Karamea (Worthy Citation1993), and the Megamania cave system in the Gunner-Heaphy River catchment (Worthy Citation1998a). There have also been reports of crested moa in Southland dunes (Worthy Citation1998b) though these have not been confirmed by ancient DNA (aDNA) analysis. Despite the abundance of crested moa remains in northwest Nelson, this taxon is one of the rarest moa species in museum collections.
Figure 1 Geographic locations within northwest Nelson, South Island, of late Holocene crested moa remains, determined by radiocarbon dating.
The youngest reported remains of crested moa thus far are only 10,165±50 yr old from Bone Cave, Canaan Road, Takaka Hill (OxA 12430, δ13C −23.174; Bunce et al. Citation2009) and date to the Pleistocene–Holocene transition, which was characterised by severe climatic and habitat change (McGlone Citation1988; Williams et al. Citation2005). This, combined with the absence of crested moa in cultural sites, has caused debate over when this species went extinct (Worthy Citation1997b; Anderson Citation2002; Tennyson & Martinson Citation2007).
Location of site and site investigation
In 1987, an in situ partial skeleton of a moa was reported from the Castle Keep Entrance of the Bulmer Cave System, on Mount Owen at 1600 m asl (Worthy Citation1989) (). A tibiotarsus (now housed in the National Museum of New Zealand Te Papa Tongawera under the accession number NMNZ S23569) was collected, but additional bones, including a femur, premaxilla, mandible, and other unidentified elements, lie deeper in the cave at the bottom of a series of vertical pitches (Worthy Citation1989).
Species identification
The bones were identified as crested moa following Worthy (Citation1988) and this identification has been confirmed by aDNA analysis. DNA extraction and Polymerase Chain Reaction (PCR) set-up were performed in a geographically isolated, dedicated aDNA laboratory at the Australian Centre for Ancient DNA following strict aDNA procedures to minimise contamination of samples with exogenous DNA (Cooper & Poinar Citation2000). DNA was extracted from 0.2–0.3 mg of bone powder following Weinstock et al. (Citation2005). To identify moa species, we used two overlapping primer pairs (185F/294R, 262F/429R) targeting the mitochondrial DNA control region (Cooper et al. Citation2001; Bunce et al. Citation2003). PCR amplification was conducted following Wood et al. (Citation2008). PCR products were sequenced in both directions and ‘blasted’ against the c. 300 moa control region sequences available on GenBank. This methodology allowed robust species identification of moa bones.
Radiocarbon dating
Radiocarbon dating of the Bulmer Cave specimen was performed at the Oxford Radiocarbon Accelerator Unit following Bronk Ramsey et al. (Citation2004a,b) and Brock et al. (Citation2010). The Bulmer Cave specimen was dated to 564±26 yr BP (544–508 cal yr BP, AD 1396–1442; ). Carbon to nitrogen atomic ratios should be within the range of 2.9–3.5, with the value for unadulterated bone 3.18. The C:N ratio of the Bulmer Cave specimen is 3.3, which gives confidence in the radiocarbon date obtained (). In addition, the most important parameter increasing confidence in the radiocarbon date is the very high collagen yield of the Bulmer Cave specimen at 15.5% compared to 20% in modern bone (Brown et al. Citation1988) (). The high value means the collagen has undergone minimal post-mortem diagenesis. The age of the Bulmer Cave specimen can be precisely calibrated because the 95.4% probability distribution falls outside the ‘wiggle’ that characterises the Southern Hemisphere calibration curve (McCormac et al. Citation2004) from c. AD 1300 to 1400 (). This wiggle makes precise calibration of radiocarbon dates during the initial period of Polynesian colonisation and subsequent environmental modification difficult because individual radiocarbon dates can intercept the calibration curve in multiple positions.
Figure 2 Probability distribution for calibrated ages of moa remains from natural and archaeological deposits. Dates were calibrated using the Southern Hemisphere calibration curve (McCormac et al. Citation2004) and plotted in Oxcal v4.1 (Bronk Ramsey Citation2009) with 95.4% confidence intervals underlined. The period between Polynesian settlement (c. AD 1280) and c. AD 1450, during which moa were hunted to extinction (purple bar; Holdaway & Jacomb Citation2000), and the pronounced wiggle in the Southern Hemisphere calibration curve c. AD 1300–1400 (orange bar) are also shown.
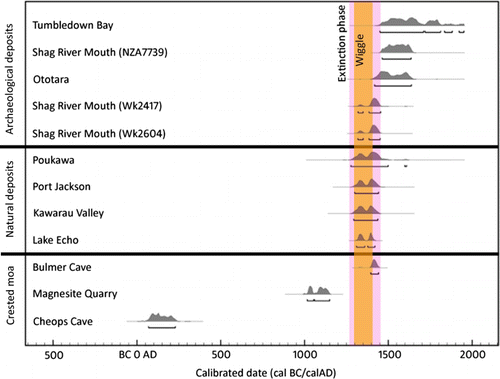
Table 1 Radiocarbon dates on crested moa remains produced in this study. Dates were calibrated using the Southern Hemisphere calibration curve (McCormac et al. Citation2004) in OxCal v4.1 (Bronk Ramsey Citation2009).
The remains of two additional crested moa (confirmed by aDNA analysis, Bunce et al. 2009; this study) were also radiocarbon dated to the late Holocene (). The first specimen (NMNZ S41344) was collected by Trevor Worthy, Aaron Worthy, Alan Tennyson and Alan Cooper on 16 April 2003 at Cheops Cave, Hodge Creek, Mount Arthur (900 m asl; ). The Cheops Cave specimen comprises an almost complete skeleton and is housed in the National Museum of New Zealand. The second specimen was discovered by Brian Muddiman in 1964 in a small cave below the Cobb Dam at Magnesite Quarry (1005 m asl; ). The specimen consisted of a right femur and a left tarsometatarsus. It is housed in Nelson Museum (NP5305.1-2).
Significance of the discovery
Post-settlement radiocarbon dates of moa
The Bulmer Cave specimen postdates the Polynesian colonisation of New Zealand which took place c. AD 1280 (Hogg et al. Citation2003; Wilmshurst et al. Citation2008) by at least 115 years, and represents the youngest reliably dated moa from a natural site (; ). The next youngest moa remains from natural sites include those from Poukawa, Hawke's Bay at 586±85 yr BP (95.4% AD 1277–1610, moa sp., δ13C−29.68, NZ3701; Anderson Citation1989a); Port Jackson, Coromandel at 605±56 yr BP (95.4% AD 1300–1443, moa sp., δ13C −23.10, NZ4883; Millener Citation1981); Kawarau Valley, Central Otago at 620±60 yr BP (95.4% AD 1294–1438, heavy-footed moa Pachyornis elephantopus (Owen, 1856), δ13C −29.29, NZ3455; Anderson Citation1989a); and Echo Valley, Fiordland at 623±28 yr BP (95.4% AD 1310–1420, δ13C−21.2, little bush moa Anomalopteryx didiformis (Owen, 1844), OxA-12729; Bunce et al. Citation2009).
The New Zealand archaeological record contains accepted radiocarbon dates on moa bone and eggshell younger than the Bulmer Cave specimen (Hogg & Higham Citation1997; Petchey Citation1997; Anderson Citation2000; Schmidt Citation2000). The three youngest dates on bone are from Ototara, North Otago at 435±70 yr BP (95.4% AD 1418–1638, stout legged moa Euryapteryx curtus (Owen, 1846), δ13C−25, NZ0754); Shag River Mouth, North Otago at 370±38 yr BP (95.4% AD 1464–1637, stout legged moa, δ13C−24, NZA7739); and from Tumbledown Bay, Canterbury at 307±85 yr BP (95.4% AD 1451–1952, South Island giant moa (Dinornis robustus Owen, 1846), δ13C−21.90, NZA0338). The youngest dates on eggshell are from Shag River Mouth: 570±45 yr BP (95.4% AD 1319–1451, δ13C−11.4, Wk2604) and 560±45 yr BP (95.4% AD 1320–1445, δ13C−15, Wk2417).
The calibrated radiocarbon dates on natural moa remains and archaeological eggshell overlap with the Bulmer Cave date but fall within the c. AD 1300–1400 wiggle of the Southern Hemisphere calibration curve so cannot be precisely calibrated. The exceptions are the two Shag River eggshell dates, which have the majority (80%–85%) of the 95.4% probability distribution outside the wiggle and therefore can be calibrated more precisely. Of the calibrated archaeological bone dates, only the Ototara date overlaps with the Bulmer Cave date, though the associated errors of all three archaeological dates are large. The above dates with the exception of the Bulmer Cave and Lake Echo dates were conducted on bone collagen, not (ultra-filtrated) pure bone gelatin (see Brown et al. Citation1988; Bronk Ramsey et al. Citation2004a,b; Brock et al. Citation2010). In addition, dates with laboratory numbers prefixed with NZ were analysed by the gas counting method that required near 1 kg of bones, so often multiple bones were combined to achieve a sufficient sample: this is the case for the Port Jackson date and is likely to be the case for the Poukawa date. The Port Jackson date was obtained from a bone sample comprising three different moa species, each with differential preservation. The non-ultra-filtrated and gas count dates should be treated with caution. The Bulmer Cave specimen therefore represents the youngest reliably radiocarbon dated moa thus far discovered.
Extinction of crested moa
The three crested moa specimens radiocarbon dated in this study () represent the first authenticated late Holocene remains of this species and confirm that crested moa did not go extinct during the Pleistocene–Holocene transition c. 10,000 yr BP as a result of severe climatic and habitat change but survived until well after Polynesian colonisation in the late thirteenth century. Dating of further crested moa remains found in high-altitude sub-alpine grassland-shrubland above 800 m asl (Worthy Citation1989), like those found recently on Mount Mytton, Cobb Valley (Jamie Wood pers. obs.), are likely to produce additional Holocene or post-settlement remains and begin to fill the early-mid Holocene radiocarbon gap in crested moa.
There is overwhelming archaeological and palaeoenvironmental evidence that overhunting and habitat destruction by early Māori were the dominant causes of the extinction of moa (Anderson Citation1989a; Wilmshurst & Higham Citation2004; Wilmshurst et al. Citation2008; McWethy et al. Citation2010). Though no verifiable crested moa remains have been found in Māori middens (Worthy Citation1997b; Anderson Citation2002; Tennyson & Martinson Citation2007), the new radiocarbon dates () suggest that it is highly probable that crested moa were hunted to extinction. Māori were hunting moa in remote subalpine areas of the South Island where crested moa was found during the early moa hunter period. O'Regan (Citation1992) radiocarbon dated the butchered remains of another sub-alpine specialist, the upland moa (Megalapteryx didinus) (Owen, 1883), from Takahe Valley rockshelter, Murchison Mountains, Fiordland (800 m asl) to 628±39 yr BP (95.4% AD 1300–1422, δ13C−23.5, NZA2227). The paucity of bone in swamp deposits in northwest Nelson and the relatively few ‘moa hunter period’ archaeological sites in the same area (Anderson Citation1989a; Worthy & Holdaway Citation1994), and the difficulty in distinguishing between crested and heavy-footed moa (P. elephantopus) in some palaeontological sites (Worthy & Holdaway Citation2002; Rawlence Citation2010), suggests crested moa are likely to have formed a cryptic component of the archaeological record.
Late survival of moa in alpine areas of New Zealand
Holdaway and Jacomb (Citation2000) hypothesised that moa became extinct within 50–100 years of Polynesian colonisation. However, this study relied heavily on modelling and ignored many accepted young post-settlement radiocarbon dates on moa bones (Anderson Citation2000). The radiocarbon dates reported here () suggest that the last surviving moa populations inhabited the relatively inaccessible subalpine areas of the South Island as has been previously hypothesised by Anderson (Citation1989b). This hypothesis is supported by the analysis of the coprophilous fungus Sporormiella Ellis & Everhart 1892, a proxy for megaherbivore biomass and extinction, from a forest soil core collected in the Murchison Mountains (Wood et al. Citation2011). This study showed that the regional ‘megaherbivore extinction’ may have occurred by the late seventeenth century AD. This late surviving megaherbivore population in alpine Fiordland no doubt included moa (O'Regan Citation1992). Our knowledge of the moa species in these late surviving alpine populations has largely come from the palaeontological investigation of caves in northwest Nelson and Fiordland (Worthy Citation1989). From this, the regional moa fauna, in order of decreasing relative fossil abundance, appears to have comprised upland moa, crested moa (restricted to northwest Nelson) and South Island giant moa, and probably represented some of the last moa species to become extinct.
The results of this study also have serious implications for hypotheses suggesting that introduced diseases (Gemmell et al. Citation2004) and human-mediated habitat loss (Didham et al. Citation2005) were the main drivers of moa extinction in contrast to overhunting or multi-causal extinction scenarios (Anderson Citation1989a; Wilmshurst & Higham Citation2004; Wilmshurst et al. Citation2008; McWethy et al. Citation2010). Introduced avian diseases have been documented to have little impact on alpine bird communities. In Hawaii, mosquito-borne diseases only affected lowland species below c. 600 m asl (Boyer Citation2008). In New Zealand, habitat destruction by early Māori was restricted mostly to lowland areas (McGlone Citation1989; Ogden et al. Citation1998). The same is true for Hawaii, where only lowland species became extinct following habitat destruction by Polynesians, while montane-alpine species survived in relatively unmodified habitat (Boyer Citation2008). The evidence for late surviving alpine moa populations supports the hypothesis that overhunting in combination with habitat destruction was the main driver of moa extinction.
Given the radiocarbon dates and discussion presented in this study, it is therefore highly probable that some of the youngest moa remains in New Zealand will be found in alpine areas of the South Island. Radiocarbon dating of further remains from these areas will help determine how long moa populations survived after initial Polynesian colonisation and will help resolve the debate over the timing of moa extinction.
Acknowledgements
We are grateful to the National Museum of New Zealand Te Papa Tongawera (Alan Tennyson) and Nelson Museum (Judith Taylor, Paula Haines-Bellamy) for permission to sample specimens. Thanks also to Jamie Wood, Trevor Worthy and Alan Tennyson for helpful discussions and comments on this manuscript. Funding was provided by an Australian Research Council Discovery Grant (20101413).
References
- Anderson , A . 1989a . Prodigious birds , Cambridge : Cambridge University Press .
- Anderson , A . 1989b . On evidence for the survival of moa in European Fiordland . New Zealand Journal of Ecology , 12 ( Supplement ) : 39 – 44 .
- Anderson , A . 2000 . Less is moa . Science , 289 : 1472 – 1474 .
- Anderson , A . 2002 . Faunal collapse, landscape change and settlement history in Remote Oceania . World Archaeology , 33 : 375 – 390 .
- Boyer , AG . 2008 . Extinction patterns in the avifauna of the Hawaiian islands . Diversity and Distributions , 14 : 509 – 517 .
- Brock , F , Higham , TFG , Ditchfield , P and Bronk Ramsey , C . 2010 . Current pretreatment methods for AMS radiocarbon dating at the Oxford Radiocarbon Accelerator Unit (ORAU) . Radiocarbon , 52 : 103 – 112 .
- Bronk Ramsey , C . 2009 . Bayesian analysis of radiocarbon dates . Radiocarbon , 51 : 337 – 360 .
- Bronk Ramsey , C , Higham , TFG , Bowles , A and Hedges , R . 2004a . Improvements to the pretreatment of bone at Oxford . Radiocarbon , 46 : 155 – 163 .
- Bronk Ramsey , C , Higham , TFG and Leach , P . 2004b . Towards high precision AMS: progress and limitations . Radiocarbon , 46 : 7 – 24 .
- Brown , TA , Nelson , DE , Vogel , JS and Southon , JR . 1988 . Improved collagen extraction by modified Longin method . Radiocarbon , 30 : 171 – 177 .
- Bunce , M , Worthy , TH , Ford , T , Hoppitt , W , Willerslev , E , Drummond , A and Cooper , A . 2003 . Extreme reversed sexual size dimorphism in the extinct New Zealand moa Dinornis . Nature , 425 : 172 – 175 .
- Bunce , M , Worthy , TH , Phillips , M , Holdaway , RN , Willerslev , E , Haile , J , Shapiro , B , Scofield , P , Drummond , A , Kamp , P and Cooper , A . 2009 . The evolutionary history of the extinct ratite moa and New Zealand palaeogeography revealed by ancient DNA . Proceedings of the National Academy of Sciences USA , 106 : 20646 – 20651 .
- Cooper , A , Lalueza-Fox , C , Anderson , A , Rambaut , A , Austin , J and Ward , R . 2001 . Complete mitochondrial genome sequences of two extinct moas clarify ratite evolution . Nature , 409 : 704 – 707 .
- Cooper , A and Poinar , HN . 2000 . Ancient DNA: do it right or not at all . Science , 289 : 1139
- Didham , RK , Ewers , RM and Gemmell , NJ . 2005 . Comment on ‘Avian extinction and mammalian introductions on oceanic islands’ . Science , 307 : 1412
- Gemmell , NJ , Schwartz , MK and Robertson , BC . 2004 . Moa were many . Proceedings of the Royal Society of London Series B , 271 ( Supplement ) : S430 – 432 .
- Gill BJ , Bell BD , Chambers GK , Medway DG , Palma RL , Scofield RP , Tennyson AJD , Worthy TH 2010 . Checklist of the birds of New Zealand, Norfolk and Macquarie Islands, and the Ross Dependency, Antarctica . 4th edition . Wellington , Ornithological Society of New Zealand and Te Papa Press . 464
- Hogg , AG and Higham , TFG . 1997 . Evidence for late Polynesian colonisation of New Zealand: University of Waikato radiocarbon measurements . Radiocarbon , 39 : 149 – 192 .
- Hogg , AG , Higham , TFG , Lowe , DJ , Palmer , JG , Reimer , PJ and Newnham , RM . 2003 . A wiggle-match date for Polynesian settlement of New Zealand . Antiquity , 77 : 116 – 125 .
- Holdaway , RN and Jacomb , C . 2000 . Rapid extinction of the moas (Aves: Dinornithiformes): model, test and implications . Science , 287 : 2250 – 2254 .
- McCormac , FG , Hogg , AG , Blackwell , PG , Buck , CE , Higham , TFG and Reimer , PJ . 2004 . ShCal04 Southern Hemisphere calibration, 0–11.0 Cal Kyr BP . Radiocarbon , 46 : 1087 – 1092 .
- McGlone , MS . 1988 . “ New Zealand ” . In Vegetation history , Edited by: Huntley , B and Webb , T III . 557 – 599 . Dordrecht : Kluwer Academic .
- McGlone , MS . 1989 . The Polynesian settlement of New Zealand in relation to environmental and biotic changes . New Zealand Journal of Ecology , 12 ( Supplement ) : 115 – 129 .
- McWethy , DB , Whitlock , C , Wilmshurst , JM , McGlone , MS , Fromont , M , Li , X , Dieffenbacher-Krall , A , Hobbs , WO , Fritz , SC and Cook , ER . 2010 . Rapid landscape transformation in South Island, New Zealand, following initial Polynesian settlement . Proceedings of the National Academy of Sciences USA , 107 : 21343 – 21348 .
- Millener PR 1981 . The Quaternary avifauna of the North Island , New Zealand. Unpublished PhD thesis , University of Auckland , New Zealand .
- O'Regan , G . 1992 . The antiquity of the Takahe Valley rock shelter site . Archaeology in New Zealand , 35 : 172 – 176 .
- Ogden , J , Basher , L and McGlone , M . 1998 . Fire, forest regeneration and links with early human habitation: evidence from New Zealand . Annals of Botany , 81 : 687 – 696 .
- Petchey , FJ . 1997 . New Zealand bone dating revisited: a radiocarbon bone discard protocol for bone . New Zealand Journal of Archaeology , 19 : 81 – 124 .
- Rawlence NJ 2010 . Palaeoecology and population demographics of the extinct New Zealand moa (Aves: Dinornithiformes) . Unpublished PhD thesis , University of Adelaide , Australia .
- Schmidt , MD . 2000 . Radiocarbon dating the end of moa-hunting in New Zealand prehistory . Archaeology in New Zealand , 43 : 314 – 329 .
- Tennyson A , Martinson P 2007 ( reprint ). Extinct birds of New Zealand . Wellington , Te Papa Press . 180
- Weinstock , J , Willerslev , E , Sher , A , Tong , W , Ho , SYW , Rubenstein , D , Storer , J , Burns , J , Martin , L , Bravi , C , Prieto , A , Froese , D , Scott , E , Xulong , L and Cooper , A . 2005 . Evolution, systematics, and phylogeography of Pleistocene horses in the new world: a molecular perspective . PLoS Biology , 3 : e241
- Williams , PW , King , DNT , Zhao , JX and Collerson , KD . 2005 . Late Pleistocene to Holocene composite speleotherm 18O and 13C chronologies from South Island, New Zealand – did a global Younger Dryas really exist? . Earth and Planetary Science Letters , 230 : 301 – 317 .
- Wilmshurst , JM and Higham , TFG . 2004 . Using rat gnawed seeds to independently date the arrival of Pacific rat and humans to New Zealand . The Holocene , 14 : 801 – 806 .
- Wilmshurst , JM , Anderson , AJ , Higham , TFG and Worthy , TH . 2008 . Dating the late prehistoric dispersal of Polynesians to New Zealand using the commensal Pacific rat . Proceedings of the National Academy of Sciences USA , 105 : 7676 – 7680 .
- Wood , JR , Rawlence , NJ , Rogers , GM , Austin , JJ , Worthy , TH and Cooper , A . 2008 . Coprolite deposits reveal the diet and ecology of the extinct New Zealand megaherbivore moa (Aves, Dinornithiformes) . Quaternary Science Reviews , 27 : 2593 – 2602 .
- Wood , JR , Wilmshurst , JM , Worthy , TH and Cooper , A . 2011 . Sporormiella as a proxy for non-mamalian herbivores in island ecosystems . Quaternary Science Reviews , 30 : 915 – 920 .
- Worthy TH 1988 . An illustrated key to the main leg bones of moas (Aves: Dinornithiformes) . National Museum of New Zealand Miscellaneous Publications Series 17 .
- Worthy , TH . 1989 . Moas of the subalpine zone . Notornis , 36 : 191 – 196 .
- Worthy TH 1993 . Fossils of Honeycomb Hill . Wellington , National Museum of New Zealand . 56
- Worthy , TH . 1994 . Late Quaternary changes in the moa fauna on the West Coast of the South Island, New Zealand . Records of the South Australian Museum , 27 : 125 – 134 .
- Worthy , TH . 1997a . Fossil deposits in the Hodge Creek Cave System, on the northern foothills of Mt Arthur, Nelson, South Island, New Zealand . Notornis , 44 : 111 – 124 .
- Worthy , TH . 1997b . What was on the menu? Avian extinction in New Zealand . Archaeology in New Zealand , 19 : 125 – 60 .
- Worthy TH 1998a . Fossil deposits in Megamania Cave, Gunner River, South Island, New Zealand . Conservation Advisory Science Notes No. 195 . Wellington , Department of Conservation .
- Worthy , TH . 1998b . The Quaternary fossil avifauna of Southland, South Island, New Zealand . Journal of the Royal Society of New Zealand , 28 : 537 – 589 .
- Worthy , TH and Holdaway , RN . 1994 . Quaternary fossil faunas from caves in Takaka valley and on Takaka Hill, northwest Nelson, South Island, New Zealand . Journal of the Royal Society of New Zealand , 24 : 297 – 391 .
- Worthy TH , Holdaway RN 2002 . The lost world of the moa: prehistoric life of New Zealand . Christchurch , Canterbury University Press . 718