Abstract
Long-term ringing data are useful for understanding population trends and migration strategies adopted by migratory bird species during migration. To investigate the patterns in demography, phenology of migration and stopover behaviour in Wood Sandpipers Tringa glareola trapped on autumn migration at Ottenby, southeast Sweden, in 1947 − 2011, we analysed 65 years of autumn ringing data to describe age-specific trends in annual trappings, morphometrics and phenology, as well as fuel deposition rates and stopover duration from recapture data. We also analysed the migratory direction of the species from recovery data. Over the years, trapping of both adults and juveniles has declined significantly. Median trapping dates were 10 July for adults and 6 August for juveniles. Average migration speed of juvenile birds was 58.1 km d−1. Adults stayed on average 3.5 days and juveniles 5.2 days, with average fuel deposition rates of 2.5 and 0.7 g day−1 respectively. Juvenile birds probably vary their strategy according to time of season and prevailing conditions. Both adults and juveniles followed the Mediterranean Flyway, but juveniles displayed significantly more southerly and significantly more scattered migratory directions.
The Wood Sandpiper Tringa glareola is a relatively common wader species breeding in Scandinavia (except the south) and further east in Finland, the Baltic States and Russia, all the way to the Bering Strait (eg Cramp & Simmons Citation1983). During spring and autumn migration, numerous Wood Sandpipers pass Ottenby, southeastern Sweden. The first south-migrating adults appear at Ottenby in mid or late June, and in July the first juveniles occur. The migration, with mostly juveniles, continues to late August and the beginning of September. Occasional birds are sometimes found later in September and in October (Kolthoff Citation1896). The migration of the Wood Sandpiper at Ottenby and in Sweden was first analysed by Myhrberg Citation(1961), using ring recoveries primarily from Ottenby Bird Observatory. He suggested that most of the Wood Sandpipers migrate from Ottenby through central Europe to northern Italy and the Camargue, France, from where they continue southwestwards to West Africa.
At Ottenby, many studies have focused on population trends, morphometrics and migration phenology of waders (eg Edelstam Citation1972, Mascher & Markström Citation1976, Holmgren et al Citation1993a, Citationb, Waldenström & Lindström Citation2001, Blomqvist et al Citation2002, Hedenström Citation2004, Helseth et al Citation2005a, Citationb, Iwajomo & Hedenström Citation2011). Furthermore, for Ruddy Turnstones Arenaria interpres (Helseth et al Citation2005b) and juvenile Common Sandpipers Actitis hypoleucos (Iwajomo & Hedenström Citation2011) population declines have been reported. At the continental level, Sanderson et al Citation(2006), while analysing population trends in European breeding bird species, showed that the Wood Sandpiper declined in population within the period 1970 to 1990 but showed no significant change from 1990 to 2000. Considering the decrease in numbers reported for similar wader species at Ottenby it is important to investigate population trends in wader species such as the Wood Sandpiper passing through Ottenby, especially using age-related data that may reveal masked changes (Iwajomo & Hedenström Citation2011).
Stopover sites provide useful information about the strategies employed by bird species during migration (Alerstam & Lindström Citation1990, Weber & Houston Citation1997, Alerstam & Hedenström Citation1998, Weber et al Citation1998, Alerstam Citation2011). Since resources are not evenly distributed along a bird's migratory pathway, it must decide when and where to refuel for the journey ahead. A bird could choose either to migrate as fast as possible, not considering the cost of carrying large fuel loads – a strategy referred to as time minimisation – or keep flight cost as low as possible by storing only as much fuel as needed to carry it to the next stopover – the energy minimisation strategy (Alerstam & Lindström Citation1990).
Migration strategy varies between age classes, as documented for wader species including Dunlin Calidris alpina (Gromadzka Citation1989), Great Knot C. tenuirostris (Battley Citation2002) and Wood Sandpiper (Wichmann et al Citation2004). Also, a change in migration strategy with time of season has been documented in White-rumped Sandpipers C. fuscicollis (Harrington et al Citation1991) and juvenile Arctic waders (Lindström et al Citation2002). However, in spite of the large data set collected for Wood Sandpipers at Ottenby since 1946, the age-related migration strategy of this species has received little attention. In this study, we have analysed 65 years of ringing and recovery data of autumn-migrating Wood Sandpipers at Ottenby and consequently describe the age-related phenology, migration route, and morphometrics of the species. In addition, we have used the patterns in fuel load of all individuals and fuel deposition rates of recaptured individuals to describe the migration strategies of adults and juveniles.
METHODS
Ringing
Ringing of autumn migrating wader birds has been carried out annually at Ottenby Bird Observatory (56°12'N 16°24'E) since 1946. The observatory is located on the southernmost point of Öland, an island in the Baltic Sea, off southeastern Sweden. Wader trapping starts in late June and continues to the end of August or into September, depending on the number of staging waders in the area (but note that, in some years, some adult Wood Sandpipers may have arrived at and even departed from Ottenby before the wader trapping started). Around 100 walk-in funnel traps of the “Ottenby model” (Bub Citation1991), placed along the shore line, are used to trap the waders every autumn. The traps are checked every full hour from dawn to dusk. Trapped birds are brought in cloth bags to the ringing laboratory (within 200 m of the traps) where they are processed and thereafter released.
Morphometrics
The birds were aged as juvenile (first calendar year), second calendar year, or adult (third calendar year or older), following Prater et al Citation(1977). However, since only 134 second-calendar-year birds were trapped (and not a single one recaptured), we grouped them with other adults and thus use the terms juvenile and adult when discussing age. Morphometric data were available for all birds trapped since 1986, and included wing length to the nearest 1 mm, using the maximum-chord method (Svensson Citation1992), and body mass to the nearest 0.1 g using a Pesola spring balance or an electronic balance. In addition, total head length (bill + head) to the nearest 1 mm (Green Citation1980) has also been measured since 1996.
The lean body mass (LBM) was estimated at 53 g for adults and 50 g for juveniles, by averaging the 20 lightest birds with the mean wing length of 128 mm and 129 mm for adult and juveniles respectively, arbitrarily assuming that these birds represent a lean state. It should be stressed that the values given are averages and that there is variation in LBM between individuals. Accordingly, estimates of fuel loads of individual birds may be imprecise, whereas population averages should be more meaningful (Helseth et al Citation2005a). The estimated values are close to published values of lean birds during the autumn migration (eg 47–50 g, Persson Citation1998; 51 g, Kvist & Lindström Citation2003; 48.6 g, Minias et al Citation2010). The LBM was used to estimate fuel load in the captured birds (expressed as percent of the LBM) as (recorded body mass − LBM)/LBM. Fuel load was averaged over five-day periods and thereafter the seasonal trend was tested separately for adults and juveniles. Data from 53 individuals retrapped within the same season were available to estimate fuel deposition rates.
In analysing body mass of trapped individuals, we corrected for body size using total head length rather than wing length as an index of body size. This is because, although both wing length and total head length were significantly related to body mass, the latter accounted for more variance than the former in a model containing age and day of capture (day number, with day 1 being 1 June) and the interaction terms wing length x age or total head length x age (linear model: wing length, F4, 3929 = 92.05, P < 0.001, R2 = 0.08; total head length, F4, 1956 = 61.24, P < 0.001, R2 = 0.11). Consequently, analyses of body mass corrected for size were based on data from 1996–2011. The slopes of the relationship between body mass and total head length differed significantly between the age classes (linear model: interaction term total head length x age, F4, 1956 = 61.24, P = 0.04). In adults, body mass was significantly related only to total head length (linear model: F1, 386 = 9.54, P = 0.002) whereas in juveniles, body mass was significantly related to total head length and to date of capture (linear model: total head length, F2, 1570 = 138.2, P < 0.001; day, P < 0.001). Therefore, we corrected for body mass using the residuals of the equations:
body mass = 0.8 x total head length + 18.2 g for adult birds, and
body mass = 1.62 x total head length + 0.24 x day - 42.7 g for juveniles.
Refuelling rate was estimated as the difference in body mass of recaptured birds between their first and last capture, divided by the number of days between these capture events. The estimated values were thereafter expressed as percent of LBM. Birds recaptured the same day as they were ringed were excluded. Note that the length of stay we have estimated here is the minimum stopover duration (Cherry Citation1982). This is because we have assumed that the individual was first trapped the day it arrived at the site and was last retrapped on the day of its departure, so that the total stopover duration may have been underestimated. When analysing trends in median passage dates, we excluded those years in which fewer than five adults or juveniles were trapped from the analysis of that corresponding age class.
We estimated the potential flight range of adult and juvenile Wood Sandpipers with mean and maximum body mass (corresponding to mean and maximum fuel loads), following the method of Castro & Myers Citation(1989). According to the method, flight range (Y, in km) is given by the formula:
Y= 96.8 x U x L1.614 x (m0 −0.464 - m1 −0.464)
where U is flight speed (m/s), L is wing length measured from the carpal joint to the tip of the longest primary (cm), m0 is body mass at the end of the flight and m1 is body mass at departure. Estimates of m0 were assumed to be equal to the respective LBM values of adult and juvenile birds. Also following Hedenström (Citation2004, see Hedenström et al Citation2002), we assumed flight speed U = 13 m/s.
All regular statistical analyses were conducted using R 2.9.0 (R Development Core Team Citation2008). In every analysis we tested for the normality of our data as well as for homogeneity of variances using Shapiro–Wilks tests and Bartlett's test, respectively. Where our data met the assumptions of normality we used parametric tests such as one-way ANOVA, independent sample t-test and Pearson's correlation analysis; otherwise we have used non-parametric tests such as Welch's t-test, Kruskal–Wallis test and Spearman's rank correlation.
Recoveries
We analysed ring recoveries of birds ringed at Ottenby, and recaptures at Ottenby of birds ringed elsewhere, from the period 1937–2010. Average migration speed was derived from all birds ringed and recovered >20 km from the ringing site within 50 days. Recoveries of presumably wintering birds (dating from December or later) as well as recoveries of uncertain finding date were excluded. Migration speed, including fuelling periods, was calculated on an individual basis (‘method A’, Hildén & Saurola Citation1982) and as a population average, the latter by dividing the total sum of distance covered by all birds by the total sum of days which had passed between ringing and recovery. This is in accordance with ‘method B’ used by Hildén & Saurola Citation(1982).
Mean direction of autumn migration was calculated for all birds recovered during autumn (July–November) and tested with Rayleigh's test. Difference between adult and juveniles in migration direction was tested with the Watson–Williams F-test, and difference in concentration with the Mardia–Watson–Wheeler test. All circular statistics were calculated using Oriana software (Kovach Citation2003).
RESULTS
Numbers ringed
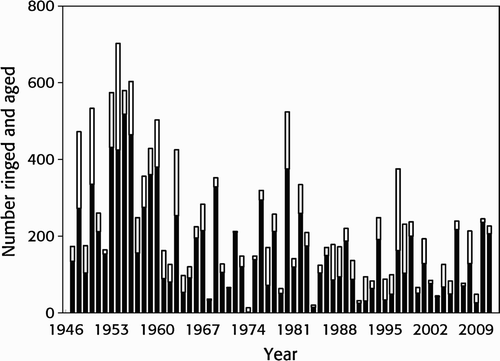
In total, 14,612 Wood Sandpipers were trapped, aged and ringed during 1947–2011. Of these 3,859 were aged as adults and 10,617 as juveniles, while the remaining 135 birds were not aged. On average 163 juveniles (sd = 122, range 2–518) and 59 adults (sd = 58.3, range 1–278) were trapped per year. The annual trapping of adults and juveniles was positively correlated (Spearman, rs = 0.48, P < 0.001, n = 65 trapping years; ). The total number of birds trapped showed a negative trend over the years (Spearman rs = -0.41, P = 0.001, n = 65 years), as did the specific age groups (Spearman: juvenile, rs = -0.44, P < 0.001, n = 65 years; adult, rs = -0.34, P = 0.006, n = 65 years).
Figure 1. Yearly total number of Wood Sandpipers Tringa glareola ringed and aged at Ottenby, Sweden, within the period 1947–2011. Juveniles are shown with solid bars and adult birds with open ones. Birds of unknown age have been excluded.
Phenology
Autumn passage of adults began on 16 June and juveniles appeared from 3 July. Autumn median passage date was 11 July for adults and 6 August for juveniles. Neither of the age classes showed any long-term trends in autumn median passage date (linear model: adult, F1, 62 = 0.487, P = 0.488; juvenile, F1, 59 = 0.741, P = 0.393).
Migration route and speed
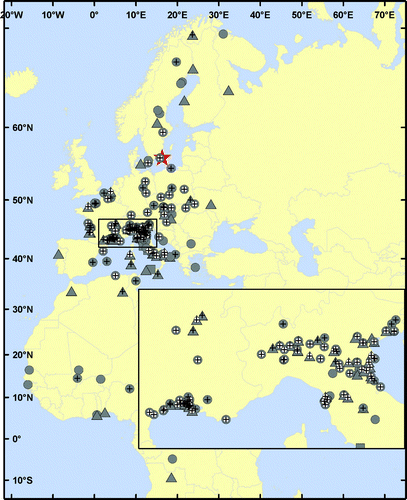
In total, there were 245 recoveries of Wood Sandpipers ringed at Ottenby during autumn, and eight recaptures during autumn at Ottenby of birds ringed elsewhere ().
Figure 2. Map of recoveries during autumn migration of Wood Sandpipers ringed at Ottenby Bird Observatory (n = 245) or ringing sites for Wood Sandpipers ringed during autumn and recovered at Ottenby Bird Observatory (n = 8). The age at the ringing/recovery site is indicated by triangles (adults), circles (juveniles) or squares (unknown). Records which met the requirements for calculations of migratory speed (see Methods) are indicated with a white circle centrally in the symbol, and those included in analyses of migratory direction (see Methods) are indicated with a black cross. Ottenby Bird Observatory is marked by a red star. The inset shows the area of N Italy and E France, where most recoveries were made. The map is drawn in Mercator projection, which displays true angles but distorts distances.
Only juvenile birds were available for speed calculations: the average southward migration speed was 58.1 km d−1 (n = 120), with the highest individual migration speed being 552 km d−1, recorded over three days. When restricting the selection, including no recoveries made later than 30 days after ringing, the average southward migration speed was 70.4 km d−1 (n = 94).
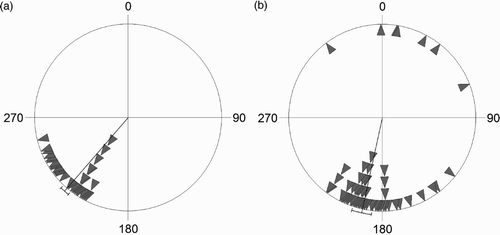
For juveniles the mean direction of autumn migration was 192.4° (circular sd = 33.7°), with a concentrated migration direction (concentration = 6.5; Rayleigh's test: Z = 87.1, P < 0.001, n = 123; ). The mean direction of adults on autumn migration was 220.8° (circular sd = 12.1°), with a highly concentrated migration direction (concentration = 23.0; Rayleigh's test: Z = 63.1, P < 0.001, n = 66). The age-dependent difference in migratory direction was significant (28.4°, Watson–Williams F1, 187 = 46.7, P < 0.001), and adults showed a significantly stronger concentration of direction (Mardia–Watson–Wheeler W = 103.4, P < 0.001).
Figure 3. Migratory direction during autumn of (a) adult (n = 66) and (b) juvenile (n = 123) Wood Sandpipers ringed at Ottenby. Each triangle represents an individual record, and the vector indicates mean migratory direction (adults 220.8° and juveniles 192.4°). The 28.4° difference in migratory direction is significant (Watson–Williams F1, 187 = 46.7, P < 0.001), as is the higher concentration of migratory direction in adults (Mardia–Watson–Wheeler W = 103.4, P < 0.001).
Morphometrics
There was no significant difference between the corrected body mass of adults (mean 63.0 g, sd 1.3 g) and juveniles (mean 62.7 g, sd 3.3 g; Welch t-test: t = -1.11, df = 1,627, P = 0.269). These means were, however, not significantly different from the means of the uncorrected values (paired t-test: adults, t = -1.41, df = 387, P = 0.161, uncorrected mean 63.6 g, sd 8.5 g, juveniles, t = -1.30, df = 1,572, P = 0.193, uncorrected mean 63.1 g, sd 8.6 g). The slopes of the trend in corrected body mass differed between the age classes (linear model: interaction term day x age, F3, 1974 = 473.8, P < 0.001). Over the season, the corrected body mass of adult birds showed a significant decline, whereas a significant increase was observed in juveniles (linear model, adults, F1, 390 = 8.726, P = 0.003, B = -0.02, n = 392; juveniles, F1, 1584 = 1,212, P < 0.001, B = 0.20, n = 1,586).
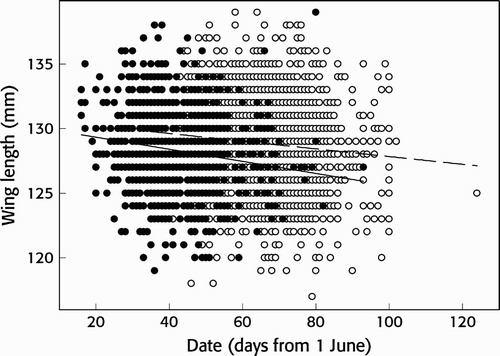
The wing length of juveniles was significantly longer than that of adult birds (juvenile mean 128.8 mm, sd 3.3 mm, n = 2,717, range 117–139; adult mean 128.3 mm, sd 3.0 mm, range 119–139, n = 1,217; Welch t-test: t = 4.84, df = 2538, P < 0.001). We tested for seasonal and long term trends in wing length of both age classes in a linear model with wing length as dependent variable, and day of capture, year and the interaction term year x day as explanatory variables. In adults, the interaction term was not significant (linear model: F3, 1213 = 19.39, P = 0.875) and so it was removed from the model and the main effects of day and year tested. Over the season there was a significant decline in the wing length of adult birds trapped (F2, 1212 = 29.09, P < 0.001, B = -0.04; ) but over the years there was a significant increase (F2, 1212 = 29.09, P < 0.001, B = 0.06). On the other hand, in juveniles, wing length of birds trapped declined significantly as the season progressed (F3, 2713 = 35.02, P = 0.034, B = -3.15; ). Although there was no significant change in wing length over the years (F3, 2713 = 35.02, P = 0.449), the long-term trend was significantly related to the date of capture (interaction term day x year, F3, 2713 = 35.02, P = 0.036).
Figure 4. Seasonal trend in wing length of a) adult (black circles, P < 0.001, B = -0.04) and b) juvenile (open circles, P = 0.034, B = -3.15) Wood Sandpipers Tringa glareola at Ottenby, Sweden.
Adults had a significantly longer total head length compared to juveniles (independent sample t-test, t = -5.15, df = 1,976, P < 0.001; adult mean 56.0 mm, sd 1.6 mm, range 52–64 mm; juvenile mean 55.5 mm, sd 1.6 mm, range 50–65 mm). There was no significant effect of the interaction term day x year on the total head length of adults (linear model: interaction term day x year, F3, 388 = 5.03, P = 0.076) or juveniles (F3, 1582 = 14.98, P = 0.118), and so we removed it from the model and tested the main effects of day and year in each age class. In juveniles, total head length declined over the season (linear model: F2, 1583 = 21.23, P < 0.001, B = -0.02) but increased over the years (linear model: F2, 1583 = 21.23, P = 0.037, B = 0.02). On the other hand, in adults, there was a significant decline over the season (linear model: F2, 389 = 5.93, P = 0.003, B = -0.02) but over the years there was no significant change (linear model: F2, 389 = 5.93, P = 0.079).
Fuel load, fuel deposition rate and length of minimum stopover
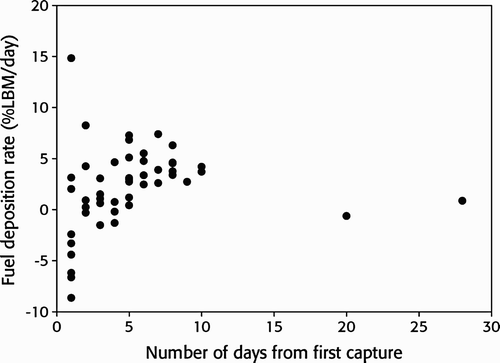
Juveniles had significantly larger fuel loads than adults (juvenile mean 24.3%, sd 17.2, adult mean 19.6%, sd 15.1; Welch t-test: t = 8.55, df = 2,632.5, P < 0.001). Maximum values were as high as 86% in juveniles and 82% in adults. The fuel load of juveniles increased over the season (Spearman's rank correlation: rs = 0.80, P < 0.001, n = 16) while that of adults showed no significant change (rs = 0.36, P = 0.184, n = 15; ). The mean time between first capture and last recapture did not differ significantly (Kruskal–Wallis χ 2 = 0.337, P = 0.563) between juveniles (5.2 days, sd 4.9 days, range 1–28, n = 49) and adult birds (3.5 days, sd 2.4 days, range 1–6, n = 4; ). The few adults recaptured more than one day after first capture stayed on average 4.3 days while juveniles stayed for 6.1 days. In total 40 (75%) of the 53 individuals recaptured within the same season increased in body mass while 13 (25%) reduced their body mass between the first capture and the last recapture (). The average mass change was higher in adults (2.5 g day−1, equivalent to 5.1% of LBM day−1, n = 4) than juveniles (0.7 g day−1, equivalent to 1.5% of LBM day−1, n = 49) but not significantly so (Kruskal–Wallis χ 2 = 0.524, P = 0.469). The maximum observed weight gain was also higher in adults (7.4 g day−1, equivalent to 14.8% of LBM day−1) than in juvenile birds (3.7 g day−1, equivalent to 8.2% of LBM day−1). We found no significant relationship between the length of stay of juveniles and their fuel load at the time of first capture (Spearman, rs = -0.015, P = 0.919, n = 49). In general, the fuelling rate of Wood Sandpipers at Ottenby was significantly related to their length of stay (Spearman, rs = 0.487, P <0.001, n = 53; ) and data were insufficient to test for age-specific relationships.
Figure 5. Seasonal trend in mean fuel load (% of lean body mass (LBM)) of a) adult (open circles, rs = -0.02, P = 0.425) and b) juvenile (black circles, rs = 0.22, P < 0.001) Wood Sandpipers Tringa glareola at Ottenby, Sweden. Mean values represent five-day averages (± 1 SE). Day 1 = 1 June.
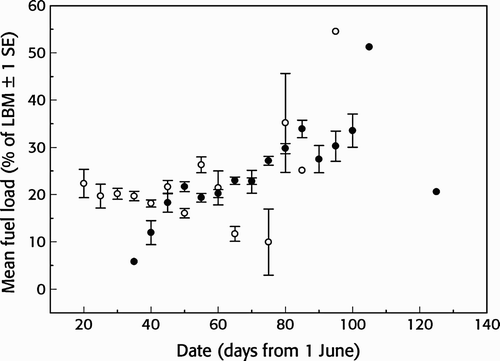
Figure 6. Fuel deposition rate (% of lean body mass (LBM) per day) in relation to the interval between first trapping and last recapture within the same season for Wood Sandpipers Tringa glareola at Ottenby, Sweden, during 1986–2011.
DISCUSSION
Numbers ringed and phenology of migration
The number of adult and juvenile Wood Sandpipers showed a negative trend over the period 1947–2011. This may be indicative of a decrease in numbers of the species owing to low breeding success or low survival, or perhaps fewer Wood Sandpipers have utilised Ottenby as a stopover site in recent years. A similar decline in population has been documented for the Ruddy Turnstone at Ottenby (Helseth et al Citation2005b).
Adult birds start to migrate through Ottenby earlier than juveniles even though there is considerable overlap later in the season. This pattern of migration is common in species which adopt the time minimisation strategy (Alerstam & Högstedt Citation1982, Włodarczyk et al Citation2007). Włodarczyk et al Citation(2007) reported that the adult Wood Sandpiper passage in central Poland begins 26 June, two weeks earlier than that of juveniles and that a similar interval was observed in the trapping of the last birds. In the present study, adult passage began 16 June, c 18 days earlier than that of juveniles and the interval between ringing the last adult and juvenile was about two weeks. Also, the difference between adult and juvenile median passage dates was 28 days. Previous studies of Wood Sandpiper autumn migration phenology at inland sites in central Europe have rendered contradictory results. Whereas Anthes Citation(2004) reported a delay in the median passage date at four wader-staging sites over the past three decades, Adamík & Pietruszková Citation(2008) found an advance in autumn passage in multiple sites in the Czech Republic and Slovakia between 1966 and 2002. Our study, however, did not conform to either of these reports, as we found no significant change in the timing of autumn migration at Ottenby, situated at the tip of an island.
Morphometrics
The average body mass (corrected for size) of adult and juvenile Wood Sandpipers trapped at Ottenby was lower than values published for birds trapped further south. For example, in Austria adult and juvenile body mass was 69.4 g and 65.9 g (Wichmann et al Citation2004), and in southern Italy it was 72.5 g and 66.9 g for adults and juveniles, respectively (Scebba & Moschetti Citation1996). Hence, it seems that Wood Sandpipers deposit more fat as they migrate southwards. That juvenile birds have significantly longer wings compared to adults may be largely due to the latter having old and worn feathers, given that many adults complete their post-breeding moult on their African wintering grounds (Ginn & Melville Citation1983). The observed pattern of decline in wing length, and total head length of both age classes as the season progressed, probably reflects that different populations, migrating at different times, differ in size or that larger individuals leave the breeding grounds earlier and so are trapped earlier in the season compared to the smaller individuals that arrive later. Also, in adult birds of this species, where both sexes show parental care, the females have been reported to depart after a few days of care so that the males carry on the responsibility (Cramp & Simmons Citation1983). With this, a sex-specific pattern should be observed in the migration of adult birds such that females, which are larger (Prater et al Citation1977, Cramp & Simmons Citation1983, Remisiewicz & Wennerberg Citation2006) and which leave the breeding ground first, would be trapped earlier in the season. We can only speculate that this is the situation for Wood Sandpipers passing Ottenby since birds ringed at the site were not sexed.
Minimum length of stay
The mean minimum length of stay of Wood Sandpipers at Ottenby, 3.5 days for adults and 5.2 days for juveniles, is similar to stopover periods recorded for other waders at Ottenby during autumn migration: the stopover period for adult Red Knots Calidris canutus was 2.8 days and 4.7 days for juveniles (Helseth et al Citation2005a), for Ruddy Turnstones 3.6 days (Helseth et al Citation2005b) and for Broad-billed Sandpipers Limicola falcinellus stopover period was 3.4 days (Waldenström & Lindström Citation2001). In Scania (southern Sweden) and central Poland, the average length of stay for Wood Sandpipers was 5.2 days (Persson Citation1998) and 5 days (Włodarczyk et al Citation2007) respectively. Wichmann et al Citation(2004) marked autumn-migrating Wood Sandpipers individually in northeastern Austria. They found that the median length of stay was 5 days for adults and 4.5 days for the juveniles. We acknowledge that only four adult birds were recaptured in this study: hence the minimum stopover may have been underestimated. In the present study a higher proportion of the juveniles was recaptured at Ottenby than of the adults. This, in addition to a longer mean individual minimum stopover length (up to 28 days in one individual), suggests that the juveniles stay for a longer time than the adults at Ottenby, unlike in Austria. We are aware that other factors can influence the recapture differences between the adults and juveniles in our data set. For example it has been suggested that adult Ruddy Turnstones are more experienced and avoid the traps, and thus are harder to recapture than juveniles (Meissner & Koziróg Citation2001). When using recapture data for estimating length of stay, the real stopover duration can be overestimated because birds that stay for a shorter period experience a lower probability of being recaptured. However, since other studies have used this method, the comparison should be adequate, and since we used an average for all birds the method ought to be useful at the population level, even if not at an individual level. Furthermore, the proportion of juveniles recaptured (4.8% at Ottenby) seems to be similar at other sites. In western Ukraine, 4% of the ringed juvenile Wood Sandpipers were caught more than once and most individuals stayed there for about two weeks (Struss Citation2011).
Fuelling rate and migration strategy
Persson Citation(1998) suggested that if the mass gain of 1.15 g day−1 in Scania represented increased fat stores alone it should enable Wood Sandpipers to fly directly to the Camargue or Gulf of Venice, which are important stopover areas before crossing the Mediterranean Sea, and maybe also the Sahara Desert. In this study we report average fuelling rates of 2.5 g day−1 and 0.7 g day−1 in adults and juveniles, respectively. It should be noted that the relatively higher fuelling rate in adult birds was estimated from only four individuals. Consequently the value should be treated with caution. The average fuelling rate of juvenile birds from this study is similar to the rate of mass gain reported for juveniles in Ukraine (0.78 g day−1; Struss Citation2011). Some limitations of estimating refuelling rate from recapture data include post-capture weight loss (Castro et al Citation1991, Atkinson et al Citation2007) and recapture probability owing to the variability in the refuelling rates of individuals, which could potentially underestimate the population refuelling rate (Minias & Kaczmarek Citation2013). Nonetheless, Minias & Kaczmarek Citation(2013) have shown that for the Wood Sandpipers, which show a non-synchronised stopover pattern and high turnover rate, estimates derived from recapture data are better than those derived from more advanced methods. Furthermore, the average fuel loads, 19.6% of LBM in adults and 24.3% of LBM in juveniles, were relatively low, considering that it can be as high as 100% of LBM for migrating waders (Alerstam & Lindström Citation1990). A similar pattern of low fuel loads has been recorded for other wader species at Ottenby during autumn migration ().
Table 1. Fuel load (% of LBM) of wader species trapped at Ottenby Bird Observatory, southeast Sweden, during autumn migration.
Theoretically, juveniles and adult birds with mean fuel loads may be able to cover a distance of 1,216 km and 974 km respectively. Based on these estimates, it seems that most individuals do not have sufficient fuel to carry them as far as the Camargue or Gulf of Venice in a single flight. Thus, these individuals will most likely make use of one or more European stopover sites after departing Ottenby. We also report that fuel load increased as the season progressed in juveniles but not in adults, as described in Red Knots (Helseth et al Citation2005a, Meissner & Kamont Citation2005) and Common Sandpipers (Iwajomo & Hedenström Citation2011). Wichmann et al Citation(2004) found strong evidence that juvenile Wood Sandpipers are more vulnerable to time and energy shortages during migration than are adults. In the present study, juveniles carried, on average, higher fuel loads than adults and also juveniles migrating later in the season had larger energy stores than individuals migrating earlier. These patterns suggest that juveniles, and in particular late individuals, may have been constrained by time; hence, they accumulated fat rapidly as is characteristic of time-minimisers. Conversely, the lower average fuelling rate of juvenile Wood Sandpipers in addition to longer mean stopover length at Ottenby suggests that the majority may use the energy-minimising strategy. Adults, on the other hand, had a relatively higher fuelling rate (one individual reaching up to 7.4 g day−1) and shorter mean stopover period, thus suggesting that they may use a time-selected migration strategy. The maximum fuelling rates obtained for both age classes, in addition to the significant relationship between fuel deposition rate and minimum length of stay, suggest that they may take advantage of good feeding opportunities and that energy assimilation capacities are as high as have been demonstrated in captive individuals (Kvist & Lindström Citation2003). According to Meissner Citation(1997) and Włodarczyk et al Citation(2007), Wood Sandpipers generally behave as time minimisers during their southward migration, but considerable variations may exist within and between seasons (Minias et al Citation2010). We conclude that, for individuals passing Ottenby, adults may be constrained by time rather than energy, whereas juveniles may adjust their strategy depending on prevailing conditions and the time of the season.
The migration speed of 58.1 km d−1 is lower than estimates from Finland (79 km d−1; Hildén & Saurola Citation1982) but comparable to calculations from Sweden (see Holmgren & Pettersson Citation1998, Fransson et al Citation2008). Compared with the migration speed of other wader species studied at Ottenby, eg 85.8 km d−1 for Red Knots (Helseth et al Citation2005a), the value estimated in our study is relatively low. The Wood Sandpipers at Ottenby migrate southward with mean directions of 220.8° and 192.4° in adults and juveniles respectively (). The adults thus follow a southwesterly route through Europe, while the juveniles migrate with a more southerly direction. The direction of the adults is highly concentrated, whereas the juveniles display a significantly wider scatter in direction, suggesting that experience is a key factor in adult migration while naive juveniles vary more in direction in their first southward journey. The difference in concentration of migratory direction is similar to findings of Holmgren & Pettersson Citation(1998). Even considering the age-dependent difference in direction, both age groups mainly follow the Mediterranean Flyway, as suggested by Myhrberg Citation(1961), rather than the East Atlantic Flyway used by most other wader species stopping over at Ottenby (eg Helseth et al Citation2005a, Citation2005b; but see Hedenström Citation2004, Fransson et al Citation2008, Iwajomo & Hedenström Citation2011). In particular, the Camargue area (France) and northern Italy seem to be frequently used as stopover sites ().
Acknowledgements
We are grateful to all the people who have been involved in bird trapping at Ottenby over the years. We would like to thank Arne Andersson who helped with data extraction. This is contribution number 271 from Ottenby Bird Observatory and number 64 from A.P. Leventis Ornithological Research Institute.
References
- Adamík , P. and Pietruszková , J. 2008 . Advances in spring but variable autumnal trends in timing of inland wader migration . Acta Ornithologica , 43 : 119 – 128 . (doi:10.3161/000164508X395225)
- Alerstam , T. 2011 . Optimal bird migration revisited . Journal of Ornithology , 152 : S5 – S23 . (doi:10.1007/s10336-011-0694-1)
- Alerstam , T. and Hedenström , A. 1998 . The development of bird migration theory . Journal of Avian Biology , 29 : 342 – 369 . (doi:10.2307/3677155)
- Alerstam , T. and Högstedt , G. 1982 . Bird migration and reproduction in relation to habitats for survival and breeding . Ornis Scandinavica , 13 : 25 – 37 . (doi:10.2307/3675970)
- Alerstam , T. and Lindström , Å. 1990 . “ Optimal bird migration: the relative importance of time, energy and safety ” . In Bird Migration , Edited by: Gwinner , E. Berlin : 331–351. Springer-Verlag .
- Anthes , N. 2004 . Long-distance migration of Tringa sandpipers adjusted to recent climate change . Bird Study , 51 : 203 – 211 . (doi:10.1080/00063650409461355)
- Atkinson , P. W. , Baker , A. J. , Bennett , K. A. , Clark , N. A. , Clark , J. A. , Cole , K. B. , Dekinga , A. , Dey , A. , Gillings , S. , González , P. M. , Kalasz , K. , Minton , C. D.T. , Newton , J. , Niles , L. J. , Piersma , T. , Robinson , R. A. and Sitters , H. P. 2007 . Rates of mass gain and energy deposition in red knot on their final spring staging site is both time- and condition dependent . Journal of Applied Ecology , 44 : 885 – 895 . (doi:10.1111/j.1365-2664.2007.01308.x)
- Battley , P. F. 2002 . Behavioural ecophysiology of migrating Great Knots , Brisbane , Australia : PhD thesis, Australian School of Environmental Studies, Griffith University .
- Blomqvist , S. , Holmgren , N. , Åkesson , S. , Hedenström , A. and Pettersson , J. 2002 . Indirect effects of lemming cycles on sandpiper dynamics: 50 years of counts from southern Sweden . Oecologia , 133 : 146 – 158 . (doi:10.1007/s00442-002-1017-2)
- Bub , H. 1991 . Bird Trapping & Bird Banding , New York : Cornell University Press .
- Castro , G. and Myers , J. P. 1989 . Flight range estimates for shorebirds . Auk , 106 : 474 – 476 .
- Castro , G. , Wunder , B. A. and Knopf , F. L. 1991 . Temperature dependent loss of mass of shorebirds following capture . Journal of Field Ornithology , 62 : 314 – 318 .
- Cherry , J. D. 1982 . Fat deposition and length of stopover of migrant White-crowned Sparrows . Auk , 99 : 725 – 732 .
- Cramp , S. and Simmons , K. E.L . 1983 . The birds of the Western Palearctic. Volume III, Waders to Gulls , Edited by: Cramp , S. and Simmons , K. E.L . Oxford : Oxford University Press .
- Edelstam , C. 1972 . The Visible Migration of Birds at Ottenby, Sweden. Vår Fågelvärld suppl. 7
- Fransson , T. , Österblom , H. and Hall-Karlsson , S. 2008 . Svensk ringmärkningsatlas , Vol. 2 , Stockholm : Naturhistoriska riksmuseet .
- Ginn , H. B. and Melville , D. S. 1983 . Moult in Birds. BTO Guide 19 , Tring : British Trust for Ornithology .
- Green , G. H. 1980 . Total head length . Wader Study Group Bulletin , 29 : 18
- Gromadzka , J. 1989 . Breeding and wintering areas of Dunlin migrating through southern Baltic . Ornis Scandinavica , 20 : 132 – 144 . (doi:10.2307/3676881)
- Harrington , B. A. , Leeuwenberg , F. J. , Resende , L. S. , McNeil , R. , Thomas , B. T. , Grear , J. S. and Martinez , E. F. 1991 . Migration and mass change of White-rumped Sandpipers in North and South America . Wilson Bulletin , 103 : 621 – 636 .
- Hedenström , A. 2004 . Migration and morphometrics of Temminck's Stint Calidris temminckii at Ottenby, southern Sweden . Ringing & Migration , 22 : 51 – 58 . (doi:10.1080/03078698.2004.9674310)
- Hedenström , A. , Alerstam , T. , Green , M. and Gudmundsson , G. A. 2002 . Adaptive variation of airspeed in relation to wind, altitude and climb rate by migrating birds in the Arctic . Behavioural Ecology and Sociobiology , 52 : 308 – 317 . (doi:10.1007/s00265-002-0504-0)
- Helseth , A. , Lindström , Å. and Stervander , M. 2005a . Southward migration and fuel deposition of Red Knots Calidris canutus . Ardea , 93 : 213 – 224 .
- Helseth , A. , Stervander , M. and Waldenström , J. 2005b . Migration patterns, population trends and morphometrics of Ruddy Turnstones Arenaria interpres passing through Ottenby in south-eastern Sweden . Ornis Svecica , 15 : 63 – 72 .
- Hildén , O. and Saurola , P. 1982 . Speed and autumn migration of birds ringed in Finland . Ornis Fennica , 59 : 140 – 143 .
- Holmgren , N. and Pettersson , J. 1998 . Recoveries of Swedish-ringed Wood Sandpipers Tringa glareola . The Ring , 20 : 91 – 93 .
- Holmgren , N. , Ellegren , H. and Pettersson , J. 1993a . Stopover length, body mass and fuel deposition rate in autumn migrating adult dunlins Calidris alpina evaluating the effects of moulting states and age . Ardea , 81 : 9 – 20 .
- Holmgren , N. , Ellegren , H. and Pettersson , J. 1993b . The adaptation of moult pattern in migratory dunlins Calidris alpina . Ornis Scandinavica , 24 : 21 – 27 . (doi:10.2307/3676405)
- Iwajomo , S. B. and Hedenström , A. 2011 . Migration patterns and morphometrics of Common Sandpipers Actitis hypoleucos at Ottenby, southeastern Sweden . Ringing & Migration , 26 : 38 – 47 . (doi:10.1080/03078698.2011.586509)
- Kolthoff , G. 1896 . Zur Herbstwanderung der nordischen Sumpfvögel über die Insel Öland , Upsala : Festschrift Wilhelm Lilljeborg .
- Kovach , W. L. 2003 . Oriana – Circular Statistics for Windows, v2 , Wales : Kovach Computing Services, Pentraeth .
- Kvist , A. and Lindström , A. 2003 . Gluttony in migratory waders – unprecedented energy assimilation rates in vertebrates . Oikos , 103 : 397 – 402 . (doi:10.1034/j.1600-0706.2003.12259.x)
- Lindström , Å. 2002 . “ Fuel deposition rates in migrating birds: causes, constraints and consequences ” . In Avian Migration , Edited by: P. , Berthold , E. , Gwinner and E. , Sonnenschein . Berlin : 307–320. Springer-Verlag .
- Mascher , J. W. and Markström , V. 1976 . Measures, weights, and lipid levels in migrating Dunlins Calidris alpina L. at the Ottenby Bird Observatory, South Sweden . Ornis Scandinavica , 7 : 49 – 59 . (doi:10.2307/3676174)
- Meissner , W. 1997 . Autumn migration of Wood Sandpipers Tringa glareola in the region of the Gulf of Gdansk . The Ring , 19 : 75 – 91 .
- Meissner , W. and Kamont , P. 2005 . Seasonal changes in the body size and body mass of Red Knots Calidris canutus during autumn migration through southern Baltic . Ornis Svecica , 15 : 97 – 104 .
- Meissner , W. and Koziróg , L. 2001 . Biometrics of Turnstone Arenaria interpres migrating in autumn through the Gulf of Gdan´sk region . Ornis Svecica , 11 : 181 – 188 .
- Minias , P. and Kaczmarek , K. 2013 . Population-wide body mass increment at stopover sites is an unreliable indicator of refuelling rates in migrating waders . Ibis , 155 : 102 – 112 . (doi:10.1111/ibi.12014)
- Minias , P. , Kaczmarek , K. , Włodarczyk , R. and Janiszewski , T. 2010 . Variation in autumn migration strategy in the first-year Wood Sandpipers Tringa glareola. Acta Ornithologica . 45 : 131 – 138 .
- Myhrberg , H. 1961 . Grönbenans (Tringa glareola) sträck genom Europa . Vår Fågelvärld , 20 : 115 – 145 .
- Persson , C. 1998 . Weight studies in Wood Sandpipers (Tringa glareola) migrating over south-western Scania in late summer and spring, with notes on related species . The Ring , 20 : 95 – 105 .
- Prater , A. J. , Marchant , J. H. and Vuorinen , J. 1977 . Guide to the Identification and Ageing of Holarctic Waders. BTO Guide 17 , Tring : British Trust for Ornithology .
- R Development Core Team . 2008 . R: a language and environment for statistical computing , Vienna , , Austria : R Foundation for Statistical Computing . www.R-project.org
- Remisiewicz , M. and Wennerberg , L. 2006 . Differential migration strategies of the Wood Sandpiper (Tringa glareola) – genetic analyses reveal sex differences in morphology and spring migration phenology . Ornis Fennica , 83 : 1 – 10 .
- Sanderson , F. J. , Donald , P. F. , Pain , D. J. , Burfield , I. J. and van Bommel , F. P.J. 2006 . Long-term population declines in Afro-Palearctic migrant birds . Biological Conservation , 131 : 93 – 105 . (doi:10.1016/j.biocon.2006.02.008)
- Scebba , S. and Moschetti , G. 1996 . Migration patterns and weight changes of Wood Sandpiper Tringa glareola in a stopover site in southern Italy . Ringing & Migration , 17 : 101 – 104 . (doi:10.1080/03078698.1996.9674124)
- Struss , I. 2011 . Migration of the Wood Sandpiper Tringa glareola in the Cholgini Ornithological Reserve, Ukraine . Wader Study Group Bulletin , 118 : 153 – 162 .
- Svensson , L. 1992 . Identification guide to European Passerines. Fourth edition , Stockholm : Lars Svensson .
- Waldenström , J. and Lindström , Å. 2001 . Migration and morphometrics of the Broad-billed Sandpiper Limicola falcinellus at Ottenby, southern Sweden, 1950–2000 . Ornis Fennica , 78 : 184 – 192 .
- Weber , T. P. and Houston , A. I. 1997 . Flight costs, flight range and the stopover ecology of migrating birds . Journal of Animal Ecology , 66 : 297 – 306 . (doi:10.2307/5976)
- Weber , T. P. , Ens , B. J. and Houston , A. I. 1998 . Optimal avian migration: a dynamic model of fuel stores and site use . Evolutionary Ecology , 12 : 377 – 401 . (doi:10.1023/A:1006560420310)
- Wichmann , G. , Barker , J. , Zuna-Kratky , T. , Donnerbaum , K. and Rössler , M. 2004 . Age-related stopover strategies in the Wood Sandpiper Tringa glareola . Ornis Fennica , 81 : 169 – 179 .
- Włodarczyk , R. , Minias , P. , Kaczmarek , K. , Janiszewski , T. and Kleszcz , A. 2007 . Different migration strategies used by two inland wader species during autumn migration, case of Wood Sandpiper Tringa glareola and Common Snipe Gallinago gallinago . Ornis Fennica , 84 : 119 – 130 .