Abstract
This study aimed to define the susceptibility of broilers to colibacillosis through quantification of clinical responses and to examine the relationship between susceptibility and growth retardation. A challenge experiment was carried out twice. In each trial, 192 chicks were challenged intratracheally with Escherichia coli (E. coli) at 7 days of age and 160 chicks served as controls. Surviving chicks were euthanized at 14 or 15 days. Parameters measured were: daily mortality, lesion scores, body weight at 1, 4, 7, 10, 12 and 14 or 15 days and feeding behaviour at 6, 11 and 13 days. The results were reproducible, and increasing susceptibility to colibacillosis was defined by four categories: chicks without lesions, chicks with airsacculitis but no systemic lesions, chicks with systemic lesions, and chicks that die. Increasing susceptibility was associated with increasing growth retardation, but growth retardation was not inevitably linked to challenge with E. coli.
Détermination de la sensibilité des poussins de chair à la colibacillose
Le but de cette étude a été la détermination de la sensibilité des poulets de chair à la colibacillose en quantifiant les réponses cliniques et en examinant la relation entre la sensibilité et le retard de croissance. Une expérimentation avec épreuve a été réalisée deux fois. Dans chaque essai, 192 poussins ont été éprouvés par voie trachéale avec Escherichia coli à l'âge de 7 jours et 160 poussins ont servi de témoin. Les poussins survivants ont été euthanasiés à l'âge de 14 ou 15 jours. Les paramètres étudiés ont été : la mortalité quotidienne, le score des lésions et le poids du corps à 1, 4, 7, 10, 12 et 14 ou 15 jours ainsi que le comportement alimentaire à 6, 11 et 13 jours. Les résultats ont été reproductibles, l'augmentation de la sensibilité à la colibacillose a été déterminée à l'aide de quatre critères : les poussins sans lésion, les poussins présentant une aérosacculite mais sans lésion systémique, les poussins avec des lésions systémiques et les poussins morts. L'augmentation de la sensibilité a été associée à l'augmentation du retard de croissance, mais ce dernier n'a pas forcément été lié à l'épreuve réalisée avec E. coli.
Bestimmung der Empfänglichkeit von Broilerküken für Kolibazillose
Ziel dieser Studie war es, die Empfänglichkeit von Broilern für Kolibazillose mittels Quantifizierung von klinischen Reaktionen zu bestimmen und die Beziehung zwischen Empfänglichkeit und Wachstumsverzögerung zu untersuchen. Ein Infektionsversuch wurde zweimal wiederholt. In jedem Versuch wurden 192 Küken am 7. Lebenstag intratracheal mit Escherichia coli infiziert, während 160 Küken als Kontrolltiere dienten. Überlebende Küken wurden am 14. oder 15. Tag euthanasiert. Als Parameter wurden die tägliche Mortalität, der Grad der Läsionen, die Körpergewichte am Tag 1, 4, 7, 10, 12 und 14 oder 15 sowie das Fressverhalten am 6., 11. und 13. Tag ermittelt. Die Ergebnisse waren reproduzierbar und es konnten vier Kategorien für eine zunehmende Empfänglichkeit gegenüber Kolibazillose definiert werden: Küken ohne Läsionen, Küken mit Aerosacculitis aber ohne systemische Alterationen, Küken mit systemischen Läsionen und Küken, die starben. Die zunehmende Empfänglichkeit war mit einer zunehmenden Wachstumsverzögerung assoziiert, aber die Wachstumsverzögerung stand nicht zwangsläufig mit der E. coli-Infektion im Zusammenhang.
Valoración de la susceptibilidad en pollos de engorde a la colibacilosis
El presente estudio pretendía determinar la susceptilidad en pollos de engorde a la colibacilosis mediante la cuantificación de las respuestas clínicas e investigar la relación entre susceptibilidad y retraso en el crecimiento. La infección experimental se repitió dos veces. En cada ensayo se infectaron 192 pollos vía intratraqueal con Escherichia coli a los 7 días de vida, y se utilizaron 160 pollos como controles. Los pollos que sobrevivieron se sacrificaron a los 14 o 15 días. Los parámetros evaluados fueron: mortalidad diaria, valoración de las lesiones, peso corporal a 1, 4, 7, 10, 12 y 14 o 15 días de vida, y conducta alimenticia a los 6, 11 y 13 días. Los resultados fueron reproducibles, y la susceptibilidad a la colibacilosis se definió en cuatro categorías: aves sin lesiones, aves con aerosaculitis pero sin lesiones sistémicas, aves con lesiones sistémicas y aves muertas. Se observó una asociación entre una mayor susceptibilidad y un mayor retraso en el crecimiento, aunque este último no se relacionó inevitablemente con la infección por E. coli.
Introduction
Colibacillosis, which is caused by Escherichia coli (E. coli), causes considerable economic and welfare problems in broilers (Goren, Citation1991; Bettelheim, Citation1994; Vandemaele et al., Citation2002b), due to its frequent occurrence and its adverse effects on growth and health. Clinical disease consists of respiratory signs, growth retardation, reduced feed intake and increased mortality (Goren, Citation1991; Vandemaele et al., Citation2002b). Airsacculitis and fibrinous polyserositis (e.g. pericarditis, perihepatitis and peritonitis) are the main gross lesions, and septicaemia, sometimes acute, is also common (Whiteman et al., Citation1989).
Good hygiene, vaccination and therapeutic treatment do not provide sufficient protection against colibacillosis. Vaccination against E. coli is limited to homologous protection, and the live vaccines used against the primary agents that provoke secondary colibacillosis may themselves act as primary agents. Therapeutic treatment is expensive, often does not result in sufficient recovery before slaughter and causes increased resistance of E. coli to antibiotics (Vandemaele et al., Citation2002a; Vandekerchove et al., Citation2004). Breeding for reduced susceptibility to colibacillosis may contribute to its prevention, but in order to do so a meaningful breeding goal and selection criteria must be defined. In other words, susceptibility to colibacillosis must be clearly defined in order to distinguish between more or less susceptible individuals. Previous studies do not provide sufficient information on the clinical responses, or the associations between responses, to provide a rational definition of susceptibility to colibacillosis, and assumptions have thus been made. A thorough investigation of responses to infection with E. coli was therefore considered necessary.
Previously, different assumptions have been made on the relative severity of lesions. Some studies have suggested a relationship between severity and the type of lesions, with systemic infections (pericarditis) being counted more severe than airsacculitis (Praharaj et al., Citation1996), and thus implying that chicks with systemic lesions are more susceptible than those with airsacculitis. This was based on the colibacillosis pathogenesis (i.e. systemic infection developing after initial infection of the respiratory system), and linked susceptibility with the infection stage. However, this does not necessarily directly reflect differences in susceptibility. Other studies have assumed that the relative severity of the lesions is not related to type of lesion and have reported lesion scores as population means based on the total scores of individuals (the sum of the lesion scores of all lesion types) (Peighambari et al., Citation2000; Matthijs et al., Citation2003). Increased knowledge on the relative severity of lesions is necessary to establish a meaningful definition of susceptibility to colibacillosis, and the association between lesions and growth retardation and feed intake.
Previous studies have shown that, on average, E. coli-infected chicks show growth retardation and that the amount of retardation is related to dose of infection (Dunnington et al., Citation1991; Maatman et al., Citation1993; Matthijs et al., Citation2003). However, these studies did not provide information on the association between growth retardation and lesion type or severity, which would facilitate a meaningful definition of susceptibility to colibacillosis. To the authors’ knowledge, there is also no information on the association between feed intake and mortality, lesion type or severity, which would also help to establish a meaningful definition of susceptibility to colibacillosis.
The aim of this study was therefore to define the susceptibility to colibacillosis in terms of mortality, lesions (airsacculitis, pericarditis and perihepatitis), growth and feeding behaviour (to provide information on feed intake) (Pym & Nichols, Citation1979; Nir et al., Citation1994). Colibacillosis was defined as clinical disease, and not merely infection with E. coli.
Materials and Methods
A challenge experiment was carried out on a population consisting of multiple broiler lines and crosses to ensure a broad genetic inference in the results. The experiment was performed twice in order to evaluate the reproducibility of the results. The results on the individual lines and crosses are the subject of another paper due to the large amount of data generated (Ask et al., Citation2006).
Chicks
Eggs were incubated and hatched for the two trials at the Spelderholt Poultry Research Centre in Beekbergen, The Netherlands. The eggs originated from six broiler lines and two crosses: two sire (A2, E3), three dam (A3, E4, E5), one slow-growing line (3), a sire cross (A2×E3) and a dam cross (A3×E4). In the text, the lines and crosses are referred to as genotypes.
Experimental design
At 1 day of age (day of hatch), the chicks were individually tagged and divided into a challenge and control group, each in four pens. In each trial, there were 192 chicks in the challenge group (48 chicks per pen), and 160 chicks in the control group (40 chicks per pen). Greater numbers were placed in the challenge group to anticipate losses due to mortality. Genotype and gender were equally represented: in each pen in the challenge group, there were three males and three females per genotype. In the control group, there were three males and two females per genotype in each of two of the four pens, and two males and three females per genotype in the other two pens.
At 7 days of age, all chicks in the challenge group were challenged (see later), and the experiment was terminated at approximately 2 weeks of age. The surviving chicks were stunned by electrocution and euthanized by bleeding: one-half of the birds in the challenge group and one-half of the control group were euthanized at 14 days of age, and the others at 15 days of age.
Housing
The challenge and control groups were kept on litter in separate, but identical, climate-controlled chambers to avoid horizontal infection of control chicks. Each pen measured 1.54×1.75 m2, with walls 0.5 m high. Feed and water was provided ad libitum. A daily schedule of 20:4 h light:dark was used, commencing with lights on at 06:00 h. Environmental temperature followed a standard schedule, starting at 34°C at 1 day of age followed by a gradual decline to 24°C at 15 days of age. The relative humidity was kept at 50%.
Challenge
All chicks in the challenge group were inoculated intratracheally with 0.3 ml of 1:100 phosphate-buffered saline solution of E. coli strain 506 (serotype O78K80) cultured in glucose peptone broth. E. coli 506 was a flumequine-resistant strain isolated from the inflamed pericardium of a commercial broiler suffering from natural colibacillosis (van Eck & Goren, Citation1991). The inoculation was carried out using a 1.0 ml syringe fitted with a blunt-ended pipette tip (catalogue number 4862; Corning, New York, USA) and the doses per bird were 106.0 colony-forming units in trial 1 and 105.8 colony-forming units in trial 2. In the first trial, 25 chicks from one pen were each given 0.5 ml E. coli inoculum (corresponding to 106.3 colony-forming units), but four chicks showed signs of suffocation within 15 min. The volume was therefore adjusted to 0.3 ml for the remainder of the chicks and those that had received 0.5 ml were omitted from the analyses. All chicks in the control group were inoculated intratracheally with 0.3 ml phosphate-buffered saline.
Recording of traits
The body weight of individual chicks was recorded at 1, 4, 7, 10 and 12 days of age. In addition, chicks in one half of the control and challenge pens were weighed at 14 days of age and the other half at 15 days of age. Mortality was recorded each morning.
Feeding behaviour was recorded by a single observer at 6, 11 and 13 days of age in both the control and challenge groups. Each pen was observed once in the morning (between 08:00 and 12:00 h) and once in the afternoon (between 13:00 and 17:00 h): the control pens were always observed first, and the challenge pens always last. An observation period lasted 20 min per pen and, at intervals of 40 to 60 sec, the identities of chicks that were eating (pecking at feed) were recorded. Individual identification was enabled through dorsal crayon stripe(s) of different colours. A different colour was used for each genotype, and to distinguish genotypes within a pen one to three stripes were applied; either from neck to tail (males) or from wing to wing (females). Observations made at 6 days of age in trial 1 were omitted from the analysis due to recording problems.
At postmortem examination gross lesions of the right and left thoracic air sac (airsacculitis), the heart (pericarditis) and the liver (perihepatitis) were scored macroscopically. Thoracic air sac lesions were considered to represent E. coli pathology of the respiratory tract, and pericarditis and perihepatitis were considered to represent systemic E. coli pathology. Lesion scoring was carried out blind and performed as described by van Eck & Goren (Citation1991) using the following scale: 0 = no lesions, 0.5 = one yellow or brown pinhead-sized spot indicative of inflammation, 1 = two or more pinhead-sized spots indicative of inflammation, 2 = thin layer of fibrinous exudate on various locations, and 3 = thick and extensive layer of fibrinous exudate.
Chicks that died during the experiment were dissected and examined macroscopically but lesions were not scored. Bacteriological examination was performed on the spleens of all chicks that died during the experiment, and the antibiotic sensitivity of the E. coli isolates was compared with that of E. coli strain 506 as described by Velkers et al. (Citation2005). The cause of death was considered to be colibacillosis if there were signs of airsacculitis, pericarditis or perihepatitis or if E. coli 506 could be isolated from the spleen.
Data analysis
Mortality and lesion traits were: incidence of mortality (in the challenge group, post-inoculation, as the percentage of total number of chicks at 7 days of age), lesions in the right and left thoracic air sac (RA and LA), pericarditis, perihepatitis, airsacculitis (the sum of RA and LA) and systemic lesions (the sum of pericarditis and perihepatitis). The feeding behaviour was defined as feeding or not feeding during each observation period, and the prevalence of chicks that did not show feeding behaviour was used as a trait. Observations of feeding behaviour on chicks that died during the experiment were also included in the analyses.
The Wilcoxon (or Kruskal–Wallis) test was used for comparisons of the incidence of mortality, the prevalence of lesions and the proportion of chicks not feeding. When sample sizes were not sufficiently large, Pearson's chi-square test was used for the comparisons.
Body weights at 1, 4, 7, 10 and 12 days of age were abbreviated as BW1, BW4, BW7, BW10 and BW12, respectively. The body weights at 14 or 15 days of age were treated as one trait, abbreviated as BW14. Body weights of chicks that died during the experiment were also included in the analyses. Body weights were adjusted by means of an analysis of variance, and the F test was applied to test for differences in body weights between trials. The model was:
The F test was also applied to test for differences in body weights between the challenge and control groups per trial, using the following model:
The t test was applied to test for differences between the control group and the following groups within the challenge group per trial: chicks without lesions, chicks with airsacculitis but no systemic lesions, chicks with systemic lesions, and chicks that died during the experiment. Tukey's adjustment (the Tukey–Kramer method) was used to correct for multiple comparisons. The model was:
Growth retardation was defined as:
Ethics
The experiment was approved by the Animal Ethics Committee (Dierexperimentencommissie, Utrecht University, The Netherlands), and chicks were handled accordingly. The Animal Ethics Committee based its decision on “De Wet op Dierproeven” (1996) and on the “Dierproevenbesluit” (1985) (http://www.nca-nl.org/).
Results
Mortality
Before the inoculation (1 to 7 days of age), eight chicks in the control group and three in the challenge group died, all for unknown reasons. In the control group, one chick died post inoculation, also for unknown reasons. Overall mortality in the challenge group amounted to 45 chicks in trial 1 and 62 in trial 2. Typical macroscopic E. coli pathology was present in all but 10 chicks in trial 1 that had died 1 day post inoculation, and it was present in all but six chicks in trial 2, of which five had died 1 day post inoculation and one died 6 days post inoculation. E. coli isolates that matched the sensitivity pattern of strain 506 were recovered from the spleens of all chicks that died during the experiment, except that from the chick in trial 2, which died 6 days post inoculation and did not show macroscopic E. coli pathology.
The total mortality incidence per trial is presented in and there was no significant difference between trials 1 and 2 (P = 0.586). The mortality per day in each trial is given in . The mortality in trial 1 showed a clear peak at 8 days of age (1 day post inoculation), and there was also a second, but less pronounced, peak between 11 and 13 days of age (4 to 6 days post inoculation). The mortality in trial 2 showed a clear peak at 13 days of age (6 days post inoculation) only. Significant differences in mortality pattern between trials were thereby apparent: at 1 day post inoculation (5.94% more mortality in trial 1 than 2, P < 0.001) and at 6 days post inoculation (5.35% less mortality in trial 1 than 2, P < 0.001).
Figure 1. Daily mortality of broiler chicks challenged with E. coli as a percentage of the total number of chicks at 7 days of age in trial 1 (black columns) and trial 2 (grey columns).
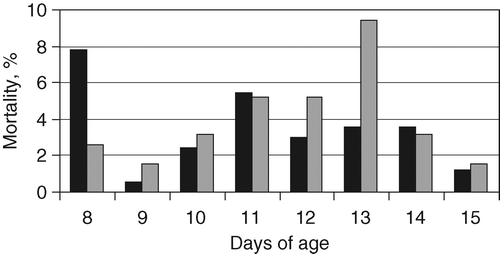
Table 1. Incidence of mortality, lesion prevalence (%) and scores of broiler chicks challenged with E. coli in two trials
Lesion traits
The prevalence of lesions in the challenge group per trial is presented in . No significant differences between trials were found in the prevalences of lesions. On average, 27% of the chicks developed colibacillosis expressed as lesions. No significant differences were found in the prevalence of airsacculitis or systemic lesions. There was a tendency to a higher prevalence of RA than LA in trial 2 (7%, P = 0.084), but not in trial 1, and no significant differences were found in the prevalence of pericarditis or perihepatitis. In both trials, the prevalence of systemic lesions, which was accompanied by at least one lesion (RA, LA, pericarditis or perihepatitis) of the highest severity (3), was significantly higher than the prevalence of airsacculitis accompanied by at least one lesion with the highest severity.
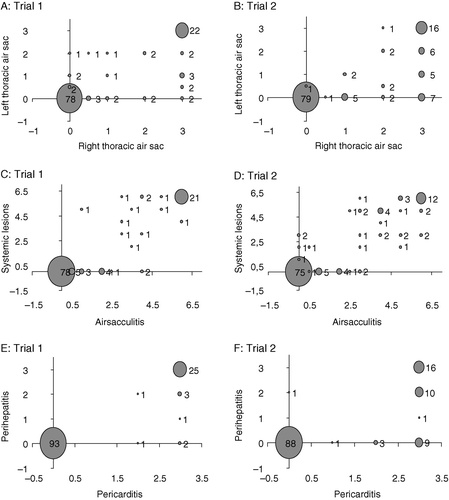
The relationship between the severity of RA and LA is illustrated in a,b. In both trials, the severity of RA and LA was related in a bimodal fashion: a high severity of LA was coupled with a high severity of RA, but not the other way around. c,d illustrates the relationship between the severity of airsacculitis and systemic lesions. In both trials, the severity of systemic lesions increased with increasing severity of airsacculitis. e,f shows the relationship between the severity of pericarditis and perihepatitis. In both trials, the severity of pericarditis and perihepatitis was related in a bimodal fashion: a high severity of perihepatitis was coupled with a high severity of pericarditis, but not the other way around.
Figure 2. Relationships between the severity of different types of lesions in broiler chicks surviving after challenge with E. coli. 2a, 2b: Relationship between the severity of lesions in the right and left thoracic air sac. 2c, 2d: Relationship between the severity of airsacculitis (lesions in the right and left thoracic air sac) and systemic lesions (pericarditis and perihepatitis). 2e, 2f: Relationship between the severity of pericarditis and perihepatitis.
Growth
At days 1, 4 and 7 (before the inoculation), the mean body weight of the control group was between 2% higher and 4% lower than that of the challenge group in trial 1 and between 0 and 3% lower than that of the challenge group in trial 2 (P < 0.100).
a (trial 1) and b (trial 2) show the body weight trends of the control and challenge group post inoculation. There was a lower body weight trend in the challenge group than in the control group. presents the growth retardation in the challenge group. The final growth retardation (at 14 days of age) was 11% in trial 1 and 8% in trial 2.
Figure 3. Body weight trends of broiler chicks challenged with E. coli based on the mean body weight (g) at 7, 10, 12 and 14 days of age. 3a, 3b: In the control group (♦) and the challenge group (▪). 3c, 3d: In the control group (♦) and in challenged birds grouped according to the presence and type of lesions (▪, no lesions; ▴, airsacculitis only; •, systemic lesions; ×, chicks that died). Standard errors, indicated by vertical bars, are too small to be seen at this scale. Observations on chicks that died during the experiment are included.
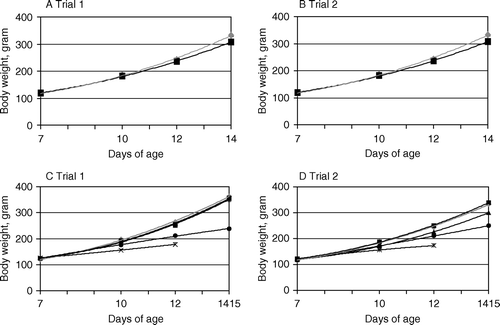
Table 2. Growth retardation (%) of broiler chicks challenged with E. coli relative to control group in two trials
c (trial 1) and d (trial 2) illustrate the body weight trends post inoculation for the control group and the challenge group divided into subgroups according to the presence and type of lesions. The body weight trends of the chicks with systemic lesions and the chicks that died during the experiment were generally lower than that of the control group. The body weight trend for the chicks without lesions, and with airsacculitis but no systemic lesions, was almost identical to that for the control group. presents the average growth retardation in the challenge group divided into subgroups according to the presence and type of lesions. The average growth retardation of the chicks without lesions, and with airsacculitis but no systemic lesions, was not significant at any age, except for that of the chicks with airsacculitis but no systemic lesions at 14 days of age in trial 2. The average growth retardation of the chicks with systemic lesions increased with time post inoculation: from 12% at 10 days of age (P < 0.001) to 34% at 14 days of age (P < 0.001) in trial 1, and from 9% at 10 days of age (P = 0.001) to 25% at 14 days of age (P < 0.001) in trial 2. The growth retardation of the chicks that died during the experiment increased with time post inoculation, resulting in an average growth retardation at 12 days of age of 33% in trial 1 and 29% in trial 2.
Feeding behaviour
presents the proportion of chicks that did not show feeding behaviour in the control and challenge groups, in total and divided into subgroups according to the presence and type of lesions. At 6 days of age, there was no significant difference between the control and challenge groups in the proportion of chicks that did not feed. At 11 and 13 days of age, the proportion of chicks that did not feed was between 11 and 18% higher in the challenge group than in the control group. In most cases, the proportion of chicks that did not feed in the control group was similar (not significantly different) to the group without lesions, or with airsacculitis but no systemic lesions. In contrast, the proportion of chicks that did not feed in the control group was in most cases much smaller than in the group with systemic lesions or that died during the experiment (between 27.3 and 60.9%; all significant with P<0.05).
Table 3. Prevalence (%) of broiler chicks, challenged with E. coli in two trials, that did not show feeding behaviour
Discussion
In the present study, the responses for mortality, lesions and growth retardation were reproducible as the differences between trials were small. The response in feeding behaviour, an increase in the prevalence of chicks not feeding from category 1 to 4, was also reproducible.
The observed mortality was assumed to be due to colibacillosis because of the macroscopic E. coli pathology, and recovery of E. coli from the spleen. The prevalence of systemic lesions and mortality in the present study were both high relative to natural colibacillosis (Matthijs et al., Citation2003), which is probably related to the relatively high inoculation dose and volume and the procedure (intratracheal inoculation rather than aerosol exposure) (Maatman et al., Citation1993; Peighambari et al., Citation2000). This discrepancy might appear to be in conflict with a meaningful definition of susceptibility to colibacillosis, but it actually does not pose a problem since it is the type and combination of responses (typical clinical signs or not) that matter rather than the magnitude of the response on a population level.
The surviving chicks without lesions did not show any growth retardation ( and c,d) or reduce their feeding behaviour (). It is therefore reasonable to assume that these chicks reflected a category of the lowest susceptibility to colibacillosis.
The group of chicks that died due to colibacillosis showed the highest level of growth retardation (where measurable; c,d), and they also reduced their feeding behaviour considerably (). This situation categorized the highest susceptibility to colibacillosis. There were also indications that there were differences in susceptibility among the chicks that died. The mortality occurred in one or two peaks (), in accordance with previous studies (Ardrey et al., Citation1968; Matthijs et al., Citation2003), and is probably related to the cause of death. The mortality peak observed at 8 days of age in trial 1 was assumed to be associated with acute septicaemia, which is characterized by E. coli in the spleen, because E. coli was indeed recovered from the spleens and no macroscopic E. coli pathology was observed. The mortality peak between 11 and 13 days of age was assumed to be associated with fibrinous polyserositis, because of the presence of lesions. Acute septicaemia may reflect higher susceptibility than does fibrinous polyserositis, because of the earlier time of death. Such ranking is not necessarily valid, however, because the causes of death may reflect different types of susceptibility rather than different magnitudes. Possible differences in susceptibility between acute septicaemia and fibrinous polyserositis could not be supported by differences in growth retardation or feeding behaviour, because neither could be measured in the chicks dying of acute septicaemia. Subdivision of mortality into two categories of different susceptibility could therefore not be supported.
The surviving chicks with gross lesions clearly reflected categories of intermediate susceptibility, but the ranking among chicks with different types or severity of lesions was not obvious. The most severe lesion scores (3) were associated with the presence of systemic lesions, because this score was virtually absent in chicks that showed airsacculitis but no systemic lesions (). This indicates, as previously suggested by Praharaj et al. (Citation1996), that systemic lesions reflect a higher susceptibility to colibacillosis than airsacculitis. High lesion scores were also given for airsacculitis (a,b), but these were also associated with systemic lesions (c,d), and it therefore cannot be excluded that severe lesions of any type reflect a severity equal to that of systemic lesions. Support for categorizing chicks with systemic lesions as being more susceptible than chicks with airsacculitis but no systemic lesions was clearly provided for by the differences in growth retardation and in feeding behaviour between the two groups ( and , and c,d). Therefore, it was considered reasonable to assume that the chicks with airsacculitis but no systemic lesions reflected a category of low, intermediate susceptibility to colibacillosis, and that the chicks with systemic lesions reflected a category of high, intermediate susceptibility.
The absence of growth retardation in the chicks without lesions and those with airsacculitis but no systemic lesions is important because it suggests that, despite the general finding that colibacillosis leads to growth retardation, growth retardation is not inevitably linked to challenge with E. coli. In theory, all chicks infected with E. coli are expected to mount an immune response, which results in reduced appetite, and thereby decreased feed intake and energy availability for growth. An immune response will also cause reallocation of resources from body reserves towards defence mechanisms (Klasing & Johnstone, Citation1991; Sonti et al., Citation1996).
There are at least two plausible explanations for why the chicks without lesions and the chicks with airsacculitis but no systemic lesions showed neither reduced feeding behaviour nor growth retardation. The first explanation is that these chicks did not elicit an immune response, or release cytokines at concentrations reaching a level that inhibits feeding behaviour (Sonti et al., Citation1996). The presence of maternal antibodies or physical factors (e.g. ciliary activity, mucus barrier) may have provided sufficient protection in these chicks rendering an immune response superfluous. Differences in maternal antibody level are probable in the present study, because the parent stock of one of the genotypes was kept in a different environment and was 20 weeks older than the others, and chicks from older parents are expected to have more maternal antibodies (Jeurissen et al., Citation2000; Parmentier et al., Citation2004). The second plausible explanation is that these chicks elicited an immune response, but their satiety and hunger mechanisms were malfunctioning and/or they continued to preferentially allocate energy towards growth rather than towards defence mechanisms. This has previously been suggested to occur in broilers (Denbow, Citation1994; Qureshi & Havenstein, Citation1994; Reddy et al., Citation2002). In order to confirm or reject these possibilities, it would be necessary to investigate immunological parameters in such chicks.
In conclusion, susceptibility to colibacillosis could be meaningfully defined in four categories with increasing susceptibility: chicks without lesions, chicks with airsacculitis but no systemic lesions, chicks with (severe) systemic lesions, and chicks that died.
Acknowledgments
The authors gratefully acknowledge Hybro B.V., Boxmeer, The Netherlands, for providing the broiler lines, and thank our colleagues and friends who helped with the experiments. A special thanks also goes to the animal caretakers, and to Sander van Voorst and Jan Hoekman for their technical support and guidance experimental planning.
References
- Ardrey , W.B. , Peterson , C.F. and Haggart , M. 1968 . Experimental colibacillosis and the development of carriers in laying hens . Avian Diseases , 12 : 505 – 511 .
- Ask , B. , van der Waaij , E.H. , Stegeman , J.A. & van Arendonk , J.A.M. (2006) . Genetic variation between broiler genotypes in susceptibility to colibacillosis . Poultry Diseases , 85 , 415 – 421 .
- Bettelheim , K.A. 1994 . “ Biochemical characteristics of Escherichia coli ” . In Domestic Animals and Humans , Edited by: Gyles , C.I. 3 – 29 . Wallingford : CABI Publishing .
- Denbow , D.M. 1994 . Peripheral regulation of food intake in poultry . Journal of Nutrition , 124 : 1349S – 1354S .
- Dunnington , E.A. , Siegel , P.B. and Gross , W.B. 1991 . Escherichia coli challenge in chickens selected for high and low antibody response and differing in haplotypes at the major histocompatibility complex . Avian Diseases , 35 : 937 – 940 .
- Goren , E. 1991 . Colibacillose bij pluimvee: etiologie, pathologie en therapie . Tijdschrift voor Diergeneeskunde , 116 : 1122 – 1129 .
- Jeurissen , S.H.M. , Boonstra-Blom , A.G. , Cornellisen , J.B.W.J. & Dijkstra , G. (2000) . Effects of antigen-specific maternal immunity on the induction of responses to the same antigen in the offspring . In Proceedings of the 6th Avian Immunology Research Group Meeting (pp. 83 – 89 ). Ithaca, NY, USA .
- Klasing , K.C. and Johnstone , B.J. 1991 . Monokines in growth and development . Poultry Science , 70 : 1781 – 1789 .
- Maatman , R. , Gross , W.B. , Dunnington , E.A. , Larsen , A.S. and Siegel , P.B. 1993 . Growth, immune response and behaviour of broiler and leghorn cockerels fed different methionine levels . Archief voor Geflügelkunde , 57 : 249 – 256 .
- Matthijs , M.G.R. , van Eck , J.H.H. , Landman , W.J.M. and Stegeman , J.A. 2003 . Ability of Massachusetts-type infectious bronchitis virus to increase colibacillosis susceptibility in commercial broilers: a comparison between vaccine and virulent field virus . Avian Pathology , 32 : 473 – 481 .
- Nir , I. , Twina , Y. , Grossman , E. and Nitsan , Z. 1994 . Quantitative effects of pelleting on performance, gastrointestinal tract and behaviour of meat-type chickens . British Poultry Science , 35 : 589 – 602 .
- Parmentier , H.K. , Lammers , A. , Hoekman , J.J. , De Vries Reilingh , G. , Zaanen , I.T. and Savelkoul , H.F. 2004 . Different levels of natural antibodies in chickens divergently selected for specific antibody responses . Developmental and Comparative Immunology , 28 : 39 – 49 .
- Peighambari , S.M. , Julian , R.J. and Gyles , C.L. 2000 . Experimental Escherichia coli respiratory infection in broilers . Avian Diseases , 44 : 759 – 769 .
- Praharaj , N.K. , Gross , W.B. , Dunnington , E.A. and Siegel , P.B. 1996 . Feeding regimen by sire family interactions on growth, immunocompetence, and disease resistance in chickens . Poultry Science , 75 : 821 – 827 .
- Pym , R.A.E. and Nichols , P.J. 1979 . Selection for food conversion in broilers: direct and correlated responses to selection for body-weight gain, food consumption and food conversion ratio . British Poultry Science , 20 : 73 – 86 .
- Qureshi , M.A. and Havenstein , G.B. 1994 . A comparison of the immune performance of a 1991 commercial broiler with a 1957 randombred strain when fed “typical” 1957 and 1991 broiler diets . Poultry Science , 73 : 1805 – 1812 .
- Reddy , M.R. , Panda , A.K. , Praharaj , N.K. , Rama Rao , S.V. , Chaudhuri , D. and Sharma , R.P. 2002 . Comparative evaluation of immune competence and disease resistance in dual purpose chicken Vanaraja and Gramapriya vis-à-vis coloured synthetic broiler . Indian Journal of Animal Sciences , 72 : 6 – 8 .
- Sonti , G. , Ilyin , S.E. and Plata-Salaman , C.R. 1996 . Anorexia induced by cytokine interactions at pathophysiological concentrations . American Journal of Physiology , 270 : R1394 – R1402 .
- Vandekerchove , D. , de Herdt , P. , Laevens , H. , Butaye , P. , Meulemans , G. and Pasmans , F. 2004 . Significance of interactions between Escherichia coli and respiratory pathogens in layer hen flocks suffering from colibacillosis-associated mortality . Avian Pathology , 33 : 298 – 302 .
- Vandemaele , F. , Vereecken , M. , Derijcke , J. and Goddeeris , B.M. 2002a . Incidence and antibiotic resistance of pathogenic Escherichia coli among poultry in Belgium . Veterinary Record , 151 : 355 – 356 .
- Vandemaele , F. , Assadzadeh , A. , Derijcke , J. , Vereecken , M. and Goddeeris , B.M. 2002b . Aviaire pathogene Escherichia coli (APEC) . Tijdschrift voor Diergeneeskunde , 127 : 582 – 588 .
- Van Eck , J.H.H. and Goren , E. 1991 . An Ulster 2C strain-derived Newcastle disease vaccine: vaccinal reaction in comparison with other lentogenic Newcastle disease vaccines . Avian Pathology , 20 : 497 – 507 .
- Velkers , F.C. , Loo , A.J. , Madin , F. and van Eck , J. H.H. 2005 . Isopathic and pluralist homeopathic treatment of commercial broilers with experimentally induced colibacillosis . Research in Veterinary Science , 78 : 77 – 83 .
- Whiteman , C.E. , Bickford , A.A. and Barnes , H.J. 1989 . Avian Disease Manual , 3rd edn , Dubuque, IA : Kendall/Hunt .