Abstract
The purpose of the present study was to measure any functional differences in peripheral blood heterophils isolated from a commercial turkey line to wild-type Rio Grande turkeys. The phagocytosis of Salmonella enteritidis, oxidative burst (OXB) and degranulation (DGR) were used as parameters of heterophil functional efficiency in these studies. Blood was collected and heterophils isolated from each line of turkeys at days 4, 7, and 14 post-hatch. On days 4 and 7 post-hatch, heterophils from Rio Grande turkeys responded to phorbol A-myristate-13-acetate with significantly greater OXB activity than commercial line A. Results from the DGR assay also revealed a greater level of activity in Rio Grande heterophils when compared with heterophils from Line A turkeys. On day 14 post-hatch, heterophils from the commercial line A responded at similar or greater levels than Rio Grande turkey heterophils in the OXB and DGR assays. No differences in the phagocytosis of S. enteritidis were observed between the lines. These results indicate that the commercial Line A turkeys may be at an immunological disadvantage during the first days post-hatch when compared with their wild-type predecessors. Based on the results of these experiments, research into the differences and similarities between the innate immune response of commercial turkey lines and wild-type turkeys may illuminate areas where commercial lines can be improved to decrease losses due to disease and to decrease pathogen contamination of turkey products while preserving performance characteristics.
Comparaison des fonctions des hétérophiles d'une souche commerciale de dindes et du type sauvage Rio Grande
Le but de cette étude a été de mesurer les différences fonctionnelles des hétérophiles du sang périphérique isolés d'une lignée commerciale de dindes et du type sauvage Rio Grande. Les paramètres d'étude de l'efficacité de la fonction hétérophile ont été: la phagocytose de Salmonella enteritidis (SE), la décharge oxydative (OXB) et la dégranulation (DGR). Le sang a été prélevé et les hétérophiles ont été isolés de chaque lignée de dinde les 4ème, 7ème et 14ème jours après éclosion. Les hétérophiles de la souche de dinde Rio Grande, récoltés les 4ème et 7ème jours après éclosion ont répondu au phorbol A-myristate 13 acetate (PMA) avec une activité OXB significativement plus élevée que ceux de la lignée commerciale A. Les résultats du test DGR ont également révélé un niveau plus élevé de l'activité des hétérophiles de la souche de dinde Rio Grande comparés à ceux de la lignée A de dinde. Les hétérophiles de la lignée commerciale A récoltés le 14ème jour après éclosion ont répondu à des niveaux similaires ou supérieurs comparés à ceux des hétérophiles de la souche Rio Grande pour les tests OXB et DGR. En ce qui concerne la phagocytose de SE aucune différence entre les souches n'a été observée. Ces résultats indiquent que la lignée commerciale A de dinde peut être désavantagée sur le plan immunologique durant les premiers jours après l'éclosion par comparaison avec la souche de type sauvage d'origine. A partir de ces résultats, la recherche des différences et des similarités entre les réponses immunitaires innées des lignées commerciales et du type sauvage de dinde peut éclairer des domaines où les lignées commerciales peuvent être améliorées pour diminuer les pertes dues aux maladies et diminuer la contamination des produits de dinde par des agents pathogènes tout en gardant les caractéristiques de performance.
Vergleich der Funktion von Heterophilen bei modernen kommerziellen und Wildtyp-Rio-Grande-Puten
Ziel dieser Studie war es, funktionelle Unterschiede zwischen peripheren Blut-Heterophilen aus einer kommerziellen Putenlinie und aus Wildtyp-Rio-Grande-Puten zu ermitteln. Als Parameter für die funktionelle Wirksamkeit von Heterophilen wurden in dieser Untersuchung die Phagozytose von Salmonella enteritidis (SE), oxidativer Ausbruch (OXB) und Degranulation (DGR) herangezogen. 4, 7 und 14 Tage nach dem Schlupf wurden von jeder Putenlinie Blutproben entnommen und die Heterophilen isoliert. Am 4. und 7. Tag nach dem Schlupf antworteten die Heterophilen aus den Rio-Grande-Puten auf Phorbol-A-Myristat-13-Acetat (PMA) mit einer signifikant höheren OXB-Aktivität als die kommerzielle Linie A. Ebenso ließen die Ergebnisse des DGR-Tests in den Heterophilen der Rio-Grande-Puten im Vergleich zu den Heterophilen der Linie-A-Puten einen höheren Aktivitätswert erkennen. Am 14. Lebenstag reagierten die Heterophilen aus der kommerziellen Linie A im OXB- und DGR-Test mit ähnlichen oder höheren Werten als die Heterophilen aus den Rio-Grande-Puten. Zwischen den beiden Linien wurden keine Unterschiede in der Phagozytose von SE festgestellt. Diese Ergebnisse weisen darauf hin, dass Puten der kommerziellen Linie A im Vergleich zu ihren Wildtyp-Vorfahren in den ersten Lebenstagen immunologisch benachteiligt sind. Basierend auf diesen Untersuchungsergebnissen könnte die weitergehende Erforschung von Unterschieden und Übereinstimmungen in der angeborenen Immunantwort von kommerziellen und Wildtyp-Puten Bereiche erkennen lassen, wo kommerzielle Linien verbessert werden könnten, um krankheitsbedingte Verluste und die Kontamination von Geflügelprodukten mit Infektionserregern bei gleichzeitiger Beibehaltung von Leistungsmerkmalen zu reduzieren.
Comparación de las funciones heterofílicas en pavos comerciales actuales y pavos salvajes de Río Grande
El objetivo de este estudio fue valorar cualquier diferencia funcional de los heterófilos en sangre periférica aislados de líneas comerciales de pavos y de pavos salvajes de Río Grande. En este estudio se utilizaron como parámetros de la eficiencia funcional de los heterófilos la fagocitosis de Salmonella enteritidis (SE), el metabolismo oxidativo (OXB) y la degranulación (DGR). Se obtuvo sangre y se aislaron los heterófilos de cada línea de pavos a los 4, 7 y 14 días post nacimiento. Los heterófilos de pavos de Río Grande respondieron con una actividad OXB significativamente mayor al acetato de forbol A-miristato 13 (PMA) en comparación con la línea comercial A. Los resultados del ensayo de DRG también mostraron un nivel más elevado de actividad en los heterófilos de pavos de Río Grande en comparación con los heterófilos de los pavos de la Línea A. A los 14 días post nacimiento, los heterófilos de la línea comercial A respondieron en los ensayos de OXD y DGR con niveles iguales o mayores que los heterófilos de los pavos de Río Grande. No se observaron diferencias en la fagocitosis de SE entre ambas líneas. Los resultados indicaron que la Línea A de pavos comercial podría estar en desventaja inmunológica durante los primeros días post-nacimiento respecto a sus predecesores salvajes. En base a los resultados obtenidos en estos experimentos, parece ser que la investigación de las diferencias y similitudes entre la respuesta inmune innata de las líneas de pavos comerciales y salvajes podría resaltar aquellas áreas en las que las líneas comerciales podrían mejorarse para reducir las pérdidas debidas a enfermedades y las contaminaciones de los productos de pavos con patógenos preservando al mismo tiempo sus características productivas.
Introduction
The innate arm of the host immune system plays a vital role in encounters with microbes. The innate system is the primary responder to initial contact with microbes, responsible for the initial control and elimination of microbial invaders that have breached the host's physical defenses. Through this initial interaction the host engages the immune machinery necessary for immediate host defense and the development of an acquired response for long-term protection against re-infection with microorganisms. The interactions between cells of the innate immune system and microbes depends on the host's recognition of highly conserved molecules that are unique to a certain class of microbes called pathogen-associated molecular patterns (Fearon & Locksley, Citation1996; Akira, Citation2001; Janeway & Medzhitov, Citation2002). These pathogen-associated molecular patterns are recognized by the cells of the innate system via pattern recognition receptors, such as the Toll-like receptors, and this recognition initiates signaling pathways within the cell that result in the production and release of cytokines, chemokines, and activation of the cell itself (Imler & Hoffman Citation2001; Janeway & Medzhitov, Citation2002; Kopp & Medzhitov, Citation2003). The recognition and subsequent release of cytokines then directs the acquired arm of the immune system, leading the response towards either a TH1, cell-mediated response or a TH2, antibody-mediated response (Medzhitov & Janeway, Citation1997a Citationb; Romagnani, Citation1992).
Observations in chickens and turkeys have revealed the host immune response has been negatively effected by selection for larger, heavier bodied, faster growing birds with highly improved feed efficiencies (Quereshi & Havenstein, Citation1994; Nestor et al., Citation1999; Li et al., Citation1999a Citationb). The traits of the modern commercial chicken and turkey allow producers to harvest larger, market-ready birds in less time than could be done previously. However, apparently the selection of commercial birds with these growth characteristics has led to a subsequent increase in susceptibility to disease and to physical maladies that have not been solved by selective breeding. In addition, the physical conditions affecting turkey livability and morbidity may also be related to selection against birds with active innate immune systems. Li et al. (Citation2001) have shown that turkeys selected for higher body weights at 16 weeks of age are different to random-bred turkeys from the same lines in some aspects of immune function. These differences included decreased non-specific phagocytic capabilities, as well as increased populations of CD4+/CD8− T cells, and an increase in mortality from bacterial infections as compared with random-bred turkeys from the same lines (Li et al., Citation1999a Citationb Citation2000a Citationb Citation2001). Nestor and others have shown through the use of lines of turkeys selected for higher body weights that these turkeys have reduced disease resistance and are more susceptible to infections with Pasteurella and Newcastle disease virus (Tsai et al., Citation1992; Nestor et al., Citation1999). These studies indicated that the acquired immune response was in large part unaffected by selection for large-bodied turkeys, although some differences were noted (Sacco et al., Citation1994; Li et al., Citation2001). Li et al. (Citation2001) reported that the phagocytic capabilities of peripheral blood phagocytes were reduced in birds selected for increased body weight, indicating some effects of selection for larger birds on the innate immune response.
Previous work in our laboratory found that heterophil functions, a major contributor to host innate immune responses, in 1-day-old to 14-day-old commercial turkeys and layer chickens was inefficient as compared with those in older birds (Lowry et al., Citation1997; Genovese et al., Citation1998 Citation2000; Wells et al., Citation1998). The inefficiency in function continued until approximately 21 days of age and was correlated with an increase in susceptibility to infection with pathogens such as Salmonella (Lowry et al., Citation1997; Genovese et al., Citation1998 Citation2000). Recently, our laboratory has found that different lines of parental broiler chickens exhibit differences in heterophil functions (Swaggerty et al., Citation2003). Increased production performance in these birds is associated with decreased innate immune functions, with the highest performing lines of birds possessing reduced innate immune functions.
Results of studies in other species have found similar results when comparing animals bred for enhanced performance characteristics with those that have not been subjected to such selection. In swine, similar differences in immune parameters have been found between different breeds of pigs (Duchet-Suchaux et al., Citation1991; Halbur et al., Citation1998; Reiner et al., Citation2002a Citationb; Clapperton et al., Citation2005). Although a definitive correlation between selection for increased performance parameters and decreased innate immune functions has not yet been established, it would appear that breeds with faster growth and larger bodies possess immune system characteristics that are different from those of smaller, slower growing pigs such as the Chinese Meishan (Duchet-Suchaux et al., Citation1991; Halbur et al., Citation1998; Reiner et al., Citation2002a Citationb; Clapperton et al., Citation2005).
Herein, we describe investigations of the innate immune response of large, heavy-bodied commercial turkey lines as compared with the response of smaller-bodied, wild-type Rio Grande turkeys to determine whether selection in modern turkeys for performance traits has affected the innate response as compared with wild-type turkeys that have not undergone such selection. Specifically, heterophil functions of large-bodied commercial lines of turkeys are compared with heterophil functions in small-bodied wild-type Rio Grande turkeys.
Materials and Methods
Turkeys
Turkeys from line A were obtained from a commercial turkey producer either as eggs or as 1-day-old poults. Eggs were incubated and hatched at our facilities. Day-of-hatch Rio Grande turkeys were obtained from Privett Hatchery (Portales, New Mexico, USA). Fertilized eggs were set in incubators (G.Q.F. Manufacturing Company, Savannah, Georgia, USA) and maintained at a dry bulb temperature of 37.5°C and a wet bulb temperature of 29.5°C. Eggs were candled at 11 days of incubation and non-fertile and non-viable eggs were disposed of. Eggs were transferred to a hatcher (Nature Form Inc., Jacksonville, Florida, USA) and maintained at a dry bulb temperature of 36.9°C and a wet bulb temperature of 32.2°C until the day of hatch. All turkeys were housed in climate-controlled rooms separately by line in floor pens with wood-shaving bedding and ad libitum access to feed and water. Additional warmth was provided by portable heat lamps. The feed met or exceeded standards established by the National Research Council (Citation1994). Experiments were conducted beginning on day 4 post-hatch to allow turkeys to acclimate to our facilities and to recover from the stress of transport.
Heterophil isolation
Heterophils were isolated from the peripheral blood of turkeys (100 turkeys per line/day) on days 4, 7, and 14 post-hatch for each of the experiments for two to three repetitions of each day using established isolation techniques (He et al., Citation2003). Briefly, ethylenediamine tetraacetic acid (EDTA)-anti-coagulated blood was mixed with 1% methylcellulose (25 cp; Sigma Chemical Co., St Louis, Missouri, USA) as a 1.5:1 ratio and was centrifuged at 25×g for 15 min. The serum and buffy coat layers were retained and suspended in Hank's balanced salt solution (HBSS 1:1) without calcium and magnesium. This suspension was layered over a discontinuous Ficoll–Hypaque gradient (specific gravity 1.077 over specific gravity 1.119). The gradient was then centrifuged at 250×g for 60 min. After centrifugation, the 1.077/1.119 interface and 1.119 band containing the heterophils were collected and washed twice in RPMI 1640 (Sigma) and resuspended in fresh RPMI 1640. The purity of the heterophil suspensions was assessed by microscopic examination of Hematology Three-Step Stain (CCRA Lab, Swedesboro, New Jersey, USA) stained cytospin (Shandon Scientific, Pittsburg, Pennsylvania, USA) smears. Cell viability counts were >95% as determined by trypan blue exclusion. Heterophil isolations obtained by this method were >95% pure. The cell concentration was adjusted to 8×106 cells/ml and stored on ice until used.
Salmonella enteritidis
A poultry isolate of S. enteritidis #97-11771 was obtained from the National Veterinary Services Laboratory (Ames, Iowa, USA). S. enteritidis was cultured in tryptic soy broth overnight at 41°C. The bacteria were pelleted (7700×g for 10 min) and washed with cold phosphate-buffered saline (PBS), centrifuged at 7700×g for 10 min, supernatant discarded, and the pellet re-suspended in 1 ml cold PBS and diluted to 1×109 colony-forming units (cfu)/ml in PBS using a Spectronic 20D spectrophotometer (Milton Roy Co., Golden, Colorado, USA) with a 625 nm reference wavelength. S. enteritidis was prepared fresh daily and kept on ice until used.
Opsonized Salmonella enteritidis
Opsonized Salmonella enteritidis (OPSE) was prepared as previously described (Ferro et al., Citation2005). Briefly, S. enteritidis (1×109 cfu/ml) were suspended in normal chicken serum (v:v [4:1]) and incubated for 60 min at 39°C on a rotary shaker. OPSE was stored at 4°C until used.
Heterophil degranulation
Degranulation was detected by quantifying the amount of β-glucuronidase activity in the culture medium following stimulation of heterophils (8×106 cells/ml) with or without OPSE for 60 min at 39°C on a rotary shaker in a 5% CO2 incubator. The reaction was stopped by transferring the tubes containing the cells to an ice bath for 5 min. Cells were centrifuged at 250×g for 10 min at 4°C. The supernatants were removed and used in the assay. Each supernatant sample (25 µl) was added to eight wells in a non-treated black CoStar flat-bottom enzyme-linked immunosorbent assay plate and incubated with 50 µl freshly prepared substrate (10 mM 4-methylumbelliferone-β-glucuronidide, 0.1% Triton X-100 in 0.1 M sodium acetate buffer) for 4 h at 41°C. The reaction was stopped by adding 200 µl stop solution (0.05 M glycine and 5 mM EDTA, pH 10.4). Liberated 4-methylumbelliferone was measured using a GENios Plus fluorescence microplate reader (355/460 nm; Tecan US Inc., Research Triangle Park, North Carolina, USA).
Heterophil oxidative burst
The heterophil oxidative burst was measured by oxidation of DCFH-DA (2′,7′-dichlorofluorescin-diacetate) to fluorescent DCF as described by He et al. (Citation2003). Heterophils were stimulated with phorbol A-myristate-13-acetate (PMA) (2.0 µg/ml; Sigma) or RPMI 1640 media for 60 min prior to measurement. Cells were placed in 96-well plates and fluorescence was measured using a GENios Plus Fluorescence Microplate Reader (485/530 nm; Tecan US Inc.).
Heterophil phagocytosis assay
Phagocytosis of live S. enteritidis was observed as previously described with modifications (Genovese et al., Citation2000). Heterophils 4×106 cells/ml in RPMI and S. enteritidis 1×108 cfu/ml were added to the heterophils (100 S. enteritidis:1 heterophil), and were incubated for 60 min at 39°C on a rocker. The samples were washed with an equal volume of RPMI and centrifuged at 190×g for 10 min and the supernatant was discarded. The heterophils were washed an additional three times with RPMI, and the pellet was resuspended in the original volume. Cytospin smears were prepared from each sample in a Shandon cytospin3 (Shandon Inc.), stained with Hematology Three stain system, and examined by light microscopy under oil immersion (100×). Results are reported as the percent heterophils containing S. enteritidis and the average number of S. enteritidis/heterophil.
Statistical analysis
Statistical analysis was performed on data using SigmaStat® statistical software (Jandel Scientific, San Rafael, California, USA). Differences between the two experimental groups were determined using Students t test.
Results
The results of turkey heterophil degranulation and oxidative burst are presented in Figures . Data are presented as separate experimental units due to the variability in activities measured during different experiments on different days. Although presented separately, the experiments consistently show that during days 4 and 7 post-hatch the observed heterophil degranulation activity, as measured by β-glucuronidase activity, and oxidative burst activity in Rio Grande turkey heterophils, expressed as relative fluorescence units, was significantly greater than the levels observed in heterophils from Line A birds (Figures ).
Figure 1. Day 4 post-hatch heterophil degranulation. Results are presented as three separate experiments. Data represent average µM per experiment±standard deviation. Heterophils were isolated from pooled peripheral blood from 100 turkeys from each line in each experiment. Heterophils were stimulated with OPSE for 1 h at 39°C. Data with different letters indicate a significant difference between groups (P < 0.001).
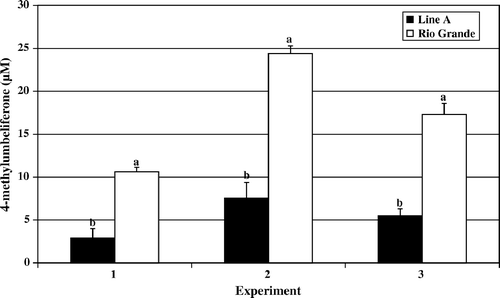
Figure 2. Day 4 post-hatch heterophil oxidative burst. Results are presented as three separate experiments. Data represent average relative fluorescence units (RFU) per experiment±standard deviation. Heterophils were isolated from pooled peripheral blood from 100 turkeys from each line in each experiment. Heterophils were stimulated with 2 µg/ml PMA for 1 h at 39°C. Data with different letters indicate a significant difference between groups (P < 0.001).
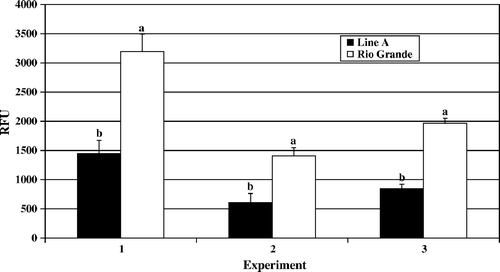
Figure 3. Day 7 post-hatch heterophil degranulation. Results are presented as three separate experiments. Data represent average µM per experiment±standard deviation. Heterophils were isolated from pooled peripheral blood from 100 turkeys from each line in each experiment. Heterophils were stimulated with OPSE for 1 h at 39°C. Data with different letters indicate a significant difference between groups (P < 0.001).
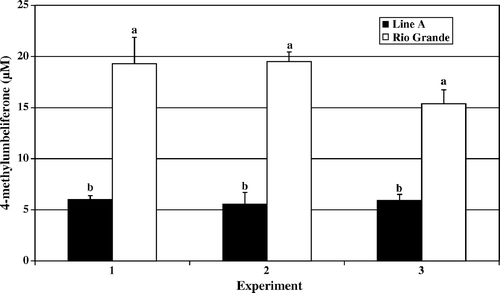
Figure 4. Day 7 post-hatch heterophil oxidative burst. Results are presented as two separate experiments. Data represent average relative fluorescence units (RFU) per experiment±standard deviation. Heterophils were isolated from pooled peripheral blood from 100 turkeys from each line in each experiment. Heterophils were stimulated with 2 µg/ml PMA for 1 h at 39°C. Data with different letters indicate a significant difference between groups (P < 0.01).
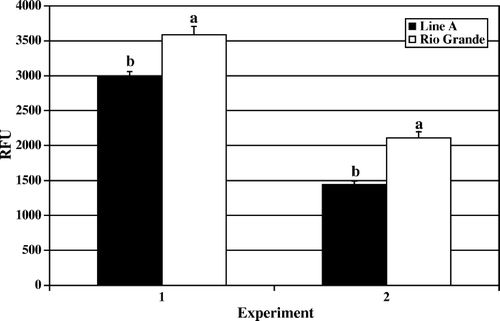
Figure 5. Day 14 post-hatch heterophil oxidative burst. Results are presented as two separate experiments. Data represent average relative fluorescence units (RFU) per experiment±standard deviation. Heterophils were isolated from pooled peripheral blood from 100 turkeys from each line in each experiment. Heterophils were stimulated with 2 µg/ml (PMA) for 1 h at 39°C. No significant differences were observed (P > 0.05).
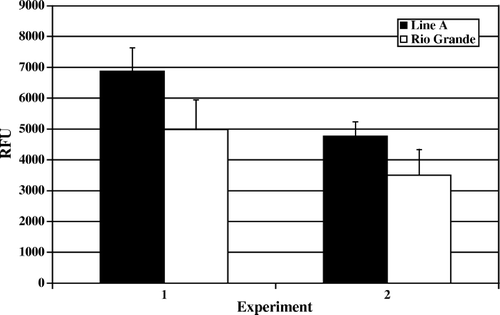
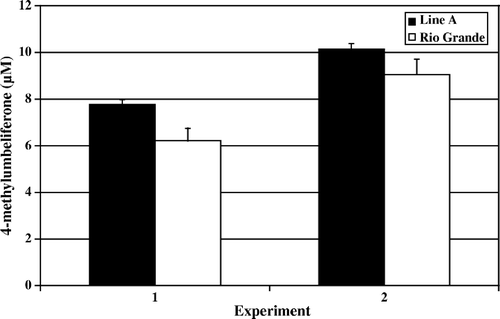
Figure 6. Day 14 post-hatch heterophil degranulation. Results are presented as two separate experiments. Data represent average µM per experiment±standard deviation. Heterophils were isolated from pooled peripheral blood from 100 turkeys from each line in each experiment. Heterophils were stimulated with OPSE for 1 h at 39°C. No significant differences were observed (P > 0.05).
Degranulation assay
In the degranulation assay, heterophils from Line A and Rio Grande turkeys were stimulated with OPSE for 1 h at 39°C. Data presented are the OPSE-stimulated levels of β-glucuronidase activity minus the control background level of activity for each line. On days 4 and 7 post-hatch, Rio Grande turkey heterophils responded with significantly (P<0.001) higher β-glucuronidase activity than did heterophils from Line A ( and ). By day 14, Rio Grande and Line A heterophils had similar levels of β-glucuronidase activity with no significant differences between the two lines of birds observed (; P>0.05).
Oxidative burst assay
The results of the oxidative burst assay, measuring the production of reactive oxygen species, followed a similar pattern of activity to that observed in the degranulation assays. In and , the data for day 4 and day 7 post-hatch are presented, respectively. Rio Grande turkey heterophils responded with a significantly elevated oxidative burst upon PMA stimulation compared with the levels recorded in Line A heterophils (day 4, P<0.001; day 7, P<0.01). Again, by day 14 post-hatch the observed levels of oxidative burst activity were increasing in Line A heterophils and remaining at a similar level to days 4 and 7 in Rio Grande heterophils ().
Phagocytosis assay
Heterophils from both lines of turkeys were isolated on days 4, 7, and 14 post-hatch. On all days assayed, there was no difference (P > 0.05) in the percentage of heterophils phagocytizing S. enteritidis (100% Line A and Rio Grande heterophils for all days) nor was there a difference in the average number of S. enteritidis phagocytized by heterophils (range, 10.1 to 10.45 S. enteritidis/heterophil) from both Line A and Rio Grande turkeys.
Discussion
Here we compare two different mechanisms heterophils employ to kill bacteria in two different lines of turkeys. Phagocytosis of S. enteritidis by heterophils from both lines was also evaluated. Commercial Line A is one of the most common lines of turkeys used in production today, while the wild-type Rio Grande turkeys have been raised to preserve their wild heritage and have not been selected for growth performance or feed efficiency and are backyard flock birds for fanciers. Wild-type Rio Grande turkeys, along with other older breeds such as Black Spanish, Blue Slate, and Narragansett turkeys, are single-breasted birds that are capable of breeding naturally. Commercial lines of turkeys are considered to be double-breasted birds that grow to a large size in a relatively short period of time and have highly improved feed efficiencies. The males of commercial lines of turkeys are bred to reach approximately 33 lbs body weight by 18 weeks of age (Leeson & Summers, Citation2005). Commercial turkeys are unable to breed naturally and are artificially inseminated.
To the authors’ knowledge, no studies on the immune system and its functions in wild-type turkeys have been reported. The Rio Grande turkeys included in these studies are from a large primary breeder that supplies rare breeds of chickens and turkeys to backyard bird fanciers and those wishing to raise unique breeds of turkeys for both consumption and as ornamentals. Likewise, no selection for growth or other production parameters has been conducted with the Rio Grande turkeys supplied by the hatchery. Although these birds are not true “wild” birds taken from their natural surroundings, their origin is from wild stock and the hatchery has attempted to preserve their wild heritage, allowing the birds to reproduce naturally.
Phagocytosis of pathogenic bacteria is the first step in the eventual elimination of invaders of the host organism. The oxidative burst produces reactive oxygen species which kill both intra-cellular and extra-cellular bacteria. Heterophil degranulation releases bactericidal products from granules inside the heterophil, often into the phago-lysosome. The results of these experiments show that heterophils from wild-type Rio Grande turkeys possess more effective killing mechanisms during the first 7 to 14 days post-hatch than do the commercial lines of turkeys. There were no differences in the phagocytosis of bacteria between the two lines. The results from commercial Line A parallel results reported earlier observing a gradual increase in heterophil functions in commercial turkeys until day 21 post-hatch (Lowry et al., Citation1997; Genovese et al., Citation1998). This previous research showed that heterophil functions of neonatal turkeys were deficient early in life and that these deficiencies in function continued until day 21 post-hatch (Lowry et al., Citation1997; Genovese et al., Citation1998 Citation2000; Wells et al., Citation1998). The reduced levels of heterophil functions observed in commercial turkey lines may suggest an inherent susceptibility to bacterial infection in young turkeys from commercial lines and these deficiencies may be a contributing factor to health problems later in life.
In the present studies, the differences in heterophil function between Line A and Rio Grande heterophil function is evident for the first 7 days of life. These data are comparable with results observed in comparison studies of heterophil functions in lines of broiler chickens conducted by our laboratory (Swaggerty et al. Citation2003). Swaggerty et al. (Citation2003 Citation2004), described similar differences between commercial broiler lines in their innate immune capabilities, and have found that the functional deficiencies observed may be related to deficiencies in proinflammatory cytokine profiles in heterophils from these broiler lines.
These results would suggest that Rio Grande turkeys would be less susceptible to infection early in life as compared with Line A birds. As the bird grows and matures, heterophil functions begin to decrease, presumably as the acquired immune system of the bird has developed and matured. In Line A birds, the heterophil functions increase over time post-hatch, but are reduced early on. These observations may suggest that selection for larger, heavier, and faster growing birds may have had a negative effect on the innate immune functions in these animals. In addition, phagocytosis of S. enteritidis did not differ between the Line A and Rio Grande heterophils over the experimental period, suggesting that the susceptibility to disease reported by other researchers in large bodied turkeys is not due to a defect in the phagocytosis of bacteria, but lies in the mechanisms used either to kill the bacteria or in the signaling events that lead to the bactericidal activities of the heterophil. Research to describe the cytokine profiles of heterophils from wild-type turkeys compared with heterophils from commercial turkey lines as well as other heterophil functions will be conducted to better understand the effects of selection on the innate and acquired immune response. We will also investigate the differences between commercial lines and wild-type lines related to susceptibility to infections with pathogens such as Salmonella.
Comparison studies of broiler strains from 1957 and 2001 showed that the 1957 strains had one-half of the mortality seen in modern strains, and much of this mortality was associated with leg problems later in production life (Cheema et al., Citation2003). In studies of broilers and in mammalian species, associations between selection for growth performance and increased feed efficiency and the subsequent effects on the innate immune system of these animals have been investigated (Duchet-Suchaux et al., Citation1991; Quereshi & Havenstein, Citation1994; Halbur et al., Citation1998; Reiner et al., Citation2002a Citationb; Swaggerty et al., Citation2003 Citation2004; Clapperton et al., Citation2005). In turkeys, it has been shown that selection for these same characteristics leads to an increase in susceptibility to disease (Sacco et al., Citation1994, Li et al., Citation1999a Citationb Citation2000a Citationb Citation2001; Nester et al., 1999). A direct genetic component for these deficiencies in the innate immune system remains to be identified.
Descriptions of defects in the intracellular signaling cascades related to pattern recognition receptors can cause increased susceptibility to bacterial diseases in mice and humans (Lee et al., Citation2005). The possibility exists that selection of modern poultry for growth and feed efficiencies, while providing excellent poultry products, has selected for changes in the genes associated with innate immune signaling cascades. These genetic changes then lead to dysfunction of the signaling events and subsequent cell functions, resulting in increased susceptibility to disease and subsequent decreases in livability. Further characterization of the innate immune functions of commercial turkey lines and wild-type turkeys, including signaling pathways and genetic components, are currently underway.
In conclusion, the present studies have described differences in aspects of the innate immune responses of commercial turkeys and wild-type Rio Grande turkeys. The differences in heterophil degranulation and production of reactive oxygen species and the lack of differences in the phagocytosis of S. enteritidis described are directly related to the host's ability to defend itself against infections with pathogens. The deficiencies noted in Commercial Line A birds would suggest an increased susceptibility to infections with pathogens and may, with further investigation, suggest a scenario for problems with morbidity and livability in these birds. It may be possible to define genetic differences in the innate system between commercial lines of birds and their wild-type predecessors, and to then select for birds that have improved innate system function early in life and that are still able to meet the performance standards of modern commercial turkey production.
References
- Akira , S. 2001 . Toll-like receptors and innate immunity . Advances in Immunology , 78 : 1 – 56 .
- Cheema , M.A. , Qureshi , M.A. and Havenstein , G.B. 2003 . A comparison of the immune response of a 2001 commercial broiler with a 1957 randombred broiler strain when fed representative 1957 and 2001 broiler diets . Poultry Science , 82 : 1519 – 1529 .
- Clapperton , M. , Bishop , S.C. and Glass , E.J. 2005 . Innate immune traits differ between Meishan and Large White pigs . Veterinary Immunology and Immunopathology , 104 : 131 – 144 .
- Duchet-Suchaux , M.F. , Bertin , A.M. and Menanteau , P.S. 1991 . Susceptibility of Chinese Meishan and European large white pigs to enterotoxigenic Escherichia coli strains bearing colonization factors K88, 987P, K99 or F41 . American Journal of Veterinary Research , 52 : 40 – 44 .
- Fearon , D.T. and Locksley , R.M. 1996 . The instructive role of innate immunity in the acquired immune response . Science , 272 : 50 – 54 .
- Ferro , P.J. , Swaggerty , C.L. , He , H. , Rothwell , L. , Kaiser , P. and Kogut , M.H. 2005 . Recombinant chicken IL-6 does not activate heterophils isolated from day-old chickens in vitro . Developmental and Comparative Immunology , 29 : 375 – 383 .
- Genovese , K.J. , Moyes , R.B. , Genovese , L.L. , Lowry , V.K. and Kogut , M.H. 1998 . Resistance to Salmonella enteritidis organ invasion in day-old turkeys and chickens by transformed t cell line produced lymphokines . Avian Diseases , 42 : 545 – 553 .
- Genovese , L.L. , Lowry , V.K. , Genovese , K.J. and Kogut , M.H. 2000 . Longevity of augmented phagocytic activity of heterophils in neonatal chickens following administration of Salmonella enteritidis-immune lymphokines to chickens . Avian Pathology , 29 : 117 – 122 .
- Halbur , P.G. , Rothschild , M.F. , Thacker , B.J. , Meng , X.J. , Paul , P.S. and Bruna , J.D. 1998 . Differences in susceptibility of Duroc, Hampshire, and Meishan pigs to infection with a high virulence strain (VR2385) of porcine reproductive and respiratory syndrome virus (PRRSV) . Journal of Animal Breeding and Genetics , 115 : 181 – 189 .
- He , H. , Farnell , M.B. and Kogut , M.H. 2003 . Inflammatory agonist stimulation and signal pathway of oxidative burst in neonatal chicken heterophils . Comparative Biochemistry and Physiology A , 135 : 177 – 184 .
- Imler , J.-L. and Hoffman , J.A. 2001 . Toll receptors in innate immunity . Trends in Cell Biology , 11 : 304 – 311 .
- Janeway , C.A. Jr. and Medzhitov , R. 2002 . Innate immune recognition . Annual Review of Immunology , 20 : 197 – 216 .
- Kopp , E. and Medzhitov , R. 2003 . Recognition of microbial infection by Toll-like receptors . Current Opinions in Immunology , 15 : 396 – 401 .
- Lee , J.Y. , Lowell , C.A. , Lemay , D.G. , Youn , H.S. , Rhee , S.H. , Sohn , K.H. , Jang , B. , Ye , J. , Chung , J.H. and Hwang , D.H. 2005 . The regulation of the expression of inducible nitric oxide synthase by Src-family tyrosine kinases mediated through MyD88-independent signaling pathways of Toll-like receptor 4 . Biochemical Pharmacology , 70 : 1231 – 1240 .
- Leeson , S. and Summers , J.D. 2005 . Commercial Poultry Nutrition , 3rd edn , Guelph , Ontario : University Books .
- Li , Z. , Nestor , K.E. , Saif , Y.M. , Bacon , W.L. and Anderson , J.W. 1999a . Effect of selection for increased body weight on mitogenic response in turkeys . Poultry Science , 78 : 1532 – 1535 .
- Li , Z. , Nestor , K.E. , Saif , Y.M. , Fan , Z. , Luhtala , M. and Vainio , O. 1999b . Cross-reactive anti-chicken CD4 and CD8 monoclonal antibodies suggest polymorphism of the turkey CD8α molecule . Poultry Science , 78 : 1526 – 1531 .
- Li , Z. , Nestor , K.E. , Saif , Y.M. and Anderson , J.W. 2000a . Antibody responses to sheep red blood cell and Brucella abortis antigens in a turkey line selected for increased body weight and its randombred control . Poultry Science , 79 : 804 – 809 .
- Li , Z. , Nestor , K.E. , Saif , Y.M. , Fan , Z. and Luhtala , M. 2000b . Flow cytometric analysis of T lymphocyte subpopulations in large-bodied turkeys and a randombred control population . Poultry Science , 79 : 219 – 223 .
- Li , Z. , Nestor , K.E. , Saif , Y.M. , Anderson , J.W. and Patterson , R.A. 2001 . Effect of selection for increased body weight in turkeys on lymphoid organ weights, phagocytosis, and antibody responses to fowl cholera and Newcastle disease-inactivated vaccines . Poultry Science , 80 : 689 – 694 .
- Lowry , V.K. , Genovese , K.J. , Bowden , L.L. and Kogut , M.H. 1997 . Ontogeny of the phagocytic and bactericidal activities of turkey heterophils and their potentiation by Salmonella enteritidis-immune lymphokines . FEMS Immunology and Medical Microbiology , 19 : 95 – 100 .
- Medzhitov , R. and Janeway , C.A. Jr. 1997a . Innate immunity: impact on the adaptive immune response . Current Opinion in Immunology , 9 : 4 – 9 .
- Medzhitov , R. and Janeway , C.A. Jr. 1997b . Innate immunity: the virtues of a nonclonal system of recognition . Cell , 91 : 295 – 298 .
- National Research Council . (1994) . Nutrient Requirements for Poultry 9th rev edn (pp. 19 – 34 ). Washington , DC : National Academy Press .
- Nestor , K.E. , Saif , Y.M. , Anderson , J.W. , Patterson , R.A. and Li , Z. 1999 . Variation in resistance to Pasteurella multocida among turkey lines . Poultry Science , 78 : 1377 – 1379 .
- Quereshi , M.A. and Havenstein , G.B. 1994 . A comparison of the immune performance of a 1991 commercial broiler with a 1957 random-bred strain when fed “typical” 1957 and 1991 broiler diets . Poultry Science , 73 : 1805 – 1812 .
- Reiner , G. , Melchinger , E. , Kramarova , M. , Pfaff , E. , Buttner , M. , Saalmuller , A. and Geldermann , H. 2002a . Detection of quantitative trait loci for resistance/susceptibility to pseudorabies virus in swine . Journal General Virology , 83 : 167 – 172 .
- Reiner , G. , Eckert , J. , Peischl , T. , Bochert , S. , Jakel , T. , Mackenstedt , U. , Joachim , A. , Daugschies , A. and Geldermann , H. 2002b . Variation in clinical and parasitological traits in Pietrain and Meishan pigs infected with Sarcocystis meischeriana . Veterinary Parasitology , 106 : 99 – 113 .
- Romagnani , S. 1992 . Induction of Th1 and Th2 responses: a key role for the “natural” immune response? . Immunology Today , 13 : 379 – 381 .
- Sacco , R.E. , Nestor , K.E. , Saif , Y.M. , Tsai , H.J. and Patterson , R.A. 1994 . Effect of genetic selection for increased body weight and sex of poults on antibody response of turkeys to Newcastle disease virus and Pasteurella multocida vaccines . Avian Diseases , 38 : 33 – 36 .
- Swaggerty , C.L. , Pevzner , I.Y. , Lowry , V.K. , Farnell , M.B. and Kogut , M.H. 2003 . Functional comparison of heterophils isolated from commercial broiler chickens . Avian Pathology , 32 : 95 – 102 .
- Swaggerty , C.L. , Kogut , M.H. , Ferro , P.J. , Rothwell , L. , Pevzner , I.Y. and Kaiser , P. 2004 . Differential cytokine mRNA expression in heterophils isolated from Salmonella-resistant and -susceptible chickens . Immunology , 113 : 139 – 148 .
- Tsai , H.J. , Saif , Y.M. , Nestor , K.E. , Emmerson , D.A. and Patterson , R.A. 1992 . Genetic variation in resistance of turkeys to experimental infection with Newcastle disease virus . Avian Diseases , 36 : 561 – 565 .
- Wells , L.L. , Lowry , V.K. , DeLoach , J.R. and Kogut , M.H. 1998 . Age-dependent phagocytosis and bactericidal activities of the chicken heterophil . Developmental and Comparative Immunology , 22 : 103 – 109 .