Abstract
Brea, M., Artabe, A. & Spalletti, L.A., December, 2008. Ecological reconstruction of a mixed Middle Triassic forest from Argentina. Alcheringa 32, 365–393. ISSN 0311-5518.
The palaeoecology of an in situ Middle Triassic forest known as the Darwin Forest, from the Paramillo Formation of Argentina, is described based on palaeobotanical, sedimentological and spatial analyses. The palaeoforest grew on an andisol soil that developed on volcaniclastic floodplain deposits. The volcanic detritus and the rhythmic amalgamation of upper flow-regime tractional deposits overlying the andisol indicate that the forest was buried rapidly by a subaerial, cool and wet pyroclastic base surge flow. The plant community was reconstructed by quantitative mapping of the fossilized stumps integrated with taxonomic and sedimentological information. The Darwin Forest had a tree density of 427–759 per ha, with an upper stratum (20–26 m) of corystosperms and a second stratum (16–20 m) of conifers. Estimated biomass is equivalent to modern dry monsoonal forest. The understorey was composed of ferns (Cladophlebis spp.). Quantitative growth-ring analysis of Araucarioxylon protoaraucana suggests that these trees were evergreen and, together with vegetation structure and sedimentary data, indicate that the forest developed under dry, subtropical, strongly seasonal conditions.

A SERIES of narrow rifts developed in west-central Argentina as a result of Triassic continental extension (Spalletti Citation2001; ). Each of these rift basins was filled with a complex mix of predominantly alluvial, fluvial, deltaic and lacustrine sediments. The volcaniclastic and sedimentary sequences of most of these basins contain well-preserved fossil plants at multiple stratigraphic levels. These assemblages provide a rich reservoir of information for reconstructing the palaeoecology and palaeophytogeography of South American Triassic vegetation and for developing a comprehensive biostratigraphic zonation of the system.
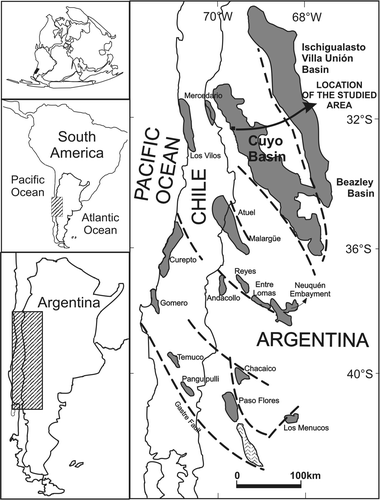
Fig. 1 Map of central Argentina and Chile showing the main Triassic basins (after Morel et al. Citation2003).
Most investigations of Argentinean Triassic floras over the past two decades have focused on the taxonomy and stratigraphic distribution of fossil plants (Stipanicic Citation2001, Zamuner et al. Citation2001, Stipanicic & Marsicano Citation2002). However, some recent studies (Spalletti et al. Citation1999, Citation2003, Artabe et al. Citation2001, Citation2003, Morel et al. Citation2003) have provided an improved understanding of floristic evolution in response to changes in Triassic depositional systems, tectonism and climate. A biostratigraphic zonation based on plant megafossils (Spalletti et al. Citation1999) has facilitated regional correlations among different Triassic basins in Argentina. Three main events punctuated the evolution of Triassic floras in southwestern Gondwana (Morel et al. Citation2003, Spalletti et al. Citation2003). Artabe et al. (Citation2003) defined several biogeographic units (realm, areas and province) within the region based on these Triassic floristic changes.
The Cuyo Basin is a 30 000 km2 elongate basin located in western Argentina between 32° and 36° S (). The basin's Triassic latitude was approximately the same as its present position (sensu Prezzi et al. Citation2001). It is the most important hydrocarbon-producing Triassic basin of western Gondwana (Moratello Citation1993, Chebli et al. Citation2001). Triassic sediments exceed 3500 m in thickness, and were deposited by multiple episodes of continental sedimentation now represented by several formations (). This study focuses on an in situ mixed forest preserved in the Paramillo Formation, Agua de la Zorra area, northwestern sector of the Cuyo Basin (, 4). This unit is composed of a 140 m thick sequence of clast-supported conglomerates, pebbly sandstones, tuffaceous sandstones and mudstones that have been described by Harrington (Citation1971), Strelkov & Álvarez (Citation1984), Massabie (Citation1985), Massabie et al. (Citation1985), Kokogian & Mancilla (Citation1989), Linares & González (Citation1990), Ramos & Kay (Citation1991), Kokogián et al. (Citation1993) and Ramos (Citation1993). These authors all interpreted the Paramillo Formation to have been deposited in high sinuosity fluvial systems. Darwin (1846), Conwentz (1885), Stappenbeck (Citation1910), Kurtz (Citation1921), Du Toit (Citation1927), Groeber (Citation1939), Windhausen (Citation1941), Harrington (Citation1971) and Stipanicic et al. (Citation1996) all carried out palaeobotanical studies on the unit. More recently, one of the authors (M.B.) described the palaeobotanical content of this unit in her doctoral thesis (1995) and illustrated two palaeocommunities in the Paramillo Formation at Agua de la Zorra area. The first one, found at locality A (Brea & Artabe Citation1999) is dominated by Apocalamitaceae. This autochthonous taphocoenosis incorporates stems and nodal diaphragms assigned to Neocalamites carrerei (Zeiller) Halle, Citation1908 and aff. Nododendron suberosum Artabe & Zamuner, Citation1991, found in close association with reproductive structures of Neocalamostachys arrondoi Brea & Artabe, Citation1999 (FL II: see ). The species probably represent dispersed organs of a single biological taxon. These monotypic horsetail palaeocommunities probably formed water-dependent, bamboo-like thickets in fluvial flood plains.
Fig. 2 Stratigraphic chart showing the position of the Paramillo Formation (ages and biozones from Morel et al. Citation2003).
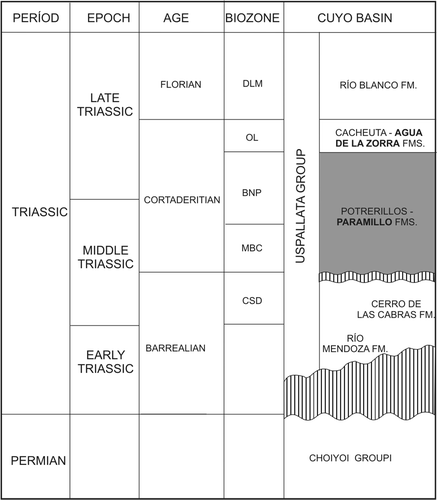
Fig. 3 Location map showing the Agua de la Zorra fossiliferous region, Mendoza province, Argentina.
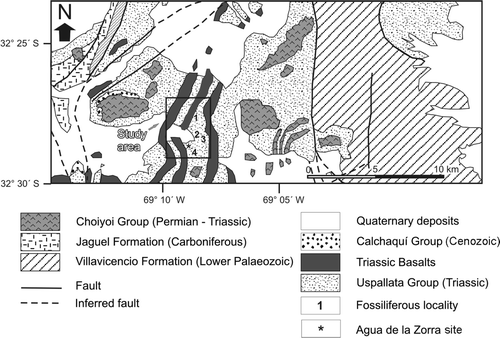
Fig. 4 Map of the study area showing the location of stumps at locality A (A), locality B (B), locality C (C) and locality D (D). Conifer and corystosperm stumps are indicated by point and asterisk symbols respectively.
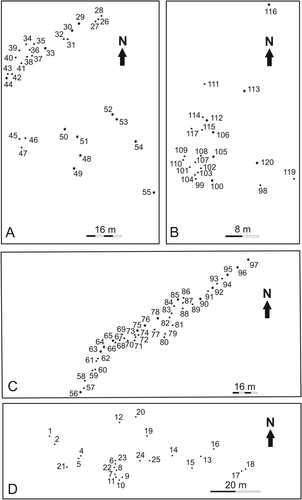
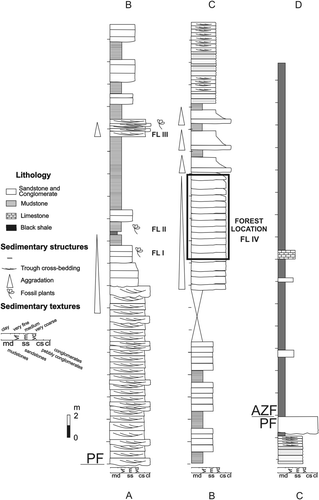
Fig. 5 Lithostratigraphic section of Paramillo and Agua de la Zorra formations at locality A, showing the main lithofacies and the position of the fossil forest level. PF = Paramillo Formation; AZF = Agua de la Zorra Formation; FL I–IV = fossiliferous levels.
The second palaeocommunity is of more significance: the Darwin Forest, first described by Darwin (1839a,b; see Ottone Citation2005) during his historic journey around the world. Initially, this fossil forest was dated as Cenozoic. However, new data reassign it to the oldest biozone of the Cortaderitian Stage (Yabeiella mareyesiaca–Scytophyllum bonettiae–Protocircoporoxylon cortaderitaensis Assemblage Biozone), following the phytozonation of Spalletti et al. (Citation1999), which corresponds to the late Middle Triassic. Other in situ petrified forests found in central western Argentina have been assigned to the late Middle Triassic (Barreal-Calingasta Basin, Cortaderita Formation), early Late Triassic (Ischigualasto-Villa Unión Basin, Ischigualasto Formation) and Late Triassic (Cuyo Basin, Río Blanco Formation): see Spalletti et al. (Citation1999, Citation2003), Artabe et al. (Citation2001, Citation2007) and Morel et al. (Citation2003). Another Middle Triassic in situ forest has been recorded at very high latitudes, in the central Transantarctic Mountains (Cúneo et al. Citation2003).
This study focuses on the in situ, mixed-fossil Darwin Forest in the Agua de la Zorra area. The Darwin Forest is reconstructed based on quantitative information (mean separation of trees, density, basal area per stump, class diameter, dominance, estimated heights and estimated biomass). Spatial data are also integrated with taxonomic-phenological information to infer the environmental conditions under which the community developed. The structural data were used to compare the vegetation with other Triassic fossil forests. Growth-ring analyses were used to determine both tree habit and palaeoclimatic conditions. Mean ring width, thinnest ring and widest ring were measured as records of annual growth conditions (Francis Citation1986). Annual Sensitivity (AS) and Mean Sensitivity (MS) were calculated to determine the degree of variability between years (Fritts Citation1976). In addition, the proportion of latewood to earlywood, the presence or absence of false rings and the cumulative sum of deviations from mean radial diameter (CSDM curve) were calculated for each growth-ring increment (Falcon-Lang Citation2000a) to assess intraseasonal conditions and leaf longevity patterns.
Material and methods
Three field studies were carried out in 1993–1994 during which four exposures of the Paramillo Formation Darwin Forest were found: localities A, B, C and D. Localities A–C (see ) are situated to the northeast of Agua de la Zorra site, whereas the locality D is located to the southeast of this site. The Paramillo Formation was thoroughly examined at the localities A and C, where the lithology was logged in three vertical sections (, 5–7). The plant fossils of each bed were recorded with respect to their quantity, taxonomic representation, distribution, orientation and state of preservation (Kidwell et al. Citation1986). These parameters allowed an estimation of the degree of transportation prior to burial (defining the assemblages as autochthonous, hypautochthonous, or allochthonous taphocoenoses). The fossil floras of beds FL I and FL III (at locality A) are represented by leaf impressions–compressions of a fern taxon (Brea Citation2000) and are considered to constitute forest understory assemblages (). The flora of FL II is a monotypic palaeocommunity dominated by Apocalmitaceae as mentioned above.
The stumps found in FL IV (at locality A), FL V (at localities B and C) and FL VI (at locality D) belong to the same in situ fossil forest that emerges at the four localities ( –). An imperfect carbonized process preserved the stumps (Poma et al. Citation2004). One hundred and twenty stumps occur in growth position over an area of 2 km2 (). The fossil forest was mapped at 1:100 scale using plane table and alidade (at locality D) and Brunton compass and tape measure (at localities A, B and C). Trunks with a diameter less than 8 cm were not considered because they may constitute parts of shrubs or branches of trees. According to Mosbrugger (Citation1990), a tree is a woody plant with a diameter of more than 7.5 cm at breast height. Wood samples were also collected for identification (Brea Citation1997, Artabe & Brea Citation2003).
Fig. 6 Lithostratigraphic section of the Paramillo and Agua de la Zorra formations at locality C, showing the main lithofacies and the position of the fossil forest level. PF = Paramillo Formation; AZF = Agua de la Zorra Formation; FL V = fossiliferous level.
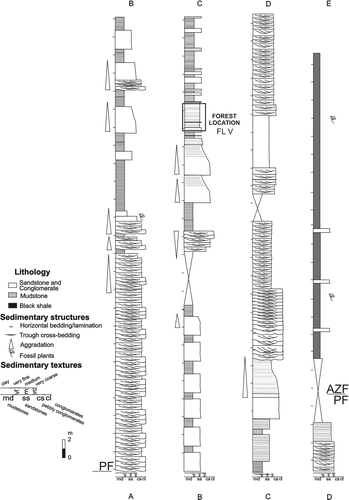
Fig. 7 Detailed log of the Paramillo Formation showing the position of the locality C fossil assemblage and main sedimentological features.

The forest structure is reinterpreted here based on palaeobotanical, sedimentological and spatial analyses. To analyse the two-dimensional distribution of the vegetation components, a spatial statistical package created to analyse crime incident location data was used. The CrimeStat II program (Levine Citation2002) provides a variety of tools to describe the overall spatial distribution, the dominant pattern of distribution and sub-regional patterns or ‘neighbourhood’ patterns within the overall distribution of studied objects. The density of the in situ permineralized stumps was calculated using the distance to its nearest neighbour for each stump and the mean distance between stumps. To identify spatial autocorrelation for the diameter and species variables, the Geary's C Statistic Index was used (In the CrimeStat II program, Levine Citation2002). Stem basal diameters were also measured for each stump and the total basal area was obtained by multiplying the number of trees per ha by the mean basal area per ha (Cottam & Curtis Citation1956). The biomass was estimated using Murray's formula (Murray Citation1927: in Mosbrugger et al. Citation1994) from stump circumference to obtain fresh weight (w f): W f = 7.08C 2.49, where C is the circumference; and dry weight (w d), where W d = 0.45W f (see Mosbrugger et al. Citation1994).
Biomass was also estimated from stump diameter (D) using the equations developed by Brown (Citation1997) for application to stems with D ≥ 5.6 cm. Two regression equations were used:
where Y T = weight, and D = diameter (cm). These equations are generally grouped according to Holdridge's life zones (Holdridge Citation1979). These values were compared with those obtained for the biomass of different life zones in extant forests (Brown & Lugo Citation1984, Murphy & Lugo Citation1986, Brown et al. Citation1991, Brandeis et al. Citation2005).
Predictive models, applied to extant conifers and angiosperms, enabled us to establish the relationship between stem diameter and height (Niklas Citation1994a). The correlation of these features can be used to reconstruct the height of fossil plants for which basal stem diameters are known or inferred (Niklas Citation1994b).
The critical buckling height (H crit), or the maximum height to which a vertical trunk can be elevated before it theoretically undergoes buckling, was estimated using Niklas' formula (see: Creber & Ash Citation2004, Artabe et al. Citation2007): H crit =C(E/ρ)1/3 D 2/3, where C is the constant of proportionality (= 0.792), E is Young's modulus (958.1 × 106 kg/m2), ρ is the bulk density of the material (tissues) used to construct the columnar stem (468.9 kg/m3), and D is the stem diameter. Finally, H crit = 95.75 D 2/3 (Creber & Ash, Citation2004). The trees never attain H crit, and for this reason, estimated tree heights were calculated on the basis of trunk diameter-to-height ratios observed in living trees (Niklas Citation1993, Citation1994a,b) using the formula: H est = 27.8D 0.430, where D corresponds to fossil stump diameters. The Safety Factor (SF) was obtained by dividing H crit by the estimated height (H est). Using SF, a more accurate estimated height (H est1) for the Darwin fossil forest trees was obtained.
The SF, which can be taken as the ratio of the maximum loading likely to be experienced during the operational (normal) loadings on a structure, is typically based on the statistical probability that certain types of loadings will occur (Niklas Citation1992).
The main purpose of growth-ring analyses is to relate ring growth patterns to palaeoclimate (Creber Citation1977, Creber & Chaloner Citation1984a). Following Fritts (Citation1976), Creber (Citation1977), Creber & Chaloner (Citation1984ab) and Schweingruber (Citation1988), growth rings were measured from thin-sections and polished blocks, then subjected to dendrological analysis. Growth rings were measured to the nearest 0.01 mm with a Bannister incremental measuring machine, capturing measurements on a microcomputer using the COMPUTA program (Stockes & Smiley Citation1968, Evans Citation1981). The COFECHA program was used to produce growth-ring series. Growth parameters determined for the gymnosperm fossil woods included the number of rings, mean ring width, maximum ring width, minimum ring width, the thinnest ring, the widest ring, the presence and number of false rings, MS and AS (Fritts Citation1976, Holmes Citation1985). The mean ring width is a useful indicator of growth, and wide rings indicate faster growth or a longer growing season (Fritts Citation1976). The most informative parameter was the variability in ring width (growth) from year to year. This was calculated in terms of AS, i.e. the difference in width between a pair of consecutive rings divided by their average width (Creber Citation1977).
The growth rings of the coniferous taxon, Araucarioxylon protoaraucana, were analysed quantitatively. Using the method described by Falcon-Lang (Citation2000a), the cumulative algebraic sum of each cell's deviation from the mean of the radial diameters was calculated for each growth-ring increment and plotted as a zero-trending curve (CSDM curve). For each ring increment, the percentage skew of the zenith of CSDM curves from the centre of the plot was calculated. Deciduous conifers give CSDM curves that are dominantly left-skewed or symmetrical, whereas evergreen conifers have dominantly right-skewed CSDM curves. On this basis, the pattern of foliage loss for Araucarioxylon protoaraucana was inferred. Because the percentage of latewood may be strongly influenced by leaf longevity in addition to reflecting the intensity of climate seasonality, two aspects of growth-ring anatomy were quantified using Falcon-Lang's technique (Citation2000b) to calculate the Ring Markedness Index (RMI). The percentage of diminution in a ring increment was determined using the percentage of diminution (x) = (b/a) 100; where a = the maximum cell diameter, and b = the maximum cell diameter – minimum cell diameter. The percentage of latewood in each growth-ring increment was calculated using the method of Creber & Chaloner (Citation1984a): percentage of latewood (y) = (d/c) 100; where d = the number of cells in each ring increment, and c = the number of cells since the CSDM curve turns to zero for the last time. The product of these two parameters was calculated to determine a Ring Markedness Index (RMI) [ = (x×y)/100].
The longest growth-ring series were analysed using the method based on the concept of ABIC (A Bayesian Information Criterion) minimization, which is a log-marginal likelihood method (Akaike Citation1980, Kumagai et al. Citation1995). This method is standardized to remove non-climatic variation. In the Past program (Hammer et al. Citation2007), the likelihood alogorithm and Akaike criterion are given for each growth-ring series.
The MS is a quantitative measure of the year-to-year variability in ring width (Fritts Citation1976), and provides the best indicator of variability in growth rate over the lifespan of the tree. Mean Sensitivity values can range from zero, i.e. no year-to-year variability, to an indeterminate maximum representing the greatest possible variability. High MS values (>0.3) represent sensitive growth and are considered diagnostic of stressful environmental conditions (Creber & Francis Citation1999), whereas complacent ring sequences (MS values <0.3) form under uniform, favourable conditions (Fritts Citation1976).
Photomicrographs were taken with a Leica DM5000 light microscope and digital camera. All specimens and microscopic slides are deposited in the Colección Paleobotánica, División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, under the acronym LPPB and pmLPPB.
Sedimentary features of the Darwin Forest deposits
The Paramillo Formation is a volcaniclastic unit composed of yellowish lithic sandstones, brownish and yellowish tuffaceous sandstones, dark grey and green shales and mudstones, and pink to reddish ash-fall tuffs. The overlying Agua de la Zorra Formation is dominated by bituminous shales and marls with subordinated intercalations of yellowish fine-grained sandstones and mudstones (, 6). Both units are intruded by several sills of olivine diabase dated as 235 ± 5 Ma (K/Ar whole age) by Ramos & Kay (Citation1991).
Brea (Citation1995) studied the upper part of the Paramillo Formation and the Agua de la Zorra Formation in detail and defined several lithofacies and facies associations. The Paramillo Formation consists of cross-bedded conglomerates, cross-bedded, plane-bedded and massive pebbly sandstones, cross-bedded, massive, plane bedded and ripple-laminated sandstones and intercalations of laminated mudstones and shales. Brea (Citation1995) interpreted these sediments to have been deposited in a high-sinuosity fluvial system in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. The Agua de la Zorra Formation is characterized by black shales and marly shales with thin and isolated intercalations of fine-grained sandstones, mudstones, and stromatolitic micrites. These deposits were accumulated in a lacustrine setting where suspension fall-out deposits and anoxic restricted hypolimnic conditions prevailed (Brea Citation1995).
The detailed measured section through the Darwin Forest deposits (made at locality C; ) shows that the volcaniclastic sediments are characterized by laterally continuous and alternating beds of massive or laminated very fine grained to fine-grained wackes intercalated with tabular units of massive and plane-bedded medium to very coarse-grained sandstones and granule conglomerates. The lithic sandstones are dominated by basaltic rock fragments accompanied by subrounded to angular, unaltered, inclusion-free andesine to labradorite plagioclase and subordinate quartz. The matrix is quite abundant in several sandstones and consists of a microcrystalline quartz-feldspar mixture and chlorite-smectite clay minerals, as indicated by XRD analysis. The mudstones and shales are composed of fine flakes of irregular interlayered chlorite-smectite clays, angular to subangular feldspars (probably plagioclase) and quartz, and submicroscopic opaque minerals.
The fine-grained sandstone beds, upon which the Darwin Forest developed, are formed of abundant ovoid to irregular nodular bodies up to 5 mm in diameter. These components are fragments of the original sediment fractured in situ by the contraction of expansive clay minerals. The nodule-rich levels host slickensides. Thin-section analysis revealed that contraction fractures are filled with recrystallized clay minerals forming a ‘chickenwire’ grid that is interpreted as a pedogenetic network (argillans). The development of these soils upon volcanic extrusive and/or pyroclastic material suggests that they could be classified as andisols: immature soils whose evolution was interrupted by renewed sediment aggradation.
The rhythmic amalgamation of upper flow-regime tractional deposits and the laterally persistent bed-geometry suggest episodic non-channelized sedimentation with rapid aggradation. The volcanic nature of detrital components and the presence of amalgamated beds with planar stratification, associated with cross-bedded sets and some undulating bedforms, indicate that the highly aggrading non-chanellized flows that buried the Darwin Forest were related to pyroclastic events. This concurs with the interpretation by Poma et al. (Citation2004), who suggested that the Darwin Forest was buried suddenly by a dilute, subaerial, cool and wet pyroclastic base surge deposit.
Darwin Forest
Three vertical sections were measured at localities A and C where the in situ fossil forest emerges (, 6, 7, 8A–E). The first one was compiled at locality A () where new fossiliferous levels (FL) were recognized in addition to the classic forest horizon: (1) FL I with Cladophlebis mesozoica Frenguelli, Citation1947, Cladophlebis mendozaensis (Geinitz) Frenguelli, Citation1947, Cladophlebis kurtzi Frenguelli, Citation1947 (Brea Citation2000); (2) FL III with poorly preserved Cladophebis species (Brea Citation2000); (3) FL IV with Araucarioxylon protoaraucana Brea, Citation1997 and Cuneumxylon spallettii Artabe & Brea, 2003. The second profile was measured at locality C where another fossiliferous level (FL V) was recognized (). A third detailed section was measured at locality C and the forest floor layer was recorded (). Finally, at locality D, only one fossiliferous level (FL VI) was found. These fossiliferous localities occur at a single stratigraphic level, so the stumps found in FL IV (locality A), FL V (localities B and C) and FL VI (locality D) belong to the same in situ fossil forest here collectively called the Darwin Forest.
Fig. 8 General view over Agua de la Zorra area. A–B, Natural longitudinal sections of tall stump moulds at locality C. C, Portion of an exposed trunk of Cuneumxylon spallettii at locality A. D, Natural cross-section of an in situ stump of Cuneumxylon spallettii at locality A. E, Natural cross-section view of in situ stump of Araucarioxylon protoaraucana at locality D. scales: A = note human figure to the left; C = 20 cm; B, D, E = 10 cm.

Fossil analysis considered taxonomic composition together with size and spatial distribution of trees, from which the fossil forest could be classified taking into account the histograms of tree diameter, vertical stature, indicating canopy position, dominance and total basal area and biomass/ha.
Taxonomic affinities of the wood
The corystosperm and coniferous in situ stumps that dominate the fossil forest have been examined previously and assigned to Cuneumxylon spallettii Artabe & Brea, Citation2003 and Araucarioxylon protoaraucana Brea, 1997. Cuneumxylon spallettii retains features of the pith, primary xylem and successive rings of secondary xylem and phloem. The pith is heterogeneous with parenchyma and polyhedral sclerenchyma cells. The primary xylem strands surrounding the pith are composed of endarch protoxylem and centrifugal metaxylem, with tracheids in radial files and a gradual transition from primary xylem to first-formed secondary xylem. The secondary xylem tracheid pitting is restricted to radial walls and is uniseriate to biseriate. Cross-fields have 2–13 circular or oval simple pits in irregular or, in some cases, vertical arrangement. Cuneumxylon spallettii was considered a corystosperm because, like Rhexoxylon, it exhibits two kinds of unusual centrifugal secondary growth. The first shows unequal activity of different portions of the cambium on the circumference of the axes; consequently, the reduction of cambial activity to certain restricted areas develops wedges of xylem that commonly split. The second pattern results in polyxylic stems following supernumerary cambial activity. The anomalous secondary growth develops secondary xylem with included phloem probably derived from successive cambia (Artabe & Brea Citation2003).
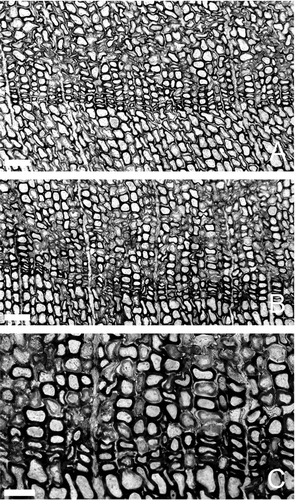
Araucarioxylon protoaraucana has features typical of conifer woods and is most similar to genera of the Araucariaceae based on the presence of tracheids with radial alternate intervascular pitting. The secondary xylem tracheids have intervascular pitting on radial and tangential walls. The tracheid pitting is bordered, uniseriate or biseriate with alternate or subopposite distribution. There is no axial parenchyma and the cross-fields have 3–9 bordered pits of araucarian-type with irregular or, in some cases, vertical arrangement (Brea Citation1997). Araucarioxylon protoaraucana has subtle growth rings (–C).
Fig. 9 Araucarioxylon protoaraucana Brea. A–C, Growth rings in transverse section. Scale bars: A–B = 100 μm; C = 60 μm. Note that growth direction is towards the bottom of each illustration.
Forest structure
Spatial analysis
The palaeovegetation of Agua de la Zorra region constituted a mixed forest of corystosperms (30–40%) and conifers (60–70%). Statistical analysis of the four mapped forest areas (localities A, B, C and D) using the Crimestat program showed that the mean nearest-neighbour distance is 3.63–4.84 m, and the density is 427 to 759 tree/ha with an average of 556 tree/ha (, 2). Nearest-neighbour index (Nni) and nearest-neighbour hierarchical spatial clustering (Nnh) were used for vegetation analysis. The Nni allows identification of tree clustering by dividing the empirical average distance to the nearest-neighbour by a spatially random expected distance. To identify groups that are spatially close, Nnh uses two criteria: the threshold distance (at Darwin Forest, the probability selected was 0.99) and the minimum number of trees (at Darwin Forest, six trees) to be included in each first-order cluster. The resulting data (see the nearest-neighbour index in ) indicate that trees are not grouped in localities B or D. The spatial distribution pattern illustrates a continuous forest. The Nni of localities A and C shows a clustered pattern, but the Nnh demostrates that there is only one major group in each stand. Therefore, despite the Nni results, we infer that the vegetation pattern is not clustered because the program only selected the best-preserved stump areas.
Table 1 . Nearest-neighbour analysis based on a sample size of 120 stumps distributed in four localities of the Agua de la Zorra region
Although the stand densities in modern conifer forests vary widely as a result of factors such as the population dynamics of the community, site disturbance regime and stand age (Pole Citation1999, Donoso Citation1993), neotropical and tropical extant forests (Pole Citation1999) show density values similar to the Darwin Forest. The Andean forests of Patagonia are defined as ‘temperate forests’ and are dominated by broad-leafed angiosperms (Nothofagus being its most important component) in association with a few conifers such as Araucaria araucana. These forests locally contain pure stands of A. araucana in drought-succeptible areas (Gallo et al. Citation2004). Typical dense Araucaria forests at Lanín National Park have a density of more than 200 trees/ha (Gallo et al. Citation2004).
The tree density of the Darwin Forest (556 trees/ha) is lower than that of the Late Triassic forest at La Elcha Mine, Río Blanco Formation, Cuyo Basin, Argentina (Artabe et al. Citation2007). The latter in situ monotypic forest of corystosperms has a higher density (1114 trees/ha) with a clustered distribution of trees. The Middle Triassic monotypic corystosperm forest found in Antarctica (Cúneo et al. Citation2003) has a density of 274 trees/ha.
Spatial autocorrelation is one of the most important concepts in spatial statistics. Spatial independence is an arrangement where no spatial relationships exist between the trees. Geary's C Statistic was used to identify spatial autocorrelation and clustering of the different species (using the CrimeStat II program: Levine, Citation2002). If trees of any one zone are spatially unrelated to any other variable, then the expected value of C is 1. At localities B and C, the ‘C’ values are close to 1 (the ‘C’ value at locality B is 0.9 and at locality C is 0.9). Therefore, corystosperms and conifers are considered to be intermingled at these localities, whereas at locality A, the C value is 0.653195 indicating spatial autocorrelation among species. This positive spatial autocorrelation at locality A suggests that corystosperms and conifers were aggregated into cohesive population groups. This is supported by the exclusive conifer presence at D locality. Thus, species are generally intermingled in the Darwin Forest but locally they form monospecific clusters.
Vertical profile
The histogram of diameter classes shows a normal distribution for corystosperms and a bimodal trend for conifers (, B; ). The normal curve indicates that the corystosperms belong predominantly to a single age cohort. The bimodal curve suggests that conifers constituted two strata of different age. When both species are plotted together, a bimodal curve is produced (), so the forest is represented by different age cohorts.
Fig. 10 Histograms of diameter class. A, Conifer stumps. B, Corystosperm stumps. C, All stumps of the forest.
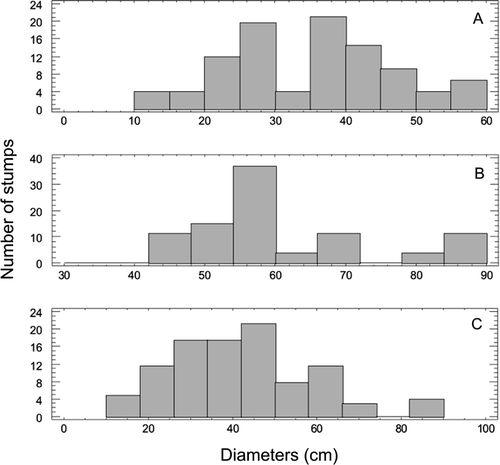
Table 2 . Structural data from the Darwin fossil forest. Basal diameters of trunks from Araucarioxylon protoaraucana Brea, Citation1997 and Cuneumxylon spallettii Artabe & Brea, Citation2003 were used to calculate the critical buckling heights (H crit), the estimated height (H est1) and the Safety Factor following Niklas (Citation1993, Citation1994a,b). Biomass is estimated using the equation of Murray (Citation1927: Method I), the subtropical dry forest equation of Brown (Citation1997: Method II) and the subtropical moist forest equation of Brown (Citation1997: Method III)
To analyse the vertical profile distribution of the vegetation, the basal diameters of trunks were used to estimate the critical bucking heights (Hcrit), the estimated height (Hest1) and the Safety Factor based on the allometric equations of Niklas (Citation1993, Citation1994a,Citationb; ).
Cuneumxylon spallettii had an average estimated tree height of 22.59 m when living. Base on the equations of Niklas (Citation1993, Citation1994a,Citationb), the maximum measured diameter corresponds to a maximum height of 29.67 m, whereas the minimum measured diameter of 0.30 m would have represented an individual up to 14.37 m tall. Araucarioxylon protoaraucana has an average estimated height of 17.91 m. A stump diameter of 0.60/0.10 m corresponds to minimum/maximum height of 8.00/26.09 m. Fifty percent of corystosperms are in the estimated height range 20.5–23.5 m, whereas 50% of conifers ranged between 16 and 20 m based on trunk diameter data (). As stature is the major factor controlling canopy position, the deduced heights of Cuneumxylon and Araucarioxylon (considering the distribution of class diameters) suggest that the forest had two strata. The highest stratum (20–24 m) had a dominance of corystosperms but also the tallest conifers. The second stratum (16–20 m) was mostly composed of conifers. A few emergent corystosperms probably reached 30 m tall.
Dominance, basal area and biomass
In vegetation studies, dominance refers to the capacity of one species to exert overriding influence upon others within the community. The dominant plant species is usually the one with the greatest biomass in the ecosystem and, therefore, controls the microclimatic conditions for all others. Dominance was quantified by Cottam & Curtis (Citation1956) and can be assessed for each species by multiplying the mean basal area by the number of trees per species per hectare. Cuneumxylon spallettii appears to be the dominant species with a dominance value of 108.62 m2/ha, whereas that for Araucarioxylon protoaraucana is just 22.48 m2/ha.
The total basal area was calculated by multiplying the number of trees per hectare by the mean basal area per hectare (Cottam & Curtis Citation1956). The total basal area values vary between the four exposures of the Darwin Forest accordingly: locality A, 26.64 m2/ha; locality B, 19.78 m2/ha; locality C, 46.06 m2/ha and locality D, 20.20 m2/ha. Considering an average density of 556 trees/ha, the total basal area is 28.12 m2/ha (). This value typically ranges between 20 and 50 m2/ha in extant forests. The monotypic corystosperm forest found in the Middle Triassic of Antarctica (Cúneo et al. Citation2003) has a total basal area value of 20.83 m2/ha, whereas for the monotypic Late Triassic La Elcha forest of Argentina, this parameter ranges from 32.67 m2/ha to 85.6 m2/ha (Artabe et al. Citation2007).
Biomass was estimated in the following three ways (see Material and methods for details). Method I (of Murray Citation1927, cited by Mosbrugger et al. Citation1994) predicted a dry weight (W d) of 92.91 tn/ha for the Darwin Forest. On this basis, Darwin Forest could be classified as a moist-dry forest or a dry forest. Using Brown's equation (Citation1997) for subtropical dry forests, the biomass estimation was 47.02 tn/ha, and Brown's equation (Citation1997) for subtropical moist forest yielded 62.06 tn/ha. The biomass for extant dry subtropical forest oscillates between 29 and 140 tn/ha (Brown & Lugo Citation1984, Murphy & Lugo Citation1986). The extant dry subtropical forests of Puerto Rico growing on volcanic soils have a biomass similar to that estimated for the Darwin Forest (Brandeis et al. Citation2005).
Structural classification: subtropical seasonal forest
The Darwin Forest was reconstructed based on the criteria outlined in the previous section together with ecological inferences based on morphlogical and anatomical characters of the whole plants. Corystosperms, like cycads, had some wood traits characteristic of plants growing in warm subtropical areas without frost or with only occasional frost. In addition, they have several attributes (secondary pycnoxyly, growth rings, sclerophylly, secretory reservoirs, thick cuticles, papillae and sunken stomata) typical of plants from seasonal climates (Petriella Citation1985, Spalletti et al. Citation2003). The included phloem and the pycnoxylic habit developed by southwestern Gondwanan Corystospermaceae might represent anatomical adaptations to semi-arid environments, and such features probably determined the remarkable success of the group during the Triassic.
The density, total basal area, biomass and canopy structure allow classification of the Darwin Forest as a subtropical seasonal forest. Almost all vegetation physiognomic types owe their origins to the distinctive climate of the region, particularly rainfall and temperature. Models of the Triassic climate of southwestern Gondwana indicate strong seasonality (Spalletti et al. Citation2003).
Growth-ring analysis
The pattern of leaf-shed may vary from evergreen to dry-season deciduous in extant subtropical seasonal (monsoon) forests. Falcon-Lang (Citation2000a) developed a method to distinguish between evergreen and deciduous gymnoperms based on CSDM curves. The growth rings of the conifer Araucarioxylon protoaraucana were analysed quantitatively using this technique. The CSDM curve was plotted and four parameters were calculated: (1) skew of CSDM curves, (2) percentage of latewood, (3) percentage of cell diminution in a ring increment and (4) Ring Markedness Index (RMI).
Deciduous conifers have CSDM curves that are dominantly left-skewed or symmetrical, whereas evergreen conifers have dominantly right-skewed CSDM curves. The CSDM curves of Araucarioxylon protoaraucana are right-skewed (mean percentage of skew +21.34%; range: +13.33% to +26.31%), suggesting that this species was evergreen.
Although the percentage of latewood has traditionally been used to infer climatic conditions (Creber & Chaloner Citation1984a,Citationb, Parrish & Spicer Citation1988, Keller & Hendrix Citation1997), its use has been regularly questioned (Bailey & Faull Citation1934, Larson Citation1967, Chapman Citation1994, Poole & van Bergen Citation2006). Falcon-Lang (Citation2000a,b) pointed out that latewood development in conifer woods could be related to foliar retention, a genetically determined characteristic. Because the percentage of latewood may be strongly influenced by both leaf longevity and the intensity of climate seasonality, Falcon-Lang (Citation2000a, Citation2005) recommended that this parameter should not be used as a climatic indicator. Nevertheless, we include its measurement here for the sake of completeness. The percentage of latewood in Araucarioylon protoaraucana is 32.62–42.85%, with a mean of 35.44%. The percentage of cell diminution varies between 68.69% and 75.60%, with an average of 72.10%.
The ring markedness index (RMI) for A. protoaraucana was in the range of 22.88–29.43%, with a mean of 25.50% (, ). Falcon-Lang (Citation2000a) used CSDM curves to determine a strong inverse linear relationship between the leaf retention and the different quantitative parameters of growth rings (percentage of latewood, percentage of diminution and ring markedness index) in a group of extant conifers. The lower values of percentage of latewood and ring markedness index correlate with the highest foliar retention in evergreen plants (Araucaria araucana: foliar longevity of 15 years), whereas higher values relate to shorter periods of leaf retention. Based on the values recorded for extant conifers, the results (percentage of skew, latewood, cell diminution and RMI values) for Araucarioxylon protoaraucana indicate a foliar retention similar to those extant conifers with 3–6 years of foliar retention ().
Fig. 11 Right-skewed CSDM curves of five growth rings and cell diameters of growth ring increment. For each ring increment, the percentage of skew for CSDM curves was calculated using Falcon-Lang's (Citation2000a) method (see ). The position at which the CSDM curve reaches the zenith represents the percentage of skew in relation to the total distance between the centre of the CSDM curve to the right of the plot.
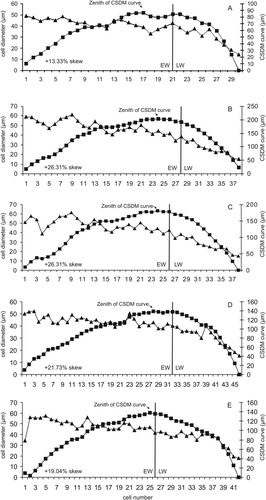
Table 3 . Results of the quantification of ring markedness parameters for Araucarioxylon protoaraucana Brea, 1997
Table 4 . Quantification of ring markedness parameters for the five extant taxa taken from the data of Falcon-Lang (Citation2000a,Citationb: see the CSDM curves in Fig. 11). The species shown in bold corresponds to the wood fossil studied in this paper
Because of their anomalous centrifugal secondary growth, the Falcon-Lang year method could not be used to infer the leaf shed pattern of Cuneumxylon. Although data for Cuneumxylon are insufficient, other Corystospermaceae (e.g. Rhexoxylon and Elchaxylon) have been interpreted as evergreen (Artabe et al. Citation2007). Petriella (Citation1978) reconstructed the ‘Dicroidium’ plant based on the consistent association of Dicroidium zuberi–Rhexoxylon piatnitzkyi found in the Ischigualasto Formation, Argentina (Archangelsky & Brett Citation1961, Archangelsky Citation1968, Citation1970). The Dicroidium plant seems to have had a palm-like habit, bearing Pteruchus pollen organs and Umkomasia ovulate structures. Spirally arranged leaf bases preserved higher on the Rhexoxylon stem (Lutz & Herbst Citation1992) agree with a palm-like reconstruction (Petriella Citation1978) incorporating a crown of leaves in the upper part of the tree. Crown trees, with persistent leaf bases, such as palms and cycads, are expected to have been typically evergreen (Tomlinson Citation1961, Henderson et al. Citation1995, Norstog & Nicholls Citation1997). The other Corystospermaceae genus mentioned above, Elchaxylon, described from the Late Triassic (Río Blanco Formation) of Argentina, is related to Rhexoxylon because its pycnoxylic wood has a unique mode of secondary xylem production resulting in polyxylic stems. However, unlike Rhexoxylon and Cuneumxylon, Elchaxylon has centrifugal secondary growth produced by a continuous cambium that generates an undivided solid pycnoxylic cylinder (Artabe & Zamuner Citation2007). This feature allows the application of the quantitative secondary wood growth-ring analysis method described by Falcon-Lang (Citation2000a,b) to infer the tree's habit (Artabe et al. Citation2007). Analysis of Elchaxylon zavattierae growth rings reveals CSDM curves with skew values ranging from +12.50 to +24.63% (mean = +17.89%, n = 4; see Artabe et al. Citation2007), thus implying that these trees could have been evergreen.
Twenty-four ring sequences ranging from 6 to 36 rings were measured from eight well-preserved fragments of fossil wood (). The total measured rings of the whole series was 428 and their presence indicates seasonality. Araucarioxylon protoaraucana has narrow, subtly demarcated growth rings (). Each growth increment is characterized by a relative wide zone of large, thin-walled earlywood cells terminated by 10–17 thick-walled latewood cells (). The presence of narrow growth rings indicates that conditions during the growing season were not strongly favourable for cell division and expansion (Creber & Chaloner Citation1984a). In addition, a thin latewood zone within a ring can be the result of water shortage at the end of the growing season, a sharp photoperiod-controlled end to the growth season or abrupt leaf shed. Therefore, the growth rings of A. protoaraucana suggest strongly seasonal conditions.
Fig. 12 Graphs showing variation in width of growth rings through ring sequences of selected fossil trees. A, specimen LPPB 12604, USM01B ring series. B, specimen LPPB 12605, USM02B ring series. C, specimen LPPB 12609, USM13A ring series. D, specimen LPPB 12610, USM15E ring series. MW = mean width, MS = mean sensitivity.
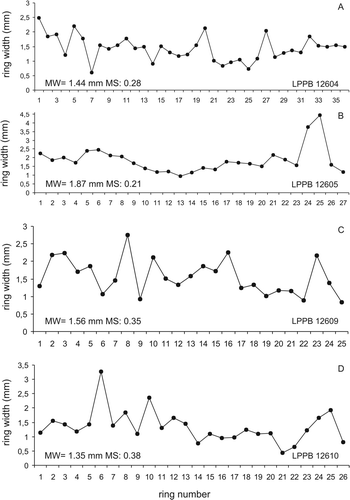
Table 5 . Summary of the tree-ring data for Araucarioxylon protoaraucana Brea, Citation1997 fossil wood
The mean ring width for A. protoaraucana is 1.49 mm; the mean minimum ring width is 0.80 mm; and the mean maximum ring width is 2.53 mm. The narrowest ring is 0.12 mm wide and the broadest is 4.44 mm. The variability in ring width may reflect either fluctuations in annual water supply or changes in the growth environment (e.g. variation in shading; ).
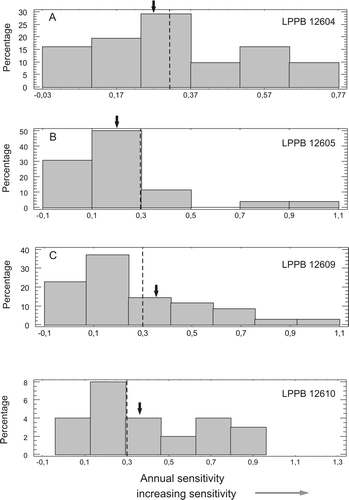
MS values (, ) range between 0.14 and 0.62, the average being 0.30. These data indicate that the growing environment was not uniform. MS values suggest that the forest community was part of a stressed ecosystem. AS ranges from 0.02 to 0.79. The largest values of AS indicate extreme climatic events (). Frequency distributions of AS of selected samples () show variations in ring thickness between adjacent years. The AS values for each tree illustrate that the variation in growth was generally low, since a high proportion of the AS values are less than 0.3. However, each tree has some AS values of the sensitive kind reflecting several periods of more marked climatic variability (Francis Citation1986). The sudden and strong pulses of volcaniclastic sedimentation alternating with periods of reduced sediment accumulation during which immature soils developed indicate that volcanism and extreme climatic events may have periodically stressed the growth of the Darwin Forest.
Fig. 13 Histograms allowing comparison of the variation in the annual sensitivities of selected fossil trees. A, specimen LPPB 12604. B, specimen LPPB 12605. C, specimen LPPB 12609. Arrows indicate the value of mean sensitivity.
The growth trends estimated by ABIC minimization methods were plotted for the longest growth-ring series. All the curves show complex trends. Even though available data are insufficient, these growth trends could indicate that competition, disturbance events, or climatic stress influenced the growth of trees in the Darwin Forest (Kumagai et al. Citation1995).
The age of conifers was estimated using the radius of stumps and growth-ring widths (Frangi Citation1976, Pole Citation1999). On the basis of 86 trunks, the average estimated age was 146 years (the maximum age was 244 years, and the minimum was 41 years). Therefore, if the oldest trees were about 250 years old, then trees of the fossil forests from the Paramillo Formation may have been younger than typical mature conifers in extant forests. According to Tortorelli (Citation1956), modern Araucaria araucana forests are typically about 300 years old (corresponding to a stump diameter of 0.40 m); the oldest trees and the tallest found near Lake Aluminé are about 1000 years old.
Conclusions
-
The in situ Middle Triassic mixed forest occurs in one fossiliferous stratum located in the Paramillo Formation (equivalent to the lower section of the Potrerillos Formation) at Agua de la Zorra region, northwestern Cuyo Basin, Mendoza province (69°12′W and 32°30′S). The Darwin Forest consists of 120 stumps in life position emerging at four localities: A, B, C and D.
-
The Paramillo Formation was deposited in a high sinuosity fluvial system, in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. Darwin Forest is located within a highly aggrading non-chanellized succession. The volcanic nature of detrital components suggests that the deposits that buried the Darwin Forest could represent a diluted, subaerial, cool and wet base surge pyroclastic flow.
-
The permineralized stumps found at Darwin Forest are referable to Cuneumxylon spallettii Artabe & Brea (Corystospermaceae) and Araucarioxylon protoaraucana Brea (Araucariaceae). The Darwin Forest was a mixed forest composed of corystosperms (30–40%) and conifers (60–70%).
-
Forest density ranges between 427 and 759 trees/ha, with an average of 556 trees/ha. In some places, corystosperms and conifers are intermingled but in others corystosperms and conifers appear to be aggregated into cohesive monotypic groups.
-
The histogram of diameter classes reveals a normal curve for the corystosperms and a bimodal curve for conifers. The normal curve indicates that the corystosperms mostly belonged to a single age cohort. The bimodal curve suggests that the conifers occurred in two strata of different age.
-
Corystosperms have a mean estimated height (H est1) of 22.59 m (14.37–29.67 m), whereas conifers are smaller with a mean estimated height (H est1) of 18 m (8–26.08 m).
-
The forest has two strata: the highest one (20–24 m) has a predominance of corystosperms and the tallest conifers; the second stratum (16–20 m) consists mostly of conifers. The forest also has emergent Cuneumxylon spallettii (corystosperm) reaching 30 m tall.
-
The biomass, stand basal area and biomass of Darwin Forest correlate with those of extant subtropical seasonal forests (dry monsoonal forests).
-
Falcon-Lang's (year) quantitative growth-ring anatomy analysis method indicated that Araucaria protoaraucana was an evergreen gymnosperm based on right-skewed CSDM curves.
-
The morpho-functional and structural analyses of Darwin Forest woods indicate that the vegetation represented a mainly evergreen dry subtropical seasonal forest.
-
The mean ring width and MS of A. protoaraucana were 1.49 mm and 0.30, respectively. Variability in ring width may have resulted from fluctuations in annual water supply or other changes in the growth environment such as shading. MS values indicate that the growing environment was stressed and not uniform. Although available data are limited, the complicated growth trends could indicate that competition, disturbance events, or climatic stress influenced the growth of trees in the Darwin Forest.
-
The average width of the A. protoaraucana trunks and their mean growth-ring widths indicate a mean age of 146 years. This suggests that the conifers of the Darwin Forest may have been composed of relatively young trees.
Acknowledgements
We would like to thank the financial support provided by the Consejo Nacional de Investigaciones Científicas y Técnicas (Project PIP 5430) and the Agencia Nacional de Promoción Científica y Tecnológica (PICT 07-08467). Mariana Brea expresses her acknowledgement to José Boninsegna, Ricardo Villalba and Fidel Roig Jr of the IANIGLA-CONICET, for their valuable help with the growth-ring analysis, and Richard Holmes, who made the use and handling of the COFECHA program possible. The authors express their acknowledgement to the editor of Alcheringa for correcting the English text.
Related Research Data
References
- Akaike , H. 1980 . “ Likelihood and Bayes procedure ” . In Bayesian Statistics , Edited by: Bernardo , J. M. , De Groot , M. H. , Lindley , D. U. and Smith , A. F.M. 143 – 166 . Valencia : University Press .
- Archangelsky , S. 1968 . Studies on Triassic fossil plants from Argentina IV. The leaf genus Dicroidium and its possible relation to Rhexoxylon stems . Palaeontology , 11 : 500 – 512 .
- Archangelsky , S. 1970 . Fundamentos de Paleobotánica Vol. 10 , 347 Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Serie técnica y Didáctica
- Archangelsky , S. and Brett , D. 1961 . Studies on Triassic fossil plants from Argentina. 1. Rhexoxylon from the Ischigualasto Formation . Philosophical Transactions of the Royal Society of London , 706B ( 244 ) : 1 – 19 .
- Artabe , A. E. and Brea , M. 2003 . A new approach to Corystospermales based on petrified stems from the Triassic of Argentina . Alcheringa , 27 : 209 – 229 .
- Artabe , A. E. and Zamuner , A. E. 1991 . Una nueva equisetal del Triásico de Cacheuta, Argentina, con estructura interna preservada . Ameghiniana , 28 : 287 – 294 .
- Artabe , A. E. and Zamuner , A. E. 2007 . Elchaxylon, a new corystosperm based on permineralized stems from the Late Triassic of Argentina . Alcheringa , 31 : 85 – 96 .
- Artabe , A. E. , Morel , E. M. and Spalletti , L. A. 2001 . “ Paleoecología de las floras triásicas argentinas ” . In El Sistema Triásico en la Argentina Edited by: Artabe , A. E. , Morel , E. M. and Zamuner , A. E. 199 – 225 . Fundación Museo de La Plata ‘Fracisco P. Moreno’, La Plata
- Artabe , A. E. , Morel , E. M. and Spalletti , L. A. 2003 . Caracterización de las provincias fitogeográficas triásicas del Gondwana extratropical . Ameghiniana , 40 : 387 – 405 .
- Artabe , A. E. , Spalletti , L. A. , Brea , M. , Iglesias , A. , Morel , E. M. and Ganuza , D. G. 2007 . Structure of a corystosperm fossil forest from the Late Triassic of Argentina . Palaeogeography, Palaeoclimatology, Palaeoecology , 243 : 451 – 470 .
- Bailey , I. W. and Faull , A. F. 1934 . The cambium and its derivative tissues. 9. Structural variability in the redwood, Sequoia sempervirens, and its significance in the identification of fossil woods . Journal of the Arnold Arboretum , 15 : 99 – 106 .
- Brandeis , T. J. and Suárez Rozo , M. R. 2005 . Effects of model choice and forest structure on inventory-based estimations of Puerto Rican Forest biomass . Caribbean Journal of Science , 41 : 250 – 268 .
- Brea , M. 1995 . Estudio de la paleoflora de la secuencia triásica de Agua de la Zorra, provincia de Mendoza , 202 Argentina : Universidad Nacional de La Plata . Ph. D. thesis, N° 642, Facultad de Ciencias Naturales y Museo
- Brea , M. 1997 . Una nueva especie fósil del género Araucarioxylon Kraus 1870 emend. Maheshwari 1972 del Triásico de Agua de la Zorra, Uspallata, Mendoza, Argentina . Ameghiniana , 34 : 485 – 496 .
- Brea , M. 2000 . Paleoflora triásica de Agua de la Zorra, Uspallata, provincia de Mendoza, Argentina: Lycophyta y Filicophyta . Ameghiniana , 37 : 199 – 204 .
- Brea , M. and Artabe , A. E. 1999 . Apocalamitaceae (Sphenophyta) triásicas de la Formación Paramillo, Agua de la Zorra, provincia de Mendoza, Argentina . Ameghiniana , 36 : 389 – 400 .
- Brown , S. 1997 . Estimating biomass and biomass change in tropical forests. FAO Forestry Paper 134 , 49 Rome : Food and Agriculture Organization of the United Nations .
- Brown , S. , Gillespie , A. J.R. and Lugo , A. E. 1991 . Biomass of tropical forests of south and southeast Asia . Canadian Journal of Forest Research , 21 : 111 – 117 .
- Brown , S. and Lugo , A. E. 1984 . Biomass of tropical forests: A new estimate based on forest volumes . Science , 223 : 1290 – 1293 .
- Chapman , J. L. 1994 . Distinguishing internal developmental characteristics from external palaeoenvironmental effects in fossil wood . Review of Palaeobotany and Palynology , 81 : 19 – 32 .
- Chebli , G. A. , Ploszkiewicz , V. and Azpiroz , G. M. 2001 . “ El sistema Triásico y los hidrocarburos ” . In El Sistema Triásico en la Argentina Edited by: Artabe , A. E. , Morel , E. M. and Zamuner , A. E. 283 – 315 . Fundación Museo de La Plata ‘Fracisco P. Moreno’, La Plata
- Conwentz , H. 1885 . Árboles fósiles del Río Negro . Boletín de la Academia de Ciencias de Córdoba , 7 : 435 – 456 .
- Cottam , G. and Curtis , J. T. 1956 . The use of distance measures in phytosociological sampling . Ecology , 37 : 451 – 460 .
- Creber , G. T. 1977 . Tree-rings: a natural data storage system . Biological Review , 52 : 349 – 383 .
- Creber , G. T. and Ash , S. R. 2004 . The Late Triassic Schilderia adamanica and Woodworthia arizonica trees of the petrified forest National Park, Arizona, USA . Palaeontology , 47 : 21 – 38 .
- Creber , G. T. and Chaloner , W. G. 1984a . “ Climatic indications from growth rings in fossil woods ” . In Fossil and Climate , Edited by: Brenchley , P. 49 – 73 . London : Wiley .
- Creber , G. T. and Chaloner , W. G. 1984b . Influence of environmental factors on the wood structure of living and fossil trees . Botanical Review , 50 : 357 – 448 .
- Creber , G. T. and Francis , J. E. 1999 . “ Tree ring analysis: palaeodendrochronology ” . In Fossil Plants and Spores: Modern Techniques , Edited by: Jones , T. and Rowe , N. 245 – 250 . London : The Geological Society .
- Cúneo , N. R. , Taylor , E. L. , Taylor , T. N. and Kring , M. 2003 . In situ fossil forest from the upper Fremouw Formation (Triassic) of Antarctica: paleoenvironmental setting and paleoclimate analysis . Palaeogeography, Palaeoclimatology, Palaeoecology , 197 : 239 – 261 .
- Darwin , C. 1839a . Narrative of the surveying voyages of His Majesty's ships Adventure and Beagle, between examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. In three volumes. Vol III. Journal and Remarks. 1832–1836 , 615 London : Henry Colburn .
- Darwin , C. 1839b . Journal of researches into the geology and natural history of the various countries visited by HMS Beagle under the command of Captain FitzRoy, R.N., from 1832 to 1836 , 519 London : Henry Colburn .
- Darwin , C. 1846 . Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, during the years 1832 to 1836 , 279 London : Smith, Elder and Co. .
- Donoso , C. 1993 . Bosques templados de Chile y Argentina. Variación, Estructura y Dinámica. Ecología Forestal , 484 Santiago de Chile : Editorial Universitaria .
- Du Toit , A. L. 1927 . The fossil flora of the Upper Karroo beds . Annals of South Africa , 22 : 289 – 420 .
- Evans , R. 1981 . COMPUTA (Measure and pulse counter, unpublished)
- Falcon-Lang , H. J. 2000a . A method to distinguish between woods produced by evergreen and deciduous coniferopsids on the basis of growth ring anatomy: a new palaeoecological tool . Palaeontology , 43 : 785 – 793 .
- Falcon-Lang , H. J. 2000b . The relationship between leaf longevity and growth ring markedness in modern conifer woods and its implications for palaeoclimatic studies . Palaeogeography, Palaeoclimatology, Palaeoecology , 160 : 317 – 328 .
- Falcon-Lang , H. J. 2005 . Global climate analysis of growth rings in woods, and its implications for deep-time paleoclimate studies . Paleobiology , 31 : 434 – 444 .
- Francis , J. E. 1986 . Growth rings in Cretaceous and Tertiary wood from Antarctica and their palaeoclimatic implications . Palaeontology , 29 : 665 – 684 .
- Frangi , J. L. 1976 . Descripción florístico-estructural de un ‘stand’ de bosque de Nothofagus dombeyi en lago Gutiérrez (Provincia de Río Negro) . Darwiniana , 20 : 577 – 585 .
- Frenguelli , J. 1947 . El género Cladophlebis y sus representantes en Argentina . Gaea , 7 : 159 – 369 .
- Fritts , H. C. 1976 . Tree Rings and Climate , 567 London : Academic Press .
- Gallo , L. , Izquierdo , F. , Sanguinetti , L. J. , Pinna , A. , Siffredi , G. , Ayesa , J. , Lopez , C. , Pelliza , A. , Strizler , N. , Gonzales Peñalba , M. , Maresca , L. and Chauchard , L. 2004 . “ Araucaria araucana forest genetic resources in Argentina ” . In Challenges in managing forest genetic resources for livelihoods: examples from Argentina and Brazil , Edited by: Vincenti , B. , Amaral , W. and Meilluer , B. 105 – 131 . Rome, , Italy : International Plant Genetic Resources Institute .
- Groeber , P. 1939 . Mapa geológico de Mendoza . Physis , 14 : 171 – 220 .
- Halle , T. G. 1908 . Zur Kenntnis der Mesozoischen Equisetales Schwedens . Kungliga Svenska Vetenskaps-Akademiens Handligar , 3 : 1 – 56 .
- Hammer , O. , Harper , D. A.T. and Ryan , P. D. 2007 . Past—Palaeontological Statistics, ver. 1.75 86 [originally released 2001, Palaeontologia Electronica 4(1)8, 1–9]
- Harrington , H. J. 1971 . Descripción geológica de la hoja 22c, ‘Ramplón’ provincias de Mendoza y San Juan. Carta Geológica–Económica de la República Argentina Escala 1:200.000 . Ministerio de Industria y Minería, Subsecretaria de Minería, Dirección de Geología y Minería, Buenos Aires, Boletín , 114 : 1 – 87 .
- Henderson , A. , Galeano , G. and Bernal , R. 1995 . Field Guide to the Palms of the Americas , 351 Princeton, NJ : Princeton University Press .
- Holdridge , L. R. 1979 . Ecología basada en zonas de vida. Instituto Interamericano de Ciencias Agrícolas , 216 Costa Rica : San José . Ed. IICA
- Holmes , R. L. 1985 . Computer-assisted quality control in tree-ring dating and measurement. A users manual for program COFECHA , 23 Tucson : University of Arizona . Laboratory of tree-ring Research
- Keller , A. M. and Hendrix , M. S. 1997 . Paleoclimatological analysis of a Late Jurassic petrified forest, southeastern Mongolia . Palaios , 12 : 282 – 291 .
- Kidwell , S. M. , Fursich , F. T. and Aigner , T. 1986 . Conceptual framework for the analysis and classification of fossil concentrations . Palaios , 1 : 228 – 238 .
- Kokogian , D. and Mancilla , O. 1989 . “ Análisis estratigráfico y secuencial de la Cuenca Cuyana ” . In Cuencas Sedimentarias Argentinas Edited by: Chebli , G. and Spalletti , L. 169 – 201 . Universidad Nacional de Tucumán, Serie Correlación Geológica 6
- Kokogián , D. , Fernández Seveso , F. and Mosquera , A. 1993 . “ Las secuencias sedimentarias triásicas ” . In Relatorio Geología y Recursos Naturales de Mendoza Edited by: Ramos , V. A. 65 – 78 . 12° Congreso Geológico Argentino y 2° Congreso de Exploración en Hidrocarburos Actas 1
- Kumagai , H. , Sweda , T. , Hayashi , K. , Kojima , S. , Basinger , J. F. , Shibuya , M. and Fukaoa , Y. 1995 . Growth-ring analysis of Early Tertiary conifer woods from the Canadian High Arctic and its paleoclimatic interpretation . Palaeogeography, Palaeoclimatology, Palaeoecology , 116 : 247 – 262 .
- Kurtz , F. 1921 . Sur l'existence d'une flore rajmahalienne dans le Gouvernement du Neuquén (Piedra Pintada, entre Limay et Colon-curá) . Revista del Museo de La Plata , 10 : 235 – 242 .
- Larson , R. R. 1967 . “ Silvicultural control on the characteristics of wood used for furniture ” . In Proceedings of the 4th TAPPI Forest Biology Conference , 143 – 150 . Quebec : Pulp and Paper Research Institute of Canada . Anon. ed
- Levine , N. 2002 . CrimeStat II: A Spatial Statistics Program for the Analysis of Crime Incident Locations (v 2.0) , Washington, DC : Ned Levine & Associates, Houston, TX, and National Institute of Justice .
- Linares , E. and González , R. 1990 . Catálogo de edades radimétricas de la República Argentina 1957–1987 1 – 628 . Asociación Geológica Argentina, Serie B (Didáctica y Complementaria), Buenos Aires 19
- Lutz , A. I. and Herbst , R. 1992 . Una nueva especie de Rhexoxylon del Triásico de Barreal, San Juan, Argentina . 7° Simposio Argentino de Paleobotánica y Palinología (Corrientes 1991). Publicación Especial de la Asociación Paleontológica Argentina , 2 : 73 – 76 .
- Massabie , A. C. 1985 . Filón capa Paramillo de Uspallata, su caracterización geológica y edad, Paramillo de Uspallata, Mendoza . Primeras Jornadas sobre Geología de Precordillera Actas , 1 : 325 – 330 .
- Massabie , A. C. , Rapalini , A. E. and Soto , J. L. 1985 . Estratigrafía del Cerro Los Colorados, Paramillo de Uspallata, Mendoza . Primeras Jornadas sobre Geología de Precordillera Actas , 1 : 71 – 76 .
- Moratello , J. 1993 . “ Cuenca Cuyana, recursos de hidrocarburos ” . In Geología y Recursos Naturales de Mendoza Edited by: Ramos , V. A. 367 – 375 . 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos Relatorio 3
- Morel , E. M. , Artabe , A. E. and Spalletti , L. A. 2003 . The Triassic floras of Argentina: Biostratigraphy, floristic events and comparison with other areas of Gondwana and Laurasia . Alcheringa , 27 : 231 – 243 .
- Mosbrugger , V. 1990 . The tree habit in land plants—a functional comparison of trunk constructions with a brief introduction into the biomechanics of trees , 161 Berlin : Springer .
- Mosbrugger , V. , Gee , C. T. , Belz , G. and Ashraf , A. R. 1994 . Three-dimensional reconstruction of an in-situ Miocene peat forest from the Lower Rhine Embayment, northwestern Germany—new methods in palaeovegetation analysis . Palaeogeography, Palaeoclimatology, Palaeoecology , 110 : 295 – 317 .
- Murphy , P. G. and Lugo , A. E. 1986 . Structure and biomass of a subtropical dry forest in Puerto Rico . Biotropica , 18 : 89 – 96 .
- Murray , C. D. 1927 . A relationship between circumference and weight in trees and its bearing on branching angles . Journal of the Society of General Physiologists , 10 : 725 – 728 .
- Niklas , K. J. 1992 . Plant Biomechanics. An Engineering Approach to Plant Form and Function , 622 Chicago : The University of Chicago Press .
- Niklas , K. J. 1993 . The scaling of plant height: a comparison among major plant clades and anatomical grades . Annals of Botany , 72 : 165 – 172 .
- Niklas , K. J. 1994a . The allometry of safety—factors for plant height . American Journal of Botany , 81 : 345 – 351 .
- Niklas , K. J. 1994b . Predicting the height of fossil plant remains: an allometric approach to an old problem . American Journal of Botany , 81 : 1235 – 1242 .
- Norstog , K. J. and Nicholls , T. J. 1997 . The Biology of the Cycads , 363 Ithaca : Cornell University Press .
- Ottone , E. G. 2005 . “ The history of palaeobotany in Argentina during the 19th century ” . In History of Palaeobotany: Selected Essays Edited by: Bowden , A. J. , Burek , C. V. and Wilding , R. 281 – 294 . Geological Society, London, Special Publications 241
- Parrish , J. T. and Spicer , R. A. 1988 . Middle Cretaceous wood from the Nanushuk Group, Central North Slope, Alaska . Palaeontology , 31 : 19 – 34 .
- Petriella , B. 1978 . La recostrucción de Dicroidium (Pteridospermopsida, Corystospermaceae) . Obra del Centenario del Museo La Plata , 5 : 107 – 110 .
- Petriella , B. 1985 . “ Caracteres adaptativos y autoecología de las Corystospermaceae ” . In 3° Congreso Latinoamericano de Paleontología (Mexico, 1984). Simposio Sobre Floras del Triásico, tardio su fitogeografia paleogeografia Edited by: Weber , R. 53 – 57 . Memorias del Instituto Geológico de la Universidade Nacional Autónomade México
- Pole , M. 1999 . Structure of a near-polar latitude forest from the New Zealand Jurassic . Palaeogeography, Palaeoclimatology, Palaeoecology , 147 : 121 – 139 .
- Poma , S. , Quenardelle , S. and Koukharsky , M. 2004 . Un flujo piroclástico del Triásico en la Formación Paramillo, provincia de Mendoza . 7° Congreso Argentino de Mineralogía y Metalogenia , : 385 – 390 .
- Poole , I. and van Bergen , P. 2006 . Physiognomic and chemical characters in wood as palaeoclimate proxies . Plant Ecology , 182 : 175 – 195 .
- Prezzi , C. , Vizán , H. and Rapalini , A. 2001 . “ Marco paleogeográfico ” . In El Sistema Triásico en la Argentina Edited by: Artabe , A. E. , Morel , E. M. and Zamuner , A. B. 255 – 1267 . Fundación Museo de La Plata ‘Francisco P. Moreno’, La Plata
- Ramos , V. A. 1993 . “ El magmatismo Triásico-Jurásico de intraplaca ” . In Relatorio Geología y Recursos Naturales de Mendoza Edited by: Ramos , V. A. 79 – 86 . 12° Congreso Geológico Argentino y 2° Congreso de Exploración en Hidrocarburos Actas 1
- Ramos , V. A. and Kay , S. M. 1991 . “ Triassic rifting and associated basalts in the Cuyo Basin, central Argentina ” . In Andean magmatism and its tectonic setting Edited by: Harmon , R. S. and Rapela , C. W. 79 – 91 . Geological Society of America, Boulder, Special Paper 265
- Schweingruber , F. H. 1988 . Tree-Rings, Basics and Applications of Dendrochronology , 273 Boston : D. Reidel .
- Spalletti , L. A. 2001 . “ Evolución de las cuencas sedimentarias ” . In El Sistema Triásico en la Argentina Edited by: Artabe , A. E. , Morel , E. M. and Zamuner , A. B. 81 – 101 . Fundación Museo de La Plata ‘Francisco P. Moreno’, La Plata
- Spalletti , L. A. , Artabe , A. E. , Morel , E. M. and Brea , M. 1999 . Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino . Ameghiniana , 36 : 419 – 451 .
- Spalletti , L. A. , Artabe , A. E. and Morel , E. M. 2003 . Geological factors and evolution of southwestern Gondwana Triassic Plants . Gondwana Research , 6 : 119 – 134 .
- Stappenbeck , R. 1910 . La Precordillera de San Juan y Mendoza . Anales del Ministerio de Agricultura, sección Geología, Mineralogía y Minería , 4 : 3 – 183 .
- Stipanicic , P. N. 2001 . “ Antecedentes Geológicos y Paleontológicos ” . In El Sistema Triásico en la Argentina Edited by: Artabe , A. E. , Morel , E. M. and Zamuner , A. B. 1 – 21 . Fundación Museo de La Plata ‘Francisco Pascasio Moreno, La Plata
- Stipanicic , P. N. and Marsicano , C. A. 2002 . Léxico estratigráfico de la Argentina, Volumen VIII: Triásico . Asociación Geológica Argentina, Serie ‘B’ (Didáctica y Complementaria) , 26 : 1 – 370 .
- Stipanicic , P. N. , Herbst , R. and Bonetti , M. I.R. 1996 . Floras Triásicas . Actas de la Academia Nacional de Ciencias. Córdoba , 11 : 127 – 184 .
- Stockes , M. A. and Smiley , T. L. 1968 . An Introduction to tree-ring dating , 73 Chicago : University of Chicago Press .
- Strelkov , E. E. and Álvarez , L. A. 1984 . Análisis estratigráfico y evolutivo de la Cuenca Triásica Mendocina-Sanjuanina . 9° Congreso Geológico Argentino Actas , 3 : 115 – 130 .
- Tomlinson , P. B. 1961 . Anatomy of the Monocotyledons. II. Palmae , 453 Oxford : Clarendon Press .
- Tortorelli , L. A. 1956 . Maderas y Bosques Argentinos , 910 Buenos Aires : Editorial Acme .
- Windhausen , H. 1941 . El ‘Rético’ del Paramillo de Uspallata (Mendoza) , Argentina : Universidad Nacional de La Plata . PhD thesis N° 28, Facultad de Ciencias Naturales y Museo
- Zamuner , A. E. , Zavattieri , A. M. , Artabe , A. E. and Morel , E. M. 2001 . “ Paleobotánica ” . In El Sistema Triásico en la Argentina Edited by: Artabe , A. E. , Morel , E. M. and Zamuner , A. B. 143 – 184 . Fundación Museo de La Plata ‘Francisco P. Moreno’, La Plata