
ABSTRACT
To assess if the nutrient supply through planted tree fallows meets crop nutrient uptake and export, N, P, K, Ca and Mg uptake and export by a groundnut/maize/cassava intercrop was compared with the nutrient uptake by three planted fallow systems (Senna spectabilis, Flemingia macrophylla, Dactyladenia barteri) and a no-tree control. Three cycles of two years fallow and one year cropping on Ultisol in southern Cameroon were studied. Fallows were slashed and burned. The fallow system had no consistent effect on nutrient uptake by individual crops. Crop nutrient uptake was most often highest in the S. spectabilis system. Nitrogen balances were generally negative due to N loss in the burn. Across three cropping cycles, the balance of fallow nutrient uptake versus total crop nutrient uptake was only in the S. spectabilis system positive for all nutrients. Nutrient export by all crops (mean of three years) was unaffected by fallow systems. The fallow nutrient uptake versus crop nutrient export balance was positive for all nutrients and systems. Planted fallows appear capable of acquiring sufficient nutrient stocks during fallow phases, covering the crops’ demand. Fallow N and K uptake and crop export declined with every fallow/cropping cycle.
Introduction
In West and Central Africa, slash and burn agriculture is still a widely used form of crop production. Increasing population densities may render the system incapable to furnish sufficient food in the long term. Planted fallows, often called improved fallows, were tested to shorten fallow phases, to improve soil fertility and to attain increased crop yields. Alley cropping was one type of these systems (Hauser et al. Citation2006), developed for continuous food production to replace slash and burn bush fallow systems. On relatively fertile Alfisols it was shown to be a suitable system, permitting continuous cereal and grain legume production (Kang et al. Citation1995a). On less fertile Ultisols and Oxisols, however, hedgerows had negative effects on crop yields (Hauser et al. Citation2006). A large proportion of alley cropping trials were conducted with a very limited number of hedgerow species (Leucaena leucocephala and Gliricidia sepium) which appear not to be suitable under all soil conditions.
In large parts of the Congo basin and specifically in southern Cameroon an intercrop of groundnut, maize and cassava is the most common subsistence system in smallholder farming systems (Büttner and Hauser Citation2003). However, groundnut, a major source of protein for the local population, has not received much attention within planted fallow and alley cropping research. Schroth et al. (Citation1995) found increased groundnut yields in alley cropping in Cote d’Ivoire on an Acrisol and concluded that groundnut should be included in the set of crops tested in alley cropping trials. A review of the literature on tree-based fallows in West and Central Africa revealed that less than 10% of the comparisons with natural fallow were made in intercropping systems, none of which had more than two crops (Hauser et al. Citation2006). Smallholder agriculture, however, is dominantly intercropping in rather complex systems with three or more crops.
In continuous alley cropping systems, the prunings are a nutrient source to the crops, if crops tolerate a mulch layer. Under Cameroonian conditions the amounts of prunings were small, rendering virtually no nutrient input to crops (Hauser Citation2008b). This might have been the major reason for frequently insignificant crop yield responses in a long-term experiment, started in 1990 and continuously cropped to a cassava/maize intercrop for five years (Hulugalle and Ndi Citation1994; Hauser et al. Citation2000). When the system was changed to a rotational ‘1 year cropping/2 years fallow’ system, biomass production and nutrient accumulation differed strongly between hedgerow species (Hauser Citation2008a) but had little effect on crop yields (Hauser Citation2008b). The cassava/groundnut/maize system requires a clean soil surface and thus can only be implemented in a labour efficient manner by slash and burn. Burning natural fallow biomass has been shown at a nearby site to have detrimental effects on maize yields (Norgrove et al. Citation2003), yet no information is available on the effects of biomass burning on intercrops including groundnut and cassava. Although it is to be assumed that the nitrogen in the biomass is almost completely lost, the questions arise (1) if the nutrient uptake into the fallow biomass within 2 years is sufficient to meet the nutrient uptake of the crops such that the soil nutrient status may remain unchanged and (2) if the nutrient uptake into the fallow has an effect on the nutrient uptake of the following crops, their yields and the according nutrient export.
This study reports the total nutrient uptake of all three crops of a groundnut/maize/cassava intercrop at harvest and the exported amount of nutrients over three complete cycles of 2 years fallowing and 1 year cropping and balances these amounts against the nutrient uptake into the living and dead aboveground biomass of the previous 2 fallow years.
Materials and methods
The experiment was established at the research farm of the International Institute of Tropical Agriculture (IITA), Humid Forest Eco-regional Centre, Mbalmayo, southern Cameroon (3°27ʹ58.33” N, 11°29ʹ11.95” E) in 1990, on land manually cleared from secondary forest. Biomass was burned in situ and remaining wood was stacked and burned again. The soil is a clayey, kaolinitic, isohyperthermic, Typic Kandiudult (Hulugalle Citation1992). Average annual precipitation is 1513 mm with a bimodal distribution. Rains start in mid-March. A short dry season follows from mid-July until the end of August. The main rainy season is from September to the middle of November.
The experiment was a randomized complete block design, replicated three times, with three hedgerow species, Senna spectabilis, Flemingia macrophylla and Dactyladenia barteri versus a no-tree control, which consisted largely of weeds of the previous cropping phase and Chromolaena odorata. Hedgerows were established from seeds in May 1990. Cropping started in April 1991. Plots measured 28 m x 6 m and had five hedgerows planted at an interrow distance of 6 m and an intrarow distance of 0.25 m. Biomass was not burned during the first 5 years of continuous maize/cassava intercropping. Further details on the establishment of the trial and the yield of the maize/cassava intercrop from 1991 to 1992 are described by Hulugalle and Ndi (Citation1994), crop yields of 1993 to 1996 were reported by Hauser et al. (Citation2000). Volunteer regrowth and weed composition were reported by Hauser (Citation2002) and biomass production and nutrient uptake of the fallows by Hauser (Citation2008a).
Land preparation and crop planting
By the end of each 2 years fallow phase, the fallow vegetation was slashed with cutlasses, left in situ to dry and burned. All dates of field operations are in . After the burn, all remaining weeds and stumps of undesired species such as C. odorata were uprooted, piled and burned again. The ash was distributed in the interrow space between the hedgerows or uniformly in the no-tree control. The entire plot of the no-tree control and the interrow space of the planted fallows was hand hoed, and a local groundnut (Arachis hypogaea) variety was seeded at a density of approximately 20 m−2 by mixing them with the hoed soil. Maize (Zea mays) cv. CMS 8704 was seeded at 1 m distance parallel to the hedgerows. Distance between maize rows was 1 m and within rows was 0.5 m with three seeds per pocket (6 plants m−2). Cassava (Manihot esculenta) cv. 8017 was planted in alternating rows with the maize starting at 0.5 m from the hedgerow with 1 m distance between in within rows (1 plant m−2). Maize was thinned to two plants per pocket (4 plants m−2) at the five- to six-leaf stage. The same crop varieties were established with the same patterns in all years.
Table 1. Dates of field operations, figures in brackets are days after planting.
Pruning
During the 1998/1999 cropping cycle, the hedgerows of S. spectabilis were pruned 4 times, on 4 May, 8 June, 18 August and 15 October; hedgerows of F. macrophylla were pruned 2 times, on 4 May and 18 August 1998. D. barteri was not pruned at all. During the 2001/2002 cropping cycle, F. macrophylla was pruned on 16 April 2001, all species were pruned on 22 May 2001, S. spectabilis was pruned on 26 June 2001, all species were pruned on 23 October 2001 and 6 February 2002. During the 2004/2005 cropping cycle, all species were pruned on 27 May 2004, 9 December 2004 and 25 April 2005.
Biomass production and nutrient content of hedgerows accumulated during fallow are reported by Hauser (Citation2008a) and pruning amounts during cropping were reported by Hauser (Citation2008b). Hedgerow biomass was cut at about 0.3 to 0.4 m height and weighed fresh. A subsample of three branches per row was retained. All branches collected from one plot were shredded to about 2 cm long pieces, then bulked into one sample, dried at 65°C to constant mass and weighed to calculate dry matter content. Thereafter, samples were ground to 0.5 mm and analysed for their nutrient concentrations.
Weeding
There was little weed germination in the first four weeks after planting. On 25 May 1998, on 3 May 2001 and 18 May 2004 all plots were manually weeded. At groundnut harvest, about 3 months after seeding, the removal of the pods went along with a removal of weeds which were piled around the cassava stands. A third manual weeding was conducted in each cropping year at the end of the second rainy season during November.
Crop harvest
Groundnuts were harvested from a transect of 0.75 m wide and 18 m long, starting in the middle of the inter-hedgerow space between the first and second hedgerow, reaching across the three inner hedgerows and ending in the middle of the inter-hedgerow space before the fifth hedgerow. All groundnuts within the transect were removed, pods detached and the straw and the pods weighed separately. Pods and straw were dried at 65°C to constant mass. Dry pods were shelled and grain and shells weighed separately. Nutrient analysis was conducted on grains and on bulked straw plus empty pods.
Maize was harvested in all inter-hedgerow spaces in the centre 4 m of each row. Cobs were removed and husked. Sub-samples of cobs and straw were dried at 65°C to constant mass. The grain dry matter was determined by shelling the dried cobs and weighing the grain. Nutrient analysis was conducted on grain and straw plus husk samples bulked across all rows and inter-hedgerow spaces.
Cassava was harvested in all inter-hedgerow spaces in the centre 4 m of each row. The above ground material was cut at ground level and separated into stem suitable as planting material and small branches including leaves. The two types of aboveground material were weighed separately and a sub-sample was retained. The tuberous roots were removed from the soil, cleaned of attached soil and weighed. Sub-samples of each type of material were dried at 65°C to constant mass. Nutrient analysis was conducted on roots, stems and branches plus leaves.
Nutrient analyses
Plant samples were digested according to Novozamsky et al. (Citation1983). Total N was determined with an ammonium sensitive electrode (Powers et al. Citation1981), and total P content was determined by the malachite green colorimetric procedure (Motomizu et al. Citation1983). Cations were determined by atomic absorption spectrophotometry.
Nutrient balances
The nutrient balances were calculated based on the total N, P, K, Ca and Mg uptake into the planted fallow species, the natural regrowth between hedgerows and in the no-tree control and the litter found by the end of the 2-year fallow cycles. The determination of these amounts is described in Hauser (Citation2008a). The term fallow biomass nutrient stocks (FBNS) includes here nutrients in the litter, volunteers (weeds) and the planted fallow species. From these amounts were subtracted the total nutrient uptake into the aboveground biomass of maize, the aboveground and the (belowground) pod and grain biomass of groundnut and the aboveground and the belowground tuberous root biomass of cassava, determined at the final harvest of each crop, to obtain the total crop nutrient uptake (CNU) and to calculate the ‘fallow biomass nutrient stock – crop nutrient uptake’ balance (FBNS – CNU balance). The ‘fallow biomass nutrient stock – crop nutrient export’ (FBNS – CNE) balance was calculated by summing up the nutrient contents in maize grain, groundnut grain and in the tuberous roots of the cassava and subtracting the total from the amounts found in the fallow biomass. The export of nutrients with cassava stems to plant new fields was not considered as the required amounts are variable, yet relatively small (estimated at 10–20% of the total) if the same area is to be planted from the stems produced on that area.
Statistical analyses
Data were analysed without transformation in SAS release 9.2, using proc GLM for comparison of treatments. Means were separated using Tukey’s honest significant difference (HSD) test.
Results
Total nutrient uptake by crops
The fallow system had a significant effect on the nutrient uptake into crop parts in 14 cases out of the 105 comparisons possible between the four fallow systems, composed of seven different crop plant parts (cassava leaves, stems and roots; maize grain and straw and groundnut grain and straw) analysed for N, P, K, Ca, Mg in three cropping cycles. None of the plant parts had a consistent response to the fallow system. In 1998, three significant differences were found, all associated with the highest uptakes in the F. macrophylla system. In 2001, seven significant differences were found, one associated with highest nutrient uptake in the S. spectabilis system, three in the D. barteri system and 3 in the F. macrophylla system. In 2004, four significant differences were found, all associated with highest nutrient uptake in the S. spectabilis system. Calcium uptake was not affected by the fallow systems. Nitrogen was affected in 2 cases and P, K and Mg were each affected in 4 cases. Detailed data are in the supplemental materials Annex 1.
The fallow systems had no consistent effect on nutrient uptake by individual crops. Cassava N, Ca and Mg uptake was not affected by fallows. Cassava P uptake was highest in the S. spectabilis system (). Groundnut N uptake was unaffected by the fallows. Groundnut P and Ca uptake were highest in the D. barteri system. Maize nutrient uptake was unaffected by the fallows. The total nutrient uptake of the three crops across the three cropping cycles, except for Mg, was highest in the S. spectabilis system, yet significantly different only for P (). Across the three cycles, cassava took up the largest amounts of N, P, K, Ca and Mg, with 54.1, 10.5, 76.6, 37.5, and 15.3 kg ha−1, respectively. Groundnut uptake was 50.7, 3.9, 37.2, 20.0 and 7.4 kg ha−1 and maize uptake was 44.3, 8.5, 52.7, 10.9 and 8.4 kg ha−1 N, P, K, Ca and Mg, respectively.
Table 2. Average crop nutrient uptake (kg ha−1) per cropping cycle in a groundnut/cassava/maize intercrop over three cropping cycles on Ultisol, Mbalmayo, Southern Cameroon.
Over the three fallow-crop cycles the total K uptake into the three crops declined from 233.7 (1998) to 149.0 (2001) and 116.8 kg ha−1 in 2004. Uptake of P declined from 28.5 (1998) to 22.6 (2001) and 17.5 kg ha−1 in 2004. Nitrogen, calcium and magnesium uptake dropped from 1998 to 2001 and increased to 2004, yet not to the level of 1998.
Nutrient export by crops
The amount of nutrients exported by all three crops was unaffected by the fallows in each cropping cycle and across the three cropping years. Exports per cycle were 67.1 kg ha−1 N (SE = 1.6), 13 kg ha−1 P (SE = 0.7), 50.9 kg ha−1 K (SE = 1.1), 5.7 kg ha−1 Ca (SE = 0.15), and 6.5 kg ha−1 Mg (SE = 0.12). The N export was constant from cycle to cycles, while the K export declined from 66.0 (1998) to 55.9 (2001) and 30.8 kg ha−1 in 2004. Nutrient exports by individual crops varied greatly (data not shown), and were, except for N, largest with cassava roots. Export of P in maize grain (5.3 kg ha−1) was marginally lower than that in cassava roots (5.4 kg ha−1). Groundnut grains exported the largest amount of N (29.2 kg ha−1).
Nutrient uptake by the fallow biomass
The amounts of nutrients accumulated in the above ground fallow biomass differed significantly in all years and across the three fallow cycles except for Mg (). Detailed data on the fallow biomass nutrient uptake by the different systems and distribution between volunteers, litter and planted tree species are reported by Hauser (Citation2008a). Across the fallow systems, the uptake of N and K decreased consistently from 1998 to 2004, while P, Ca and Mg increased from 1998 to 2001 and then decreased slightly in 2004, yet not to the same level as in 1998. The changes of P, Mg and Ca uptake over the fallow phases followed the same pattern in all fallow systems, being lowest in 1998 highest in 2001 and dropping slightly in 2004 (). However, K uptake increased from 1998 to 2001 and remained the same in 2004 in the no-tree control, while it decreased from 1998 to 2001 and 2004 in the planted fallow systems. N uptake decreased consistently in the Senna system, yet peaked in 2001 in all other systems.
Table 3. Nutrient uptake (kg ha−1) into all fallow biomass in four different fallow systems over a 2 years fallow phase on Ultisol, Mbalmayo, Southern Cameroon.
Nutrient cycling through pruning application
The tree species were pruned at different frequencies and dates in each cropping cycle. Dactyladenia barteri did not produce any prunings in 1998 thus made no contribution to nutrient inputs. In the following years, the input remained low compared with the other tree species (). The cropping cycle was considered to have two phases, the early phase during which the maize and the groundnut were growing and a second phase after the maize and groundnut harvest (at around 100 DAP) during which only the cassava remained in the plots. Due to the generally low nutrient acquisition of D. barteri, the total nutrient input and the input during the early phase was negligible. Flemingia macrophylla contributed the highest proportion of nutrients from prunings in the early phase, compared with S. spectabilis and D. barteri.
Table 4. Average nutrient input (kg ha−1) through pruning application in the early phase (up to 66 DAP), total nutrient input through prunings and the percentage of total nutrients applied in the early phase over three cropping cycles on Ultisol, Mbalmayo, Southern Cameroon.
Fallow biomass nutrient stock – crop nutrient uptake balance
Due to the assumed complete loss of all N in the fallow biomass during the burn, the nitrogen balance between N in fallow biomass and the crop N uptake was generally negative, if the potential N-fixation of the groundnut and in the F. macrophylla prunings is not considered.
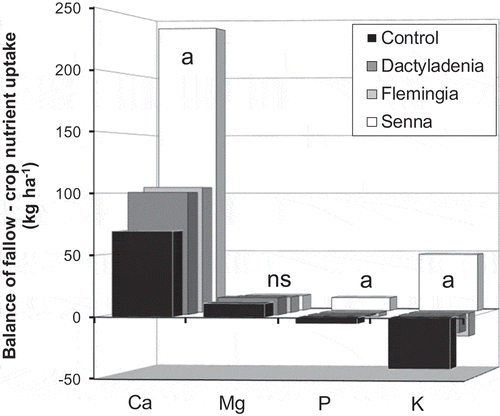
In the 1998 cropping cycle 10 of the 16 nutrient balances of fallow biomass nutrient stocks (FBNS) versus crop nutrient uptake (CNU) were negative, with P, K and Mg balances being negative in three of the fallow systems (). In the 2001 cropping cycle two balances were negative, both for K. In the 2004 cropping cycle all balances were positive. The FBNS versus CNU balance of Mg was unaffected by the fallows in each cycle and across the cycles (). The P and Ca FBNS versus CNU balance was significantly affected by the fallows in each cycle and across the three cycles (). The K FBNS versus CNU balance was significantly affected by the fallows in the 1998 and 2004 cycles and across the three cycles. The average balance across the three cycles () was positive for Ca and Mg in all fallows, with a Ca balance in the S. spectabilis system being significantly more positive than in all other fallows. The P and K balances were positive only in the S. spectabilis system and were significantly different from all other fallows.
Table 5. Fallow biomass nutrient stock versus the crop nutrient uptake balance (kg ha−1) by fallow system and year.
Figure 1. Average balance of fallow biomass nutrient stocks versus crop nutrient uptake per fallow/cropping cycle over three complete cycles. Mbalmayo, southern Cameroon. Senna columns labelled with the letter a are significantly different from all other fallow systems.
Fallow biomass nutrient stock – crop nutrient export balance
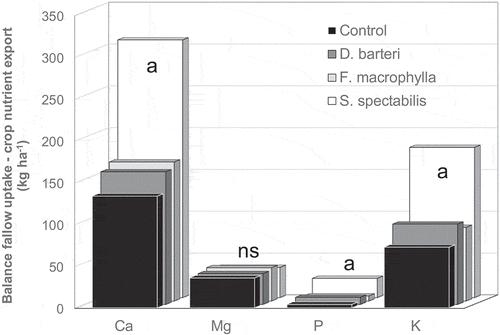
As for the N in fallow biomass versus the crop N uptake balance, the N in fallow biomass versus crop N export balance was generally negative. In 2001 and 2004 (not shown) and on average across the three fallow/cropping cycles the average balance of FBNS versus crop nutrient export (CNE) was in all systems and for all nutrients positive (). Only in 1998 was the FBNS – CNE balance of P in the no-tree control negative. Except for Mg, all FBNS – CNE balances were significantly higher in the S. spectabilis system than all other systems without differences between the other systems. Unlike for the FBNS – CNU balance, there was no increase in the FBNS – CNE balances over the cycles.
Figure 2. Average balance of fallow nutrient stock versus crop nutrient export per fallow/cropping cycle over three complete cycles. Mbalmayo, southern Cameroon. Senna columns labelled with the letter a are significantly different from all other fallow systems.
Correlations
Nutrient stocks in fallow aboveground biomass previous to cropping were not related to individual and total crop nutrient uptake.
Discussion
The concept of high fallow biomass production, accumulating large nutrient stocks, which could translate into high crop nutrient uptake and potentially high crop yields, could not be confirmed in this study. The low proportion of about 13% significant fallow system effects on the nutrient uptake into crop parts, the fact that no crop part experienced a consistent significant fallow system effect and the fact that fallow nutrient uptake was not correlated with crop nutrient uptake indicates that fallow nutrient uptake has no direct discernible effect on crop nutrient uptake. The fact that certain crops or crop parts had higher nutrient uptake after a specific fallow indicates that fallow systems may need to be selected for specific cropping systems, intercrops or individual crops. Identifying best fitting fallow species for specific crops may require the use of more specific criteria than high fallow biomass production and nutrient accumulation. The quality of the slashed material had strong effects on a maize crop in Tanzania (Kimaro et al. Citation2008) and in Malawi (Makumba and Akinnifesi Citation2008), yet only when slash was retained. Previous work in this trial has shown that high fallow biomass and nutrient uptake had little effects on crop yields (Hauser Citation2008b). There are no comparable data from other rotational planted tree fallow/cropping systems.
The failure of the fallow systems to cause a larger proportion and consistent significant effects on crop nutrient uptake may be related to the biomass management by burning, specifically for N. Burning biomass, however, is inevitable for the groundnut/maize/cassava intercrop, the most common field type in southern Cameroon (Büttner and Hauser Citation2003). For nutrients retained in the ash (P, K, Ca, Mg), a longer fallow phase or more fallow – crop cycles may be required to cause more frequent and consistent positive significant effects on crop nutrient uptake and potentially on crop yields. The shift of the number of significantly highest crop part nutrient uptakes from F. macrophylla in 1998 to S. spectabilis in 2004 may be an indication of the F. macrophylla system declining and the S. spectabilis system maintaining fallow nutrient accumulation or facilitating nutrient uptake by the crops through other mechanisms. The absence of cases of highest nutrient uptake in the no-tree control system, which accumulated the least nutrients, corroborates this assumption.
The most important aspect of the planted tree fallows appears to be the generally positive nutrient balances of fallow biomass nutrient stocks versus the nutrient export through the crops’ produce. Not considering N, as well as losses of the other nutrients through run-off, leaching and displacement as fly ash during the burn, it appears that the planted fallows, particularly S. spectabilis is capable to increase the nutrient stocks held in biomass during the fallow phase compared with the no-tree control. Except for N, all fallow species appear to acquire sufficient amounts of nutrients to balance the crops’ above ground plus cassava storage roots’ nutrient uptake.
While crops do not solely rely on nutrients supplied through the decomposing fallow biomass or ash from the burn, the balances drawn here indicate that the planted fallows are capable to accumulate and store in biomass the amounts of nutrients required by the crops. This means that these nutrients are actually acquired from the soil and kept in a form relatively safe from losses, until the fallow is cleared and the nutrients are released to the soil from the ash or lost (N) in the burn. For a legume, as the groundnut, which requires Ca in the topsoil to be taken up through the pod walls to form grains, the acquisition of Ca and enrichment of the topsoil should over a number of fallow – crop cycles lead to higher and more stable yields (Wolt and Adams Citation1979; Mandimba and Kilo Citation1997; Wendt Citation2002).
Nitrogen demanding crops such as the maize may over time face deficiencies due to N losses by burning. Over the three cropping cycles from 1998 to 2004 the maize grain yields declined, yet not consistently (Hauser Citation2008b). Potential advantages of an N-fixing fallow species such as F. macrophylla may become relevant over more cropping cycles. However, in a slash and burn system the N input from fixation potentially available to crops will be limited to the legume’s below ground plant parts and decomposition and to some extent to amounts provided through prunings. However, in this trial N supply from F. macrophylla prunings before the maize harvest was on average 19.6 kg ha−1 and cycle, thus a relatively small amount that needs to undergo the decomposition and transfer processes before it could benefit the maize. Banful et al. (Citation2008) determined F. macrophylla decomposing slower than Chromolaena odorata and Pueraria phaseoloides under southern Cameroonian conditions with about 65% of the N released from the leaves at 20 days after incubation. This reduces the N supply from F. macrophylla prunings to around 13 kg ha−1 at around 80 DAP, relatively late to have an effect on the maize yield. Henrot and Brussaard (Citation1997) found that 31% of the mass loss in F. macrophylla was attributed to soil macrofaunal activity, thus N contained in this portion would be removed from the decomposition process and become available later and through other pathways, likely to further reduce the amount of N available to the maize.
The potassium demanding cassava had consistently declining root yields over the three cropping cycles and highest yields in the F. macrophylla system. Although the K uptake of cassava was highest in the S. spectabilis system, this did not translate into highest yields. Either N was more deficient for cassava production or the S. spectabilis imposed too strong competition on the cassava, obliterating the advantage of the apparently better nutrient supply. The latter appears highly likely because the cassava had a strong negative spatial response to the proximity of the S. spectabilis hedgerows in 1998 and 2001 with yields lowest in the rows next to the hedgerows (Hauser Citation2008b).
The positive nutrient balances of the planted fallow systems need to be considered with caution. Due to the application of prunings and the different lengths of growing period of maize and groundnut compared with the cassava and due to the fact that crop residues were retained in the plots, the balances drawn by simple addition of all crops’ nutrient uptakes, may comprise a portion of nutrients that were cycled from prunings and from the early harvested crops to the cassava within the overall growing period. One pathway would have been the uptake of nutrients by the maize and groundnut from early phase prunings, of which a portion would have remained in the plots as crop residues, providing nutrients to the cassava. This would result in an overestimation of the total crop nutrient uptake, leading to more positive FBNS – CNU balances, yet would not affect the crop nutrient export data and FBNS – CNE balances.
The generally negative balances for N based on N exports against the complete loss of N from fallow biomass, would be less negative if N fixed by the groundnut is considered. However, to my knowledge, there are no data on N fixation by groundnut on the acid Ultisols of southern Cameroon. Estimates from the literature range from around 40 to 200 kg ha−1 fixed N (Toomsan et al. Citation1995; Okito et al. Citation2004), providing around 72–89% of the crop’s N uptake (Dakora et al. Citation1987). Due to the low biomass production of the groundnut and the low total N amounts in the crop (average across years and fallow systems of 50.7 kg ha−1) and assuming a proportion of N contributed by fixation of 72% the amounts of N added by fixation would have been 20.4, 37.6 and 29.5 kg ha−1 in 1998, 2001 and 2004, respectively. The N balances would need to be upward adjusted by these amounts, yet would remain negative.
Mackensen et al. (Citation1996) determined losses of 90% N, 30% P, 35% K, 20% Ca, and 30% Mg with the burn. Such losses vary with the intensity of the burn; thus, the fuel load and the weather conditions specifically wind speed during the burn. Wind deposition of P, K, Ca and Mg cannot be excluded but are unlikely to have been as high as reported by Mackensen et al. (Citation1996) because the fuel loads were small and the material was not completely dry thus the burn had a relatively low intensity. However, if the data of Mackensen et al. (Citation1996) are used as a worst-case scenario, more FBNS – CNU balances would be negative, whereby the K balances would be most severely affected leading to negative FBNS – CNU balances in all years and all fallow systems equivalent to an additional average loss of 56 kg ha−1 K per cycle across the fallow systems. The second most affected element would be P with negative FBNS – CNU balances in all years and systems except the S. spectabilis system in 2001 and 2004. Mg balances would be negative in all systems in 1998. Considering that K uptake by the fallow and crops was declining with time, it is to be assumed that K losses occurred either during the burn and/or after through leaching.
The more positive balances with time were largely caused by a decrease of the crop nutrient uptake and not through an increased fallow nutrient uptake. As such the systems may be in a general condition of declining nutrient stocks in the biomass. This appears specifically the case for K and P which had most frequently negative FBNS – CNU balances. The decline of the amounts of N in the fallow biomass indicates in addition a potential decline in the S. spectabilis system, due to the relatively large amounts of N taken up and lost in the burn. Senna spectabilis, although a legume, does not fix N, thus draws on soil N resources and through biomass burning, caused the largest N losses.
The decreasing K uptake into the fallow biomass of the planted hedgerow systems may have to be interpreted as an indicator of the systems moving towards a K deficiency situation. In the S. spectabilis system this would be aggravated by the concurrent decrease in the N uptake into the fallow biomass.
Conclusion
It appears that the (conceptual) functions of these agroforestry systems provide the services that could render these systems sustainable from a nutrient cycling and supply point of view within one fallow – cropping cycle. However, the negative trends in fallow biomass accumulation and crop nutrient uptake indicate that over time these systems may become nutrient deficient. The failure of systems with higher nutrient supply from fallow biomass to produce higher crop yields demands the development of management systems for the trees that would reduce or eliminate competition during the cropping phase, ensure that no other negative effects of the trees affect the crop yields (allelopathic substances), yet allow rapid recovery and high nutrient uptake in the fallow phase. Reasons for declining fallow nutrient uptake require further investigations into the pathways of nutrient losses and methods to reduce or prevent them. Alternatively, nutrient losses might be compensated for by fertilizer application or addition of other nutrient sources while maximizing the cycling.
GAGS-2019-0363-File003.xlsx
Download MS Excel (27.7 KB)Supplemental data
Supplemental data for this article can be accessed here.
Disclosure statement
No potential conflict of interest was reported by the author.
Related Research Data
References
- Banful B, Ofori K, Kumaga F, Hauser S, Ndango R. 2008. Decomposition and nutrient release patterns of Pueraria phaseoloides, Flemingia macrophylla and Chromolaena odorata leaf residues in tropical land use systems. Ghana J Agric Sci. 41:11–22.
- Büttner U, Hauser S. 2003. Farmers’ nutrient management practices in indigenous cropping systems in southern Cameroon. Agr Ecosyst Environ. 100:103–110. doi:10.1016/S0167-8809(03)00178-6.
- Dakora FD, Aboyinga RA, Mahama Y, Apaseku J. 1987. Assessment of N2 fixation in groundnut (Arachis hypogaea L.) and cowpea (Vigna unguiculata L. Walp) and their relative N contribution to a succeeding maize crop in Northern Ghana. MIRCEN J Appl Microb. 3(4):389–399. doi:10.1007/BF00935697.
- Hauser S. 2002. Production and spatial distribution of volunteer biomass between multipurpose tree hedgerows after two years of fallow. Agroforestry Systems. 55:139–147. doi:10.1023/A:1020586711333.
- Hauser S. 2008a. Biomass production, nutrient uptake and partitioning in planted Senna spectabilis, Flemingia macrophylla, and Dactyladenia barteri fallow systems over three fallow/cropping cycles on Ultisol. Arch Agron Soil Sci. 54:423–438. doi:10.1080/03650340802070919.
- Hauser S. 2008b. Groundnut/cassava/maize intercrop yields over three cycles of planted tree fallow/crop rotation on Ultisol in southern Cameroon. Biol Agric Hortic. 25:379–399. doi:10.1080/01448765.2008.9755063.
- Hauser S, Ndi JN, Hulugalle NR. 2000. Yields of maize/cassava intercrops grown with hedgerows of three multipurpose trees on an acid Ultisol of Cameroon. Agroforestry Systems. 49:111–122. doi:10.1023/A:1006351025490.
- Hauser S, Nolte C, Carsky RJ. 2006. What role can planted fallows play in humid and sub-humid West Africa? Nutr Cycl Agroecosys. 76:297–318. doi:10.1007/s10705-005-5630-4.
- Henrot J, Brussaard L. 1997. Determinants of Flemingia congesta and Dactyladenia barteri mulch decomposition in alley-cropping systems in the humid tropics. Plant Soil. 191:101–107. doi:10.1023/A:1004274115587.
- Hulugalle NR. 1992. Contributory factors to soil spatial variability in an Ultisol I. burning vegetation residues in heaps during land clearing. Commun Soil Sci Plant Anal. 23:1859–1869. doi:10.1080/00103629209368709.
- Hulugalle NR, Ndi JN. 1994. Changes in soil properties of a newly-cleared Ultisol due to establishment of hedgerow species in alley cropping systems. J Agr Sci-Cambridge. 122:435–443. doi:10.1017/S002185960006737X.
- Kang BT, Hauser S, Vanlauwe B, Sanginga N, Atta-Krah AN. 1995. Alley farming research on high base status soils. In: Kang BT, Osiname OA, Larbi A, editors. Alley farming research and development. Ibadan: Alley Farming Network for Tropical Africa; p. 25–39.
- Kimaro AA, Timmer VR, Chamshama SAO, Mugasha AG, Kimaro DA. 2008. Differential response to tree fallows in rotational woodlot systems in semi-arid Tanzania: post-fallow maize yield, nutrient uptake, and soil nutrients. Agr Ecosyst Environ. 125:73–83. doi:10.1016/j.agee.2007.11.007.
- Mackensen J, Hoelscher D, Klinge R, Foelster H. 1996. Nutrient transfer to the atmosphere by burning of debris in eastern Amazonia. Forest Ecol Manag. 86:121–128. doi:10.1016/S0378-1127(96)03790-5.
- Makumba W, Akinnifesi FK. 2008. Mineralization and N-use efficiency of tree legume prunings from fertilizer tree systems and low quality crop residues in Malawi. Afr J Biotechnol. 7:3266–3274.
- Mandimba G, Kilo C. 1997. Effect of liming on the performance of groundnut grown on acid soil in Congo. International Arachis Newsletter. 17:66–67.
- Motomizu S, Wakimoto P, Toei K. 1983. Spectrophotometric determination of phosphate in river waters with molybdate and malachite green. Analyst (London). 108:361–367. doi:10.1039/AN9830800361.
- Norgrove L, Nkem JN, Hauser S. 2003. Separating the agronomic effects of burning and biomass removal in a typical slash and burn rotation system in southern Cameroon. Biol Agric Hort. 21:367–381. doi:10.1080/01448765.2003.9755279.
- Novozamsky I, Houba VJG, van Eck R, van Vark W. 1983. A novel digestion technique for multi-element plant analysis. Commun Soil Sci Plant Anal.. 14:239–248. doi:10.1080/00103628309367359.
- Okito A, Rodrigues Alves BJ, Urquiaga S, Boddey RM. 2004. Nitrogen fixation by groundnut and velvet bean and residual benefit to a subsequent maize crop. Pesq Agropec Bras. 39(12):1183–1190. doi:10.1590/S0100-204X2004001200004.
- Powers RF, Van Gent D, Townsend RF. 1981. Ammonia electrode analysis of nitrogen in microKjeldahl digests of forest vegetation. Commun Soil Sci Plant Anal. 12:19–30. doi:10.1080/00103628109367123.
- Schroth G, Balle P, Peltier R. 1995. Alley cropping groundnut with Gliricidia sepium in Cote d’Ivoire: effects on yields, microclimate and crop diseases. Agroforestry Systems. 29:147–163. doi:10.1007/BF00704883.
- Toomsan B, McDonagh JF, Limpinuntana V, Giller KE. 1995. Nitrogen fixation by groundnut and soyabean and residual nitrogen benefits to rice in farmers’ fields in Northeast Thailand. Plant Soil. 175:45–56. doi:10.1007/BF02413009.
- Wendt JW. 2002. Groundnut response to ash, phosphorus, lime and tillage in southern Cameroon. Biol Agric Hortic. 20:187–199. doi:10.1080/01448765.2002.9754964.
- Wolt JD, Adams F. 1979. Critical levels of soil and nutrient solution calcium for vegetative growth and fruit development in Florunner peanuts. Soil Sci Soc Am J. 43:1159–1165. doi:10.2136/sssaj1979.03615995004300060020x.