
Abstract
A new species of the genus Zetorchella (Oribatida: Caloppiidae) is described from soil of Côte d’Ivoire. Zetorchella cotedivoirensis Ermilov n. sp. differs from Zetorchella deleoni (Higgins, Citation1966) by the presence of phylliform interlamellar and dorsal notogastral setae and nine pairs of notogastral setae. A revised generic diagnosis and an identification key to species of Zetorchella from the Afrotropical region are presented.
http://zoobank.org/urn:lsid:zoobank.org:pub:677FAD4B-781F-4AE1-96B0-1D41428A2BE2
Introduction
The oribatid mite genus Zetorchella (Acari: Oribatida) of the family Caloppiidae was proposed by Berlese (Citation1916) with Chaunoproctus cancellatus Pearce, Citation1906 as type species. According to Subías’s catalogue (2004, updated version 2018), it comprises 23 species, which collectively are distributed in the Afrotropical and Oriental regions.
Among the material collected from soil of Côte d’Ivoire, we found a new species of Zetorchella. The main goals of the article are: (a) to describe and illustrate the new species, (b) to propose a revised generic diagnosis, (c) to give an identification key to species of Zetorchella from the Afrotropical region.
Material and methods
The study was conducted in 2007 in primary forest located in Oumé region of mid-West Côte d’Ivoire, precisely in the village of Goulikao. The climate is subequatorial with bimodal rainfall patterns (March to June and September to October). The vegetation is very dense and even luxuriant. Man-made activities are very weak and limited to some tracks. The undergrowth is also dense with lianas and dead wood. Ferralitic soils are regularly distributed in the forest. Soil cores were collected in June 2007 (rainy season) with a steel corer (5 cm diameter) at two extreme layers (0–5 cm and 35–40 cm). Soil mites were extracted by using the Berlese–Tullgren funnels. Extraction lasted 10 days. The detailed collection locality and habitat for the new species is given in the “Material examined” section.
Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the ventral plate. Notogastral width refers to the maximum width of the notogaster. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulas for leg setation are given in parentheses according to the sequence trochanter–femur–genu–tibia–tarsus (famulus included). Formulas for leg solenidia are given in square brackets according to the sequence genu–tibia–tarsus.
Drawings were made with a camera lucida using a Leica transmission light microscope “Leica DM 2500”.
Morphological terminology used in this article follows that of F. Grandjean: see Travé and Vachon (Citation1975) for references, Norton (Citation1977) for leg setal nomenclature, and Norton & Behan–Pelletier (Citation2009), for overview.
The following abbreviations are used: lam – lamella; tlam – translamella; slam – sublamella; Al – sublamellar porose area; tu – tutorium; ro, le, in, bs, ex – rostral, lamellar, interlamellar, bothridial and exobothridial setae, respectively; bo – bothridium; hr – humeral ridge; D – dorsophragma; P – pleurophragma; c, la, lm, lp, h, p – notogastral setae; Aa, A1, A2, A3, A4 – notogastral porose areas; ia, im, ip, ih, ips – notogastral lyrifissures; gla – opisthonotal gland opening; cs – circumgastric scissure; csb – circumgastric sigillar band; a, m, h – subcapitular setae; or – adoral seta; v, l, d, cm, acm, ul, sul, vt, lt – palp setae; ω – palp and leg solenidion; cha, chb – cheliceral setae; Tg – Trägårdh’s organ; Pd I, Pd II – pedotecta I, II, respectively; 1a, 1b, 1c, 2a, 3a, 3b, 3c, 4a, 4b, 4c – epimeral setae; dis – discidium; cp – circumpedal carina; g, ag, an, ad – genital, aggenital, anal and adanal setae, respectively; iad – adanal lyrifissure; p.o. – preanal organ; p.a. – leg porose area; σ, φ – leg solenidia; ɛ – leg famulus; v, ev, bv, l, d, ft, tc, it, p, u, a, s, pv, pl – leg setae.
Systematics
Family Caloppiidae Balogh, Citation1960
Genus Zetorchella Berlese, Citation1916
Type species: Chaunoproctus cancellatus Pearce, Citation1906
Revised generic diagnosis
Adult
Prodorsum. Rostrum rounded. Lamellae of medium length, broad, without cusps. Translamella, sublamellae and sublamellar porose areas present. Tutoria long, ridge-like. Rostral, lamellar and interlamellar setae well-developed: ro setiform, inserted laterally on prodorsum close to distal ends of tutoria; le setiform, inserted on lamellae ends; in setiform, bacilliform or phylliform, located in interlamellar region. Bothridial setae of medium size to short, clavate, rarely bacilliform. Bothridia cup-like. Exobothridial setae setiform, thin. Surface of anterior part of prodorsum usually with reticulate ornamentation. Notogaster. Anterior margin distinct or not developed medially, posterior margin rounded. Humeral regions often with quadrangular processes and strong ridges. With five pairs of very small porose areas, Aa represented by one pair. Ten [exception: nine, p3 absent] pairs of notogastral setae well-developed, setiform, bacilliform or phylliform. Surface of notogaster usually densely foveolate. Gnathosoma. Subcapitular setae setiform. Palps with setation 0–2–1–3–9(+ω). Solenidion of palp tarsi bacilliform, attached to eupathidium. Chelicerae chelate-dentate, with two setiform setae. Lateral podosomal and epimeral regions. Pedotecta I and II represented by small laminae. Discidia and circumpedal carinae present. Borders of sejugal apodemes fused medially with each other and anterior margin of genital aperture. Epimeral setal formula: 3–1–3–3. Anogenital region. Six pairs of genital, one pair of aggenital, two pairs of anal and three [exception: two] pairs of adanal setae short, setiform. Adanal lyrifissures located close and lateral to anal plates. Marginal porose area absent or present. Surface of region usually densely foveolate. Legs. Tridactylous. Dorsoparaxial porose areas on femora I–IV and on trochanters III, IV and ventral porose areas in basal parts of tarsi and distal parts of tibiae well visible.
Juvenile instars
Not known.
Zetorchella cotedivoirensis Ermilov n. sp.
Diagnosis
Body size: 365–415 × 249–298. Notogaster covered with tuberculate cerotegument; dense foveolate ornamentation absent. Rostral and lamellar setae setiform, barbed; interlamellar setae shorter than ro and le, phylliform, barbed. Bothridial setae clavate, barbed. Nine pairs of notogastral setae present: p2 short, slightly thickened, barbed; other setae of medium size, phylliform, barbed; p3 absent. Epimeral and anogenital setae setiform, thin, slightly barbed. Two pairs of adanal setae developed. Marginal porose area absent. Leg tarsi I with 19 setae, l” absent.
Description
Measurements. Body length: 415 (holotype: ♀), 365–415 (9 paratypes: 4♀♀, 5♂♂); notogaster width: 298 (holotype: ♀), 249–298 (9 paratypes: 4♀♀, 5♂♂). No difference between females and males in body size.
Integument (). Body color brown. Body surface densely microfoveolate (visible under high magnification) and microgranulate (granules rounded or elongated, diameter and length up to 1). Anterior part of prodorsum reticulate. Notogaster covered with tuberculate cerotegument (diameter of tubercles up to 6) forming specific ornamentation (four longitudinal bands); dense foveolate ornamentation absent. Posterior parts of notogaster and ventral plate with strong sinuous ridges forming partially unclear polygonal pattern.
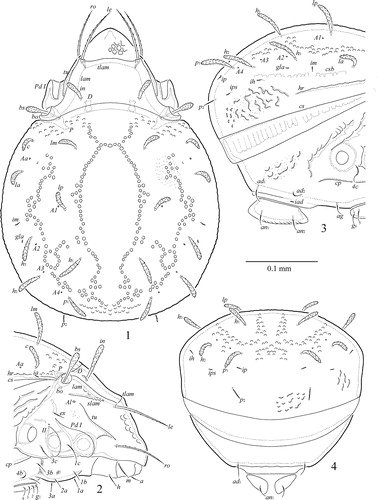
Figures 1–4 Zetorchella cotedivoirensis Ermilov n. sp. Adult: 1, dorsal view; 2, anterior part of body (legs I–III not illustrated), lateral view; 3, posterior part of body (leg IV not illustrated), lateral view; 4, posterior view.
Figures 5–11 Zetorchella cotedivoirensis Ermilov n. sp. Adult: 5, ventral view (gnathosoma and legs not illustrated); 6, subcapitulum, ventral view; 7, subcapitular lip, right, ventral view; 8, palp, right, antiaxial view; 9, chelicera, left, paraxial view; 10, leg I, without trochanter, left, paraxial view; 11, leg IV, left, antiaxial view.
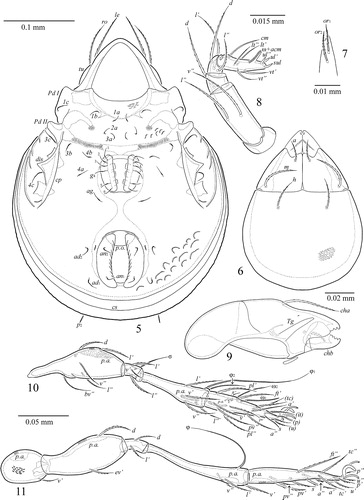
Prodorsum ( and ). Rostrum broadly rounded. Lamellae slightly shorter than half of prodorsum (measured in lateral view). Translamella complete, thick, straight. Sublamellae thin, distinctly shorter than lamellae. Sublamellar porose areas rounded (4). Tutoria as long as lamellae, ridge-like with one short additional ventral ridge. Rostral (73–82) and lamellar (82–86) setae setiform, barbed, ro slightly thinner than le. Interlamellar setae (53–65) phylliform, barbed. Bothridial setae (41–53) clavate, barbed, stalks and heads similar in length. Exobothridial setae (12–16) setiform, thin, barbed.
Notogaster (). Anterior margin present, but slightly visible medially. Humeral processes not developed. Humeral ridges strong. Nine pairs of notogastral setae present: p2 (14–16) setiform, slightly thickened, barbed; other setae (45–49) phylliform, barbed; p3 and their alveoli absent. Five pairs of porose areas (2) poorly visible; Aa located between insertions of setae c and la, A1 close to lp, A2 close to h3, A3 close h2, A4 close to p1. All lyrifissures distinct; ia located lateral to insertions of setae c, im anterior to h3, ip between p1 and p2, ih and ips in lateral position, anterior to p2. Opisthonotal gland openings located lateral to h3. Circumgastric scissure and circumgastric sigillar poorly visible.
Gnathosoma (). Subcapitulum longer than wide (102–106 × 82–86). Subcapitular setae setiform, barbed; m (28–32) longer and thicker than h (20) and a (16). Adoral setae (12) setiform, barbed. Palps (length 69–73) with typical setation 0–2–1–3–9(+ω). Postpalpal setae (6) spiniform, smooth. Chelicerae (length 102–106) with two setiform, barbed setae, cha (41) longer than chb (24). Trägårdh’s organ of chelicerae elongate triangular.
Epimeral and lateral podosomal regions (). Epimeral setae setiform, thin, slightly barbed; 1c, 3c, 4c (28–32) longer than others (14–16). Pedotecta II trapezoid in ventral view. Discidia triangular. Circumpedal carinae long, directed to pedotecta II.
Anogenital region (). Six pairs of genital, one pair of aggenital, two pairs of anal and two pairs of adanal setae, similar in length (14–16), setiform, thin, slightly barbed. Adanal lyrifissures located close and parallel to anal plates. Marginal porose area absent. Ovipositor is typical for Caloppiidae (but with coronal setae; e.g., Ermilov & Liao Citation2017), elongated (196 × 65), blades (65) shorter than length of distal section (beyond middle fold; 131). Each of the three blades with four smooth setae, ψ1 ≈ τ1 (36) setiform, ψ2 ≈ τa ≈ τb ≈ τc (16) thorn-like. Six coronal setae spiniform (4).
Legs ( and ). Morphology of leg segments, setae and solenidia generally typical for species of Zetorchella (e.g., Ermilov et al. Citation2010). Median claw thicker than laterals, all serrate on dorsal side; lateral claws each with small tooth ventrodistally. Dorsoparaxial porose areas on femora I–IV and on trochanters III, IV and ventral porose areas in basal parts of tarsi and distal parts of tibiae well visible. Formulas of leg setation and solenidia: I (1–5–3–4–19) [1–2–2], II (1–5–2–4–15) [1–1–2], III (2–3–1–3–15) [1–1–0], IV (1–2–2–3–12) [0–1–0]; homology of setae and solenidia indicated in .
Table 1 Leg setation and solenidia of adult Zetorchella cotedivoirensis Ermilov n. sp. [Roman letters refer to normal setae, Greek letters to solenidia (except ɛ = famulus). Single prime (′) marks setae on the anterior and double prime (″) setae on the posterior side of a given leg segment. Parentheses refer to a pair of setae].
Type material examined
Holotype (♀) and 9 paratypes (4♀♀, 5♂♂): Côte d'Ivoire, Oumé region, precisely in the village of Goulikao, 6°31’N, 5°30’W, 200 m a.s.l., primary forest, ferralitic soil, June 2007 (rainy season), collected by Julien K.N’Dri.
Depository
The holotype (♀; in ethanol with drop of glycerol) and 2 paratypes (1♀, 1♂; in ethanol with drop of glycerol): Senckenberg Institute, Görlitz, Germany; 7 paratypes (3♀♀, 4♂♂; in ethanol with drop of glycerol): Tyumen State University Museum of Zoology, Tyumen, Russia.
Etymology
The specific name cotedivoirensis refers to Côte d'Ivoire, where the holotype and paratypes were collected.
Remarks
Zetorchella cotedivoirensis Ermilov n. sp. is morphologically most similar to Z. deleoni (Higgins, Citation1966) from Guyana in having tuberculate ornamentation on the notogaster, but differs by the presence of phylliform interlamellar and dorsal notogastral setae (vs. bacilliform) and nine pairs of notogastral setae, p3 absent (vs. ten pairs, p3 present).
Key to known species of Zetorchella from the Afrotropical region
Notogaster without foveolae, but covered by the tuberculate cerotegument forming specific ornamentation; nine pairs of notogastral setae present (p3 absent); body size: 365–415 × 249–298Zetorchella cotedivoirensis Ermilov n. sp. Distribution: Côte d'Ivoire.
– Notogaster with dense foveolae and without tuberculate cerotegument; ten pairs of notogastral setae present (p3 present)2
Interlamellar and dorsal notogastral setae phylliform3
– Interlamellar and dorsal notogastral setae bacilliform or rod-like4
Bothridial setae shorter than dorsal notogastral setae, stalk shorter than head; notogastral setae p2 and p3 shorter and narrower than others; body size: 397–494 × 276–350Zetorchella rugosa (Mahunka, Citation1992). Distribution: Senegal.
– Bothridial and dorsal notogastral setae similar in length, stalk not shorter than head; notogastral setae p2 and p3 not shorter and not narrower than others; body size: 408–520 × 327–400Zetorchella pedestris Berlese, Citation1916 (=Chaunoproctus crinitus Karppinen, Citation1966; =Caloppia papillata Balogh, Citation1958) (see also Balogh Citation1960; Mahunka Citation1991). Distribution: Afrotropical region.
Bothridial setae bacilliform; body size: 785–830 × 585–615Zetorchella longipilosa (Mahunka, Citation1973). Distribution: Zimbabwe.
– Bothridial setae clavate5
Stalk of bothridial setae shorter than head; notogastral setae p2 and p3 bacilliform; body size: 394 × 296Zetorchella semirugosa (Mahunka, Citation2011). Distribution: Madagascar.
– Stalk of bothridial setae longer than head; notogastral setae p2 and p3 setiform6
Notogastral setae c longer than bothridial setae; body size: 837 × 610 Zetorchella vargai (Balogh, Citation1959). Distribution: Afrotropical region.
– Notogastral setae c shorter than bothridial setae 7
Notogastral setae c slightly shorter than bothridial setae and distinctly longer than notogastral setae p2 and p3; body size: 713–813 × 514–564Zetorchella nortoni Ermilov, Sidorchuk & Rybalov, Citation2010. Distribution: Ethiopia.
– Notogastral setae c distinctly shorter than bothridial setae and similar in length to notogastral setae p2 and p38
Anterior notogastral margin developed; notogastral setae rod-like; body size: 590–600 × 430–470Zetorchella basilewskyi (Balogh, Citation1958) (see also Balogh, Citation1960).
Distribution: Congo.
– Anterior notogastral margin not developed; notogastral setae bacilliform; body size: 465 × 320Zetorchella minor (Balogh, Citation1958) (see also Balogh, Citation1960). Distribution: Congo.
Acknowledgements
We cordially thank two anonymous reviewers for the valuable comments. Moreover, we are grateful to the farmers from Goulikao for allowing the study in their fields and the field assistance of technicians during the work.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Balogh J. 1958. Oribatides nouvelles de l'Afrique tropicale. Revue de Zoologie et de Botanique Africaines. 58(1/2):1–34.
- Balogh J. 1959. Some oribatid mites from Eastern Africa (Acari: Oribatidae). Acta Zoologica Academiae Scientiarum Hungaricae. 5(1/2):13–23.
- Balogh J. 1960. Descriptions complémentaires d’Oribates (Acari) d’Angola et du Congo Belge (Ière série). Companhia de Diamantes de Angola, Lisboa. 51:89–105.
- Berlese A. 1916. Centuria prima di Acari nuovi. Redia. 12:19–67.
- Ermilov SG, Liao J-R. 2017. Contribution to the knowledge of the oribatid mite genus Brassiella (Acari: Oribatida: Caloppiidae). Biologia. 72(9):1041–1048.
- Ermilov SG, Sidorchuk EA, Rybalov LB. (2010) Zetorchella nortoni, a new species of oribatid mite from Ethiopia (Acari: Oribatida: Caloppiidae). Acarina. 18(1):61–65.
- Higgins HG. 1966. Two new South American mites (Acari: Oribatei). Proceedings of the Biological Society of Washington. 79:13–16.
- Karppinen E. 1966. Some new oribatids (Acar., Oribatei) from Central Africa. Annales Entomologicae Fennicae. 32(4):275–281.
- Mahunka S. 1973. Neue und interessante Milben aus dem Genfer Museum XI. Neue und wenig bekannte Oribatiden aus Rhodesien (Acari). Archives des Sciences. 26(3):205–225.
- Mahunka S. 1991. Notes, additions and redescriptions of the oribatid species of Berlese (Acari). Acta Zoologica Hungarica. 37(1–2):27–58.
- Mahunka S. 1992. New and interesting mites from the Geneva Museum LXIII. A survey of the oribatid fauna of Senegal (Acari: Oribatida). Revue suisse de Zoologie. 99(3):673–712.
- Mahunka S. 2011. New and little known oribatid mites from Madagascar (Acari: Oribatida), III. Opuscula Zoologica Budapest. 42(1):43–66.
- Norton RA. 1977. A review of F. Grandjean’s system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal DL, editor. Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry; p. 33–61.
- Norton RA, Behan-Pelletier VM. 2009. Oribatida. Chapter 15. In: Krantz GW, Walter DE, editors. A manual of acarology. Lubbock: Texas Tech University Press; p. 430–564.
- Pearce NDF. 1906. VII – On some Oribatidae from the Sikkim Himalaya. Journal of the Royal Microscopical Society. 5:269–273.
- Subías LS. 2004. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles). Graellsia. 60 (número extraordinario):3–305. Online version accessed in January 2018, 605 pp.; http://escalera.bio.ucm.es/usuarios/bba/cont/docs/RO_1.pdf
- Travé J, Vachon M. 1975. François Grandjean. 1882–1975 (Notice biographique et bibliographique). Acarologia. 17(1):1–19.