
Abstract
This review presents a summary of the influences of floods on river ecology, both instream and on the adjacent floodplain, mostly in a Canadian context. It emphasizes that ecological impacts and benefits can be highly dependent on flood-generation processes and their magnitude and timing. In Canada, floods can occur under open-water or ice-influenced river conditions. The ecological impacts of floods generated from ice jamming are particularly relevant in Canadian ecosystems due to the potentially higher water levels produced and suspended sediment concentrations that can be detrimental to instream aquatic habitat, but beneficial to floodplains. Large floods provide a major source of physical disturbance. Moderate floods with shorter return periods can be beneficial to aquatic habitats by providing woody debris that contributes to habitat complexity and diversity, by flushing fine sediments and by providing important food sources from terrestrial origins. Floods also influence water-quality variables such as sediment loads, metals and pH, which further influence river ecology. This review points out important links between floods and habitat connectivity and refugia, and the ability of aquatic resources to recover from flood disturbances. The floodplain vegetation also influences bank erosion and channel configuration, as well as many processes contributing to the river ecology, such as allochthonous inputs of carbon, preventing bank erosion, food-web dynamics and other effects. Many species of riparian trees are dependent on floods for reproduction, and floods can limit competitive encroachment of upland vegetation. Floods play a critical role in deltaic environments where high flows provide nutrient supplies and overflows maintain water balances within these environments. Two substantial factors that will likely influence future flood regimes in Canada are climate change and flow regulation. Future research should focus on enhancing our understanding of how floods affect river ecology, including under winter conditions, to better manage important natural resources.
Cette revue présente un sommaire des impacts des crues sur l’écologie des rivières, tant au niveau du cours d’eau que dans la plaine inondable, et ce dans un contexte canadien. L’emphase est mise sur le fait que les impacts négatifs et positifs des crues dépendent des processus qui les génèrent, de même que de l’amplitude et de l’occurrence de ces événements. Au Canada, les crues peuvent se produire en eau libre ou en présence de glace. Les impacts écologiques des crues générées par des embâcles sont particulièrement importants et leur compréhension dans un contexte écologique est pertinente, étant donnés les hauts niveaux d’eau et les concentrations de sédiments élevées produits durant de tels événements. Ces conditions peuvent êtres nuisibles aux habitats aquatiques, mais bénéfiques pour les plaines inondables. Les grandes crues sont une cause primordiale de la perturbation du milieu physique. Les crues modérées, avec des périodes de retour plus petites, peuvent être bénéfiques aux habitats aquatiques puisqu’elles peuvent amener des débris d’arbres dans le cours d’eau, ce qui contribue à la diversité et la complexité des habitats. De plus, ce type de crue permet d’évacuer des sédiments fins et peuvent faire une contribution importante de nourriture d’origine terrestre. Les crues influencent aussi la qualité de l’eau, incluant des variables comme les charges sédimentaires, les métaux, le pH, qui modulent l’écologie lotique. Cette revue identifie les liens entre les crues et la connectivité des habitats et des refuges, de même que la capacité des ressources aquatiques à se rétablir suite aux perturbations associées aux débits extrêmes. La forêt située dans la plaine inondable joue un rôle dans la morphométrie du chenal et dans plusieurs processus qui contribuent à l’écologie de la rivière, tels que les intrants allochtones de carbone, la minimisation de l’érosion des berges, la dynamique de la chaîne trophique, etc. Plusieurs essences d’arbres de la zone riparienne dépendent des crues pour leur reproduction et ces mêmes crues peuvent limiter la propagation d’autres végétaux qui poussent habituellement sur les terres plus hautes. Les crues jouent un rôle critique dans les environnements des deltas, aux embouchures de certains cours d’eau. Dans ces milieux, les crues et les débordements sont des éléments qui font en sorte que l’équilibre hydrologique est maintenu et que les apports en nutriments sont suffisants. Deux facteurs majeurs vont fort probablement influencer le régime des crues au Canada: les changements climatiques et la régularisation des débits par la présence de barrage. Dans l’avenir, les chercheurs devraient se pencher sur l’amélioration de notre compréhension des aspects écologiques des crues, surtout durant les embâcles, afin de mieux informer les gestionnaires et les intervenants en conservation des ressources naturelles.
Introduction
Flood dynamics in Canada are governed by several hydrometeorological factors that include rainfall events, snowmelt and a combination of rain on snow that generally results in high water levels (Watt Citation1990). Floods can also be associated with river ice-breakup processes to produce extremely high waters. As such, floods can have a significantly different effect on the environment when generated under open-water versus ice-jamming conditions (Prowse and Culp Citation2003). An in-depth review of physical processes and regional aspects of floods in Canada is also presented in Buttle et al. (this issue).
Streamflow has a key role in driving the structure and function of riparian and aquatic communities, including connectivity to floodplain habitats (Hynes Citation1970). This is especially evident during extreme events where flood and low flows are often identified as the primary sources of ecological disturbance (Poff and Ward Citation1989). Accordingly, floods have played a major role in the structuring of stream ecosystems by modifying many physical, chemical and biological processes in rivers across Canada. Flood processes have direct and indirect effects on aquatic organisms (i.e. fish and invertebrates) and riverine vegetation as a result of erosion, sedimentation, bedload movement, etc. (Waters Citation1995). Seasonal flooding events bring important nutrients, carbon and material to the stream channel (e.g. woody debris and additional allochthonous material), essential in the overall function of riverine ecosystems. Occasional floods are equally important to the maintenance of a diverse and vibrant riparian ecosystem (Tabacchi et al. Citation1998).
However, floods have been observed to be detrimental in some cases (e.g. survival of juvenile fishes, invertebrates, etc.). For instance, a study by Cunjak et al. (Citation1998) demonstrated that mid-winter floods can adversely impact the survival of juvenile Atlantic salmon. A study by Milner et al. (Citation2013) showed that a major flood significantly impacted pink salmon populations of Wolf Point Creek (Alaska). For example, the number of spawners dropped from over 14,000 (pre-flood) to less than 500 (post-flood), and juveniles were equally affected. That study also showed that both spawners and juveniles recovered to pre-flood densities and returns within two generations (i.e. 4 years). Fausch et al. (Citation2001) showed that the timing of floods and the emergence of rainbow trout had a significant influence on the invasion success of some fish species. For instance, in regions where the timing of floods did not coincide with the native fish emergence, invasion success was high, whereas in regions where the floods coincided with the emergence of the native fish, invasion success was low (Fausch et al. Citation2001). Conversely, Peters et al. (Citation2006) demonstrated the importance of ice-jam flooding to recharging desiccated wetland basins of the Peace–Athabasca Delta (Alberta), a system that is influenced by climate and flow regulation effects. Additionally, Cordes et al. (Citation1997) used dendrochronological data to show that poplar and cotton wood regeneration in riparian zones of the Red Deer River (Alberta) was strongly associated with major floods.
This paper provides an overview of the ecological aspects of floods in Canada (e.g. water quality, habitat connectivity, ice jams, etc.), a vast region (~10 million km2) of North America that covers eight ecozones and five major physiographic regions. More specifically, the key objectives are to: (1) synthesize the state of knowledge of the role of open-water and ice-jam flooding on river and floodplain ecology; and (2) identify threats and knowledge gaps for future ecological flood studies in Canada. This information will ultimately provide valuable knowledge for both water and fisheries managers that will contribute to better protection and management of these important natural resources.
Floods and river ecology
Floods occur when the river flow exceeds the bankfull discharge (Qbf), which also represents the effective or channel-forming discharge (Leopold Citation1994), and spills water onto the adjacent floodplains (Gurnell Citation1997). For American rivers, the Qbf commonly approximates the QT2, where QT2 is a flood of a 2-year recurrence interval or the maximum flow that is reached on average every 2 years (Leopold Citation1994). For some Canadian rivers the channels are proportionally larger, with channel widening that may be due to ice scour. Subsequently, Qbf can be higher and around the QT5 for some rivers (Smith Citation1979; Kellerhals and Church Citation1980; Polzin and Rood Citation2006), but there have been limited field assessments of the impacts of ice on channel forms (Morse and Hicks Citation2005). The bankfull recurrence interval is dependent on geographic location and the flood frequency characteristics of the stream (Watt Citation1990). Under open-water conditions, the areas of riparian inundation increase with increasing magnitude and duration of flood flows, and the floodplain is typically fully inundated with the QT100, or 1-in-100-year flood (Gurnell Citation1997; Kalischuk et al. Citation2000).
In cold regions of the world, such as Canada, floodwaters can be generated under ice-influenced and open-water conditions. Although not as widely studied as the open-water period, the literature suggests that floating ice/ice breakup is a key component in freshwater systems that creates and controls aquatic habitats (Prowse and Culp Citation2003; Peters et al. Citation2014a). River ice breakups can occur under mild air temperature conditions where the ice generally decays in place and does not cause jamming, termed a thermal breakup. In contrast, when a breakup is caused by a sudden increase in flow/water level and occurs under thick and still hard ice conditions, the breakup is called a mechanical breakup and the potential for ice jamming is high. The jamming of ice may present significant resistance and obstruction to the intense freshet flow, leading to extremely high river stage at considerably less discharge than under open-water conditions (Figure ) (see Beltaos Citation2014 for more details on river ice breakup). In general, the study of river ice processes and hydraulics is a relatively young field of research (Hicks Citation2008), as is the study of winter dynamics of stream ecosystems (Huusko et al. Citation2007). For instance, it is only recently that the effects of ice have been explicitly incorporated in the development of sustainable ecological flow needs approaches in Canada (Peters et al. Citation2014a).
Figure 1. Annual peak water level vs. discharge under breakup conditions in the lower Peace River at Peace Point hydrometric station 70 km upstream of Peace Delta, headwaters of the Mackenzie River Basin, northern Alberta (updated from Peters et al. Citation2006). Data are presented for the pre-regulation (1962–1967), filling of Williston Reservoir (1968–1971), and post-regulation (1972–2008) periods. Dashed line depicts the magnitude of the historical peak open-water flow event (modified from Peters et al. Citation2014a). masl: metres above sea level.
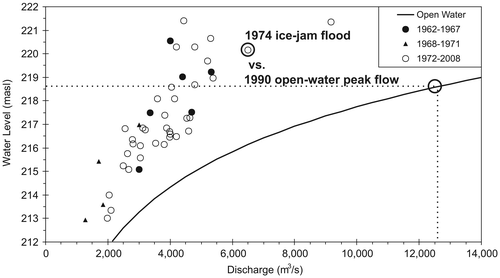
The relative importance of flood mechanisms across Canada was recently investigated by von de Wall (Citation2011). His study demonstrated that under the present climate regime, ~32% of the rivers were dominated by annual extreme peak water levels generated under river ice breakup, ~45% open-water and ~23% mixed conditions. These results are consistent with a report that an estimated 40% of the floods in Canada over the years 1983–1987 were caused by ice-jam events (see Peters et al. Citation2014a). As will be demonstrated in the present study, ice jamming in combination with large spring freshet snowmelt runoff is often the only mechanism capable of inundating elevated portions in a floodplain, particularly in northern deltaic environments (Peters et al. Citation2006; Lesack and Marsh Citation2010).
Role of habitat connectivity and ecological recovery during floods
It has been suggested in the literature that the ecological integrity of river ecosystems depends on their natural dynamic character (Richter et al. Citation1996), as well as habitat connectivity (Altermatt et al. Citation2011). A seminal paper by Poff et al. (Citation1997) synthesized the scientific knowledge to derive the “natural flow paradigm,” which emphasizes the critical role of intra- and inter-annual variations in flow characteristics in sustaining and protecting native biodiversity and ecosystem integrity in river systems. The authors identified five ecologically relevant components of the hydrological regime, namely the magnitude, frequency, duration, timing and rate of change of hydrological events. These components of the hydrograph, presented in Figure , can be used to characterize the range of flows and specific hydrologic occurrences, such as flood events. Notably, the magnitude, frequency, duration and timing of floods are of particular importance in river ecosystems (e.g. productivity, response to habitat availability, etc.) (Poff et al. Citation1997). Some aspects of floods can be beneficial (e.g. removing fine sediments) whereas other may present an impact (e.g. scouring of redds). In fact, the physical structure and habitats are largely defined by flood processes within the channel and between the channel and floodplain.
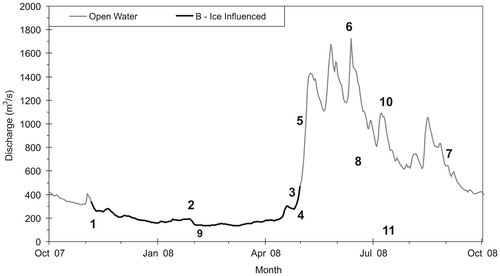
Figure 2. Hydrograph showing open-water and ice-influenced streamflow conditions observed on the lower Athabasca River just below Fort McMurray in the headwaters of the Mackenzie River Basin over the 2008 water year. Also shown on the figure are key hydrograph variables: (1) fall freeze-up magnitude and timing, (2) ice-influenced season, (3) spring freshet initiation date and magnitude, (4) spring breakup magnitude and timing, (5) hydrograph rise rate, (6) peak flow magnitude and timing, (7) hydrograph fall rate, (8) open-water season, (9) low-flow magnitude and timing, (10) number of times the hydrograph reverses direction, and (11) baseflow (source of hydrometric data, Environment Canada Citation2013; modified from Peters et al. Citation2014a).
Floods play an important role in the delivery of food and energy to streams, especially to small streams where the amount of sunlight is limited. For example, much of the energy in large rivers comes from sunlight, thus contributing to the photosynthetic and biological activity; however, much of the energy in small streams come from external, or allochthonous, sources (i.e. nutrients, terrestrial plant debris composed of leaves, needles, and twigs, etc.) from the nearby floodplain/riparian habitat during high-water events (Hynes Citation1975). This energy source coming from the drainage basin is critical for aquatic invertebrates and fungal populations, which in turn provide the basic diet for many species of fish. Accordingly, floods are crucial to the food web dynamics of small streams and large rivers. In recognition of this, Junk et al. (Citation1989) put forward the “flood pulse concept,” a conceptual framework that highlights the importance of annual floods in shaping ecosystem structure and contributing to a healthy river ecosystem. This concept, extended by Tockner et al. (Citation2000), is considered to be one of the most important aspects influencing the biological productivity of riverine ecosystems. In the flood pulse concept, floods are seen as important attributes linking terrestrial sources to stream productivity. As flooding occurs, water levels increase within the floodplain area and provide nutrients and organic matter that tend to increase the stream primary production.
Habitat connectivity under different flow regimes, including extreme flows, is critical to the overall ecological integrity and subsequent management and restoration of river corridor ecosystems (Pringle Citation2001; Reich and Lake Citation2014). In fact, the structure and function of communities and their ability to recover from disturbances are directly associated with their dispersal ability and the degree of connectivity with other habitats (Altermatt et al. Citation2011; Gallardo et al. Citation2014). The spatial and temporal variability in connectivity between the main channel and the floodplain is highly complex (Pringle Citation2001; Amoros and Bornette Citation2002). To support the development of directed management strategies (see the section on potential threats to flood regimes, with ecological implications for more discussion), the approach of Gallardo et al. (Citation2014) could be applied, where they identified groupings of hydrological connectivity for streams and rivers along a gradient from low connectivity (i.e. only connected under extreme flood events) to intermediate connectivity (i.e. connected during high flows) to high connectivity (i.e. connected during low-return flood events such as bankfull). This connectivity gradient is related to the spatial availability of water, nutrients and sediment within the floodplain ecosystem (e.g. Trigg et al. Citation2013; Larsen et al. Citation2015) In addition, using variables that quantify hydrological variability (e.g. stability of flows, flashiness of flows), connectivity and flow predictability, researchers have applied classification approaches to identify groups of rivers to allow direct comparison of ecological patterns and responses to hydrological variability across large and diverse geographical areas (Poff and Ward Citation1989; Kennard et al. Citation2010; Monk et al. Citation2011).
Flow variability shapes ecological communities through taxon-specific responses and associated adaptation of key biological traits to flow variability along the hydrological connectivity gradient (Poff Citation1997; Lake Citation2000) (Table ). For example, in the case of invertebrates, overall community-level taxon richness often peaks at intermediate hydrological connectivity and disturbance where generalist and specialist taxa coexist (Ward and Stanford Citation1983; Tockner et al. Citation2000). More specifically, the taxon richness of Ephemeroptera and Plecoptera often peaks at sites with high hydrological connectivity because of their high dissolved oxygen requirements (Usseglio-Polatera and Tachet Citation1994). This knowledge allows for the development of diagnostic indices to assess flow impacts for ecological flow strategies using the structure of the benthic macroinvertebrate community to understand antecedent flow conditions (Armanini et al. Citation2011; Peters et al. Citation2012).
Table 1. Examples of habitat and biological response under increasing hydrological disturbance.
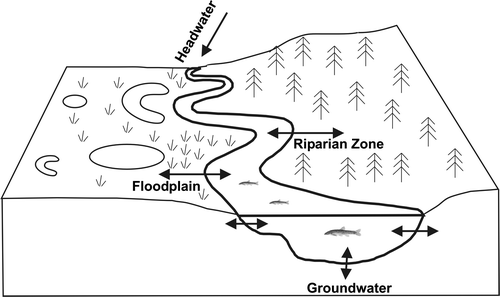
Flow variability within the river system can result in a naturally shifting mosaic of available habitat patches and variable patterns/timing of connectivity (lateral, vertical and longitudinal) with adjacent habitats, such as floodplain wetlands and oxbow lakes (Stanford et al. Citation2005; Whited et al. Citation2007) (Figure ). The timing, duration and extent of ice in northern rivers affect the degree of aquatic habitat isolation and potential refugia availability, leading to shifts in community assemblages following flood disturbance events. For instance, in their seminal paper describing the riverscape concept, Fausch et al. (Citation2002) explained that reduced winter freezing in pools, associated with higher flow conditions, results in greater habitat connectivity that allow for recolonization of previously disconnected habitats. Hydrological connectivity is especially relevant in deltaic environments which largely depend on flood flows and occasional major floods associated with ice jams to provide waters to important habitats (see below for further discussion). High-flow events can cause high hydraulic stress followed by habitat restructuring and resulting in increased mortality to the benthic communities (Death Citation2010; McMullen and Lytle Citation2012; Robinson Citation2012). However, flow refugia patches, characterised by low hydraulic stress during a high flow event (e.g. backwaters, side channels and woody debris; Hax and Golladay Citation1998; Matthaei et al. Citation1999; Schwartz and Herricks Citation2005), are often maintained during high flows and allow for the persistence of aquatic communities (Palmer et al. Citation1995; Sueyoshi et al. Citation2014).
Figure 3. The four-dimensional nature of the riverine ecosystem (Ward Citation1989; Ward and Standford Citation1989): longitudinal connectivity along the entire length of a stream; periodic lateral connectivity to the floodplain and/or riparian zone (terms often used for the same streamside areas), and the resulting exchange of water, sediment, organic matter, nutrients and organisms; vertical connectivity between the atmosphere, streamflow and groundwater; temporal connectivity of continuous physical, chemical and biological interactions over time. As outlined in the text, the floodplain provides a mosaic of aquatic habitats, such as oxbow lakes and wetlands, and potential floodwater storage areas.
Aquatic taxa demonstrate resistance to hydrological disturbance events, for example the ability to move (actively or passively) among habitat patches to flow refugia, or resilience through dispersal of new recruits from adjacent refugia (Lancaster Citation2000). The ability to recolonize at the local scale following a flood disturbance can be rapid depending on connectivity to refugia and the local species pool (Mackay Citation1992; Lake Citation2000; Rice et al. Citation2010; Bertrand et al. Citation2013), via different mechanisms including longitudinal (i.e. drift, crawling; e.g. Layzer et al. Citation1989; Downes et al. Citation2005; Rios-Touma et al. Citation2012), lateral (i.e. from floodplain habitats) and vertical (i.e. from the hyporheic zone) (Olsen and Townsend Citation2005) pathways, or from adult dispersal and ovipositing (Tronstad et al. Citation2007).
Redd scouring can be associated with adverse flood conditions. Lapointe et al. (Citation1999) suggested that a 15% increase in Atlantic salmon egg pocket scouring occurs as floods increase from frequent recurrence to more extreme values, i.e. with return periods over 100 years. Fish are vulnerable not only at the egg or larval stage but also during the emergence period, i.e. when they come out of the gravel and their swimming ability is still limited. During the period of emergence, fry (0+) can be washed away by high flows during the first few weeks before their swimming ability improves and they start feeding. Flood timing in conjunction with the intragravel temperature (which will determine the timing of emergence) is important. Studies have shown that high spring floods have generally resulted in lower population density of some non-native fishes (e.g. brown trout) in southern Hokkaido, Japan (Kawai et al. Citation2013). In fact, the study showed that emerging brown trout were most negatively affected by floods.
Floods and water quality
High flows may affect the natural dynamic character of rivers and habitat connectivity, as well as streamflow water quality. Floods can modify the chemical composition of the water column, the relative surface to groundwater contributions, erosional processes and the suspended sediment load (Glover and Johnson Citation1974; Vegas-Vilarrubia and Herrera Citation1993), potentially leading to shifts in the structure and function of the aquatic communities (e.g. Thomaz et al. Citation2007). High-flow events also influence sediment processes, sediment delivery (and corresponding loads), particulate organic matter, nutrients, potentially increasing the resistance of aquatic ecosystems to future disturbances (Amoros and Bornette Citation2002; Leigh and Sheldon Citation2008; McMullen and Lytle Citation2012; Gallardo et al. Citation2014).
During flood events, the sediment load and transport can lead to important geomorphological changes to the river environment and impact on aquatic organisms (i.e. fish and invertebrates) (e.g. Milner et al. Citation2013). The suspended sediment load (sediment within the water column) tends to have a greater direct impact on fish by reducing their capacity to process oxygen through gill abrasion, as well as impacting the capacity of fish to capture their prey (Waters Citation1995). In many instances, flood events are associated with suspended sediment concentrations (SSC) that exceed 80 mg/L, which is considered a threshold above which aquatic resources are likely impacted (Hynes Citation1973). In addition, the Canadian Council of Environment Ministers’ guidelines stipulate that an SSC increase of 25 mg/L above the background level may be detrimental to aquatic life. SSCs in rivers are characterized by a hysteresis effect, i.e. different SSCs during the rising and falling limbs of the flood hydrographs for a given flow value (e.g. Sawada and Johnson Citation2000). Consequently, the range of SSCs that fish and invertebrates are exposed to and must withstand differs greatly even within a specific flood event. Robertson et al. (Citation2006) conducted an extensive review of the impacts of sediments on fish and fish habitat in Canada. Among others, they noted physiological impacts of high SSCs on fish that included elevated blood sugar, gill flaring, increased cardiac output and decreased resistance to disease. The concentration thresholds that trigger such physiological impacts vary with fish species.
Floodwaters can also mobilize fine sediment delivered from the catchment, washload, bank erosion and instream resuspension. For example, sand and debris deposited in pools and riffles by winter and spring floods can have an impact on fish populations because of the destruction of their habitat (Saunders and Smith Citation1965; Elwood and Waters Citation1969; Pearsons et al. Citation1992; Curry and MacNeill Citation2004; Wohl et al. Citation2015). When floodwaters recede, settling of fine sediments will occur in river reaches with low gradient and velocity. Under such conditions, fine sediments (material < 2 mm) will generally deposit within the stream channel environment. If the fine sediment exceeds 15–20% of the total substrate matrix, then both the eggs and the emergence of fry (young of the year fish) are negatively impacted (Peterson and Metcalfe Citation1981).
In addition to the sediment load (suspended and within the stream substrate), other water-quality variables of importance to the river ecology vary in concentration during floods. Certain chemical constituents tend to become diluted during high-flow events, while others increase in concentration or load as material is flushed from the surrounding basin to the river (Kurtenbach et al. Citation2006). For instance, most dissolved ions tend to dilute during high flows (e.g. calcium (Ca2+), specific conductance, etc.) whereas other variables such as suspended sediment (see above), dissolved organic carbon and some metal concentrations tend to increase with discharge (Feller and Kimmins Citation1979; Caissie et al. Citation1996). Water-quality variability associated with floods is, to a certain extent, a normal occurrence, and aquatic organisms have adapted to withstand such natural variability. However, the chemical composition of flood flows that influence aquatic organisms can also be a function of land use (e.g. forested vs. agricultural), especially for nutrient loading (Poor and McDonnell Citation2007). High flows lower than bankfull discharge can also impact aquatic resources, particularly in urban and agricultural areas during high summer precipitation events. In the absence of beneficial management practices, both sediments and chemicals (nitrogen-rich fertilizer, pesticides, etc.) are flushed from the landscape to the stream during precipitation events. Such events have been found to cause fish kills in highly agricultural areas (e.g. Prince Edward Island), and these events are often associated with rivers reaching anoxic levels due to the high influx of nutrients. These events have resulted in increased primary production and high oxygen demand, thus resulting in anoxic events.
pH also demonstrates high variability during flood events. The timing and severity of the pH depression and concentration of metals (e.g. aluminum) during the spring freshet can adversely affect salmonid populations (Campbell et al. Citation1992; Dennis and Clair Citation2012). The impact of pH depression is more pronounced in low-buffering-capacity (low-alkalinity) environments. For instance, studies have shown the Atlantic salmon is particularly vulnerable to spring pH depression which has limited salmonid production in rivers of southwest Nova Scotia (Watt Citation1987).
Floods influenced by river ice breakup/ice jamming can also directly affect water quality. The role of ice breakup and ice-jam floods is addressed separately below because water quality under ice-breakup processes can be different than under open-water conditions. The ecological impact of river ice breakup-induced floods is relatively poorly understood compared to the open-water floods. Nevertheless, the reader is referred to Prowse (Citation2001a, Citation2001b) and Prowse and Culp (Citation2003) for more in-depth syntheses of the biological, hydrologic, geomorphic and water-quality aspects of river ice ecology.
Role of ice breakup and ice-jam floods
In general, the high flow and rapid changes in water stage, velocities, temperature, SSCs and mechanical action of ice (intense scouring) can combine to make breakup events an erosive cause of change for river channels, banks and adjacent riparian zones (Scrimgeour et al. Citation1994; Prowse and Culp Citation2003). Similar to open-water situations, sediment concentrations have been reported to increase; however, the increase is generally higher by an order of magnitude during river ice breakup periods (Beltaos et al. Citation1994; Milburn and Prowse Citation1998). Repeated deposition of sediment-laden floodwater at elevations higher than the open-water maximum stage can stabilize and elevate floodplains (e.g. Stewart-Schumm Citation1985). Under extremely high water conditions, flow can be redirected into ancillary channels (MacKay et al. Citation1974), make a diversion channel the principal channel (King and Martini Citation1984) or create an entirely new channel; distributary channel systems in rivers deltas are particularly prone to shifting (Dupre and Thompson Citation1979; Timoney Citation2013).
The severity and frequency of ice-breakup floods can affect the riparian and aquatic vegetation communities found in and along the channels, including extensive floodplain areas (Prowse and Culp Citation2003). A demarcation line can often be seen between mature trees along the upper banks and less permanent vegetation below. Localized damage to vegetation can occur from ice push/scour and, in extreme cases, denudation of vegetation. Depending on site exposure, scour damage, severity of and frequency of ice jamming, re-colonization can result from the re-introduction of species from adjacent undisturbed riparian zones and/or via regrowth from the penetrating organs of plants with a widespread root structure (e.g. willows). As outlined in Prowse and Culp (Citation2003), the greatest diversity of vegetation in the riparian zone should occur in cases where moderate-severity events or a mix of events of varying severity occur, whereas sites affected only by thermal breakup events will experience a more biologically mediated ecological succession of plant communities down the river banks. As the vegetation community moves towards dominance by the mature sere, species diversity and biological productivity decrease. As will be shown later, river-ice breakup can also affect vegetation type and structure on floodplains well away from the upper limit of ice scour via the spread of overbank flooding over extensive lowland areas.
There is some evidence that aquatic macrophytes are not always affected by ice-breakup disturbance. For example, an unusual ice jam on the St. Claire River in eastern Canada in April 1984 was observed to delay the growth of submerged macrophytes, which was attributed to the attenuation of light by the packed ice; yet by September the total biomass, abundance of taxa and their frequency were similar to those observed in the previous year (Nichols et al. Citation1989). Moreover, the action of dynamic breakups has a greater potential to deplete algal biomass in the rivers compared to thermal breakups that would largely leave algal communities undisturbed during the spring (Scrimgeour et al. Citation1994). This study also stated that it is reasonable to assume that severity of breakup could affect the timing of peak algal biomass, abundances of individual species and taxonomic community composition.
In contrast to the open-water period, the effects of ice-jam flooding on benthic invertebrates and fish have not been widely studied in Canada or internationally, which is likely partly due to the difficulty in undertaking observations in very difficult conditions and often remote locations. The information available from the literature regarding benthic organisms suggests that the trauma induced by river ice breakup is temporary (Bradt and Wieland Citation1981). Avoidance behaviour is one possible reason for the apparent resilience of some invertebrates, which can use the within-substrate interstitial spaces and hyporheic habitats (Clifford Citation1969; Müller et al. Citation1976; Harper Citation1981). Notably, the movement of fish (white sucker, carp and brown trout) during flooding and ice breakup was observed in the Grand River (Ontario) (Brown et al. Citation2001). These authors also tracked fish to backwater habitats and stated that ice breakup and accompanying flooding resulted in several fish being stranded on the floodplain. The stranding of fish in dewatered floodplain areas, following ice-jam release and the rapid decline in water levels, was also noted by Needham and Jones (Citation1959). A study by Cunjak et al. (Citation1998) in eastern Canada revealed that a mid-winter breakup event caused a major decline in survival of Atlantic salmon, especially egg and juvenile. Overall, ice-jam flood events have the potential to negatively affect fish communities. For example, under ice-jamming conditions, some rivers have shown poorer overwinter survival of juvenile fishes. This was observed in the Little Southwest Miramichi River after a significant ice jam in April of 1994, and where Atlantic Salmon parr densities were lower than the previous years following the ice-jamming event (D. Caissie, unpublished data).
Similar to open-water floods (see above), ice-jam flood events are also known to affect a number of water-quality parameters, such as water temperature, dissolved oxygen (DO), nutrients and movement of sediments (Prowse and Culp Citation2003). For instance, turbulent mixing can cause DO levels to temporarily rise to supersaturation levels (Tilsworth and Bateman Citation1982; Milburn and Prowse Citation2000). Large amounts of dissolved/particulate organic material are entrained by floodwaters from the river banks and floodplain into the channel (a major source of energy to lotic food webs), and can rapidly metabolize with increased water temperatures and lead to an important decrease in DO (Hynes Citation1970; Brunskill et al. Citation1973; Hou and Li Citation1987). Large input of organic material and sediment deposited in the channel after an ice-jam flood event can also impair primary production, reduce habitat and create difficulties in feeding and filtering for benthic organisms (Prowse and Culp Citation2003).
Floods and floodplain ecology
Ecological links to floodplain habitats
Many floodplain zones throughout North America and around the Northern Hemisphere naturally support riparian woodlands where cottonwoods and riparian-adapted poplar trees were often dominant species (Figure ) (Cooke and Rood Citation2007). Across Canada, three cottonwoods are widely distributed, with the black cottonwood (Populus trichocarpa) in Pacific drainages, the closely related balsam poplar (P. balsamifera) in boreal regions, and the prairie or eastern cottonwood (P. deltoides) in the prairie and southeastern zones. Riparian willows are also common, and these shrubs or small trees are related to and share ecophysiological characteristics with cottonwoods (Stromberg et al. Citation2007).
Figure 4. The flooded floodplain forest along the Oldman River in southwestern Alberta during a 10-year flood (QT10), 19 June 2014. Features include: (1) the river channel; (2) an eroding cut-bank along the outside of a meander; (3) a submerged meander lobe point bar with young cottonwood saplings from the prior flood of 2005; (4) flooded groves of narrowleaf and prairie cottonwoods and their hybrids; (5) exposed terraces above the floodplain that are used for livestock pasture (lower right) or cultivated crops (upper; not shown); and (6) the hillslopes that lack trees due to insufficient local precipitation. In wetter regions, terraces and hillslopes are wooded, generally by conifers in many Canadian ecoregions.

Cottonwoods are keystone species for the riparian woodlands and provide the foundation for the floodplain forest ecosystems (Naiman et al. Citation2005). The trees influence channel configuration by resisting bank erosion, and contribute to the fishery and aquatic ecosystem with leaf litter that enriches the aquatic food web, and woody debris that provides aquatic habitat and further influences channel patterns (Gurnell Citation1997; Rood et al. Citation2005). Cottonwood groves provide rich habitats for birds and other wildlife, and offer shade and shelter for livestock (Naiman et al. Citation2005). Riparian woodlands are preferred areas for human recreation, and unfortunately also for homes and other buildings that are subsequently vulnerable to the eventual flood events.
The cottonwoods and willows are generally obligate riparian plants, ecological specialists that are well adapted to the physically dynamic floodplain zones (Scott et al. Citation1997; Rood et al. Citation2007). The willows and especially the cottonwoods are dependent upon dynamic river flows, and particularly flood events, for sexual reproduction that is essential for periodic woodland replenishment and also required for genetic recombination for long-term adaptation (Scott et al. Citation1996; Cooke and Rood Citation2007; Stromberg et al. Citation2007). The high flows enable woodland reproduction by: (1) excluding the competitive encroachment from flood-intolerant upland vegetation; (2) driving the erosion, transport and deposition of alluvial sands, gravels and cobbles to create and expand point and lateral bars that are the primary seedling colonization sites; and (3) saturating the suitable elevational bands and providing subsequent gradual post-flood recession that is essential for seedling survival (Figure ) (Scott et al. Citation1996; Kalischuk et al. Citation2000; Rood et al. Citation2007).
As adaptations to the dynamic floodplain environments, cottonwoods have evolved a specialized r-selection reproductive strategy. They produce vast numbers of tiny seeds that have cottony enclosures for initial wind dispersal and subsequent floating on the river surface and deposition in bands along the saturated stream banks, positions that are optimal for establishment (Kalischuk et al. Citation2000; Polzin and Rood Citation2006). The seeds are released over short intervals typically during the late-spring post-peak flow recession, and with their small size the seeds are viable for only a few weeks. Thus, their life history is closely coordinated with the natural river flow regime, and subsequently floodplain forests are adapted to and dependent upon floods (Scott et al. Citation1997; Auble and Scott Citation1998; Rood et al. Citation2007).
Dendrochronological studies assess cottonwood tree age distributions and these have revealed pulses of recruitment that followed major flood events, commonly with a recurrence interval of 10 years or greater (Scott et al. Citation1997; Samuelson and Rood Citation2004). Complementary analysis has involved the direct observation of cottonwood and willow recruitment following major floods, and this confirms the quantitative association whereby larger floods typically enable more extensive bank and floodplain disturbance and cottonwood recruitment (Gurnell Citation1997; Kalischuk et al. Citation2000). The seedling recruitment follows not only in the flood year but also in the subsequent few years, until the barren colonization sites that are formed by the erosion of vegetation and sediments and sediment deposition are occupied by native riparian plants or, unfortunately, invasive exotic plants (Stromberg et al. Citation2007; Polzin and Rood Citation2006). It is now understood that it is not only the flood magnitude but also the post-flood pattern that is critical for the essential occasional replenishment of floodplain forests (Rood et al. Citation2005). The flood and post-flood sequences thus naturally produce pulses of recruitment of riparian woodlands, consistent with the broader flood pulse concept for riverine ecosystems (Junk et al. Citation1989; Bovee and Scott Citation2002).
Emerging from this advancing understanding of the coordination between river floods and floodplain forests, a current focus in river science and restoration involves the deliberate regulation of ecological flows to encourage the replenishment of cottonwoods and other native riparian plants. Following damming, flood flow attenuation has reduced floodplain dynamics and woodland replenishment along many rivers (Friedman et al. Citation1998; Wilding et al. Citation2014), and while it would generally be unacceptable to intentionally deliver flood flows, there are attempts to restore more dynamic flow patterns with seasonal variation that better resembles the natural flow regime (Poff et al. Citation1997; Stromberg et al. Citation2007). Further, when floods do occur, such as following exceptionally heavy and widespread rain events, there can be efforts to manage the post-flood flows to deliver gradual stage recession (“flow ramping”) that allows the elongating roots of the new cottonwood and willow seedlings to track the receding moisture zone (Kalischuk et al. Citation2000). Simultaneous land-use management is also essential, and livestock or recreation use of the recruitment zones should be limited during the colonization phase after flood events to avoid seedling destruction. Following from these principles, there have been some major restoration successes along extensively regulated rivers in semi-arid ecoregions, and this provides promise for the conservation and restoration of other floodplain forests across Canada and worldwide (Molles et al. Citation1998; Rood et al. Citation2005; Stromberg et al. Citation2007).
The development and testing of ecological flow guidelines to restore and maintain riverine and floodplain ecosystems have become increasingly important in recent decades (e.g. Caissie and El-Jabi Citation2003; Peters et al. Citation2012; Linnansaari et al. Citation2013). These studies have shown the importance of ecological flows, and particularly high flushing flow targets below dams. The objective of having such short-duration floods (e.g. a few days to weeks) is to maintain channel morphology as well as to flush fine sediments. To help maintain a deltaic ecosystem in a cold regions, the potential of enhancing the extent of ice-jam flooding in a downstream deltaic floodplain environment via a small reservoir release was demonstrated in the Peace River Delta system during the spring of 1996 (Prowse et al. Citation2006).
Floods and deltaic environments
The importance of flooding for maintaining the health of freshwater riparian ecosystems, specifically deltas, has become widely studied over the last 30 years (e.g. Mackenzie River Basin Committee [MRBC] Citation1981; Marsh and Lesack Citation1996; Prowse and Conly Citation2002; Brock et al. Citation2008; Lesack and Marsh Citation2010; Wiklund et al. Citation2012). Derived from river-borne sediments at the edge of a lake or ocean, deltas are formed by a complex of aquatic, semi-aquatic and terrestrial zones, such as estuaries, beaches, dunes, distributary channels, levees, bays, lakes, wetlands, meadows and woods. For instance, the Mackenzie River basin, the largest and longest river system in Canada, includes two major freshwater floodplains, the climate- and flow regulation-influenced Peace–Athabasca Delta (PAD) and Slave River Delta (SRD), and the relatively pristine estuarine Mackenzie River Delta (MRD) (MRBC Citation1981). Figure presents an example of the spatial extent of a very large spring ice-jam flood of the PAD that occurred in 1974, versus the historical extreme high flow (under open-water conditions) on the Peace River in June 1990 that did not overflow into the delta (see Figure for a difference exceeding 1.5 m under the ice-jam flood of 1974).
Figure 5. Landsat satellite images of the Peace–Athabasca Delta in northern Alberta: (a) early May 1974, showing the extensive ice-jam generated overland flow (blue) and ice (white); (b) late June 1990, following the all-time high open-water flow event measured on the lower Peace River that resulted in less than bankfull flow conditions (blue) and no apparent flooding of perched basins (green areas in Zone 1). As highlighted in Figure 1, the ice-jam event generated backwater levels > 1.5 m higher with considerably less discharge than the peak flow event that did not overflow the river bank in Zone 1 (modified from Peters et al. Citation2014a).

A prominent feature of the deltas in the Mackenzie River Basin is an abundance of shallow, productive, macrophyte-dominated wetland and lake basins with low to moderate phytoplankton abundance that experience varying degrees of surface water connectivity to the main flow system (Peters et al. Citation2006; Wolfe et al. Citation2007; Sokal et al. Citation2008; Squires et al. Citation2009; Lesack and Marsh Citation2010). In general, low-elevation basins are potentially flooded annually and for relatively long durations, while basins perched at higher elevations are flooded infrequently and for relatively shorter durations (Lesack and Marsh Citation2010). As discussed above for the riverine ecosystem, the ecological integrity of deltaic floodplains depends on their natural dynamic character, which includes occasional surface water connectivity to the main flow system. It is important to mention that in more populated areas of southern Canada, there are examples where human interventions to accommodate urban, agricultural and industrial development (i.e. drainage, flood protection works, channel confinement, and dredging) have noticeably impacted lateral connectivity, such as in the case of the Fraser River Delta (Barrie and Currie Citation2000; Evans-Ogden et al. Citation2008).
In Canada and throughout the world, delta floodplains are recognized as ecologically and culturally important freshwater landscapes that are strongly regulated by variations in river flow/level and floodwater connectivity (e.g. Lesack and Marsh Citation2010). Seasonal and periodic flood events have been shown to play an important role in the water balance, light, environment, habitat availability, nutrient supply, productivity and community composition of the basin receiving water (van der Valk and Bliss Citation1971; Squires and Lesack Citation2002, Citation2003; Squires et al. Citation2002, Citation2009; Junk and Wantzen Citation2004; Wolfe et al. Citation2007; Brock et al. Citation2008; Wantzen et al. Citation2008; Sokal et al. Citation2010). The dynamics of wetlands and lakes within deltaic floodplains, in terms of their overall biological structure and productivity, is dependent on occasional floodwater input, deposition of material and flushing during high-river-stage events.
For example in the PAD, river floodwaters that entered delta wetland/lake basins were observed to elevate concentrations of suspended sediments, total phosphorus (TP), sulphate (SO4) and dissolved silica (DSi), and reduce concentrations of total Kjeldahl nitrogen (TKN), dissolved organic carbon (DOC) and most ions (Wiklund et al. Citation2012). Soon after an event, water clarity increased as the suspended sediments and TP settled out of the water column, with no appreciable change to DOC, SO4, TKN and ions. After flood waters subside and lakes become disconnected from the river, limnological conditions will diverge because of local basin differences (Tockner et al. Citation2000; Junk and Wantzen Citation2004; Thomaz et al. Citation2007). Over the years, generally semi-arid climate conditions (evaporation greater than precipitation) influencing the water balance of wetland/lake basins leads to an increased concentration in most nutrients and ions. In the SRD, phytoplankton standing crop, measured as chlorophyll-a concentration (chl-a), was highest in deltaic lake basins that did not flood because water column nutrient concentrations were highest in these lakes (Sokal et al. Citation2008, Citation2010). Wiklund et al. (Citation2012) concluded that the above results demonstrated that regular flooding was not required to maintain high nutrient concentrations.
Farther to the north in the MRD, delta lake basins with extended surface connection times to river water with multiple water level resets per year to the spill elevation yielded lakes with a high degree of similarity (Lesack and Marsh Citation2010). These authors stated that the combination of deltaic floodplain wetlands and lakes arranged in an intermittently connected continuum may be an important mechanism driving distinctive habitat productivity and biodiversity of aquatic communities in the MRD relative to the surrounding landscape. Similarly to the semi-arid PAD, ponded water levels usually decline until basins dry up or are reset to the spill elevation by floodwater input (Marsh and Lesack Citation1996; Peters et al. Citation2006). Without floodwater inputs to inland lakes and wetlands, deltaic environments desiccate and transition into terrestrial environments. Wetting and drying cycles brought about by ranges in the magnitude and frequency of ice-induced floods are important for overall vegetation diversity and productivity (Lesack et al. Citation1991; Prowse and Conly Citation2002). With the appropriate inundation and drawdown intervals, vegetation is maintained at an early successional stage, forming some of the most biologically productive habitat in the world (MRBC Citation1981).
The frequency, duration and depth of floodwaters have been observed to influence vegetation community composition in deltaic environments. For instance, the interval of spring ice-jam flooding in the MRD was considered to be a major factor responsible for seed production and germination of white spruce in elevated areas (Pearce et al. Citation1988). In the PAD, drawdown after short duration spring ice-jam and/or summer floods exposes a moist mineral seedbed that facilitates willow establishment and expansion, while persistent deep floodwater might act to decrease willow cover (Timoney Citation2013). Aquatics such as pondweeds, duckweeds and coontails tended to rise with flooding and then decline with water drawdown. The latter phase is usually necessary for seed germination of marsh plants such as sedges, grasses, bulrush, cattail, arrowhead and willows (Weller Citation1987).
Overall, the pulsing of water abundance drives habitat cycles favoured by wildlife that inhabits and/or seasonally visits deltas of the Mackenzie River Basin. For instance, the dominant marsh sedge areas of the PAD help support the largest free-ranging wood bison herd in the world. The mixed marsh areas of sedge, spangletop, cattail and bulrush provide ideal muskrat habitat (Timoney Citation2013). The deltas are central to the major waterfowl flyways of North America, with deltaic wetland areas providing ideal habitat influenced by wet–dry cycles. For instance, Butterworth et al. (Citation2002) concluded that the response of waterbirds to the 1996–1997 flood events and the subsequent decline of water levels in the PAD supports the hypothesis that floods are crucial to waterfowl use of the delta. Thus, it is important to protect or restore the natural variability of discharge and the lateral connectivity in deltaic environments.
Potential threats to flood regimes, with ecological implications
One potential threat to flood regimes is climate change. A national climate assessment revealed that Canada has on average warmed by more than 1.3°C since 1948 (about twice the global average), and experienced generally wetter conditions, with mean precipitation across the country increasing by about 12% (Warren and Egginton Citation2008). In particular, with winter and spring warming, snowmelt has been observed to commence earlier in certain regions, especially in western Canada, leading to advancement in the seasonal river flow regime (Rood et al. Citation2008). Influences on overall river discharge have been variable, following apparently increasing precipitation that was countered by increased evaporation (Zhang et al. Citation2000, Meehl et al. Citation2007).
With respect to floods, it had been anticipated that global warming could intensify the water cycle, e.g. more intense precipitation and potentially more prolonged droughts (Meehl et al. Citation2007). More intense precipitation could produce heavier and more extensive rain events that could increase river flooding (Milly et al. Citation2002). This is the climate intensification hypothesis that predicts more frequent and more severe floods, although it is recognized there will always be substantial regional variation (Mailhot et al. Citation2012). In contrast to this prediction, analyses of the historic flood records have not detected widespread increases in either flood magnitude or frequency. This was the case for the contiguous United States, for which the only regionally significant pattern over the century-long historic records was a decrease in flood magnitude in the American southwest (Hirsch and Ryberg Citation2012).
For Canada, Cunderlik and Ouarda (Citation2009) investigated patterns over three decades (1974–2003) and found decreasing magnitudes of spring snowmelt floods for some streams, and little evidence for change in rainfall floods, which can occur in autumn or winter. Covering a similar time period, a national assessment by Monk et al. (Citation2011) found only 3% of hydrometric stations experienced increasing, vs. 11% significant declining (p < 0.05), trends in peak runoff magnitude. Regarding spring breakup flood conditions, von de Wall (Citation2011) observed a general advancement (earlier timing) and declines in the magnitude of peak breakup water levels since 1969 for available station data across Canada. This lack of historical evidence for flood intensification in Canada and the United States is consistent with an analysis of the broader international literature by Kundzewicz et al. (Citation2013) that extended from the assessment by the Intergovernmental Panel on Climate Change (IPCC) (Field et al. Citation2012). Relative to ecological impacts, there have been occasional major floods in the recent past, and it is likely that generally similar patterns of flood frequency and severity will persist at least into the near future.
A potentially changing climate may bring about significant shifts to a river’s dominant flood-generation mechanism, such as a shift from an ice-breakup-dominated flood regime to a mixed or to an open-water peak flow-dominated flood regime. For instance, Beltaos et al. (Citation2006) estimated that under a warmer and wetter climate projected for the 2080s, the Peace River ice season is likely to be reduced by 2 to 4 weeks, the ice cover would be slightly thinner and mid-winter thaws would deplete the snowpack, leading to a severe reduction in the frequency of spring ice-jam flooding in the PAD. The ecological implications of less frequent ice-induced recharge of highly perched deltaic basins are a longer interval of drying and accelerated loss of aquatic habitat because open-water flood levels will likely not be capable of reaching these basins (see Figures and ) (Peters et al. Citation2006). Conversely, in eastern Canada, the available recent historical evidence suggests the Saint John River is becoming subject to more frequent mid-winter jams and higher April flows, both of which augment the potential for major jamming (Beltaos Citation2002) and associated ecological impacts (discussed above). Note that flow regulation also has the potential to reduce the occurrence of spring ice-jam floods as a result of higher fall/winter flows to meet hydroelectric power demands when combined with declining spring runoff (e.g. Beltaos et al. Citation2006; Beltaos Citation2014).
Flow regulation is another important feature of many Canadian rivers that can dramatically modify floods (e.g. magnitude and duration) and corresponding flood processes (e.g. flushing of fines), given that over 10,000 dams exist in the country. The ecological effects of dams will potentially be very different depending on the size of these dams and reservoirs, and their management patterns. The ecological implications of maintaining floods in rivers to conserve these important attributes have often been overlooked in many dam construction and ecological flow studies. Much of the Canadian research (as elsewhere) on the impact of flow regulation on river ecosystems has focused on developing methods to establish ecological flow guidelines for the protection and conservation of aquatic species, including flushing flows or floods, as mentioned above (e.g. Caissie and El-Jabi Citation2003; Linnansaari et al. Citation2013). Flushing flows are relatively high flows that can range between 200% of mean annual flow (Tennant Citation1976) to flows close to or exceeding the bankfull discharge. Also, the duration of these high flows is important. Short-duration high flows (a few hours to a day) will generally flush away fine sediments, and improve spawning as well as egg-incubation gravel conditions and success, whereas longer duration high flows (several days) are generally required when channel maintenance is required (i.e. maintaining channel morphology and characteristics).
In a review of the impact of flow regulation on fish, Murchie et al. (Citation2008) concluded that although there is clear evidence that flow regulation modifies fish habitat, fish response varies widely in amplitude and direction. They also contributed a number of suggestions for improving research related to the impact of flow regulation on fish habitat, including an increase in studies conducted on non-salmonid species, the inclusion of different biological levels (from cellular to ecosystem) and the need to document fish response during the dynamic phases of flow increase. They also mentioned that more research is required on the thermal impact of flow regulation.
Research on the impacts of floods in a regulated context has often focused on hydropeaking and fish, with a clear emphasis on salmonids (Murchie et al. Citation2008). For instance, Scruton et al. (Citation2005) conducted experiments during summer and winter and identified a number of potential impacts of hydropeaking on salmonids, including decreased home ranges, stranding, freezing and depleted energy reserves. Murchie and Smokorosky (Citation2004) monitored movement of brook trout (Salvelinus fontinalis) and walleye (Sander vitreus) in the Magpie River (Ontario). They found that the peak in fish activity corresponded to the peak in flows. Potential causes of increased activity during flood periods include relocation to refugia, increased energy expenditure to maintain position in the water column or on the bottom, and increased feeding associated with greater food availability because of increased drift. In addition, Armanini et al. (Citation2014) demonstrated clear shifts in macroinvertebrate community responses as quantified by the Canadian Ecological Flow Index at hydropeaking sites as compared to reference sites. These studies demonstrate the importance of maintaining floods in order to protect the natural character and dynamic of river systems.
When designing ecological flow guidelines and flood restoration projects, it is important that river scientists and managers understand that fluvial systems have changed and will continue to change through time. For instance, morphological changes to rivers and delta channels have been found to play a key role in flood level recurrence. Studies of some American rivers revealed that although the peak discharge had not changed with time, the associated flood level was markedly increased as a result of river bed adjustments from natural processes and/or various human interventions (James Citation1999; Stover and Montgomery Citation2001; Pinter and Heine Citation2005). In the Athabasca Delta, Wolfe et al. (Citation2008) inferred from paleolimnological studies that an engineered meander cut-off led to reduced flooding and a natural channel bifurcation led to an increased flooding recurrence of adjacent floodplain lakes.
Given the number, diversity and broadening scope of stressors potentially affecting the magnitude and timing of flood events across Canada, effective protection, conservation and restoration of flood processes in riverine and connected floodplains will need to be accomplished using an ecosystem-based management approach, which in some cases involves a watershed scale and multiple jurisdictions. For example, in recognition that rock-fill weirs were not capable of raising water levels into the more elevated floodplain areas of the PAD (Peters et al. Citation2006), hydroelectric reservoir flow releases to enhance flooding in the downstream delta during ice breakup and open-water season were proposed as management options worthy of further assessment (Prowse et al. Citation2006; Peters et al. Citation2014b). Another example of a transboundary, science-based management approach is in the St Mary River system, where Rood et al. (Citation2015) developed a novel twofold restoration strategy comprised of modified dam operations to deliver functional flood flows and direct seeding to enable cottonwood colonization.
The above studies highlight an international shift in the last 20 years or so towards managing river systems based on a better understanding of natural processes, e.g. making space for water in England (Defra Citation2005), room for the river in the Netherlands, the fluvial territory in Spain (Ollero Citation2010), freedom space in France (Piegay et al. Citation2005) and Quebec (Biron et al. Citation2014). Such management approaches promote river–floodplain connectivity and the concept of resilience of fluvial systems, and, as such, move away from traditional engineering approaches for flood protection. Trigg et al. (Citation2013) highlight the importance of a well-connected floodplain to provide flood protection via flow storage, and the risk created by engineered structures (e.g. dykes) which disrupt surface water connectivity. For example, in western Canada, the Okanagan River Restoration Initiative (Okanagan Basin Water Board [OBWB] Citation2015) and Mamquam River Floodplain Restoration Project (MRFRP Citation2015) were initiated in the early 2000s to connect portions of channelized and dyked rivers to their historic floodplain via restoration of meanders, wetland and pond habit, and riparian vegetation.
Conclusions
It is generally recognized in the literature that the ecological integrity of river ecosystems is greatly influenced by their natural dynamic hydrological character, and, notably, extreme events play a role. This Canadian overview paper demonstrates the important roles of flood flows and overflow events and how these potentially influence ecological, geochemical and physical processes of both in-channel and adjacent floodplain zones. In Canada, floods can occur under open-water and ice-influenced conditions. Floods generated from ice jamming are particularly relevant due to the potentially higher water levels produced and the erosive power, but the ecological effects of these events remain poorly understood relative to those occurring during the ice-free period. The ecological impacts and/or benefits of floods can be highly dependent on the flood generation processes, and on their magnitude and timing, which should be considered when designing ecological restoration projects, and remedial or adaptation strategies.
Large floods, such as those generated under ice-jam conditions or those with high return periods (see Buttle et al. Citation2015 in this issue for examples of major floods across Canada since 1995), provide a major source of physical disturbance and suspended sediment movement that can be detrimental to instream aquatic habitat, but beneficial to floodplain ecology. Moderate floods with relatively low return periods can be particularly beneficial to aquatic habitats by providing woody debris that contributes to habitat complexity and diversity, by flushing fine sediments and by providing important food sources from terrestrial origins. In addition to sediment concentration and loads, floods also influence water-quality parameters, such as metals and pH depression, which further influence river ecology.
This overview paper also recognizes the importance of linkages between floods and habitat connectivity and refugia, and the ability of aquatic resources to recover from flood disturbance. Additionally, the floodplain forest plays a role in channel configuration, as well as in many other processes contributing to the river ecology, such as allochthonous inputs of carbon, moderating bank erosion, food-web dynamics and other influences. Several species of trees are dependent on floods for reproduction, and floods can limit competitive encroachment of upland vegetation. Floods play a critical role in deltaic environments where high flows and overflow connect to delta lakes and wetlands, which is important in sustaining water balances (ponded water) and influencing nutrient supplies. Incorporating flood-ecology understanding into basin-scale management approaches is advocated for maximizing the resiliency of riverine and floodplain ecosystems to anthropogenic stressors (e.g. Biron et al. Citation2014).
Significant factors that have already produced changes and will likely influence future flood regimes in Canada are river flow regulation, land-use change, urban development and climate change. Following from the potential changes to the magnitude and timing of floods resulting from these environmental stressors, it is recommended that future research efforts should focus on enhancing our understanding of the ecological aspects of floods, and especially under ice-jam conditions. This new information will prove invaluable in preserving, protecting and managing the natural resources provided by riverine ecosystems.
Acknowledgements
We thank the four anonymous reviewers for their detailed and constructive comments on a previous version of this manuscript.
References
- Altermatt, F., A. Bieger, F. Carrara, A. Rinaldo, and M. Holyoak. 2011. Effects of connectivity and recurrent local disturbances on community structure and population density in sexperimental metacommunities. PloS ONE 6(4): e19525.
- Amoros, C., and G. Bornette. 2002. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshwater Biology 47: 761–776.
- Armanini, D. G., N. Horrigan, W. A. Monk, D. L. Peters, and D. J. Baird. 2011. Development of a benthic macroinvertebrate flow sensitivity index for Canadian Rivers. River Research and Applications 27: 723–737.
- Armanini, D. G., A. Idigoras Chaumel, W. A. Monk, J. Marty, K. Smokorowski, M. Power, and D. J. Baird. 2014. Benthic macroinvertebrate flow sensitivity as a tool to assess effects of hydropower related ramping activities in streams in Ontario (Canada). Ecological Indicators 46: 466–476.
- Auble, G. T., and M. L. Scott. 1998. Fluvial disturbance patches and cottonwood recruitment along the Upper Missouri River, Montana. Wetlands 18: 546–556.
- Barrie, J. V., and R. G. Currie. 2000. Human impact on the sedimentary regime of the Fraser River Delta, Canada. Journal of Coastal Research 16: 747–755.
- Beltaos, S. 2002. Effects of climate on mid winter ice jams. Hydrological Processes 16: 789–804.
- Beltaos, S. 2014. Comparing the impacts of regulation and climate on ice-jam flooding of the Peace-Athabasca Delta. Cold Regions Science and Technology. http://dx.doi.org/10.1016/j.coldregions.2014.08.006.
- Beltaos, S., B. C. Burrell, and S. Ismail. 1994. Ice and sedimentation processes in the Saint John River, Canada. In Proceedings of the IAHR Ice Symposium, Vol. 1, pp. 11–21. Trondheim: International Association for Hydraulic Research.
- Beltaos, S., T. Prowse, B. Bonsal, R. MacKay, L. Romolo, A. Pietroniro, and B. Toth. 2006. Climatic effects on ice-jam flooding of the Peace-Athabasca Delta. Hydrological Processes 20: 4031–4050.
- Benke, A. C. 2001. Importance of flood regime to invertebrate habitat in an unregulated river–floodplain ecosystem. Journal of the North American Benthological Society 20: 225–240.
- Bertrand, K. N., M. R. Whiles, K. B. Gido, and J. N. Murdock. 2013. Influence of macroconsumers, stream position, and nutrient gradients on invertebrate assemblage development following flooding in intermittent prairie streams. Hydrobiologia 714: 169–182.
- Biron, P. M., T. Buffin-Bélanger, M. Larocque, G. Choné, C.-A. Cloutier., M.-A. Ouellet, S. Demers, T. Olsen, C. Desjarlais, and J. Eyquem. 2014. Freedom space for rivers: A sustainable management approach to enhance river resilience. Environmental Management 54: 1056–1073.
- Bovee, K. D., and M. L. Scott. 2002. Implications of flood pulse restoration for Populus regeneration on the upper Missouri River. River Research and Applications 18: 287–298.
- Bradt, P. T., and G. E. Wieland. 1981. A comparison of the benthic macroinvertebrate communities in a trout stream: Winter and spring 1973 and 1977. Hydrobiologia 77: 31–35.
- Brock, B. E., B. B. Wolfe, and T. W. D. Edwards. 2008. Spatial and temporal perspectives on spring break-up flooding in the Slave River Delta. NWT. Hydrological Processes 22 (20): 4058–4072.
- Brown, R. S., G. Power, and S. Beltaos. 2001. Winter movements and habitat use of riverine brown trout, white sucker and common carp in relation to flooding and ice break-up. Journal of Fish Biology 59: 1126–1141.
- Brunskill, G. J., D. M. Rosenberg, N. B. Snow, G. L. Vascotto, and R. Wagemann. 1973. Ecological studies of aquatic systems in the Mackenzie–Porcupine drainages in relation to proposed pipeline and highway developments. Vol. 1 and 2. Environmental–Social Committee, Northern Pipelines, Task Force on Northern Oil Development, Report 73-40 (Vol. 1) and Report 73-41 (Vol. 2). Ottawa: Information Canada.
- Butterworth, E., A. Leach, M. Gendron, B. Pollard, and G. R. Stewart. 2002. Peace–Athabasca Delta waterbird inventory program: 1998–2001 Final Report. Edmonton: Ducks Unlimited, 40 pp. + appendices.
- Buttle, J. M., D. M. Allen, D. Caissie, B. Davison, M. Hayashi, D. L. Peters, J. W. Pomeroy, S. Simonovic, A. St-Hilaire, and P.H. Whitfield. This issue. Flood processes in Canada, regional and special aspects. Canadian Water Resources Journal.
- Caissie, D., and N. El-Jabi. 2003. Instream flow assessment: From holistic approaches to habitat modelling. Canadian Water Resources Journal 28(2): 173–183.
- Caissie, D., T. L. Pollock, and R. A. Cunjak. 1996. Variation in stream water chemistry and hydrograph separation in a small drainage basin. Journal of Hydrology 178: 137–157.
- Campbell, P. G. C., H. J. Hansen, B. Dubreuil, and W. O. Nelson. 1992. Geochemistry of Quebec North Shore salmon rivers during snowmelt: Organic acid pulse and aluminum mobilization. Canadian Journal of Fisheries and Aquatic Sciences 49(9): 1938–1952.
- Clifford, H. F. 1969. Limnological features of a northern brownwater stream, with special reference to the life histories of the aquatic insects. American Midland Naturalist 82: 578–597.
- Cooke, J. E. K., and S. B. Rood. 2007. Trees of the people: The growing science of poplars in Canada and worldwide. Canadian Journal of Botany 85: 1103–1110.
- Cordes, L. D., F. M. R. Hughes, and M. Getty. 1997. Factors affecting the regeneration and distribution of riparian woodlands along a northern prairie river: The Red Deer River, Alberta Canada. Journal of Biogeography 24(5): 675–695.
- Cunderlik, J. M., and T. B. M. J. Ouarda. 2009. Trends in the timing and magnitude of floods in Canada. Journal of Hydrology 375: 471–480.
- Cunjak, R. A., T. D. Prowse, and D. L. Parrish. 1998. Atlantic salmon (Salmo salar) in winter: “The season of parr discontent”? Canadian Journal of Fisheries and Aquatic Sciences 55 (suppl. 1): 161–180.
- Curry, R. A., and W. S. MacNeill. 2004. Population-level responses to sediment during early life in brook trout. Journal of the North American Benthological Society 23(1): 140–150.
- Death, R. G. 1996. The effect of patch disturbance on stream invertebrate community structure: The influence of disturbance history. Oecologia 108: 567–576.
- Death, R. G. 2008. The effects of floods on aquatic invertebrate communities. In Aquatic insects: Challenges to populations, eds. J. Lancaster and R. A. Briers, 103–121. Wallingford, UK.
- Death, R. G. 2010. Disturbance and riverine benthic communities: What has it contributed to general ecological theory? River Research and Applications 26: 15–25.
- Defra. 2005. Making space for water, taking forward a new government strategy for flood and coastal erosion risk management in England. Department of Food and Rural Affairs. http://coastaladaptationresources.org/PDF-files/1329-Making-space-forwater.pdf (accessed June, 2015).
- Dennis, I. F., and T. A. Clair. 2012. The distribution of dissolved aluminum in Atlantic salmon (Salmo salar) river of Atlantic Canada and its potential effect on aquatic populations. Canadian Journal of Fisheries and Aquatic Sciences 69: 1174–1183.
- Downes, B. J., A. Bellgrove, and J. L. Street. 2005. Drifting or walking? Colonisation routes used by different instars and species of lotic, macroinvertebrate filter feeders. Marine and Freshwater Research 56: 815–824.
- Dupre, W. R., and R. Thompson. 1979. The Yukon Delta: A model for deltaic sedimentation in an ice-dominated environment. Proceedings of the 11th Annual Offshore Technology Conference, 657–664. Houston, TX: American Institute of Mining, Metallurgical and Petroleum Engineers.
- Elwood, J. W., and T. F. Waters. 1969. Effects of flood on food consumption and production rates of a stream brook trout population. Transactions of the American Fisheries Society 98: 253–262.
- Environment Canada. 2013. Water Survey of Canada HYDAT archival database of water information collected through the National Hydrometric Program. http://www.ec.gc.ca/rhc-wsc/default.asp?lang=En&n=4EED50F1-1 (accessed January, 2014).
- Evans-Ogden, L. J., S. Bittman, D. B. Lank, and F. C. Stevenson. 2008. A review of agricultural land use by shorebirds with special reference to habitat conservation in the Fraser River Delta, British Columbia. Canadian Journal of Plant Science 88: 71–83.
- Fausch, K. D., Y. Taniguchi, S. Nakano, G. D. Grossman, and C. R. Townsend. 2001. Flood disturbance regimes influence rainbow trout invasion success among five holarctic regions. Ecological Applications 11(5): 1438–1455.
- Fausch, K. D., C. E. Toregersen, C. V. Baxter, and H. W. Li. 2002. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes. Bioscience 52(6): 483–498.
- Feller, M. C., and J. P. Kimmins. 1979. Chemical characteristics of small stream near Haney in Southwestern British Colombia. Water Resources Research 15(2): 247–258.
- Field, C. B., V. Barros, T. F. Stocker, D. Qin, D. J. Dokken, K. L. Ebi, M. D. Mastrandreaet al. 2012. Managing the risks of extreme events and disasters to advance climate change adaptation. Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC). Cambridge: Cambridge University Press.
- Friedman, J. M., W. R. Osterkamp, M. L. Scott, and G. T. Auble. 1998. Downstream effects of dams on channel geometry and bottomland vegetation: Regional patterns in the Great Plains. Wetlands 18: 619–633.
- Gallardo, B., S. Doledec, A. Paillex, D. B. Arscott, F. Sheldon, F. Zilla, S. Merigoux, E. Castella, and F. A. Comin. 2014. Response of benthic macroinvertebrates to gradients in hydrologic connectivity: A comparison of temperate, subtropical, mediterranean and semiarid river floodplains. Freshwater Biology 59: 630–648.
- Glover, B. J., and P. Johnson. 1974. Variations in the natural chemical concentration of river water during flood flows, and the lag effect. Journal of Hydrology 22: 303–316.
- Gurnell, A. 1997. The hydrological and geomorphological significance of forested floodplains. Global Ecology and Biogeography Letters 6: 219–229.
- Harper, P. P. 1981. Ecology of streams at high latitudes. In Perspectives in running water ecology, eds. M.A. Lock and D.D. Williams, 313–337. New York: Plenum Press.
- Hax, C. L., and S. W. Golladay. 1998. Flow disturbance of macroinvertebrates inhabiting sediments and woody debris in a prairie stream. American Midland Naturalist 139(2): 210–223.
- Hicks, F. 2008. An overview of river ice problems: CRIPE07 guest editorial. Cold Regions Science and Technology 55: 175–185.
- Hirsch, R. M., and K. R. Ryberg. 2012. Has the magnitude of floods across the USA changed with global CO2 levels? Hydrological Sciences Journal 57: 1–9.
- Hou, R., and H. Li. 1987. Modelling of BOD–DO dynamics in an ice-covered river in northern China. Water Resources 21: 247–251.
- Huusko, A., L. Greenberg, M. Stickler, T. Linnansaari, M. Nykanen, T. Vehanen, S. Kolkonen, P. Louhi, and K. Alfredsen. 2007. Life in the ice lane: The winter ecology of stream salmonids. River Research and Applications 23: 469–491.
- Hynes, H. B. N. 1970. The ecology of running waters. Toronto: University of Toronto Press, 555 pp.
- Hynes, H. B. N. 1973. The effects of sediment on the biota in running waters. In Proceedings of Hydrology Symposium on Fluvial Processes and Sedimentation, 652–663. Edmonton: University of Alberta.
- Hynes H. B. N. 1975. The stream and its valley. Verhandlungen des Internationalen Verein Limnologie 19:1–15.
- James, A. 1999. Time and the persistence of alluvium: River engineering, fluvial geomorphology, and mining sediment in California. Geomorphology 31: 265–290.
- Junk, W. J., and K. M. Wantzen. 2004. The flood pulse concept: New aspects, approaches, and applications- an update. In Proceedings of the 2nd Large River Symposium (LARS), eds. R. Welcomme and T. Petr, 117–149. Bangkok: RAP Publication.
- Junk, W. J., P. B. Bayley, and R. E. Sparks. 1989. The flood pulse concept in river–floodplain systems. Canadian Special Publication of Fisheries Aquatic Sciences 106: 110–127.
- Kalischuk, A. R., S. B. Rood, and J. M. Mahoney. 2000. Environmental influences on seedling growth of cottonwood species following a major flood. Forest Ecology and Management 144: 75–89.
- Kawai, H., N. Ishiyama, K. Hasegawa, and F. Nakamura. 2013. The relationship between the snowmelt flood and the establishment of non-native brown trout (Salmo trutta) in streams of the Chitose River, Hokkaido, northern Japan. Ecology of Freshwater Fish 22: 645–653.
- Kellerhals, R., and M. Church. 1980. Comment on “Effects of channel enlargement by river ice processes on bankfull discharge in Alberta, Canada” by D. G. Smith. Water Resources Research 16: 1131–1134.
- Kennard, M. J., B. J. Pusey, J. D. Olden, S. J. Mackay, J. L. Stein, and N. Marsh. 2010. Classification of natural flow regimes in Australia to support environmental flow management. Freshwater Biology 55: 171–193.
- King, W. A., and I. P. Martini. 1984. Morphology and recent sediments of the lower anastomosing reaches of the Attawapiskat River, James Bay, ON: Sedimentary Geology 37: 295–320.
- Kundzewicz, Z. W., S. Kanae, S. I. Seneviratne, J. Handmer, N. Nicholls, P. Peduzzi, R. Mechler, et al. 2013. Flood risk and climate change: Global and regional perspectives. Hydrological Sciences Journal 59: 1–28.
- Kurtenbach, A., S. Moller, A. Krein, and W. Symader. 2006. On the relationship between hydrographs and chemographs. Hydrological Processes 20: 2921–2934.
- Lake, P. S. 2000. Disturbance, patchiness and diversity in streams. Journal of the North American Benthological Society 19(4): 573–592.
- Lancaster, J. 2000. Geometric scaling of microhabitat patches and the efficacy as refugia during disturbance. Journal of Animal Ecology 69: 442–457.
- Lapointe, M., B. Eaton, S. Driscoll, and C. Latulippe. 1999. Modelling the probability of salmonid egg pocket scour due to floods. Canadian Journal of Fisheries and Aquatic Sciences 57: 1120–1130.
- Larsen, L. G., J. W. Harvey, and M. M. Maglio. 2015. Mechanisms of nutrient retention and its relation to flow connectivity in river-floodplain corridors. Freshwater Science 34(1): 187–205.
- Layzer, J. B., T. J. Nehus, W. Pennington, J. A. Gore, and J. M. Nestler. 1989. Seasonal variation in the composition of drift below a peaking hydroelectric project. Regulated Rivers: Research and Management 3(1): 29–34.
- Leigh, C., and F. Sheldon. 2008. Hydrological changes and ecological impacts associated with water resource development in large floodplain rivers in the Australian tropics. River Research and Applications 241: 251–1270.
- Leopold, L. B. 1994. A view of the river. Cambridge, MA: Harvard University Press.
- Lesack, L. F. W., R. E. Hecky, and P. Marsh. 1991. The influence of frequency and duration of flooding on the nutrient chemistry of Mackenzie Delta lakes. In Mackenzie Delta: Environmental interactions and implications of development, NHRI Symposium No. 4, ed. P. Marsh and C. S. L. Ommanney, 19–36. Saskatoon: National Hydrology Research Institute, Environment Canada.
- Lesack, L. F. W., and P. Marsh. 2010. River-to-lake connectivities, water renewal, and aquatic habitat diversity in the Mackenzie River Delta. Water Resources Research 46: W12504. doi:10.1029/2010WR009607.
- Linnansaari, T., W. A. Monk, D. J. Baird, and R. A. Curry. 2013. Review of approaches and methods to assess environmental flows across Canada and internationally. Canadian Science Advisory Secretariat document 2012-039, vi + 73 pp.
- Lorang, M. S., and F. R. Hauer. 2003. Flow competence and streambed stability: An evaluation of technique and application. Journal of the North American Benthological Society 22(4): 475–491.
- MacKay, D. K., D. A. Sherstone, and K. C. Arnold. 1974. Channel ice effects and surface water velocities from aerial photography of Mackenzie River break-up. In Hydrological aspects of northern pipeline development. Task Force on Northern Oil Development, Environmental–Social Program, Northern Pipelines, Report 74-12, 71–107. Ottawa: Information Canada.
- Mackay, R. 1992. Colonization of lotic macroinvertebrates: A review of processes and patterns. Canadian Journal of Fisheries and Aquatic Sciences 49(3): 617–628.
- Mackenzie River Basin Committee (MRBC). 1981. Alluvial ecosystems. Mackenzie River Basin Study Supplement 2. MRBC, Governments of Canada, Alberta, British Columbia, Saskatchewan, Northwest Territories and Yukon Territory, 129 pp. Ottawa, Canada.
- Mailhot, A., I. Beauregard, G. Talbot, and S. Biner. 2012. Future changes in intense precipitation over Canada assessed from multi-model NARCCAP ensemble simulations. International Journal of Climatology 32: 1151–1163.
- Mamquam River Floodplain Restoration Project (MRFRP). 2015. Background information: The Mamquam River Floodplain Restoration Project. http://squamish.ca/assets/PDF/3.14.7-Fisheries-Project-Background-Information.pdf (accessed June, 2015).
- Marsh, P., and L. F. W. Lesack. 1996. The hydrologic regime of perched lakes in the Mackenzie Delta: Potential responses to climate change. Limnology and Oceanography 41: 849–856.
- Matthaei, C. D., K. A. Peacock, and C. R. Townsend. 1999. Scour and fill patterns in a New Zealand stream and potential implications for invertebrate refugia. Freshwater Biology 42(1): 41–57.
- Matthews, W. J., E. Marsh-Matthews, R. C. Cashner, and F. Gelwick. 2013. Disturbance and trajectory of change in a stream fish community over four decades. Oecologia 173 (3): 955–969.
- McMullen, L. E., and D. A. Lytle. 2012. Quantifying invertebrate resistance to floods: A global-scale meta-analysis. Ecological Applications 22(8): 2164–2175.
- Meehl, G. A., T. F. Stocker, W. D. Collins, P. Friedlingstein, A. T. Gaye, J. M. Gregory, and A. Kitoh. 2007. Global climate projections. Climate Change 3495: 747–845.
- Milburn, D., and T. D. Prowse. 1998. Observations on the role of an ice cover in sediment deposition in a northern delta. In Ice in surface waters, Proceedings of the 14th IAHR Ice Symposium, ed. H. T. Shen, 189–196. Potsdam, New York, Rotterdam: A. A. Balkema.
- Milburn, D., and T. D. Prowse. 2000. Observations on some physical–chemical characteristics of ice breakup. Journal of Cold Regions Engineering 14: 214–223.
- Milly, P. C. D., R. T. Wetherald, K. A. Dunne, and T. L. Delworth. 2002. Increasing risk of great floods in a changing climate. Nature 415: 514–517.
- Milner, A. M., A. L. Robertson, M. J. McDermott, M. J. Klaar, and L. E. Brown. 2013. Major flood disturbance alters river ecosystem evolution. Nature Climate Change 3(2): 137–141.
- Molles, M. C., C. S. Crawford, L. M. Ellis, H. M. Valett, and C. N. Dahm. 1998. Managed flooding for riparian ecosystem restoration. BioScience 48: 749–756.
- Monk, W. A., D. L. Peters, R. A. Curry, and D. J. Baird. 2011. Quantifying trends in indicator hydroecological variables for regime-based groups of Canadian rivers. Hydrological Processes 25: 3086–3100.
- Morse, B., and F. Hicks. 2005. Advances in river ice hydrology 1999–2003. Hydrological Processes 19: 247–263.
- Müller, K., H. Mendl, M. Dahl, and A. Müller-Haeckel. 1976. Biotopwahl und colonization cycle von plecopteren in einem subarktischen gewässersystem. Entomologisk Tidskrift 97: 1–6.
- Murchie, K. J., and K. E. Smokorowski. 2004. Relative activity of brook trout and walleye in response to flow in a regulated river. North American Journal of Fisheries Management 24(5): 1050–1057.
- Murchie, K. J. P. E. Hair, C. E. Pullen, T. D. Redpath, H. R. Stephens, and S. J. Cooke. 2008 Fish response to modified flow regimes in regulated rivers: Research methods, effects and opportunities. River Research and Applications 24(2): 197–217.
- Naiman, R. J., H. Décamps, and M. E. McClain. 2005. Riparia: Ecology, conservation, and management of streamside communities. Amsterdam: Elsevier.
- Needham, P. R., and A. C. Jones. 1959. Flow, temperature, solar radiation, and ice in relation to activities of fishes in Sagehen Creek, California. Ecology 40: 465–474.
- Nichols, S. J., D. W. Schloesser, and P. L. Hudson. 1989. Submersed macrophyte communities before and after an episodic ice jam in the St. Clair and Detroit rivers. Canadian Journal of Botany 67: 2364–2370.
- Okanagan Basin Water Board (OBWB). 2015. Okanagan River Restoration Initiative. http://www.obwb.ca/wcqi_project/okanagan-river-restoration-initiative-3-grants/ (accessed June, 2015).
- Ollero, A. 2010. Channel changes and floodplain management in the meandering middle Ebro River, Spain. Geomorphology 117: 247–260.
- Olsen, D. A., and C. R. Townsend. 2005. Flood effects on invertebrates, sediments and particulate organic matter in the hyporheic zone of a gravel-bed stream. Freshwater Biology 50(5): 839–853.
- Palmer, M. A., P. Arensburger, P. S. Botts, C. C. Hakenkamp, and J. W. Reid. 1995. Disturbance and the community structure of stream invertebrates: Patch-specific effects and the role of refugia. Freshwater Biology 34: 343–356.
- Pearce, C. M., D. McLennan, and L. D. Cordes. 1988. The evolution and maintenance of white spruce woodlands on the Mackenzie Delta, NWT, Canada. Holarctic Ecology 11: 248–258.
- Pearsons, T. N., H. W. Li, and G. A. Lamberti. 1992. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Transactions of the American Fisheries Society 121(4): 427–436.
- Pelletier, E., B. Deflandre, C. Nozais, G. Tita, G. Desrosiers, J.-P. Gagné, and A. Mucci. 1999. Crue éclair de juillet 1996 dans la région du Saguenay (Québec). 2. Impact sur les sédiment et le biote de la baie des Ha! Ha! et du fjord du Sagnenay. Canadian Journal of Fisheries and Aquatic Sciences 56: 2136–2147.
- Peters, D. L., W. A. Monk, and D. J. Baird. 2014a. Cold-regions hydrological indicators of change (CHIC) for ecological flow assessment. Hydrological Sciences Journal 59: 502–516.
- Peters D. L., R. R. Shrestha, M. A. Schnorbus, W. A. Monk, and D. J. Baird. 2014b. Ecological flows in a changing climate: Case study of the Peace River. Paper presented at the Paper presented at the 2014 Joint Scientific Congress of the Canadian Geophysical Union and Canadian Soil Society, Banff Alberta, Canada, 4–7 May 2014.
- Peters, D. L., D. J. Baird, W. A. Monk, and D. G. Armanini. 2012. Establishing standards and assessment criteria for ecological instream flow needs in agricultural regions of Canada. Journal of Environmental Quality 41: 41–51.
- Peters, D. L., T. D. Prowse, A. Pietroniro, and R. Leconte. 2006. Flood hydrology of the Peace-Athabasca Delta, northern Canada. Hydrological Processes 20: 4073–4096.
- Peterson, R. H., and J. L. Metcalfe. 1981. Emergence of Atlantic salmon fry from gravels of varying composition: A laboratory study. Canadian Technical Report Fisheries Aquatic Sciences. 1020. St. Andrews, NB: Department of Fisheries and Oceans, Biological Station, 15 pp.
- Piegay, H., S. E. Darby, E. Mosselman, and N. Surian. 2005. A review of techniques available for delimiting the erodible river corridor: A sustainable approach to managing bank erosion. River Research and Applications 21: 773–789.
- Pinter, N., and R. A. Heine. 2005. Hydrodynamic and morphodynamic response to river engineering documented by fixed-discharge analysis, Lower Missouri River, USA. Journal of Hydrology 302: 70–91.
- Poff, N. L. 1997. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. Journal of the North American Benthological Society 16: 391–409.
- Poff, N. L., and J. V. Ward. 1989. Implications of streamflow variability and predictability for lotic community structure: A regional analysis of streamflow patterns. Canadian Journal of Fisheries and Aquatic Sciences 46: 1805–1818.
- Poff, N. L., J. D. Allan, M. B. Bain, J. R. Karr, K. L. Prestegaard, B. D. Richter, R. E. Sparks, and J. C. Stromberg. 1997. The natural flow regime: A paradigm for river conservation and restoration. BioScience 47(11): 769–784.
- Polzin, M. L., and S. B. Rood. 2006. Effective disturbance: Seedling safe sites and patch recruitment of riparian cottonwoods after a major flood of a mountain river. Wetlands 26: 965–980.
- Poor, C. J., and J. J. McDonnell. 2007. The effects of land use on stream nitrate dynamics. Journal of Hydrology 332: 54–68.
- Pringle, C. M. 2001. Hydrologic connectivity and the management of biological reserves: A global perspective. Ecological Applications 11: 981–998.
- Prowse, T. D. 2001a. River-ice ecology. I: Hydrologic, geomorphic, and water-quality aspects. Journal of Cold Regions Engineering 15(1): 1–16.
- Prowse, T. D. 2001b. River-ice ecology. II: Biological aspects. Journal of Cold Regions Engineering 15 (1): 17–33.
- Prowse, T. D., and F. M. Conly. 2002. A review of hydro-ecological studies of the Northern River Basins Study, Canada: Part 2, Peace-Athabasca Delta. River Research Application 18: 447–460.
- Prowse, T. D., and J. M. Culp. 2003. Ice breakup: A neglected factor in river ecology. Canadian Journal of Civil Engineering 30(1): 128–144.
- Prowse, T. D., S. Beltaos, J. T. Gardner, J. J. Gibson, R. J. Granger, D. L. Peters, A. Pietroniro, and L. Romolo. 2006. Climate change, flow regulation, and land-use effects on the hydrology of the Peace–Athabasca–Slave system; finding from the Northern Rivers Ecosystem Initiative. Journal of Environmental Monitoring and Assessment 113: 167–197.
- Reich, P., and P. S. Lake. 2014. Extreme hydrological events and the ecological restoration of flowing waters. Freshwater Biology. doi:10.1111/fwb.12508.
- Rice, S. P., J. Lancaster, and P. Kemp. 2010. Experimentation at the interface of fluvial geomorphology, stream ecology and hydraulic engineering and the development of an effective, interdisciplinary river science. Earth Surface Processes and Landforms 35: 64–77.
- Richter, B. D., J. V. Baumgartner, J. Powell, and D. P. Braun. 1996. A method for assessing hydrological alteration within ecosystems. Conservation Biology 10: 1163–1174.
- Rios-Touma, B., N. Prat, and A. C. Encalada. 2012. Invertebrate drift and colonization processes in a tropical Andean stream. Aquatic Biology 14(3): 233–246.
- Roach, K. A. 2013. Environmental factors affecting incorporation of terrestrial material into large river food webs. Freshwater Science 32(1): 283–298.
- Robertson, M. J., D. A. Scruton, R. S. Gregory, and K. D. Clarke. 2006. Effect of suspended sediment on freshwater fish and fish habitat. Canadian Technical Report of Fisheries and Aquatic Sciences, 2644. v + 37 pp.
- Robinson, C. T. 2012. Long-term changes in community assembly, resistance, and resilience following experimental floods. Ecological Applications 22(7): 1949–1961.
- Rood, S. B., L. A. Goater, J. M. Mahoney, C. M. Pearce, and D. G. Smith. 2007. Flood, fire, and ice: Disturbance ecology of riparian cottonwoods. Canadian Journal of Botany 85: 1019–1032.
- Rood, S. B., S. Kaluthota, K. M. Gill, E. J. Hillman, S. G. Woodman, D. W. Pearce, and J. M. Mahoney. 2015. A twofold strategy for riparian restoration: Combining a functional flow regime and direct seeding to re-establish cottonwoods. River Research and Applications. doi:10.1002/rra.2919.
- Rood, S. B., J. Pan, K. M. Gill, C. G. Franks, G. M. Samuelson, and A. Shepherd. 2008. Declining summer flows of Rocky Mountain rivers: Historic hydrology and probable impacts on floodplain forests. Journal of Hydrology 349: 397–410.
- Rood, S. B., G. M. Samuelson, J. H. Braatne, C. R. Gourley, F. M. R. Hughes, and J. M. Mahoney. 2005. Managing river flows to restore floodplain forests. Frontiers in Ecology and the Environment 3: 193–201.
- Samuelson, G. M., and S. B. Rood. 2004. Differing influences of natural and artificial disturbances on riparian cottonwoods from prairie to mountain ecoregions in Alberta, Canada. Journal of Biogeography 31: 435–450.
- Saunders, J. W., and M. W. Smith. 1965. Changes in a stream population of trout associated with increased silt. Journal of Fisheries Research Board of Canada 22(2): 395–404.
- Sawada, M., and P. G. Johnson. 2000. Hydrometeorology, suspended sediment and conductivity in a large glacierized basin, Slims River, Yukon Territory, Canada (1993–94). Arctic 53(2): 101–117.
- Scharbert, A., and J. Borcherding. 2013. Relationships of hydrology and life-history strategies on the spatio-temporal habitat utilisation of fish in European temperate river floodplains. Ecological Indicators 29: 348–360.
- Schwartz, J. S., and E. E. Herricks. 2005. Fish use of stage-specific fluvial habitats as refuge patches during a flood in a low-gradient Illinois stream. Canadian Journal of Fisheries and Aquatic Sciences 62(7): 1540–1552.
- Scott, M. L., G. T. Auble, and J. M. Friedman. 1997. Flood dependency of cottonwood establishment along the Missouri River, Montana, USA. Ecological Applications 7: 677–690.
- Scott, M. L., J. M. Friedman, and G. T. Auble. 1996. Fluvial process and the establishment of bottomland trees. Geomorphology 14: 327–339.
- Scrimgeour, G. J., T. D. Prowse, J. M. Culp, and P. A. Chambers. 1994. Ecological effects of river ice break-up: A review and perspective. Freshwater Biology 32: 261–275.
- Scruton, D. A., C. J. Pennell, M. J. Robertson, L. M. N. Ollerhead, K. D. Clarke, K. Alfredsen, A. Harby, and R. S. Mckinley. 2005. Seasonal response of juvenile Atlantic salmon to experimental hydropeaking power generation in Newfoundland, Canada. North American Journal of Fisheries Management 25: 964–974.
- Smith, D. G. 1979. Effects of channel enlargement by river ice processes on bankfull discharge in Alberta, Canada. Water Resources Research 15: 469–475.
- Sokal, M. A., R. I. Hall, and B. B. Wolfe. 2008. Relationships between hydrological and limnological conditions in lakes of the Slave River Delta (NWT, Canada) and quantification of their roles on sedimentary diatom assemblages. Journal of Paleolimnology 39: 533–550.
- Sokal, M. A., R. I. Hall, and B. B. Wolfe. 2010. The role of flooding on interannual and seasonal variability of lake water chemistry, phytoplankton diatom communities and macrophyte biomass in the Slave River Delta (Northwest Territories, Canada). Ecohydrology 3: 41–54.
- Squires, M. M., and L. F. W. Lesack. 2002. Water transparency and nutrients as controls on phytoplankton along a flood-frequency gradient among lakes of the Mackenzie Delta, western Canadian Arctic. Canadian Journal of Fisheries and Aquatic Sciences 59: 1339–1349.
- Squires, M. M., and L. F. W. Lesack. 2003. The relation between sediment nutrient content and macrophyte biomass and community structure along a water transparency gradient among lakes of the Mackenzie Delta. Canadian Journal of Fisheries and Aquatic Sciences 60: 333–343.
- Squires, M. M., L. F. W. Lesack, and D. Huebert. 2002. The influence of water transparency on the distribution and abundance of macrophytes among lakes of the Mackenzie Delta, Western Canadian Arctic. Freshwater Biology 47: 2123–2135.
- Squires, M. M., L. F. W. Lesack, R. E. Hecky, S. J. Guildford, P. Ramlal, and S. N. Higgins. 2009. Primary production and carbon dioxide metabolic balance of a lake-rich arctic river floodplain: Partitioning of phytoplankton, epipelon, macrophyte, and epiphyton production among lakes on the Mackenzie Delta. Ecosystems 12: 853–872.
- Stanford, J. A., M. S. Lorang, and F. R. Hauer. 2005. The shifting habitat mosaic of river ecosystems. Internationalen Vereinigung für Theoretische und Angewandte Limnologie Verhandlungen 29: 123–136.
- Stewart-Schumm, S. A. 1985. Patterns of alluvial rivers. Annual Review of Earth and Planetary Sciences 13: 5.
- Stover, S. C., and D. R. Montgomery. 2001. Channel change and flooding, Skokomish River, Washington. Journal of Hydrology 243: 272–286.
- Stromberg, J. C., V. B. Beauchamp, M. D. Dixon, S. J. Lite, and C. Paradzick. 2007. Importance of low-flow and high-flow characteristics to restoration of riparian vegetation along rivers in arid south-western United States. Freshwater Biology 52: 651–679.
- Sueyoshi, M., D. Nakano, and F. Nakamura. 2014. The relative contributions of refugium types to the persistence of benthic invertebrates in a seasonal snowmelt flood. Freshwater Biology 59: 257–271.
- Tabacchi, É., D. L. Correll, R. Hauer, G. Pinay, A.-M. Planty-Tabacchi, and R. Wissmar. 1998. Development, maintenance and role of riparian vegetation in the river landscape. Freshwater Biology 40: 497–516.
- Tennant, D. L. 1976. Instream flow regimens for fish, wildlife, recreation and related environmental resources. Fisheries 1: 6–10.
- Thomaz, S. M., L. M. Bini, and R. L. Bozelli. 2007. Floods increase similarity among aquatic habitats in river–floodplain systems. Hydrobiologia 579: 1–13.
- Tilsworth, T., and P. L. Bateman. 1982. Changes in Chena River water quality. The Northern Engineer 14: 29–37.
- Timoney, K. P. 2013. The Peace-Athabasca Delta – Portrait of a dynamic ecosystem, Edmonton: The University of Alberta Press, 95 pp.
- Tockner, K., F. Malard, and J. V. Ward. 2000. An extension of the flood pulse concept. Hydrological Processes 14: 2861–2883.
- Trigg, M. A., K. Michaelides, J. C. Neal, and P. D. Bates. 2013. Surface water connectivity dynamics of a large-scale extreme flood. Journal of Hydrology 505: 138–149.
- Tronstad, L. M., B. P. Tronstad, and A. C. Benke. 2007. Aerial colonization and growth: Rapid invertebrate responses to temporary aquatic habitats in a river floodplain. Journal of the North American Benthological Society 2(3): 460–471.
- Usseglio-Polatera, P., and H. Tachet. 1994. Theoretical habitat templets, species traits, and species richness: Plecoptera and Ephemeroptera in the Upper Rhone River and its Floodplain. Freshwater Biology 31(3): 357–375.
- van der Valk, A. G., and L. C. Bliss. 1971. Hydrarch succession and net primary production of oxbow lakes in central Alberta. Canadian Journal of Botany 49: 1177–1199.
- Vegas-Vilarrubia, T., and R. Herrera. 1993. Effects of periodic flooding on the water chemistry and primary production of the Mapire systems (Venezuela). Hydrobiologia 262: 31–42.
- von de Wall, S. 2011. An assessment of the river ice break-up season in Canada. MSc thesis, Department of Geography, University of Victoria, 84 pp.
- Wantzen, K. M., W. J. Junk, and K. Rothhaupt. 2008. An extension of the floodpulse concept (FPC) for lakes. Hydrobiologia 613: 151–170.
- Ward, J. V. 1989. The four dimensional nature of lotic ecosystems. Journal of the North American Benthological Society 8: 2–8.
- Ward, J. V., and J. A. Stanford. 1983. The intermediate disturbance hypothesis: An explanation for biotic diversity in lotic ecosystems. In dynamics of lotic ecosystems, ed. T. D. Fontaine and S. M. Bartell, 347–356. Ann Arbor, MI: Ann Arbor Science Publishers.
- Ward, J. V., and J. A. Stanford. 1989. Riverine ecosystems: The influence of man on the catchment dynamics and fish ecology. Canadian Special Publication Fish and Aquatic Sciences 106: 56–64.
- Warren, F. J., and P. A. Egginton. 2008. Background information. In From impacts to adaptation: Canada in a changing climate 2007, ed. D. S. Lemmen, F. J. Warren, J. Lacroix and E. Bush, 27–56. Ottawa: Government of Canada.
- Waters, T. F. 1995. Sediment in streams: Sources, biological effects, and control., Bethesda, MD: American Fisheries Society, Monograph 7, 251 pp.
- Watt, W. D. 1987. A summary of the impact of acid rain on Atlantic Salmon (Salmo salar) in Canada. Water, Air and Soil Pollution 35: 27–35.
- Watt, W. E. 1990. Hydrology of floods in Canada: A guide to planning and design. Associate Committee on Hydrology: National Research Council Canada. 245 pp. Ottawa, ON.
- Weller, M. W. 1987. Freshwater marshes: Ecology and wildlife management. Minneapolis, MN: University of Minnesota Press.
- Whited, D. C., M. S. Lorang, M. J. Harner, F. R. Hauer, J. S. Kimball, and J. A. Stanford. 2007. Climate, hydrologic disturbance, and succession: Drivers of floodplain pattern. Ecology 88(4): 940–953.
- Wiklund, J. A., R. I. Hall, and B. B. Wolfe. 2012. Timescales of hydrolimnological change in floodplain lakes of the Peace-Athabasca Delta, northern Alberta, Canada. Ecohydrology 5: 351–367.
- Wilding, T. K., J. S. Sanderson, D. M. Merritt, S. B. Rood, and N. L. Poff. 2014. Riparian responses to reduced flood flows: Comparing and contrasting narrowleaf and broadleaf cottonwoods. Hydrological Sciences Journal 59: 605–617.
- Wohl, W., B. P. Bledsoe, R. B. Jacobson, N. L. Poff, S. L. Rathburn, D. M. Walters, and A. C. Wilcox. 2015. The natural sediment regime in rivers: Broadening the foundation for ecosystem management. BioScience. doi:10.1093/biosci/biv002.
- Wolfe, B. B., T. L. Karst-Riddoch, R. I. Hall, and T. W. D. Edwards. 2007. Classification of hydrological regimes of northern floodplain basins (Peace–Athabasca Delta, Canada) from analysis of stable isotopes (δ18O, δ2H) and water chemistry. Hydrological Processes 21: 151–168.
- Wolfe, B. B., R. I. Hall, T. W. D. Edwards, S. R. Vardy, M. D. Falcone, C. Sjunneskog, R. Sylvestre, S. McGowan, R. R. Leavit, and P. van Driel. 2008. Hydroecological responses of the Athabasca Delta, Canada, to changes in river flow and climate during the 20th century. Ecohydrology 1: 131–148.
- Zhang, X., L. A. Vincent, W. D. Hogg, and A. Niitsoo. 2000. Temperature and precipitation trends in Canada during the 20th century. Atmosphere-Ocean 38(3): 395–429.
- Zimmermann, E. M., and R. G. Death. 2002. Effect of substrate stability and canopy cover on stream invertebrate community. New Zealand Journal of Marine and Freshwater Research 36: 537–545.