Abstract
Nine isolates of Alternaria causing black rot of ‘Fortune’ mandarin were obtained from a commercial orchard in the Murcia region of south-east Spain. The isolates, according to their morphological and physiological characteristics, were identified as A. alternata. Analysis of DNA sequences of the internal transcribed spacers 1 (ITS1) and 2 (ITS2) of the ribosomal DNA corroborated the morphological identification. In this work, we evaluated the pathogenicity of each of the nine isolates on fruits and leaves as well as fruit susceptibility to the pathogen at different growth stages. Studies were performed on both wounded and unwounded fruits and leaves. Disease development was faster in wounded than in unwounded organs, one and two days after inoculation, respectively. Differences in pathogenicity were observed among the A. alternata isolates, depending on the plant organ. Immature fruits were more susceptible; fruit infection tolerance increased with maturity, for both wounded and unwounded fruits. These findings could contribute to more-successful phytosanitary management of the disease.
Résumé
Neuf isolats d'Alternaria ont été obtenus à partir de feuilles et fruits affectés par la tache noire d'une plantation commerciale de mandarine ‘Fortune’, dans la région de Murcie (au sud-est de l'Espagne). Ces isolats, d'après ces caractéristiques morphologiques y physiologiques, ont été identifiés comme A. alternata. L'analyse des séquences ITS1 et ITS 2 (internal transcribed spacers) de l'ADN ribosomale a confirmé leur identification morphologique. Ce travail a évalué: a) la pathogénicité de chacun des neuf isolats dans les feuilles et les fruits, et b) la susceptibilité des fruits dans des différents stades de développement. Les études ont été réalisées sur des feuilles et des fruits blessés et non blessés. Le développement de la lésion a évolué plus rapidement dans les organes végétaux blessés que dans les non blessés; un et deux jours plus tard après l'inoculation, respectivement. On a observé aussi que la pathogénicité des isolats dépend de l'organe dans lequel se développe. Par ailleurs, les fruits immatures sont plus sensibles à l'A. alternata puis la tolérance à la maladie augmente avec la maturité du fruit, aussi bien pour les fruits blessés que pour les non blessés. Ces résultats peuvent contribuer à améliorer le traitement de la maladie dans les champs.
Introduction
Members of the genus Alternaria are responsible for substantial losses in Citrus, especially in pre-harvest fruits. Alternaria brown spot, caused by Alternaria alternate (Fr.) Keissl. is an important disease of tangerines and their hybrids, affecting leaves, twigs and immature fruit (Pegg, Citation1966; Canihos et al., Citation1999). Several studies have revealed that, in the process of natural infection, the susceptibility to A. alternata depends on the citrus species. Among the most-affected cultivated varieties are hybrids of the mandarin ‘Dancy’, particularly ‘Fortune’, although other cultivars for which ‘Dancy’ is a direct or indirect parent, such as the tangelo ‘Minneola’ or the mandarin ‘Nova’, are also affected. Several varieties of grapefruit, as well as mandarin ‘Emperor’, and the hybrids ‘Murcott’, ‘Orlando’, ‘Fairchild’, and ‘Page’ are also affected. Satsumas and clementines are slightly affected while lemon and lime cultivars are not affected (Pegg, Citation1966; Gardner et al., Citation1986; Kohmoto et al., Citation1991; Reis et al., Citation2007). Previous work (Ortuño et al., Citation2008) described how a complex signalling pathway may be involved in the differences observed in the defence mechanism of Citrus against A. alternata; in addition to different secondary metabolites, the ethylene produced during infection probably participates in this.
The objectives of this work were (i) to determine the pathogenicity of nine isolates of A. alternata on both fruits and leaves of ‘Fortune’ mandarin, by simulating two field scenarios, namely intact and injured surfaces (wounded/unwounded); and (ii) to ascertain the susceptibility of ‘Fortune’ mandarin fruits to A. alternata at different stages of their development. The results will aid in the reduction of pathogen selection pressure and will allow more successful management of the disease.
Materials and methods
Plant material
‘Fortune’ mandarin leaves (40–55 mm wide) and fruits (80–100 mm long) at development stages 16–18 and 78–79, respectively (Finn et al., Citation2007), were used for evaluation of isolate pathogenicity. To study their susceptibility to A. alternata, fruits were picked at different development stages, being 24–60 mm in diameter (90 to 360 days after anthesis). The leaves and fruits were collected in a section of a commercial orchard, without pesticide pressure, situated in Campo de Cartagena (Murcia, Spain).
Isolation of the fungus
Nine isolates of Alternaria were obtained from the ‘Fortune’ mandarin orchard, between March 2006 and October 2007. Six (Alt1, Alt3, Alt5, Alt6, Alt10 and Alt12) were obtained from leaves and the others (Alt13, Alt14 and Alt15) from fruits. Leaves and pieces of fruits showing Alternaria brown spot symptoms were placed in Petri dishes with PDA (potato dextrose agar) medium and incubated at 25 ± 1 °C, in the dark.
Colonies were transferred to new Petri dishes with PDA and monosporic cultures were obtained for each of the nine isolates through the following procedure: one drop of conidial suspension (1 × 106 mL−1) was spread on a Petri dish with PDA and incubated under the above-described conditions. After 24 h, germinating spores were separated under a stereoscopic microscope, placed individually in Petri dishes with PDA, and incubated under the same previously described conditions. Seven-day cultures, obtained by monosporic production, were used in all studies.
Molecular characterization
Genomic DNA was extracted from each isolate using a modified Chelex method (Walsh et al., Citation1991). Briefly, this consisted of crushing fungal mycelia taken from the surface of PDA dishes in 40 μL of stirred 5% Chelex solution (BioRad). This homogenate was incubated at 54 °C for 15 min and at 99 °C for 3 min. After incubation at –20 °C for 10 min, the suspension was centrifuged at 13 200 rpm for 10 min. The mycelial debris was discarded and the DNA was stored at –20 °C, if not used immediately.
Analysis of rDNA ITS
The PCR primers employed to amplify the internal transcribed spacers of rDNA were ITS 5 (GGAAGTAAAAGTCGTAACAAGG) and ITS 4 (TCCTCCGCTTATTGATATGC) (White et al., Citation1990). These primers amplify the partial end of the 18S rRNA gene, the entire ITS1, the 5.8S rRNA gene, ITS2 and the initiation of the 28S rRNA gene. The PCR assays were conducted using 2.0 μL of template DNA (50 ng μL−1), 0.3 μL of Ecotaq polymerase 5U μL−1 (Ecogen), and 1.2 μL of each primer (10 μM), in a total reaction volume of 25 μL. The PCR conditions were an initial 5 min at 94 °C, followed by 30 s at 94 °C, 1 min at 56 °C, and 30 s at 72 °C for 35 cycles, with a final 7 min at 72 °C (GeneAmp PCR System 2400 Perkin Elmer Applied Biosystems, Massachusetts, USA). The DNA fragments of ≈ 600 bp were purified using a QIAquick Gel Extraction Kit (Qiagen) and then cloned using the TOPO TA Cloning Kit for Sequencing (Invitrogen), following the manufacturers' protocols. Plasmids were purified using the PureLink Quick Plasmid Miniprep Kit (Invitrogen) and two individual clones per isolate were chosen for sequencing in both directions, at SECUGEN SL (Servicio de Secuenciación, Centro de Investigaciones Biológicas, CIB, Madrid, Spain).
Phylogenetic analysis
Phylogenetic and molecular evolutionary analyses were conducted using MEGA version 4.0 (Tamura et al., Citation2007). Multiple sequence alignments were performed with CLUSTAL W (Thompson et al., Citation1994), available at http://www.ebi.ac.uk/clustalw/. The BLAST program was used to search for homologous Alternaria spp. sequences in the NCBI databases (http://www.ncbi.nlm.nih.gov/). The phylogenetic tree was inferred using the Neighbour-Joining method (Saitou & Nei, Citation1987) and the Kimura 2-parameters model. The consensus tree was inferred from 2000 bootstrap replicates.
Pathogenicity studies
Fungal pathogenicity was evaluated by measuring the growth of lesions produced after inoculation of leaves and fruits with the isolates. Prior to inoculation, fruits and leaves were immersed in 0.5% NaClO for 5 min. Then, they were washed with sterile water and dried with sterile cellulose paper. The inoculation procedure was as follows: small discs (4 mm diameter) were cut from 7-day-old PDA cultures of the nine isolates of Alternaria. One disc was inverted onto the surface of a fruit or leaf, held in place by Parafilm ‘M’ plastic film (Pechiney Plastic Packaging, Chicago), and placed in a sterile tray (34 × 24 cm). Each tray was bagged up (using a disinfected, 4-mm-thick polyethylene bag) and incubated at 24 ± 1 °C, with a 14-h photoperiod.
Twenty fruits and leaves per isolate were inoculated. Ten of them were inoculated directly, on an intact surface, and the others after wounding (3–4 mm deep) with a sterile scalpel. The incubation period was 7 d for leaves and 25 d for fruits. Periodic observations were made during these periods; each time, the growth of the lesions was measured. Non-inoculated fruits and leaves, either with or without damage, were used as controls: they were sprayed with sterile, distilled water. The inoculations were performed under sterile conditions, in a laminar flow cabinet.
Susceptibility of fruit studies
Fruit susceptibility to A. alternata at different growth stages was evaluated by measuring lesion growth after inoculation. Fruits were picked from July 2006 (24-mm diameter) to April 2007 (60-mm diameter) and inoculated with the Alt6 isolate, because of its high and homogeneous pathogenicity on fruits compared with the other isolates. The inoculations were carried out on wounded and unwounded fruits, 10 fruits in each case. The inoculation procedure and the period and conditions of incubation were as described above.
Statistical analysis
The values are given as means ± standard error. The data were analysed by a General Linear Model (GLM). Duncan's Multiple Range Test (DMRT) was used for the analysis of the variance in the data of – All statistical analyses were performed using Statgraphics 5.0 software.
Results and discussion
Identification of Alternaria isolates
The ITS region of the rDNA was sequenced for the nine Alternaria isolates used in our study. The length of the entire ITS1, the 5.8S rRNA gene, and ITS2 was around 550 bp. The alignment of sequences did not reveal any sequence variation; they showed complete or almost-complete homogeneity. The sequences of ITS regions obtained here were homologous to other A. alternata sequences associated with Citrus spp. in the NCBI databases: HQ846574.1, JF440581.1, JF835905.1, JN541212.1, GU073112.1, JN618076.1, JF835906.1, JF835812.1, JN107734.1 and AB667801.1.
The phylogenetic tree inferred from our ITS sequences and their homologues in the NCBI databases showed that all isolates cluster together in a single group (data not shown) but separate from Alternaria tenuissima, the species causing black rot in mandarin hybrids (Simmons, Citation1999). The high genetic homogeneity among all the sequences corroborated our morphological and physiological studies: all the isolates belonged to A. alternata.
Pathogenicity of isolates on fruits
Six of the nine isolates studied could be considered pathogens on unwounded fruits because they caused black rot symptoms (). Based on the diameters of the lesions they produced, we distinguished three groups of isolates differing in aggressiveness: low (Alt1 and Alt3), medium (Alt6 and Alt12) and high (Alt5 and Alt14). Differences in the development of the lesions caused by these isolates were observed. In , only one representative isolate per group (Alt1, Alt5 and Alt6) was considered in the analyses, for a clearer understanding. shows that the expansion of the lesions exhibited a lag during the first two days, then between days 2 and 5 it was at its greatest, with a different diameter for each isolate, and from day 5 onwards the lesion expansion slowed.
Fig. 1. Pathogenicity of Alternaria alternata isolates Alt1, Alt3, Alt5, Alt6, Alt10, Alt12, Alt13, Alt14 and Alt15 on unwounded fruits of ‘Fortune’ mandarin. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
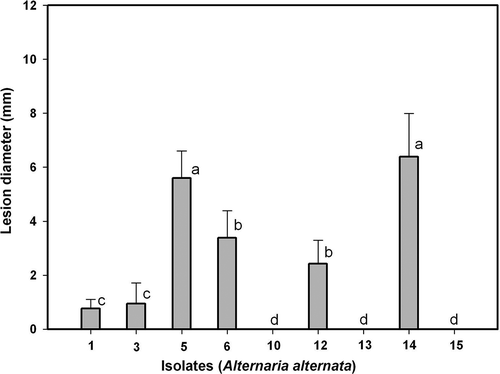
Fig. 2. Lesion development caused by three Alternaria alternata isolates on unwounded fruits of ‘Fortune’ mandarin: Alt 1(•), Alt5 (○) and Alt6 (Δ). The data represent mean values of lesion diameter (mm) at different days post-inoculation. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
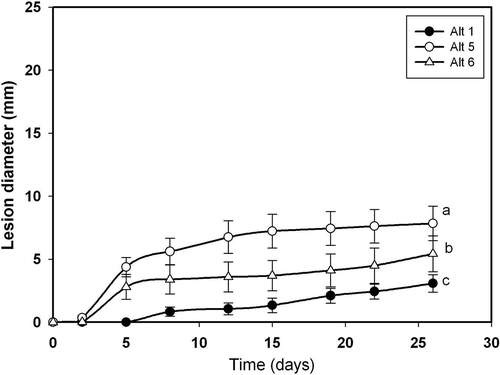
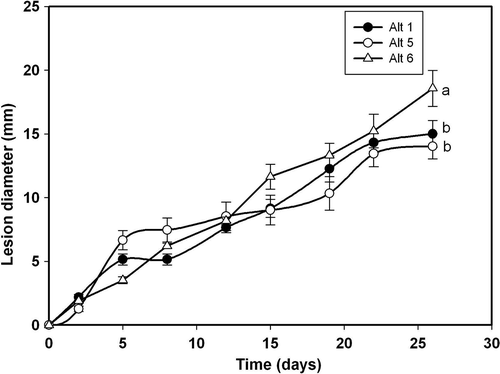
In relation to disease development on wounded fruits (), all isolates were pathogenic except Alt10, which produced lesions very reduced in size. In this respect, the pathogen isolates can be grouped as slightly aggressive (Alt15), moderately aggressive (Alt1 and Alt13), aggressive (Alt3, Alt5, Alt6 and Alt12) and highly aggressive (Alt14). To follow the development of lesions on wounded fruits, we considered the same three isolates used for unwounded fruits (Alt1, Alt5 and Alt6). Lesion diameter increased linearly with time (). In this case, there was no lag phase as described above for the unwounded fruits. Final lesion diameters were > 15 mm for isolate Alt6 and around 12 mm for isolates Alt5 and Alt1, after 25 days of incubation of wounded fruits (), while the diameters did not reach 10 mm in unwounded fruits ().
Fig. 3. Pathogenicity of Alternaria alternata isolates Alt1, Alt3, Alt5, Alt6, Alt10, Alt12, Alt13, Alt14 and Alt15 on wounded fruits of ‘Fortune’ mandarin. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
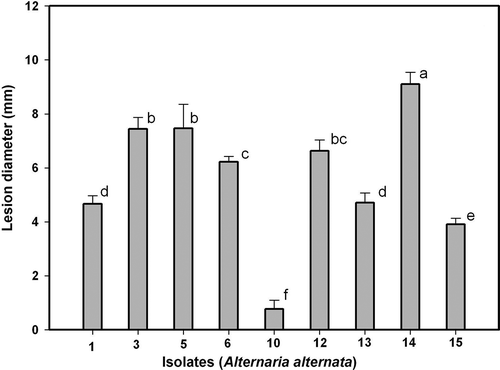
Fig. 4. Lesion development caused by three Alternaria alternata isolates on wounded fruits of ‘Fortune’ mandarin: Alt 1(•), Alt5 (○), and Alt6 (Δ). Data represent mean values of lesion diameter (mm) at different days post-inoculation. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
Pathogenicity of isolates on leaves
Only three isolates inoculated on unwounded leaves were pathogenic (); of these, Alt3 and Alt5 were aggressive and Alt1 was very aggressive. On the other hand, on wounded leaves (), isolates Alt1, Alt3, Alt5, Alt6, Alt12, Alt13, Alt14 and Alt15 can be considered pathogenic, as they produced lesions of different sizes. and show the progression of lesions in wounded and unwounded leaves: as for fruits, isolates Alt1, Alt5 and Alt6 were used to analyse the progress.
Fig. 5. Pathogenicity of Alternaria alternata isolates Alt1, Alt3, Alt5, Alt6, Alt10, Alt12, Alt13, Alt14 and Alt15 on unwounded leaves of ‘Fortune’ mandarin. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
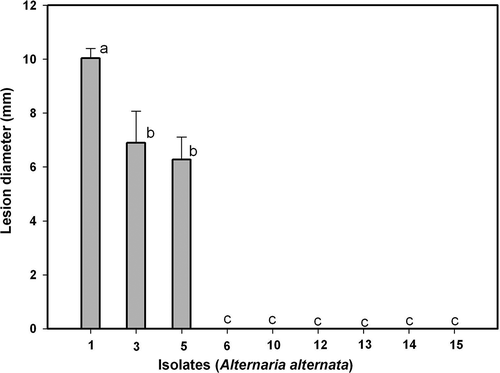
Fig. 6. Pathogenicity of Alternaria alternata isolates Alt1, Alt3, Alt5, Alt6, Alt10, Alt12, Alt13, Alt14 and Alt15 on wounded leaves of ‘Fortune’ mandarin. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
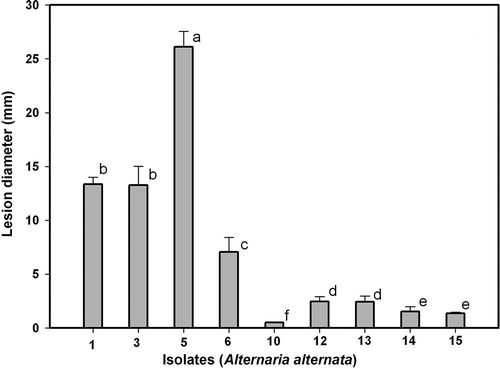
Fig. 7. Lesion development caused by three Alternaria alternata isolates on unwounded leaves of ‘Fortune’ mandarin: Alt 1(•), Alt5 (○) and Alt6 (Δ). Data represent mean values of lesion diameter (mm) at different days post-inoculation. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
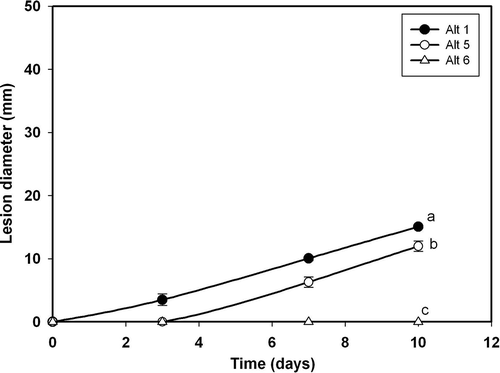
Fig. 8. Lesion development caused by three Alternaria alternata isolates on wounded leaves of ‘Fortune’ mandarin: Alt 1(•), Alt5 (○) and Alt6 (Δ). Data represent mean values of lesion diameter (mm) at different days post-inoculation. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
Isolate Alt6 did not infect or produce any necrosis on unwounded leaves (). The lesions expanded faster on wounded than on unwounded leaves ( and ). Isolate Alt6 infected and grew more slowly than the other isolates, while Alt5 was the most virulent (). Regardless of the isolate, the lesions were larger on wounded than on unwounded leaves. The diameter of the lesions increased in a curvilinear fashion on wounded leaves () and linearly on unwounded leaves ().
The differences observed among the A. alternata isolates regarding the initial disease values in fruits and leaves, which can be determined from the progress curves shown in , , and , suggest that the isolates have differing degrees of virulence. In the case of fruits, isolate Alt14 was the most virulent, followed by Alt5, Alt6 and Alt12. In all cases, the lesion diameter was greater on wounded than on unwounded fruits (see , ). Isolates Alt3 and Alt1 showed a similar lesion size (somewhat lower than for Alt14, Alt5, Alt6 and Alt12), both producing lesions of very small diameter on unwounded fruits. Isolates Alt13 and Alt15 showed a higher virulence on wounded fruits that was similar or slightly lower, and failed to initiate infection on unwounded fruits (, ). Lower disease was observed for isolate Alt10, which produced only a small lesion on wounded fruits ().
In the case of leaves, isolate Alt1 was the most virulent, followed by Alt3 and Alt5. All three produced lesions of greater diameter on wounded than on unwounded leaves (, ). A gradual reduction in disease was observed for isolates Alt6, Alt12, Alt13, Alt14 and Alt15 and none of them produced symptoms on unwounded leaves (). Lower disease was observed when the leaves were inoculated with isolate Alt10 ().
The results show clear differences in the susceptibility of fruits and leaves to the different A. alternata isolates assayed, since the most pathogenic on fruits (Alt14) was hardly pathogenic on leaves (, ). Similarly, isolate Alt1 was very pathogenic on leaves but hardly at all on fruits (, ).
Fruit susceptibility at different development stages
The mean diameter of the lesions produced by isolate Alt6, from July (90 days after anthesis) to April (360 days after anthesis) is shown in The lesion diameter decreased rapidly during July and August, coinciding with the linear phase of fruit growth, and remained constant from August to October, when the fruit reached their maximum size. Subsequently, the lesion diameter decreased gradually as the fruit reached maturity and changed colour. The same pattern was observed on wounded fruits, although the lesion diameters were greater than those of unwounded fruits (, b). From December onwards, the wounded fruit showed a high degree of resistance to this isolate. Increased tolerance of fruits to A. alternata during their advancement towards maturity has been reported previously (Pegg, Citation1966; Gardner et al., Citation1986; Reis et al., Citation2006), but resistance has not (Vicent et al., Citation2004).
Fig. 9. Diameters of the lesions caused by Alternaria alternata isolate Alt6, five days after its inoculation in unwounded (a) and wounded (b) fruits of ‘Fortune’ mandarin. Fruits were harvested from July (90 days after anthesis) to April (360 days after anthesis), covering different developmental stages. Vertical bars denote ± SE, when larger than symbols. Values not sharing a common superscript letter are significantly different (P < 0.05).
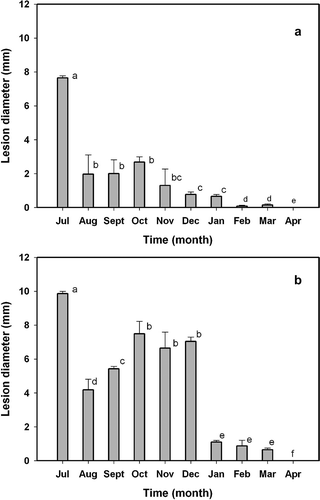
The results obtained here suggest that the degree of tolerance or susceptibility to A. alternata depends on the virulence of the fungal isolate, the presence or absence of wounds, and the stage of development of the fruits. According to our results and previous reports (Pegg, Citation1966; Gardner et al., Citation1986; Reis et al., Citation2006), it seems necessary to design control strategies to protect fruits at the beginning of development, before the change of colour, rather than at full maturity when symptoms are evident.
Acknowledgements
This research was supported by the Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (PR06‐07 and POI07‐010), the Consejería de Agricultura, Agua y Medio Ambiente de la Región de Murcia (AGR/11621/08) and the Fundación Séneca, Agencia de Ciencia y Tecnología de la Región de Murcia (08676/PI/08).
References
- Canihos , Y. , Prever , T.L. and Timmer , L.W. 1999 . Temperature, leaf wetness, and isolate effects on infection of ‘Minneola’ tangelo leaves by Alternaria sp . Plant Dis. , 83 : 429 – 433 .
- Finn , G.A. , Straszewski , A.E. and Peterson , V. 2007 . A general growth stage key for describing trees and woody plants . Ann. Appl. Biol. , 151 : 127 – 131 .
- Gardner , J.M. , Kono , Y. and Chandler , J.L. 1986 . Bioassay and host-selectivity of Alternaria citri toxins affecting rouge lemon and mandarins . Physiol. Mol. Plant Pathol. , 29 : 293 – 304 .
- Kohmoto , K. , Akimitsu , K. and Otani , H. 1991 . Correlation of resistance and susceptibility of Citrus to Alternaria alternata with sensitivity to host-specific toxins . Phytopathology , 81 : 719 – 722 .
- OrtuñO , A. , Nemsa , I. , Alvarez , N. , Lacasa , A. , Porras , I. , GarcíA-LidóN , A. and Del RíO , J.A. 2008 . Correlation of ethylene synthesis in citrus fruits and their susceptibility to Alternaria alternata pv . citri. Physiol. Mol. Plant Pathol. , 72 : 162 – 166 .
- Pegg , K.G. 1966 . Studies of a strain of Alternaria citri Pierce, the causal organism of brown spot of ‘Emperor’ mandarin . Queensland J. Agric. Anim. Sci. , 23 : 15 – 18 .
- Reis , R.F. , De Goes , A. , Mondal , S.M. and Timmer , L.W. 2006 . Effectiveness of fungicides and susceptibility of fruit and leaves of tangerine, tangor and tangelos to infection by Alternaria alternata, the cause of brown spot . Summa Phytopathol. , 32 : 11 – 12 .
- Reis , R.F. , De Almeida , T.F. , Stuchi , E.S. and De Goes , A. 2007 . Susceptibility of citrus species to Alternaria alternata, the causal agent to the Alternaria brown spot . Scient. Horticul. , 113 : 336 – 342 .
- Saitou , N. and Nei , M. 1987 . The neighbor-joining method: a new method for reconstructing phylogenetic trees . Mol. Biol. Evol. , 4 : 406 – 425 .
- Simmons , E.G. 1999 . Alternaria themes and variations (236–243). Host specific toxin producers . Mycotaxon , 70 : 325 – 369 .
- Tamura , K. , Dudley , J. , Nei , M. and Kumar , S. 2007 . MEGA 4, Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0 . Mol. Biol. Evol. , 24 : 1596 – 1599 .
- Thompson , J.D. , Higgins , D.G. and Gibson , T.J. 1994 . CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice . Nucl. Acids Res. , 22 : 4673 – 4680 .
- Vicent , A. , Badal , J. , Asensi , M.J. , Sanz , N. , Armengol , J. and GarcíA-Jiménez , J. 2004 . Laboratory evaluation of citrus cultivars susceptibility and influence of fruit size on ‘Fortune’ mandarin to infection by Alternaria alternata pv . citri. Eur. J. Plant Pathol. , 110 : 245 – 251 .
- Walsh , P.S. , Metzger , D.A. and Higuchi , R. 1991 . Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material . Biotechniques , 10 : 506 – 512 .
- White , T.J. , Bruns , T. , Lee , S. and Taylor , J. 1990 . “ Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics ” . In PCR Protocols: A Guide to Methods and Applications , Edited by: Innis , M.A. , Gelfand , D.H. , Shinsky , J.J. and White , T.J. 315 – 322 . San Diego , , Calif : Academic Press .