
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Hybrid wheat shows promise to increase yield potential, but many questions remain about the hybrid system. One of the most important is how hybrids will perform in terms of diseases such as Fusarium head blight (FHB). It is unknown if a hybrid produced from a resistant by susceptible cross will express acceptable resistance levels. This experiment tested hybrids produced from four resistant parents crossed with a common susceptible parent (2398). The results indicated that the hybrids were resistant compared with the susceptible parent and mid-parent, while they were less resistant compared with their respective resistant parent. The hybrids were between the mid-parent and resistant parent in resistance to FHB traits. This may indicate that the resistance in the hybrids may be controlled by partial dominance gene action. The findings also indicated that accumulation of more resistance genes provided improved resistance to FHB. In conclusion, successful hybrids with a good level of FHB resistance could be produced through crossing parents with contrasting resistance to this disease.
Résumé: Le blé hybride promet d’accroître le potentiel de rendement, mais plusieurs questions subsistent quant au système d’hybridation. Une des plus importantes est à savoir comment les hybrides se comporteront à l’égard de maladies telle la brûlure de l’épi causée par le fusarium (BEF). Nous ignorons si un hybride résultant du croisement résistant x réceptif affichera des taux de résistance acceptables. Cette expérience a testé des hybrides produits à partir de quatre parents résistants croisés avec un parent réceptif commun (2398). Les résultats ont indiqué que les hybrides étaient résistants, comparativement au parent et au parent moyen réceptifs, tandis qu’ils étaient moins résistants que leur parent résistant respectif. Quant à la résistance aux caractères de la BEF, les hybrides se situaient entre le parent moyen et le parent résistant. Cela peut indiquer que la résistance chez les hybrides peut être gérée par l’action génétique en dominance partielle. Les résultats ont également indiqué que l’accumulation de plusieurs gènes de résistance offrait une résistance accrue à la BEF. Pour terminer, les hybrides efficaces possédant un bon taux de résistance à la BEF pourraient être produits en croisant des parents dont la résistance à cette maladie varie.
Introduction
Wheat (Triticum aestivum L.) is one of the most important crops in the world and certainly in North Dakota. In 2018, North Dakota-planted spring wheat totalled 2.65 million hectares and was valued over 1.6 USD billion (USDA/NASS, Citation2019). One of the most limiting factors in wheat production is the spike disease Fusarium head blight, known colloquially as scab. Several Fusarium species can cause scab and the most important species in North Dakota (ND) is F. graminearum (Friskop et al. Citation2018). This disease can cause yield reduction up to 70% under conditions conducive to the disease, which are temperatures of about 25ºC accompanied by moist, humid air (>85%) extending for 36 to 72 hours during anthesis and seed development (Bai & Shaner Citation1994; Osborne & Stein Citation2007; McMullen et al. Citation2012). The fungus also produces mycotoxins, such as deoxynivalenol (DON), which can cause health concerns in humans and livestock. Generally, the highest DON concentration that most grain elevators in the USA typically accept is around 2ppm. This is based on the United States Food and Drug Administration’s recommendation of 1ppm in finished wheat products (U.S. FDA, Citation2010), and the assumption that DON concentration is reduced in grain blending and traditional roller milling.
Management of Fusarium head blight (FHB) relies on an integrated strategy of chemical management with fungicides and varietal resistance, with the latter being the most important tool. One of the first FHB resistance quantitative trait loci (QTL) identified was Qfhs.ndsu-3 BS (Fhb1), derived from ‘Sumai 3’, which was introduced by Waldron et al. (Citation1999). Breeders have produced resistant cultivars containing Fhb1 such as NDSU cultivars ‘Alsen’ and ‘Faller’, and Syngenta’s cultivar ‘Soren’ (Browne Citation2007). ‘Sumai 3’ and Fhb1 are not the only resistance source. For example, US cultivars such as ‘Freedom’ and ‘Ernie’ express resistance unlinked to ‘Sumai 3’. Complete resistance or immunity have not been found in any cultivar worldwide. The important economic impacts and health concerns that are caused by this disease necessitate ongoing resistance-breeding efforts.
Wheat varieties are typically released as inbred lines because wheat is a self-pollinating crop. However, advances in chemical hybridizing agents (CHAs) and cytoplasmic male sterility (CMS) systems have renewed the possibility of hybrid wheat becoming commercially viable. Hybrid wheat has shown yield advantages over pure-line cultivars. Small plot research studies have shown yield increases of 5–15% over the better parent using parents from the same class; however, drill-sown trials show yield increases at or over 20% (Singh et al. Citation2010; Whitford et al. Citation2013). When using parents from differing classes of wheat, such as soft and hard red, heterosis levels reach nearly 30% (Singh et al. Citation2010). An efficient hybrid system could help generate a yield boost in wheat that will help feed the world’s increasing population.
Questions remain about the effectiveness of the hybrid production system. One question is how disease resistance will be affected by the hybrid system. Miedaner et al. (Citation2017) found that hybrids, on average, expressed FHB resistance greater than the mean of their parents. While dominance and epistatic gene action has been observed, FHB resistance is quantitatively inherited showing an abundance of additive effects (Miedaner et al. Citation2017). Knowledge of the type of resistance gene action could help formulate crossing strategies. Complete dominance of all FHB resistance genes would be desirable to hybrid breeders because the F1 progeny obtained by pairing a resistant parent with a susceptible parent would have acceptable levels of resistance. Hybrids produced from two parents expressing resistance genes were reported to exhibit resistance levels most similar to that of the better parent (Miedaner et al. Citation2017). However, it is currently unknown if a hybrid produced from only one resistant parent will express sufficient resistance to be commercially acceptable. What is commercially acceptable will be subjective, but resistance more similar to the resistant parent would be desirable. Therefore, the objectives of this experiment were to evaluate the FHB resistance of F1 hybrids and their parents by measuring visual symptoms, Fusarium damaged kernels, grain volume weight and DON accumulation, when the hybrids and parents were grown in the field across multiple environments using artificial inoculation.
Materials and methods
Parental genotypes and hybrid production
F1 hybrids were created by crossing four moderately FHB-resistant cultivars to (‘Alsen’, ‘Faller’, ‘Glenn’, and ‘SY Soren) with a common susceptible cultivar (‘2398’). Resistant parents were selected based on the combination of resistance alleles they were known to carry (). In the NDSU variety selection guide, ‘Faller’ and ‘Glenn’ are rated as a four, and ‘SY Soren’ is a seven for FHB score on a scale of 1–9, where a 1 is highly resistant and a 9 is highly susceptible (Ransom et al. Citation2019). Cultivar ‘2398’ is a variety released by the North Dakota Agricultural Experiment Station (NDAES) and developed by the Pioneer Hi-Bred HRSW program in the 1980’s. It has been used historically as a highly susceptible check in the NDSU scab nursery as referenced in registrations for ‘Faller’, ‘Glenn’, and ‘Alsen’ (Frohberg et al. Citation2006; Mergoum et al. Citation2006, Citation2008).
Table 1. Descriptions of the parents that were used for hybrid production including Fusiarum head blight (FHB) resistance alleles, year of cultivar release, pedigree, and developing institution
All hybrid seed was produced by hand emasculations and pollinations in the NDAES greenhouse in Fargo, ND. Reciprocal crosses were made and tested, and when no differences between any reciprocal cross were detected, they were combined into a single genotype for analysis. Hybrid seed was bulk harvested by specific parent by parent crosses in Spring 2018. Seed production of parents and of hybrids was repeated in Fall 2018 and Spring 2019. The identity of the parental genotypes was verified using molecular markers commonly used in the marker assisted selection programme at NDSU during the summer of 2018. Markers associated with loci mentioned in (Fhb3A, Fhb3B, Fhb5A) along with Lr34 (a leaf rust resistance gene) were used to confirm the identity of parental genotypes. Marker analysis was done by the USDA-ARS genotyping laboratory at Fargo, ND. Three seeds of each F1 hybrid were also checked using the previously mentioned markers to confirm heterozygosity at each locus. Visual verifications of hybridity were also made during the field trials. Plant height and anthesis date were measured to distinguish between F1 hybrids and inbred lines. Both traits were expected to be intermediate to the parents. Plant height was recorded at physiological maturity.
Field experiments and data recording
The field experiments were organized in a randomized complete block design (RCBD) with six replications in 2018 and three replications in 2019. In addition to the parental lines and the hybrids, two checks were included in the experiments and repeated three times in each replication. The repeated checks were ‘ND2710’ (PI 633 976) as the resistant check and ‘WB Mayville’ (PI 661 061) as the susceptible check. Plots consisted of three-seed hill plots. The experiment was planted using a four-row planter with 30 cm spacing between rows and 58 cm between plots. Experimental plots were surrounded by a border of ‘WB Mayville’. All agronomic practices were applied in accordance with regional practices for each of the locations. In addition to herbicide application, hand weeding was completed to remove weeds that the herbicides did not control. No fungicide was applied to the experimental plots or surrounding border plots.
Fusarium graminearum colonized corn grain was used as inoculum to ensure a reoccurring source of spores. For Langdon and Prosper, we used inoculum composed of 20 isolates (ten 3A-DON producing and ten 15A-DON producing isolates) collected throughout North Dakota. The inoculum was prepared in accordance with the method described by Balut et al. (Citation2013) in the Plant Pathology laboratory of North Dakota State University. For Sabin, the colonized corn grain inoculum was prepared by BASF and contained one 3A-DON producing isolate and one 15A-DON producing isolate. Fusarium inoculum was spread by hand at multiple growth stages to ensure a high infection rate. In 2018, the inoculation was conducted twice at early jointing (Feekes 7) and early boot stages (Feekes 10). In 2019, inoculum was applied three times at tillering (Feekes 3), early jointing (Feekes 7), and early booting (Feekes 10) or early heading (Feekes 10.3) stages. In each location, the total combined rate of corn grain inoculum was 61 g m−2.
Experimental plots were irrigated using an overhead mist irrigation system, with the first irrigation beginning at the first inoculation. Irrigation water was supplied in one to two minutes durations per hour for four days per week. In Langdon, the irrigation operated 12-h during daytime. The Prosper irrigation system operated on a 24-h basis. During periods of excessively dry or wet conditions, the irrigation system was adjusted to ensure that moisture was not limiting. Misting methods used at Sabin included an application of water for 30 min late in the afternoon every other day.
The disease incidence and severity were assessed for all hill plots around milk/dough development stages (Feekes 11.1–11.2) and those values were used to create a FHB index. Disease incidence was calculated by counting the number of diseased heads out of 10 randomly selected heads and expressed as a percentage (Khorzoght et al. Citation2010). Ten infected heads were also evaluated for disease severity by counting total diseased spikelets per head and dividing that by the total spikelets in the head, expressed as a percentage (Khorzoght et al. Citation2010; Balut et al. Citation2013). The disease index was calculated as below according to Khorzoght et al. (Citation2010).
Post-harvest evaluations of FHB
At physiological maturity (Feekes 11.4), all heads from each plot were harvested and dried for four days at 16°C. The Langdon experiment in 2018 was not rated or harvested due to lack of Fusarium infection. Harvested heads were threshed on a whole plot basis using a stationary thresher (Sparc Machine Design, Swift Current, SK), with minimum air flow to retain as many Fusarium damaged kernels as possible. Pieces of chaff were removed by hand after threshing.
Seed lots for each individual plot were evaluated visually for Fusarium damaged kernels (FDK) following Jones & Mirocha (Citation1999), with the FDK value being expressed as percentage of damaged kernels in the sample. A sample from each plot was evaluated for deoxynivalenol (DON) accumulation, given in parts per million (ppm), at DON testing laboratories (University of Minnesota in 2018 and NDSU in 2019). Samples were ground using a UDY Cyclone Sample Mill model 3010–014 (UDY Corporation; Fort Collins, CO). The mill was cleaned between each sample using a brush, forced air, and vacuum to ensure no cross-contamination between samples. Samples were grouped by genotype in order to minimize contamination and the mill cleaned extra thoroughly when changing to a new genotype. The mill was allowed to cool after every samples to avoid any sample damage through contact with hot rotor blades.
A DISK rating was calculated to observe the cumulative effect of multiple traits. It is a summation of four individual ratings (DON, incidence, severity, and FDK) to generate a single numeric rating. Higher weight is given to FDK and DON because they measure economic effects of the disease. A high FDK can lead to reduced grain yield or test weight discounts. High DON may result in grain being rejected at the point of sale. The DISK values were calculated as shown below according to the calculation procedure of Bissonnette et al. (Citation2018).
Statistical analysis
Combined and individual environment analysis of variance were conducted using PROC GLM of SAS 9.4 (SAS Institute, Cary, NC). The SAS PROC MIXED was applied to make several contrasts including resistant parents versus their hybrids, susceptible parent versus hybrid, and mid-parent versus hybrid. Significant differences between compared groups were declared at P ≤ 0.05. PROC MIXED was also used to generate LSMEANS for the parents, hybrids, and the two checks. These LSMEAN values per environment for FHB Index, FDK, DON, and DISK were used to make clustering of 11 genotypes into groups. The cluster analysis was done in JMP Genomics (JMP® Genomics Citation1989-2019) using Ward’s minimum variance method with standardized data.
Results
Verification of identity of genotypes
Molecular markers were used to verify the identity of each cultivar, especially to confirm hybridity, and to ensure that the resistance alleles were consistent with the published literature. Resistance alleles known to be contained in ‘Alsen’, ‘Faller’, and ‘SY Soren’ were consistent in our screening (). ‘Glenn’ and ‘2398’ had a null allele for all markers in the screening. The hybrids expressed both resistant and alternate alleles for all the FHB resistance loci, as expected per the respective pedigrees. For instance, the 2398*Alsen hybrids appeared heterozygous for all three markers. There also were no anomalies in the Lr34 data, plant height, or anthesis data that caused discarding of any plots (data not included).
Analysis of variance
The genotypic and environmental main effects exhibited highly significant differences (P < 0.01) for all the four traits related to FHB resistance (). The genotypic effect was also highly significant (P < 0.01) in the individual environment analysis (Supplemental ). The interaction of the genotype with environment was also highly significant, which may imply that the genotypes responded differently in resistance across those environments. Despite the significant interaction effects, the susceptible genotypes showed consistently high values for the four traits compared to the resistant genotypes and the hybrids (). The heritability estimates were 0.66 for FHB Index, 0.90 for FDK, 0.36 for DON, and 0.66 for DISK.
Table 2. Combined analysis of variance across the four environments (PRO 18, LANG 19, PRO 19, SAB 19) for four Fusarium head blight (FHB) resistance-related traits (FHB Index, Fusarium Damaged Kernels, DON concentration, DISK score)
Table 3. Least square means and least significant difference (LSD) mean separation for the four response traits (Fusarium head blight (FHB) Index, Fusarium Damaged Kernels, DON concentration, DISK score) of the 11 genotypes included in this study, across all four locations
Comparison of the hybrids with their respective parents
We compared the hybrids with the resistant and susceptible parents used to produce the hybrids as well as the mid-parent value () to evaluate how the hybrids performance in FHB resistance. Here we discuss the results of these comparisons by individual hybrid.
Table 4. Contrast analyses of hybrid genotypes with the resistant, susceptible, and mid-parent value for the four Fusarium head blight (FHB) resistance-related traits, by location and combined across locations
Performance of Alsen Hybrid
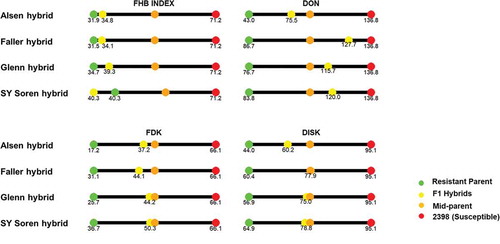
The hybrid created from ‘Alsen’ and ‘2398’ was found to be significantly different from both parents in the four FHB resistance-related traits (FHB index, FDK, DON, and DISK) in individual environments or combined across environments (). The hybrid exhibited less resistance than the resistant parent (‘Alsen’), where the hybrid had about 3%, 20%, 33ppm, and 16 higher values for FHB Index, FDK, DON, and DISK, respectively ( and Supplemental ). However, the hybrid was found to be more resistant than the susceptible parent (‘2398’). This was evident from highly significant (P < 0.01) differences in contrast analysis between ‘2398’ and the hybrid (), and generally the hybrid led to the reduction of FHB index, FDK, DON, and DISK by 36%, 29%, 61 ppm, and 35, respectively (). The ‘Alsen’-based hybrid also exhibited significantly low (P < 0.01) values for all the four FHB resistance-related traits compared to the mid-parent values.
Performance of Faller Hybrid
The performance of the ‘Faller’ hybrid was similar to that of the ‘Alsen’ hybrid (). The ‘Faller’ hybrid exhibited increased resistance compared to the susceptible parent (‘2398’) and the mid-parent. However, it showed more susceptibility compared to the resistance parent (‘Faller’). Overall, the hybrid led to a reduction of FHB index, FDK, DON, and DISK by 40%, 22%, 9 ppm, and 17, respectively; compared to cultivar ‘2398’ ( and Supplemental ). However, it showed higher values () for those FHB resistance-related traits compared to the resistant parent (‘Faller’), implying that the hybrid was not comparable in resistance to ‘Faller’. This was consistent with the other genotypes.
Performance of Glenn Hybrid
In almost all environments, the resistant parent (‘Glenn’) showed lower values compared with the hybrid for all the FHB resistance-related traits, where the differences were shown to be significant (P < 0.05) in most cases (). However, the susceptible parent (2398) and the mid-parent were highly susceptible compared to the Glenn-based hybrid. Glenn was generally lower compared to the hybrid in FHB Index, FDK, DON, and DISK by 5%, 19%, 39ppm, and 18, respectively ( and Supplemental ). The Glenn-based hybrid resulted in a reduction of FHB Index, FDK, DON, and DISK by 32%, 22%, 21ppm, and 20, respectively; compared to the susceptible parent (‘2398’).
Performance of SY Soren Hybrid
The ‘SY Soren’-based hybrid showed inconsistency in the level of resistance compared to the resistant parent, susceptible parent, and mid-parent (). The resistant parent (‘SY Soren’) can be categorized as more resistant compared to the hybrid when FDK, DON, and DISK are considered, but not when FHB Index was taken into consideration. The hybrid exhibited more resistance almost always compared to the susceptible parent (‘2398’). Compared to the mid-parent, the hybrid generally showed more susceptibility based on FHB Index and DISK but higher resistance based on FDK and DON.
Clustering of the genotypes based on resistance to FHB
Clustering of the 11 genotypes using the FHB resistance-related traits (FHB Index, FDK, DON, and DISK) for each of the four environments resulted in three clear groups (). Three of the resistant parents (‘Alsen’, ‘Glenn’, and ‘Faller’), the resistant check (ND2710), and one of the hybrids (2398*Alsen) were clustered together. As expected, the genotypes in cluster 2 exhibited the highest resistance compared to the other two clusters (). The third cluster was composed of four of the genotypes (‘SY Soren’, 2398*Faller, 2398*Glenn, and 2398*SY Soren). The two susceptible genotypes (‘2398’ and ‘WB Mayville’) were grouped together and were shown to be highly susceptible.
Fig. 1 Cluster analysis using Ward’s minimum variance method to organize the eleven genotypes into three clusters based on the FHB Index, Fusarium Damaged Kernels, DON concentration, and DISK score, across the four environments
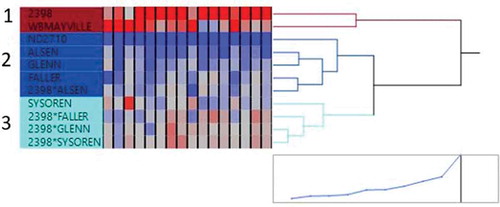
Table 5. Mean and standard deviation of wheat genotypes grouped under the three clusters created using four Fusarium head blight (FHB) resistance-related traits (FHB Index, Fusarium Damaged Kernels (FDK), DON concentration and DISK score)
Discussion
The main objective of this experiment was to compare the resistance of hybrids with their respective parents. In addition, we also discussed how the genetic composition of the parents affected their resistance to FHB. Our evaluation for each hybrid indicated that the hybrids were generally more resistant than the susceptible parent, but significantly less resistant than the resistant parent. Compared with the mid-parent values, the hybrids exhibited more resistance to FHB, as the values for the four FHB resistance-related traits were lower in the hybrids compared with mid-parent values. In general, the resistance level of hybrids was between the mid-parent and the resistant parent (), implying that resistance to FHB could be controlled by partial dominance gene action. Previously, Miedaner et al. (Citation2017) reported that FHB resistance is quantitatively inherited with a high frequency of additive effects and less of dominance and epistatic actions.
Fig. 2 FHB Index, Fusarium Damaged Kernels, DON concentration and DISK score of four F1 hybrids, their moderately resistant parent, the susceptible parent (‘2398’), and mid-parent value, across four environments
Assuming only additive effects, the accumulation of additional resistance alleles should confer increasing levels of FHB resistance. Based on the number of resistance alleles, the resistant parents should order as ‘Alsen’ most resistant, followed by ‘Faller’, and then ‘SY Soren’ and ‘Glenn’ (). For all the four FHB resistance-related traits, ‘Alsen’ had the lowest (most resistant) while ‘Faller’ had the second lowest values. This result may indicate that the accumulation of resistance genes could increase resistance. Glenn has historically been rated highly resistant despite not containing a marker for any of the known resistance genes (ElDoliefy Citation2015).
In general, hybrids in this experiment did not have the same resistance level as the resistant parents. Even if the resistance of hybrids was less than the resistant parent, the alleles from the resistant parent provided the hybrids a good level of resistance to FHB. This suggests that successful hybrids could be developed using only one resistant parent. In practice, the use of a susceptible parent as one of the parents in hybrid production is justifiable if the susceptible parent provides favourable alleles for other traits of breeding importance. Otherwise, it is important to use resistant parental lines for hybrid production. To our knowledge, this study is the first attempt to evaluate the resistance level in hybrids produced from susceptible by resistant crosses. Thus, further studies need to be done using a different set of parents to conclusively understand the feasibility of hybrids from parental lines having contrasting resistance levels for FHB.
Conflict of interest
The authors declare no conflict of interest from funding provided by Corteva Agriscience nor BASF for their collaboration and hosting a research location
Supplemental_File.docx
Download MS Word (20.9 KB)Supplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
References
- Bai GH, Shaner G. 1994. Scab of wheat: prospects of control. Plant Dis. 78(8):760–764. doi:10.1094/PD-78-0760.
- Balut AL, Clark AJ, Brown-Guedira G, Souza E, Sanford DA. 2013. Validation of Fhb1 and QFhs. nau-2DL in several soft red winter wheat populations. Crop Sci. 53:934–945. doi:10.2135/cropsci2012.09.0550.
- Bissonnette KM, Kolb FL, Ames KA. 2018. Effect of wheat cultivar on concentration of Fusarium mycotoxins in wheat stems. Plant Dis. 102:2539–2544. doi:10.1094/PDIS-12-17-2034-RE.
- Browne RA. 2007. Components of resistance to fusarium head blight (FHB) in wheat detected in seed-germination assay with Microdochium majus and the relationship to FHB disease development and mycotoxin accumulation from Fusarium graminearum infection. Plant Pathol. 56:65–72. doi:10.1111/j.1365-3059.2006.01509.x.
- ElDoliefy AEA. 2015. Molecular mapping of fusarium head blight resistance in two adapted spring wheat cultivars [Doctoral Dissertation]. Retrieved from ProQuest Dissertations & Theses (Order No. 3685801). @ North Dakota State University; ProQuest Dissertations & Theses Global. (1664843621). https://ezproxy.lib.ndsu.nodak.edu/login?url=https://search.proquest.com/docview/1664843621?accountid=6766
- Friskop A, Zhong S, Breuggeman R. 2018. Fusarium head blight (Scab) of small grains. NDSU Extension Service Bulletin. PP804 (Revised). https://www.ag.ndsu.edu/publications/crops/fusarium-head-blight-scab-of-small-grains/pp804.pdf
- Frohberg RC, Stack RW, Olson T, Miller JD, Mergoum M. 2006. Registration of ‘Alsen’ wheat. Crop Sci. 46:2311–2312. doi:10.2135/cropsci2005.12.0501.
- JMP® Genomics. Version 9.0. 1989-2019. Cary (NC): SAS Institute Inc.
- Jones RK, Mirocha CJ. 1999. Quality parameters in small grains from Minnesota affected by fusarium head blight. Plant Dis. 83:506–511. doi:10.1094/PDIS.1999.83.6.506.
- Khorzoght EG, Soltanloo H, Ramezanpour SS, Arabi MK. 2010. Combining ability analysis and estimation of heterosis for resistance to head blight caused by Fusarium graminearum in spring wheat. AJCS. 4(8):626–632.
- McMullen M, Bergstrom G, Wolf ED, Dill-Macky R, Hershman D, Shaner G, Van Sanford D. 2012. A unified effort to fight an enemy of wheat and barley: fusarium head blight. Plant Dis. 96(12):1712–1728. doi:10.1094/PDIS-03-12-0291-FE.
- Mergoum M, Frohberg RC, Stack RW, Olson T, Friesen TL, Rasmussen JB. 2006. Registration of ‘Glenn’ wheat. Crop Sci. 46:473–474. doi:10.2135/cropsci2005.0287.
- Mergoum M, Frohberg RC, Stack WR, Rasmussen JB, Friesen TL. 2008. Registration of ‘Faller’ wheat. J Plant Regist. 2:224–229. doi:10.3198/jpr2008.03.0166crc.
- Miedaner T, Schulthess AW, Gowda M, Reif JC, Longin CFH. 2017. High accuracy of predicting hybrid performance of fusarium head blight resistance by mid-parent values in wheat. Theor Appl Genet. 130:461–470. doi:10.1007/s00122-016-2826-8.
- Osborne LE, Stein JM. 2007. Epidemiology of fusarium head blight on small-grain cereals. Int J Food Microbiol. 119:103–108. doi:10.1016/j.ijfoodmicro.2007.07.032.
- Ransom J, Green A, Simsek S, Friskop A, Breiland M, Friesen T, Liu Z, Zhong S, Rickertsen J, Eriksmoen E, et al. 2019. North Dakota hard red spring wheat variety trial results for 2019 and selection guide. NDSU Extension Service & NDAES. https://www.ag.ndsu.edu/publications/crops/north-dakota-hard-red-spring-wheat-variety-trial-results-for-2019-and-selection-guide/a574-hrsw-19.pdf
- Singh SK, Chatrath R, Mishra B. 2010. Perspective of hybrid wheat research: a review. Indian Journal of Agricultural Sciences. 80(12):1013–1027.
- United States Department of Agriculture National Agriculture Statistics Service [USDA/NASS]. 2019. 2018 state agriculture overview – North Dakota. https://www.nass.usda.gov/Quick_Stats/Ag_Overview/stateOverview.php?state=NORTH%20DAKOTA
- United States Food and Drug Administration. 2010. Guidance for industry and FDA: advisory levels for deoxynivalenol (DON) in finished wheat products for human consumption and grains and grain by-products used for animal feed. https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-fda-advisory-levels-deoxynivalenol-don-finished-wheat-products-human
- Waldron BL, Moreno-Sevilla B, Anderson JA, Stack RW, Frohberg RC. 1999. RFLP mapping of QTL for fusarium head blight resistance in wheat. Crop Sci. 39:805–811. doi:10.2135/cropsci1999.0011183X003900030032x.
- Whitford R, Fleury D, Reif JC, Garcia M, Okada T, Korzun V, Langridge P. 2013. Hybrid breeding in wheat: technologies to improve hybrid wheat seed production. J Exp Bot. 64(18):5411–5428. doi:10.1093/jxb/ert333.