Abstract
Lake Mead has experienced drought conditions since 2000 and the introduction of invasive quagga mussels since about 2004. Empirical evidence from the Great Lakes region suggests a potential for energy transfer to the benthos and away from planktivorous fish through competition between quagga mussels and filter-feeding, herbivorous zooplankton. We describe Daphnia dynamics from 2043 analyses performed on the 3 basins of subtropical Lake Mead during 2000–2009. Daphnia were generally most numerous in shallower areas strongly influenced by nutrient inputs and with decreased clarity and higher chlorophyll a (chl-a) concentrations. Daphnia populations increased in mid- to late winter and early spring prior to the spawning of gamefish populations and rapidly decreased with increasing predation pressure, water temperature and cyanobacteria populations. Concurrent with the reduction in Daphnia populations in late spring, quagga veliger abundance rapidly increased, declined in summer, and peaked again in the fall. Quagga veligers in Lake Mead were generally most numerous in open water areas with maximum depth, greater clarity and low chl-a. Although mean Daphnia abundance since the beginning of drought conditions and the establishment of quagga mussels is lower in Boulder Basin, the temporal and spatial dynamics of the zooplankton community remain comparable to previously described historic patterns in all 3 basins of Lake Mead.
Lake Mead is experiencing conditions that typically stress aquatic ecosystems (drought and introduction of dreissenid mussels). Reservoir levels began to drop dramatically in 2000 in this subtropical reservoir and are now near a 40-year low. Little is known about the impacts of climatic changes and resulting periods of drought to top-down, bottom-up, and abiotic controls that structure qualitative and quantitative characteristics of plankton communities. Studies describing the zooplankton response to drought are particularly limited (CitationMcGowan et al. 2005, CitationHavens et al. 2007). Quagga mussels (Dreissena rostriformis bugensis) became established in Boulder Basin subsequent to drought conditions. Quagga mussels were first confirmed in January 2007, but likely colonized around 2004 (CitationMcMahon 2007). Since becoming established, quagga mussels have expanded throughout all basins of Lake Mead. Given their ability to attain high biomass, combined with intrinsically high clearance rates, dreissenid mussels can modify established trophic linkages by grazing phytoplankton and altering patterns in nutrient recycling (CitationVanderploeg et al. 2002, Citation2009, CitationWilson and Sarnelle 2002). Extensive research on the ecological impacts of invasive dreissenid mussels exists for north temperate lakes (i.e., Great Lakes region, Europe), but the ultimate ecological impact of invasive dreissenid mussels on subtropical reservoirs with differing thermal and hydrological regimes is unknown.
Our objective was to examine a long-term database (2000–2009) for evidence that Daphnia spp. have been significantly affected by drought and subsequent invasion of quagga mussels. Previous studies of the zooplankton community of Lake Mead have been limited in scope and duration (CitationBurke 1977, CitationPaulson and Baker 1983, CitationWilde 1984, CitationAxler et al. 1988, CitationWilde and Paulson 1988, CitationSollberger and Paulson 1991, CitationLoomis 2009).
Methods
Sample locations
Lake Mead was at near full capacity at the beginning of the study, but water elevation progressively dropped over the subsequent 10 years. The end-of-month elevation and content as of January 2000 was 1214.26 ft, 25.0 million acre-feet (97% capacity). The end-of-month elevation and content as of December 2009 was 1096.30 ft, 11.2 million acre-feet (43% capacity). Major surface inflows to Lake Mead include the Colorado River (∼97%), the Virgin and Muddy rivers, and the Las Vegas Wash (CitationHoldren et al. 2006). Boulder Basin is the most downstream portion of the reservoir and collects the combined flows of Overton Arm and the Colorado River and receives nonpoint surface discharges as well as the treated effluent from the City of Las Vegas. The hydrodynamics and thermal regime of Lake Mead are complex (CitationLaBounty and Burns 2005). Monitoring of epilimnetic zooplankton populations of the pelagic region in the 3 basins of Lake Mead was conducted from 2000 through 2009. Samples were collected from established sampling stations in the 3 basins, Boulder Basin (n = 8), Colorado River (n = 8) and Overton Arm (n = 6; ). Sampling transects for each basin included gradients from stations near inflows to open, deeper water. The US Bureau of Reclamation (Denver Technical Services Center) has collected approximately 20 samples monthly from Boulder Basin and Overton Arm since 2000 and the Colorado River since 2001. The US Bureau of Reclamation (Lower Colorado Office) has collected approximately 20 samples quarterly from all 3 basins of Lake Mead since November 2006. The Southern Nevada Water Authority has collected samples weekly from approximately 6 stations in Boulder Basin since 2000.
Figure 1 Location of Lake Mead zooplankton sampling stations (CR = Colorado River, LVB = Las Vegas Bay, VR = Virgin River). ▴ = Colorado River Stations; • = Overton Arm Stations; • ▪ Boulder Basin Stations.
Limnological characteristics of Boulder Basin, Overton Arm and the Colorado River and details of collection and analyses of chlorophyll a (chl-a) and physical parameters are described elsewhere (e.g., CitationLaBounty and Burns 2005, Citation2007, CitationHoldren et al. 2006). The trophic state of Lake Mead is mostly mesotrophic with the exception of Boulder Basin, which is classified as supertrophic in the inner basin (located immediately downstream from the Las Vegas Wash) and mesotrophic in the middle and outer basins (CitationLaBounty and Burns 2005). Lake Mead is stratified throughout the majority of the year. Mixing of the epilimnion occurs during a 2 month period (Jan–Feb) to a depth of at least 60–70 m (CitationLaBounty and Horn 1997, CitationLaBounty and Burns 2005, Citation2007). Average water temperature in the epilimnion ranges from approximately 12 C in early February to approximately 27 C in early August (CitationLaBounty and Burns 2005).
Zooplankton collection and analyses
The epilimnion was operationally defined based on collection methodology. Zooplankton samples were limited to either the top 30 m for stations deeper than 30 m or from the top of the water column to the bottom for stations <30 m deep (Southern Nevada Water Authority's collections were from the top 40 m or to the bottom). CitationMueller and Horn (2004) documented that the vast majority of zooplankton in Lake Mead are found within the top 40 m of the water column in the open water stations. They also documented that selection of this range for sampling negated any artifacts related to vertical migration of zooplankton because most would be within this range regardless of sampling time. Zooplankton samples were collected with a Wisconsin-style plankton net with 64 or 80 μm mesh, which would underestimate the rotifer population (CitationChick et al. 2010), but given the importance of daphnids in the ecosystem it is unlikely that epilimnetic rotifer populations were significant (CitationVanni 1986). Samples were immediately preserved with Lugol's solution and placed on ice after collection.
Appropriate aliquots were placed in an Utermohl chamber, and zooplankton were counted at 100× magnification on a customized Wilovert inverted microscope until at least 200 individuals were tallied (cladocerans, copepods, rotifers, veligers). Identification followed CitationEdmundson (1959), CitationRuttner-Kolisko (1974), CitationStemberger (1979), and CitationPennak (1989). The Zebra Mussel Information System (ZMIS) developed and distributed by the US Army Corp of Engineers was used for identification of Dreissena veligers, and enumeration and identification was conducted at 100× (USACE 2002).
Biomass estimates were based on established length–width relationships (CitationDumont et al. 1975, CitationMcCauley 1984, CitationLawrence et al. 1987). The lengths or the lengths and widths of each species were measured equal to 10 for common species and less for more rare taxa. More detailed descriptions of biomass estimation methodology are given in CitationHavens et al. (2007) and Havens and Beaver (2010). In accordance with CitationMcCauley (1984), biomass was computed for the appropriate number of individuals for each sample location, and the arithmetic mean biomass was multiplied by the species abundance to produce a species biomass for each sample.
Data analyses
The study included 2043 zooplankton analyses over the 10 year sampling period. Because most stations were sampled multiple times in a single month, individual station values for a given month were averaged (22 station means). A grand mean was computed for each of the individual flows (Boulder Basin, Overton Arm, Colorado River) based on station means. Chl-a values (top 0–5 m of the water column) and Secchi disk transparency were obtained from the Southern Nevada Water Authority Members’ database and were computed as described for zooplankton.
Results
Chl-a concentrations and Daphnia abundance and biomass
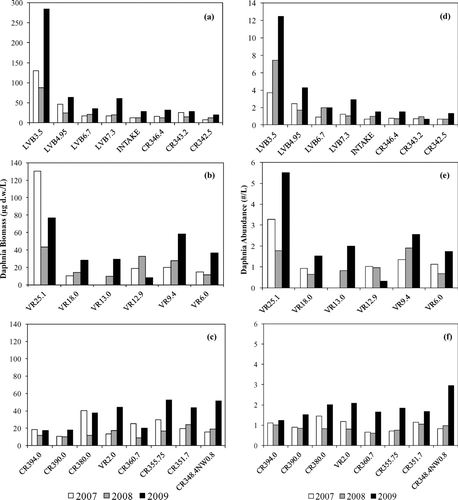
Basin-wide means for Daphnia abundance (D. pulex and D. galeata) and chl-a concentrations from 2000 through 2009 each demonstrate seasonal trends (). Peaks in chl-a concentrations consistently occurred in Boulder Basin, Overton Arm and the Colorado River during summer and were lowest during winter. Daphnia densities consistently peaked in winter and early spring in each basin. Boulder Basin demonstrated declining trends in both chl-a concentrations and Daphnia abundance during the 10 year study. The less productive Overton Arm and Colorado River stations did not demonstrate a declining pattern in chl-a concentrations. Basin-wide mean Daphnia biomass estimates for the period November 2006 through December 2009 reflect the winter to early spring peak seen in the abundance data (). Daphnia accounted for approximately 70% of the mean total zooplankton biomass for the Lake Mead system (Boulder Basin = 75%, Overton Arm = 69%, Colorado River = 75%). Boulder Basin demonstrated higher mean Daphnia biomass in comparison with Overton Arm and the Colorado River (Boulder Basin = 28.0 μg d.w./L, Overton Arm = 18.4 μg d.w./L, Colorado River =13.8 μg d.w./L). All basins experienced larger Daphnia biomass in 2009 compared to values in 2007 and 2008. An evaluation of the importance of proximity of nutrient inflows to Daphnia biomass and abundance () showed that Boulder Basin (LVB 3.5) and Overton Arm (VR25.1) stations located immediately downstream from the inflows displayed more Daphnia biomass and abundance for the period January through May for all 3 years under study. The Colorado River stations did not exhibit this relationship, and in 2009 the downstream stations were actually higher than stations near the inflow.
Figure 2 Chl-a (μg/L) and Daphnia abundance (#/L; n = 1872) for (a) Boulder Basin (n = 981); (b) Overton Arm (n = 323); and (c) Colorado River (n = 568) from Jan 2000–Dec 2009. ![]()
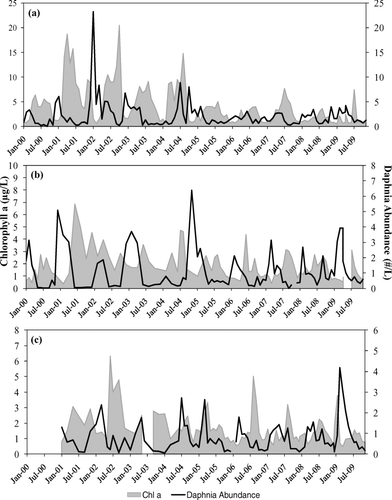
Figure 3 Daphnia biomass (μg d.w./L) for (a) Boulder Basin (n = 515); (b) Overton Arm (n = 183); and (c) Colorado River (n = 369) during Oct 2006–Dec 2009. ▪ = Daphnia biomass (μg d.w./L); □ = total zooplankton biomass (μg d.w./L).
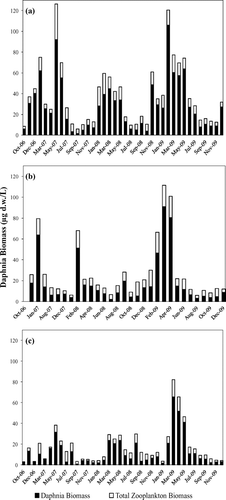
Figure 4
Daphnia biomass (μg d.w./L; a = Boulder Basin, b = Overton Arm, c = Colorado River) and abundance (#/L; d = Boulder Basin, e = Overton Arm, f = Colorado River) for the sampling stations within Boulder Basin (n = 255), Overton Arm (n = 80) and Colorado River (n = 148) during the period Jan–May for 2007, 2008, and 2009. □ = 2007; ![]()
Quagga veliger abundance and distribution
Quagga veligers were first detected in zooplankton samples collected at CR 355.75 (the Narrows) in November 2006, and adults were confirmed at the Las Vegas Boat Harbor and at Lake Havasu, a downstream reservoir, in January 2007. Boulder Basin and Colorado River stations experienced peaks in fall 2007 (). Quagga veliger abundance in Overton Arm in fall 2007 was modest; however, by fall of 2008 the mean abundance exceeded the other basins (>30/L). A midsummer decrease in mean quagga veliger abundance occurred in Boulder Basin and Colorado River 2007 and 2008 and was followed by increased abundance until October. In 2008 and 2009, quagga veligers displayed a clear bimodal seasonality with spring and fall pulses that corresponded to water temperatures of 18–20 C in the top 5 m of the water column in this study and other studies of Lake Mead (CitationLaBounty and Burns 2005). By 2009 the densities of veligers decreased in Boulder Basin and Overton Arm but maintained the bimodal seasonality observed in 2007 and 2008. Colorado River veliger abundance in 2009 was similar to 2007 and 2008. Quagga veligers were most numerous in open water areas with maximum depth, greater clarity, low chl-a concentrations (; ) and the largest amount of submerged geological features (CitationTwichell et al. 1999, CitationTwichell and Cross 2004).
Discussion
Comparison with previous studies on Lake Mead
Previous Lake Mead research indicates that the response of the phytoplankton community to nutrient loadings has been the single most important factor regulating the seasonal, spatial and temporal abundance of zooplankton in the phosphorus (P)-limited reservoir. Established temporal and spatial patterns in primary productivity and phytoplankton populations include maxima near point source nutrient inputs (e.g., Las Vegas Wash [Boulder Basin], Virgin and Muddy rivers [Overton Arm]), decreasing with distance from the inflows (e.g., CitationLaBounty and Horn 1997, CitationHoldren et al. 2006). Phytoplankton populations typically peak from July through September. Similar temporal and spatial patterns in zooplankton and fish populations in Lake Mead have been ascribed to variations in nutrient inputs (nutrient abatement–nutrient inflows) and associated gradients in phytoplankton populations (e.g., CitationLaBounty et al. 2004, CitationMueller and Horn 2004).
Consistent with previous studies, Daphnia populations in this study were low during summer, increased in fall, reached maximum in winter–early spring and then decreased again during spring. The highest Daphnia densities occurred in the inner Las Vegas Bay followed by inflow stations in the Overton Arm. Daphnia densities consistently decreased down-reservoir from the inflows for these 2 basins, and densities were lowest in the main basin stations. In contrast, the more rapidly moving Colorado River inflow stations did not follow this pattern. The importance of nutrient inputs to the trophic linkages in Lake Mead is underscored by the results of the in situ fertilization of Overton Arm in the late 1980s which resulted in a greater than 4-fold increase in Daphnia pulex abundances and improved lipid storage and egg production rates (CitationAxler et al. 1988). Recently, CitationLoomis (2009) documented the trophic linkage between the forage fish base (threadfin shad) and zooplankton in Las Vegas Bay and Overton Arm of Lake Mead (also see CitationWilde and Paulson 1988).
North temperate zone studies
Ecological impacts of dreissenid mussels in north temperate systems include reduced phytoplankton populations and less efficient energy transfer to higher trophic levels (CitationConroy and Culver 2005, CitationGarton et al. 2005). Filter-feeding competition between dreissenid mussels and both macrozooplankton and microzooplankton for food resources (phytoplankton and bacteria) has adversely impacted fisheries in the Great Lakes and resulted in energy transfer to the benthos and away from planktivorous fish (CitationBridgeman et al. 1995, CitationMacIsaac et al. 1995, CitationNalepa et al. 2008, CitationFahnenstiel et al. 2010). Empirical evidence from the Great Lakes region indicates that dreissenid grazing activities can also have a positive impact on cyanobacteria (i.e., Microcystis aeruginosa) in low-nutrient, P-limited lakes (P < 25 μg/L) despite reductions in P-loadings to the systems (CitationVanderploeg et al. 2002, CitationRaikow et al. 2004, CitationKnoll et al. 2008). Dreissenid mussels exhibit differential grazing on algal species (e.g., CitationLavrentyev et al. 1995) and prefer hard, deep and colder habitats, but with increasing depth they will colonize sands and silty sands (CitationMills et al. 1996).
Figure 5 Quagga veliger abundance (#/L) for (a) Boulder Basin (n = 515); (b) Overton Arm (n = 183); and (c) Colorado River (n = 369) during Oct 2006–Dec 2009. ![]()
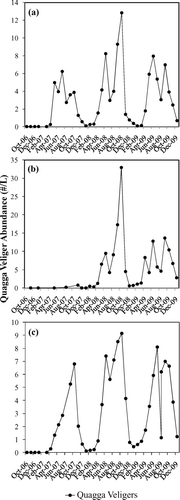
Figure 6 Abundance of quagga veligers (organisms/L) at the sampling locations on (a) October 2007, (b) October 2008, and (c) October 2009. Sampling station CR343.2 was not sampled in October 2009. Sampling station VR12.9 was not sampled on the selected dates.
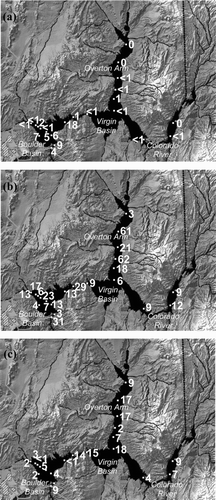
Table 1 Characteristics of zooplankton sampling stations. For each station, values represent means for chlorophyll a (n = 1506) and Secchi depth (n = 1148).
Importance of temperature and season
Life history patterns of phytoplankton populations, Daphnia and the major forage fish in Lake Mead, threadfin shad (Dorosoma petenense), are strongly coupled (CitationWilde and Paulson 1988, Citation1989) and at least partially dependent on water temperature. Following thermal overturn of the epilimnion in winter (∼12 C in Jan–Feb), the phytoplankton community of Lake Mead is dominated by high-quality algal food resources such as cryptomonads and chrysophytes, which are replaced by chlorophytes in the spring (18–20 C) and increased cyanobacteria in the summer (>25 C; LaBounty et al. 2004; Beaver et al. unpublished data). Life history traits for Daphnia pulex include a strong relationship between body-size, fecundity and feeding efficiency, with optimal temperatures around 20 C and decreasing population and metabolic status at higher temperatures (CitationLynch 1977, CitationRellstab and Spaak 2009). Elevated summer epilimnetic temperatures in Lake Mead favor a phytoplankton community dominated by low-quality food resources (i.e., cyanobacteria), and Daphnia populations are greatly reduced. Threadfin shad spawn concurrently with the spring Daphnia pulse from March through June (CitationWilde and Paulson 1989), and their annual productivity is strongly related to total phosphorus (TP) and chl-a concentrations (CitationNey 1996). Summer temperatures in the epilimnion of Lake Mead (>25 C) are also suboptimal for dreissenid mussels (CitationSpidle et al. 1995), and lower food quality (i.e., cyanobacteria) may increase their respiration rates and energy expenditures resulting in reduced reproduction and filter-feeding rates (CitationStoeckmann 2003).
Conclusions
Because of their contribution to total zooplankton biomass, Daphnia are the key intermediaries in energy transfer to higher trophic levels in Lake Mead. Largest populations of Daphnia were associated with nutrient-rich and higher primary production inflow areas, while quagga veligers were most numerous in open water areas. Impacts of invasive quagga mussels on food web dynamics in Lake Mead are unclear. Conventional thought that dreissenid mussel grazing will result in major ecological change outside the north temperate zone is unsubstantiated by our data. Lake Mead is a monomictic reservoir with contrasting thermal, hydrological and ecological conditions to those of the north temperate ecosystems that form the basis for empirical observations on the impacts of dreissenid mussels. Thermal overturn of the epilimnion is limited to winter months, and grazing impacts of quagga mussels are more intensive in well-mixed systems than thermally stratified systems (CitationAckerman et al. 2001, CitationIdrisi et al. 2001, CitationKissman et al. 2010, CitationFahnenstiel et al. 2010). The success of quagga mussels in the Lake Mead ecosystem could be measured by reduction in epilimnetic phytoplankton and zooplankton populations and increased importance of cyanobacteria. Chl-a concentrations and Daphnia abundance have decreased only in Boulder Basin since drought conditions and subsequent invasion by quagga mussels; however, interpretation of the ecological significance is confounded by simultaneous reductions in nutrient loadings from improved wastewater treatment and hydrologic inflows (CitationLaBounty and Burns 2007, CitationLaBounty 2008). Nevertheless, the well-described trophic level interactions beginning with thermal overturn in winter and subsequent pulses of high-quality phytoplankton, Daphnia and fish populations in the spring will likely persist subject to continued, localized nutrient inflows.
Acknowledgments
This work is a continuation of research initiated by Dr. James LaBounty. His critical insights and forethought on the importance of the functioning of the Lake Mead ecosystem have provided all researchers with a better understanding of the complexities of the ecosystem. The authors are grateful to 3 anonymous reviewers for their constructive comments.
References
- Ackerman , J D , Loewen , M R and Hamblin , P F . 2001 . Benthic-pelagic coupling over a zebra mussel reef in western Lake Erie. . Limnol Oceanogr , 46 : 892 – 904 .
- Axler , R P , Paulson , L , Vaux , P , Sollberger , P and Baepler , D H . 1988 . Fish aid: the Lake Mead fertilization project. . Lake Reserv Manage. , 4 ( 2 ) : 1 – 12 .
- Bridgeman , T B , Fahnenstiel , G L , Lang , G A and Nalepa , T F . 1995 . Zooplankton grazing during the zebra mussel (Dreissena polymorpha) colonization of Saginaw Bay, Lake Huron. . J Great Lakes Res , 21 : 567 – 573 .
- Burke , T A . 1977 . The limnetic zooplankton community of Boulder Basin, Lake Mead in relation to the metalimnetic oxygen minimum. MS Thesis , Las Vegas (NV) : University of Nevada .
- Chick , J H , Levchuk , A P , Medle , K A and Havel , J H . 2010 . Underestimation of rotifer abundance a much greater problem than previously appreciated. . Limnol Oceanogr: Methods , 8 : 79 – 87 .
- Conroy , J D and Culver , D A . 2005 . Do dreissenid mussels affect Lake Erie ecosystem stability processes? . Am Midl Nat , 153 : 20 – 32 .
- Dumont , H J , Van de Velde , I and Dumont , S . 1975 . The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. . Oecologia , 19 : 75 – 97 .
- Edmundson , W T . 1959 . Freshwater Biology. 2nd ed. , New York (NY) : Wiley-Interscience .
- Fahnenstiel , G , Pothoven , S , Vanderploeg , H , Klarer , D , Nalepa , T and Scavia , D . 2010 . Recent changes in primary production and phytoplankton in the offshore region of southeastern Michigan. . J Great Lakes Res , 36 : 20 – 29 .
- Garton , D W , Payne , C D and Montoya , J P . 2005 . Flexible diet and trophic position of dreissenid mussels as inferred from stable isotopes of carbon and nitrogen. . Can J Fish Aquat Sci. , 44 ( 62 ) : 1119 – 1129 .
- Havens , K E and Beaver JR . “ Forthcoming ” . In Composition, size and biomass of zooplankton in large productive Florida lakes , Hydrobiologia .
- Havens , K E , East , T L and Beaver , J R . 2007 . Zooplankton response to extreme drought in a large subtropical lake. . Hydrobiologia , 589 : 187 – 198 .
- Holdren , G C , Horn , M J and Lieberman , D M . 2006 . The limnology of Boulder Basin, Lake Mead, AZ-NV, 2005 report of findings , Denver , CO : US Bureau of Reclamation Technical Memorandum 8220-06-15 .
- Idrisi , N , Mills , E L , Rudstam , L G and Stewart , D J . 2001 . Impact of zebra mussels (Dreissena polymorpha) on the pelagic lower trophic levels of Oneida Lake, New York. . Can J Fish Aquat Sci , 58 : 1430 – 1441 .
- Kissman , C E , Knoll , L B and Sarnelle , O . 2010 . Dreissenid mussels (Dreissena polymorpha and Dreissena bugensis) reduce microzooplankton and macrozooplankton biomass in thermally stratified lakes. . Limnol Oceanorgr. , 55 ( 5 ) : 1851 – 1859 .
- Knoll , L B , Sarnelle , O , Hamilton , S K , Kissman , C EH , Wilson , A E , Rose , J B and Morgan , M R . 2008 . Invasive zebra mussels (Dreissena polymorpha) increase cyanobacteria toxin concentrations in low-nutrient lakes. . Can J Fish Aquat Sci , 65 : 448 – 455 .
- LaBounty , J F . 2008 . Secchi transparency of Boulder Basin, Lake Mead, Arizona-Nevada: 1990-2007. . Lake Reserv Manage , 24 : 207 – 218 .
- LaBounty , J F and Burns , N M . 2005 . Characterization of Boulder Basin, Lake Mead, Nevada-Arizona, USA - Based on analysis of 34 limnological parameters. . Lake Reserv Manage , 21 : 277 – 307 .
- LaBounty , J F and Burns , N M . 2007 . Long-term increases in oxygen depletion in the bottom waters of Boulder Basin, Lake Mead, Nevada-Arizona, USA. . Lake Reserv Manage. , 23 : 69 – 82 .
- LaBounty , J F , Holdren , G C and Horn , M J . 2004 . The limnology of Boulder Basin, Lake Mead, Arizona-Nevada: Report of Findings 1990-1998 , Denver , CO : US Bureau of Reclamation Technical Memorandum 8220-04-10 .
- LaBounty , J F and Horn , M J . 1997 . The influence of drainage from the Las Vegas Valley on the limnology of Boulder Basin, Lake Mead, Arizona-Nevada. . Lake Reserv Manage. , 13 : 95 – 108 .
- Lavrentyev , P J , Gardner , W S , Cavaletto , J F and Beaver , J R . 1995 . Effects of zebra mussels (Dreissena polymorpha Pallas) on phytoplankton and planktonic protozoa in Saginaw Bay, Lake Huron. . J Great Lakes Res. , 21 : 545 – 557 .
- Lawrence , S G , Malley , D F , Findlay , W J , MacIver , M A and Delbaere , I L . 1987 . Method for estimating dry weight of freshwater planktonic crustaceans from measures of length and shape. . Can J Fish Aquat Sci , 44 : 264 – 274 .
- Loomis , E M . 2009 . Trophic interactions associated with introduction of the invasive quagga mussel in Lake Mead, Nevada. MS Thesis. , Las Vegas (NV) : University of Nevada .
- Lynch , M . 1977 . Fitness and optimal body size in zooplankton populations. . Ecology , 58 : 763 – 774 .
- MacIsaac , G L , Lonnee , C L and Leach , J H . 1995 . Suppression of microzooplankton by zebra mussels: importance of mussel size. . Freshwater Biol , 34 : 379 – 387 .
- McCauley , E . 1984 . “ The estimation of the abundance and biomass of zooplankton in samples. ” . In A manual for the assessment of secondary productivity in fresh waters , Edited by: Downing , J A and Rigler , F H . 228 – 265 . Oxford , UK : Blackwell Sci .
- McGowan , S , Leavitt , P R and Hall , R I . 2005 . A whole-lake experiment to determine the effects of winter droughts on shallow lakes. . Ecosystems , 8 : 694 – 708 .
- McMahon , R F . 2007 . Analysis of the shell length distributions of quagga mussel (Dreissena rostriformis bugensis) taken from Lakes Mead and Mohave on the Colorado River drainage during Jan-Mar 2007 , US Bureau of Reclamation .
- Mills , E L , Rosenberg , G , Spidle , A P , Ludyanskiy , M , Pligin , Y and May , B . 1996 . A review of the biology and ecology of the quagga mussel (Dreissena bugensis), a second species of freshwater dreissenid introduced into North America. . Am Zool , 36 : 271 – 286 .
- Mueller , G and Horn , M J . 2004 . Distribution and abundance of the pelagic fish in Lake Powell, Utah and Lake Mead, Arizona-Nevada. . West N Am Naturalist , 64 : 306 – 311 .
- Nalepa , T F , Fanslow , D F and Lang , G A . 2008 . Transformation of the offshore benthic community in Lake Michigan: recent shift from the native amphipod Diporeia spp. to the invasive mussel Dreissena rostriformis bugensis. . Freshwater Biol , 54 : 466 – 479 .
- Ney , J J . 1996 . Oligotrophication and its discontents: effects of reduced nutrient loading on reservoir fisheries. . Am Fish Soc , 16 : 285 – 295 .
- Paulson , L J and Baker , J R . 1983 . Interrelationships among nutrients, plankton and striped bass in Lake Mead. , Las Vegas (NV) : University of Nevada . Lake Mead Limnological Research Center Technical Report 10
- Pennak , R W . 1989 . Fresh-water invertebrates of the United States: protozoa to mollusca. 3rd ed. , New York (NY) : John Wiley and Sons .
- Raikow , D F , Sarnelle , O , Wilson , A E and Hamilton , S K . 2004 . Dominance of the noxious cyanobacterium Microcystis aeruginosa in low-nutrient lakes is associated with exotic zebra mussels. . Limnol Oceanogr , 49 : 482 – 487 .
- Rellstab , C and Spaak , P . 2009 . Lake origin determines Daphnia population growth under winter conditions. . J Plankton Res , 31 : 261 – 271 .
- Ruttner-Kolisko , A . 1974 . Plankton rotifers, biology and taxonomy. . Die Binnengewasser , 26 : 1 – 146 .
- Sollberger , P J and Paulson , L J . 1991 . Littoral and limnetic zooplankton communities of Lake Mead, Nevada-Arizona, USA. . Hydrobiologia , 237 : 175 – 184 .
- Spidle , A P , Mills , E L and May , B . 1995 . Limits to tolerance of temperature and salinity in the quagga mussel (Dreissena bugensis) and the zebra mussel (Dreissena polymorpha). . Can J Fish Aquat Sci , 52 : 2108 – 2119 .
- Stemberger , R S . 1979 . “ A guide to rotifers of the Laventian Great Lakes ” . In Environmental Monitoring and Support Laboratory , Cincinnati , OH : U.S. Environmental Protection Agency .
- Stoeckmann , A . 2003 . Physiological energetics of Lake Erie dreissenid mussels: a basis for the displacement of Dreissena polymorpha by Dreissena bugensis. . Can J Fish Aquat Sci , 60 : 126 – 134 .
- Twichell , D C and Cross , V A . 2004 . “ Mapping the floor of Lake Mead (Nevada and Arizona); preliminary discussion and GIS data release ” . In US Geological Survey Open-File Report 03-320 http://pubs.usgs.gov/of/2003/of03-320/. Accessed 19 May 2009
- Twichell , D C , Cross , V A , Rudin , M and Parolski , K F . 1999 . Surficial geology and distribution of post-impoundment sediment of the western part of Lake Mead based on a sidescan sonar and high-resolution seismic-reflection survey US Geological Survey Open-File Report
- [USACE] US Army Corps of Engineers . 2002 . Zebra Mussel Information System (ZMIS) , USACE. Engineer Research and Development Center . http://el.erdc.usace.army.mil/zebra/zmis. Accessed 2 Jan 2007
- Vanderploeg , H A , Joengen , T H and Liebig , J R . 2009 . Feedback between zebra mussel selective feeding and algal composition affects mussel condition: did the regime changer pay a price for its success? . Freshwater Biol , 54 : 47 – 63 .
- Vanderploeg , H A , Liebig , J R , Carmichael , W C , Agy , M A , Johengen , T A , Fahnenstiel , G L and Nalepa , T F . 2002 . Zebra mussel (Dreissena polymorpha) selective filtration promoted toxic Microcystis blooms in Saginaw Bay (Lake Huron) and Lake Erie. . Can J Fish Aquat Sci , 55 : 1208 – 1221 .
- Vanni , M J . 1986 . Competition in zooplankton communities: suppression of small species by Daphnia pulex. . Limnol Oceanogr , 31 : 1039 – 1056 .
- Wilde , G R . 1984 . Seasonal and spatial heterogeneity in the limnetic zooplankton community of Lake Mead. MS thesis , Las Vegas (NV) : University of Nevada .
- Wilde , G R and Paulson , L J . 1988 . Food habits of young-of-year largemouth bass in Lake Mead and Lake Mohave, Arizona-Nevada. . Great Basin Nat , 48 : 485 – 488 .
- Wilde , G R and Paulson , L J . 1989 . Temporal and spatial variation in pelagic fish abundance in Lake Mead determined from echo grams. . Calif Fish Game. , 75 ( 4 ) : 218 – 223 .
- Wilson , A E and Sarnelle , O . 2002 . Relationship between zebra mussel biomass and total phosphorus in European and North American lakes. . Arch Hydrobiol , 153 : 339 – 351 .
