Abstract
Recently Shen et al. have argued that the Ediacaran faunas from Avalon-Charnwood [580–560 million years ago (MA)], the White Sea-Flinders Range (560–550 MA) and Namibia (550–543 MA) occupied the same morphospace even though these faunas differed in species composition, ecology, biogeography and age. The traits they used to characterise these faunas could not distinguish between important promorphological features such as radial vs. bilaterian and unitary vs. colonial animals. Their inappropriate assignment of symmetry properties led to the homogenisation of morphospace in these different faunas. Another way to examine shifts in morphospace involves sorting out radial vs. bilaterian and unitary vs. colonial genera in terms of their time of first appearance in the fossil record. While genera with both kinds of symmetry properties and unitary and colonial animals were present during the early Ediacaran, there was a large proportional increase in new bilaterian genera and a decrease in colonial genera beginning between 560–550 MA. The increase in bilaterians may have been associated with the ability to exploit food sources in and on the substrate of shallow water environments.
Introduction
Ediacaran animal fossils have a wide distribution and vary in age from about 620 million years ago (MA) to the beginning of the Cambrian (543 MA). There are four major assemblages: the Yangtze Gorge (620–543 MA), Avalon-Charnwood (580–560 MA), the White Sea-Flinders Range (560–550 MA), and the Nama (550–543 MA) in addition to a number of smaller assemblages ((Fedonkin et al. Citation2007a). In a recent study, Shen et al. (Citation2008) have argued that three of these assemblages, the Avalon-Charnwood, the White Sea-Flinders, and the Nama occupied similar morphospaces even though they differed to a large extent in species composition, ecological setting, biogeography and age. This claim is questionable in view of the fact that there was only one putative bilaterian in the Avalon-Charnwood fauna and a significant increase of new bilaterians starting after 560 million years (Narbonne Citation2005). If a morphospace analysis cannot distinguish between motile bilaterian taxa and tethered colonial taxa it is not doing its job.
This study re-examines the criteria Shen et al. (Citation2008) used to collect and categorise their data in order to expose the pitfalls inherent in the information used for their morphospace analysis. It also documents the appearance of bilaterian genera and the decline in colonial genera during the Ediacaran to provide a quantitative assessment of the changes in relative generic numbers in these subgroups but also in their morphological disparity during this period. These data indicates that there was a shift in morphospace during the Ediacaran associate starting between 550–560 MA.
Methods
Time of first appearance of Ediacaran genera and their geological setting
A number of procedures have been used to infer time of first appearance of Ediacaran genera. One or more formations at a given site frequently have an absolute date. The most trustworthy dates are based on U–Pb zircon dating associated with volcanism; however, some of these dates have a large standard error. A few dates are based on Pb–Pb isotope ratios; these measurements are inherently less accurate and generally have a larger standard error than U–Pb dates. If a fossil is in a formation that is bracketed by two relatively close dates, one can be fairly confident of its date. A fossil can be in a formation that is above or below a dated formation; in this case, the date is less certain. In cases where there is not a good date for a site but where elements of the fauna are similar to a dated site, the dated site has sometimes been used to assign a date to the site where there is a paucity of information. In some cases, the fossil site may lie immediately under a formation with characteristic Cambrian fossils. Because the base of the Cambrian is dated, these Ediacaran fossils can be dated with a fair degree of certainty. Dating of fossils in Table is done in most cases within 10 million year intervals. Figure gives the geographical sites and formations, where the fossils listed in Table first appeared and indicates the inferred age of these formations. If a date is fairly certain, it is given superscript ‘a’; if it is less certain, it is given superscript ‘b’ and if it is an educated guess, it is given superscript ‘c’ in Table .
Table 1. Time of first appearance, and type of Ediacaran genus.
Figure 1 Time line showing the ages of the sites where Ediacaran fossils were collected and where the age of the deposit has been established by measuring the products of radioactive decay. An ‘ × ’ marks the U–Pb zircon dates and a ‘0’ marks the Pb–Pb dates. The superscipt identifies the authors who established the dates: (1) (Condon et al. (Citation2005), (2) Zhang et al. (Citation2005), (3) Barford et al. (Citation2002), (4) Chen et al. (Citation2004), (5) Bemus (Citation1988), (6) Bowring et al. (Citation2003a), (7) Bowring et al. (Citation2003b), (8) Compston et al. (Citation2002), (9) Martin et al. (Citation2000), (10) Grazhdankin (Citation2004), (11) Preiss (Citation2000) and (12) Grotzinger (Citation1995). A solid line indicates that the date for a given formation or a member in the formation is relatively secure; while a dashed line indicates that the dating is not secure.
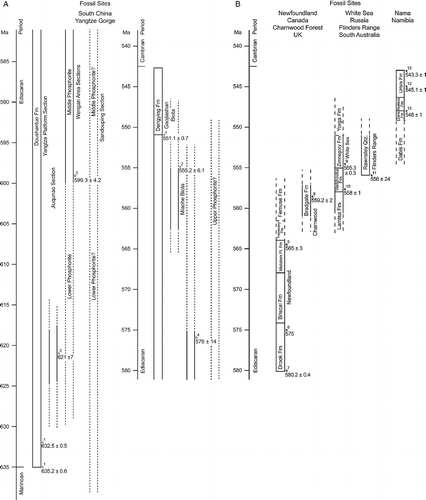
The Ediacaran fauna in the Yangtze Gorge is found in two formations. The upper Dengying Formation extends from the base of the Cambrian to about 551 MA (Condon et al. Citation2005; Zhang et al. Citation2005). The Doushantuo Formation extends from the base of the Dengying formation to about 635 MA. The Marinoan glaciation, which was a global event, was coming to an end at the base of the Doushantuo Formation (Condon et al. Citation2005; Zhang et al. Citation2005). All of the fossiliferous sites in both formations appear to have been shallow marine basins. Fossils found primarily in the lower part of the Dengying Formation are referred to as the Gaojiashan biota; a number of these fossils have thin mineralised shells. There are three fossil containing phosphorite strata within the Doushantuo Formation. At these sites, the soft parts of organisms have been phosphatised. Preservation can be superb allowing one to see tissues and cellular detail; this is the Ediacaran Lagerstätten. Anywhere between 1 and 3 phosphorite containing strata are found in all of the extended sections of the Doushantuo Formation that have been examined. The upper Phosophorite unit is frequently referred to as the Miaohe biota and lies just below the Dengying Formation. Its upper boundary is well defined, however, its lower boundary has not been determined; it may extend to 560 MA. At Weng'an the Doushantuo Formation can be about 40 m thick (Dornbos et al. Citation2006). In some sections, the region just above the Middle Phosophorite unit has been dated at 576 MA and the region just below this unit has been dated at 599 MA (Barford et al. Citation2002; Chen et al. Citation2004); the dating of this part of the formation is poorly constrained. The upper part of the Basal Phosphorite unit terminates some time before the Middle Phosphorite unit and begins after 621 MA. The dating of this unit is even less well constrained than the middle unit. Another set of fossiliferous sections associated with Sandouping is 188 m thick (Tang et al. Citation2006); none of its fossiliferous units have been dated, however all three phosphoirite containing strata are present.
The Avalon fauna occurs in strata on the east coast of Newfoundland and an almost identical fauna is found in Charnwood Forest in the English midlands. The Ediacaran is truncated at the basal end of the Drook Formation by the Gaskiers glaciation (ca. 580 MA), which appears to have been a regional event. Four dates constrain the ages of these sites, including the Drook (Bowring et al. Citation2003a, Citation2003b), Mistaken Point (Bemus 1988) and Bradgate Formations (Compston et al. Citation2002). The top ends of the Bradgate and Femeusse Formations are not constrained. It has been assumed that there is a partial chronological overlap between one or more formations in the Avalon fauna and the Charnwood fauna. Ecologically, both of these faunas existed in relatively deep water below the photic zone on the continental shelf (Narbonne Citation2005). All of the fossils at these sites are casts.
The Russian White Sea and Australian Flinders Range faunas are also quite similar; both faunas lived in shallow near-shore environments; all of the fossils at these sites are casts. There are two dates that constrain the age of the Verkhovka Formation; at the same time they define the top of the Lamtsa and the base of the Zimnegory Formations (Martin et al. Citation2000; Grazhdankin Citation2004) at the White Sea sites. The date of 556 MA defines the base of the Rawnsley Quartzite in the Flinders Range (Preiss Citation2000); however, this date has a large standard deviation associated with it. The top of the Rawnsley Quartzite and the Yorga Formations are not constrained. The fact that there is a significant overlap of the White Sea and Flinders Range faunas suggests that they were largely contemporaneous.
The Nama fauna of Namibia also lived in a shallow water near shore environment; the fossils at this site are either casts or composed of shell. It is constrained by three dates (Grotzinger et al. Citation1995). The upper end of the Urusis Formation, which abuts the lower Cambrian is dated at 543 and 545 MA and the base of the Nudaus and boundary of the Zaris Formation is dated at 548.8 MA. The bottom of the Dabis, which is the oldest formation, is not constrained by a date. At both the Yangtze Gorge and Nama sites there is a shelly index fossil, Cloudina that is only found in the strata just below the Cambrian boundary (Geyer and Uchman Citation1995). At Nama this genus extends from the top of the Urusis to the Dabis Formation (Vickers-Rich Citation2007).
There are additional sites where dates are not available, but the formation lies just below the Cambrian and Cloudina is present. These include the Blueflower Formation in the Canadian northwest, the Lower Woods Formation in the great basin of the US, the Tamango Formation of Brazil, the Yerbal Formation of Uruguay and the Deschiling Formation of Bhutan.
Frequently, a given Ediacaran genus is found at more than one stratigraphic level at a given site or at multiple sites. An atlas of Precambrian metazoans in (Fedonkin et al. Citation2007a) was used in Table to establish the earliest geographic site and formation of a given genus. The earliest record for a given genus does not always provide the best description of the genus, as a consequence, the reference for the genus is frequently based on material that dates from a later horizon.
Morphological characterisation of Ediacaran genera
Fossils were characterised as either unitary individuals or colonies composed of multiple individuals (zooids). Unitary individuals or zooids that make up a colony can be either radially or bilaterally symmetrical. A radially symmetrical individual has only one axis of symmetry which is frequently referred to as its anterior–posterior axis. Bilaterally, symmetrical individuals have an anterior–posterior axis and a plane that bisects the dorsal and ventral region that extend along the anterior–posterior axis; this plane divides the organism into left and right halves along its anterior–posterior axis. Extant radially symmetrical animals are composed of two cell layers; an outer epithelium and a gastrodermis. Extant bilaterians are composed of an outer epidermis, a gastrodermis and an intermediate tissues layer between them, the mesoderm that forms the muscles and the excretory system. The distinction between radial and bilateral symmetry is robust. During embryogenesis in extant bilaterians, the anterior–posterior axis is always specified prior to the specification of the dorsal–ventral axis (Slack Citation1991).
Three classes of fossils make up the Ediacaran fauna: body fossils, trace fossils and embryos. Body fossils provide the most information on the morphology of a given organism. Trace fossils of trails made by animals moving through a substrate or feeding have frequently been used to assign that fossil to the Bilateria because it takes muscle, a mesodermal derivative, operating on a hydrostatic skeleton to make a track. In a number of cases, the body fossil that was responsible for making a given trace is present with its trace. However, there are circumstances involving living animals where traces have been generated by an organism that was not a bilaterian (Jensen et al. Citation2005; Bengston and Rasmussen Citation2009). Two additional problems are associated with trace fossils: (1) short morphologically simple traces may be artifacts and (2) because trace fossils have a relatively simple morphology it is easy to confuse one ichnogenus for another. Jensen et al. (Citation2006) have compiled and assessed the published records on Ediacaran trace fossils. Almost all of the trace fossil citations in Table are based on their records. There are plausible claims for cleavage stages, gastrula stages and larval fossils (Li et al.Citation1998; Chen et al. Citation2000; Chen et al. Citation2006). However, in most cases these stages do not contain enough information to indicate whether the developing animal would be radially or bilaterally symmetrical. Because a fossil of a ’gastrula’ is essentially a snapshot of an individual at a given stage of development, that particular fossil may only have two germ layers while at slightly later stage of the development that species may have three germ layers. This would change the diagnosis of the embryo from a radial to a bilaterian form. Usually, asexual reproduction does not begin until after embryogenesis is over; therefore it cannot be diagnosed on the basis of an embryonic stage.
A given fossil was only used in Table , if it could be dated with some degree of confidence, had characters that could be used to distinguish unitary and colonial forms and a statement could be made about its symmetry properties.
For each genus a reference to a figure showing a typical fossil and description of the genus is given. If there was more than one species in the genus, the species used was named. The reference was used to obtain an approximate measure of the size of an average member of a genus. When no measurements were given in the reference, the length and width or diameter of the fossil from the figure was measured where possible. This is only an approximate measure of the size of a typical member of a genus since different species within a genus can differ in size. Within a species there are also size differences within a population; in a number of cases these size differences represent different growth stages. In colonial organisms this measurement was made on the size of the largest zooids in the colony. The habitat of each genus: sessile, motile or pelagic is also given and the approximate number of specimens of that genus that have been examined. A number of genera are only known from one or a small number of specimens; one has less confidence in a generic description based on a small specimen number. In the categories Morphology and Habitat in Table , ‘a’ indicates that the judgment is supported by the fossil evidence, ‘b’ indicates a judgment that is only partially supported by the evidence and ‘c’ indicates a judgment that is an educated guess.
Results
Critique of the work of Shen et al. (2008)
The data set for the Shen et al. (Citation2008) paper was published as supporting online material. Fifty morphological characters were used to quantify the symmetry properties and morphology of species from three Ediacaran faunas. The characters used (# in parenthesis) delimited overall shape (6), symmetry (7), structure of the central region (23), structure of the intermediate region (7), structure of the peripheral region (3) and other structures (4). The two most important character sets are overall shape and symmetry because they are used to characterise every fossil.
There is a formal language zoologists use to characterise the symmetry properties of animals and there is simple descriptive language. Shen et al. (Citation2008) have opted for simple descriptive language. For example, the symmetry term ‘radial’ means going out from a centre, while ‘bilateral’ indicates extending out on each side of a median line. For zoologists an extant multicellular animal is either radially symmetrical or it is a bilaterian (Beklemishev Citation1969). These categories also accommodate the Ediacaran fauna (Fedonkin Citation1990a). A radially symmetric animal has an anterior–posterior axis; each of these axial landmarks differs in morphology and the animal is symmetrical along the axis separating the two end points. Bilaterally, symmetrical animals have an anterior–posterior axis in addition to dorsal and ventral sides that differ and extend along this axis. The plane that defines the dorsal–ventral sides also defines the left and right sides of the animal. When descriptive language is used, the term bilateral refers to an organism that has left-right symmetry but does not take into account dorsal–ventral differences. One problem involved in using the informal language of the authors to describe fossils is that the description is not commensurate with a statement about whether or not a fossil is radially symmetrical or a bilaterian. The morphological characters necessary to make a distinction between a radially symmetrical animal and a bilaterian are not included in the authors data matrix and they do not discuss these morphological categories.
Another problem in Shen et al. (Citation2008) is based on their treatment of unitary individuals and colonies composed of zooids that were generated by asexual reproduction. All of the individuals in a given colony can be either radially or bilaterally symmetrical, however, in almost all Ediacaran colonies, while one can locate where the zooids were, in most cases it is not possible to specify their symmetry properties because of their small size. The symmetry properties of a colonial species are defined by the symmetry properties of its zooids. While the shape of the colony as a whole can be described as having a given kind of symmetry, these symmetry properties may not be the same as the symmetry properties of the zooids that make up the colony. Shen et al. (Citation2008) do not make a distinction between colonies and unitary individuals even though colonies make up a significant part of the Ediacaran biota (Fedonkin Citation1990b). The symmetry properties of the whole colony are treated as if they were equivalent to the symmetry properties of a unitary individual. Since many colonial species have a bilateral colony morphology, they are lumped with disparate unitary bilaterians that also exhibit bilateral symmetry. This effectively makes their morphospaces similar.
In addition to these major methodological errors, there are categories of information that the authors have purposely excluded that would help to establish differences between radial and bilaterian taxa. Most bilaterians are capable of locomotion, while locomotion on substrates in radially symmetrical animals either does not occur or is restricted. There are a number of cases where a trail is coupled with a fossil animal. This kind of information can be used to define the front end of the animal in cases where both ends appear to be the same. There are also a number of structural differences that emerge when both the dorsal and ventral sides of a given fossil are examined and internal structures can become visible in compressed fossils that are potentially useful for establishing dorsal–ventral and anterior–posterior polarity differences. Different growth stages of a fossil genus can be used in order to make inferences about the generation of structural features from a growth zone.
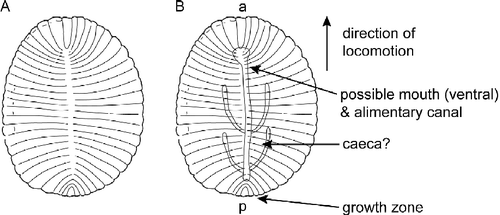
These excluded categories of information can change the morphospace diagnosis of a fossil. One example involves the bilaterian Dickinsonia (Figure ). According to Shen et al. (Citation2008), this genus is defined by the following character states: overall shape – oval, symmetry – bilateral and bipolar, and structures in the central region – central groove plus primary branches (segments) curved toward both ends. As Dickinsonia moved it could leave resting traces on its substrate. The set of resting traces that follow a body fossil are oriented in the same way as the body fossil allowing one to infer that the body fossil moved with only one bilateral end in front (Gehling et al. Citation2005). As Dickinsonia grew it added superficial segmental structures to its external epithelium. Segments are only added to one end of the animal (Runnegar Citation1992). This end is opposite the front end of the animal during locomotion. Studies of compressed specimens have revealed structures interpreted as an anterior ventral mouth-pharynx that connects to an alimentary canal (the central groove of Shen et al. Citation2008) and paired caecae that extend anterior laterally from the alimentary canal (Jenkins Citation1992; Dzik and Ivantsov Citation2002). Some of these fossils show characteristic wrinkles on part of their surface, indicating muscle contractions, muscle is indicative of bilaterian status (Runnegar Citation1982). This additional information shows that the animal had an anterior–posterior axis and that there are dorsal–ventral differences along this axis; in other words it was a bilaterian (Figure ). It is important to note that Shen et al. (Citation2008) would have not been able to infer that this genus was a bilaterian because they do not use a character set for defining dorsal–ventral differences.
Figure 2 A) Drawing of Dickinsonia showing the bipolar features that were used by (Shen et al. (Citation2008). (B) Drawing of Dickinsonia showing that it has an anterior (a)–posterior (p)-axis, and dorsal and ventral sides based on inferences about the polarity of locomotion, the pattern of growth and the anatomy of internal and ventral organs.
In a number of cases, separated parts of an Ediacaran body fossil have been assigned to different taxa. This was a frequent occurrence for sessile colonial forms where the holdfast that tethers the colony to the substrate was preserved while the frond-like structures with the zooids was lost. After this condition was recognised (Gehling et al. Citation2000) a number of disc shaped fossils were identified as holdfasts (see atlas of Precambrian fossils in Fedonkin et al. Citation2007a). Shen et al. (Citation2008) have included these holdfasts in their morphospace analysis. There are 32 cases that were used in Shen's analysis of the Avalon fauna, 31% of these cases are holdfasts; there are 195 cases in their White Sea-Flinders Range fauna, 36% of these are holdfasts; there are 32 cases in the Nama fauna, 32% of them are holdfasts. Since holdfast morphospace is rather uniform it dampens the impact of the morphospace values of intact fossils.
Time of first appearance of Ediacaran genera
One can get a picture of the changes in the Ediacaran fauna through time and changes in morphospace by noting when members of this fauna first appear in fossil record and the promorphological nature of these genera (Table ).
600–615 MA: There are two fossils from the lowest fossiliferous unit of the Doushantuo formation. One fossil is a crown group poriferan. Thin sections of these fossils show monaxial spicules with associated sclerocytes. There is an epidermis with porocytes and an internal spongocoel. There is suggestive evidence that these animals were engaged in asexual reproduction. These fossils were found in association with embryos and a possible parenchymella type sponge larva. Recent work on the distribution of a sponge biomarker in Ediacaran and Cryogenian strata has provided evidence for the appearance of sponges just prior to the Marinoan glaciation at ca. 635 MA (Love et al. Citation2009).
The other fossil resembles the ichnogenus Planolites. This type of trace is found on sediment surfaces. Its round cross-section suggests peristaltic locomotion which is associated with a bilaterian body plan. Given the fact that there is only one representative of this ichnogenera it is not a compelling case for an early Ediacaran bilaterian.
580–590 MA: This time interval contains a crown group cnidarian body fossil that resembles part of a modern thecate hydrozoan polyp colony (Chen et al. Citation2002). Two zooids are connected by a stolon. Underneath the perisarc there appears to be an epithelial layer that surrounds a gastric region. There are also embryos with two germ layers associated with the body fossils that resemble some hydrozoan gastrulae, however, these gastrulae do not have enough diagnostic features to give them a phylum identity. Chen et al. have also described the fossil Ramitubus (which Chen et al. refer to as Sinocylocyalicus). This tubular fossil, which can reproduce asexually, is composed of a series of compartments enclosed in an extracellular matrix. Only the terminal compartment fed. In one case, two tentacles emerge from the top compartment of a sectioned fossil. The skeleton of this fossil has the same organisation of a tabulate coral, however, the evidence that it represents a crown group Anthozoan cnidarian is not as compelling as it is in the case of the hydrozoan because the body of the fossil was not sectioned even though it can be seen in outline. Rambitubus has also been found in younger strata in the Doushantuo Formation that comprise the Miaohe biota (Xiao et al. Citation2002). Another genus, Funsia (Droser and Gehling Citation2008) from the Ediacara of Australia appears to be an enlarged version of Rambitubus.
There are cases involving a body fossil, a trace fossil, and embryos during this time interval that indicate the presence of bilaterians. The most convincing case involves the body fossil Vernanimalcula (Chen et al. Citation2004a). A population of these small fossils has been examined in section. They have three germ layers, an anterior mouth and a through gut. They have clear-cut anterior–posterior a dorsal–ventral axes and anterior dorsal structures that may have had a sensory function. Bengston and Budd (Citation2004) have argued that this fossil genus is an artifact on the basis of both taphonomic and diagenetic alterations. Chen et al. (Citation2004b) replied that they have a number of cases that show the complex morphology that they report and as a consequence the possibility that one is dealing with an artifact is slim.
The trace fossil ?Cruziana has parallel edges and faint transverse ridges at the bottom of a shallow longitudinal furrow. Only one case of this ichnogenus has been found. It resembles Cruziana, a trace fossil, typical of the early Cambrian, that is thought to have been generated by an arthropod (Jensen Citation2003).
One class of cleavage stage fossil embryos has been found with polar lobes at the 2 and 4 cell stages of early embryogenesis (Chen et al. Citation2006). The polar lobe is a transitory structure that forms from the vegetal pole of the egg during cleavage and shunts cytoplasm to one of the blastomeres at the four-cell stage. The descendants of this cell play a major role in setting up the dorsal–ventral axis of the embryo in some genera of extant lophotrochozoan animals (Freeman and Lundelius Citation1992). These embryos appear to be derived from different species because the egg sizes of embryos producing polar lobes vary.
In addition to Ramitubus, there are three other tubular genera, Sinocyclocylicus, Quadratitutbus and Crassitubus that first make their appearance during this period. Sinocyclocylicus and Quadratitutbus have internal cross walls; however, in these three genera it is not clear if the animals that lived in the tube had radial or bilateral symmetry. All three genera were probably sessile.
570–580 MA: Four colonial and one unitary genera appeared during this period, all of these genera were sessile. In colonial forms, there is a central vane; primary branches come off the central vane, secondary branches come off the primary branches, third order branches can come off the secondary branches and zooids (frondlets) form on third order branches. (See Brazier and Antcliffe (Citation2009) for the nomenclature used to describe branching and Narbonne et al. (Citation2009) for photographs of zooids of these frond-like animals.) Because of the small size of the zooids one cannot say with certainty whether or not they are radially or bilaterally symmetrical. Trepassia was formerly called Charnia wardi. These frond-like colonial animals are referred to as rangeomorphs. Thectardis has a cone-like morphology with an attachment site at the apex of the cone. Other genera with this morphology appear later in the Ediacaran fossil record.
560–570 MA: During this period one bilaterian and one radially symmetrical unitary genus appeared in the fossil record. Epibaion was a large flattened bilaterian that presumably made its living moving over and grazing on surfaces. Feeding trails left behind by these animals, directly adjacent to a body fossil help to define their anterior–posterior axis. (This genus resembles Dickinsonia which is found in the 550–560 MA interval.) It appears that during life the base of Inaria was partly covered by the substrate and there was an apical extension of the animal with a retractable feeding apparatus that probably had a tentacle-like morphology. The other seven genera were sessile colonial organisms with a rangeomorph body plan with zooids distributed along their terminal branches; it is not clear whether their zooids were radially or bilaterally symmetrical.
550–560 MA: During this period 22 bilaterians, 15 unitary genera that do not have the overt characteristics of bilaterians and eight colonial genera appeared in the fossil record.
Two of the bilaterians appear to be members of stem lineages that may have evolved to form extant phyla. Parvanocornia has a dorsal shield that covers the body; there is a ventral median region that paired ?limbs emerge from; it closely resembles and has a growth trajectory similar to the Cambrian arthropod Skania (Lin et al. Citation2006). Another bilaterian, Kimberella, is a putative stem group mollusc (Fedonkin et al. Citation2007c). This organism has a stiff non-mineralised dorsal shell with a characteristic ornamentation. A ventral foot is present; locomotory traces are associated with the foot in some specimens. There is an anterior proboscis that could be extended beyond its shell; there can be feeding traces associated with some animals suggesting the presence of hard claws or a rasp-like feeding structure associated with the proboscis. The animal has an internal axial structure interpreted as an alimentary canal. In the groove between the shell and the foot there are bilaterally arranged ridges on each side of the animal that could have functioned in respiration.
The affinities of the other bilaterians to modern animal phyla are more problematic. There have been a number of attempts to assign Ediacaran genera to groups on the basis of their characters; however, there have been no formal cladistic studies of these animals. A group of bilaterians that first appear in the fossil record at this time had an anterior head region and a set of somites which made up the posterior part of their body and are referred to as Vendomorpha. The genera included in this group include, Cyanorus, Vendia, Karakhtia, Onega, Pseudovendia, Spriggina, Lossinia, Armillifera,Yorgia Archaeaspius and Praecambridium. Some of the members of this group including Cyanorus and Vendia had a tubular digestive system that ran along the animal's anterior–posterior axis with bilateral caeca that extended from the digestive tube to the somites that make up the posterior part of the body. Yorgia left locomotory traces. Temnoxa which had a head region and an unsegmented body region probably also belongs to this group.
Another group of bilaterians are referred to as the Dipleurozoa; it includes Dickinsonia, Andivia and Solza from the interval 550–560 MA and Epibaion from the interval 560–570 MA. In these species, cephalisation is minimal. The organisation of Dickinsonia has previously been discussed in the critique of the work of Shen et al. Andivia and Solza had non-mineralised dorsal shells and a ventral foot. There are five worm-like genera; four of these genera are trace fossils. Another genus labelled ‘bilaterian’ has a through gut with a bulb at one end and appears to have been in the process of making a locomotory trace.
The unitary genus Palaeophragmodictya is a poriferan with a reticulate mesh of spicules and a centrally placed osculum. There is a bag-like unitary fossil Bonata that appeared to be attached to and partly covered by the substrate. Inaria from 560–570 MA has a similar morphology; it was also embedded in the substrate. There is a set of disc shaped genera that may have been attached to the substrate that had higher order radial symmetry; five genera, Albumares, Anfesta, Skinnera, Triforillonia and Tribrachidium exhibited triradial symmetry, one low cone-shaped genus, Conomedusites exhibits tetraradial symmetry and one genus Arkarua exhibits pentameral symmetry. Vendoconularia has a conical form and resembles Thectardis from the interval 570–580 MA. Three disc-like genera with tentacles may have been pelagic: Veprina, Eoporpita and Rugoconites. Both Calyptrina and Protoconites lived in tubes so that it is not possible to say if they were radially symmetrical or bilaterians.
Ramellina and Gehlingia are colonies with multiple zooids along their arms that extend from its central axis. The rangeomorph colonies that first appeared between 560 and 580 MA are also organised in this way. Pteridinium, Pambikalbae, Phyllozoon and Swartpuntia have a central axis with vanes made up of double rows of closely packed tubular elements extending out of it. The ends of these tubular elements furthest from the insertion of the vanes on the axis are open and presumably contained the zooids. While the sessile tubular colonial genus Funisia is much larger than Ramitubulus it had the same body plan and reproduced asexually in the same way. Ventogyrus is a polar form composed of three radial modules organised around an axis. Each module has a large basal chamber and is divided into two lateral compartments that are divided into smaller chambers of zooids. Because of the orientation of members of this genus and their organisation they are thought to have been pelagic. The zooids of these colonial animals could either be radially or bilaterally symmetrical.
543–550 MA: Thirteen bilaterians, four unitary genera with radial symmetry and one colonial genus with radial symmetry made their appearance in the fossil record during this period. In addition, seven unitary tubular genera, two colonial tubular genera and four colonial genera appeared; in these cases, it is not clear if the organisms were radially symmetrical or bilaterians One new trait that appeared during this period was biomineralised shells.
Among the bilaterians the genera Paravendia and possibly Tamga were vendomorphs. Windermeria and Ivovicia were members of the Dipleurozoa. Platypholinia and Vladimissa were flatworm-like and might be considered as dipleurzoans. Archaeichnium was a tubular fossil with a wall composed of grains of sediment; it left locomotory trails. Protechiuris was an elongated body fossil with a narrow spatulate anterior end, a flattened lobe on one side and a convex lobe on the other side. There are five trace fossils; three are from the Blue Flower Formation in Northwest Canada. Two of these trace fossils, Palaeophycus and Torrowangea, are also present at Nama where these trace fossils are associated with the late Precambrian index fossil Cloudina.
All four of the unitary radially symmetrical genera, Nemiana, Ernietta, Vaveliksia and Namacalathus were sessile. Nemiana lived in close proximity to its conspecifics. Ernietta was partly buried in its substrate. Namacalathus are relatively small animals, the body of which is connected to a stalk attached to the substrate; parts of the animal were mineralised. Namapoikia is a cnidarian that built large calcareous reefs as such it is a radially symmetrical genus that reproduced asexually. Valveliksia has a holdfast and an elongated radially symmetrical bag-like body.
The unitary tubular genera are: Soldadotubulus, Onuphionella, Sinotubulities, Gaojiashania, Ningqiangella, Conotubus and Chenella. Sinotubulities lived in a mineralised tube that was open at both ends (Chen et al. Citation2008). The two colonial tubular genera, Waltheria and Cloudina also had mineralised shell walls. These tubular genera could have been either bilaterally or radially symmetrical animals.
Three of the colonial frondose fossils, Paracharnia, Rangea and Nasepia were rangeomorphs. Corumbella had vanes composed of tubes attached to a central axis with zooids at the end of each tube; its organisation is similar to Pteridinium and Swartpuntia.
Discussion and conclusions
Shen et al. (Citation2008) have reported that the full range of Ediacaran morphospace was established in the Avalon fauna and that these morphospace parameters remained essentially unchanged in the younger White Sea-Flinders range and Nama faunas. They have also suggested that there was a special set of circumstances analogous to those that might have caused the Cambrian explosion that generated the Avalon fauna.
The morphospace analysis that Shen et al. (Citation2008) did was flawed because the character set used was selected without considering the possible promorphologies of the animals whose fossils were being examined. They made no attempt to distinguish between unitary and colonial animals. The symmetry characters they used could not distinguish between animals that are radially symmetrical and bilaterians. The authors make the statement that they ‘make no inference on phylogenetic homology and functional biology of the coded characters’; this approach led to data sets that were ambiguous because a given character state could have different meanings in different contexts.
Three practices the authors used tended to homogenise the morphospace envelopes for the fauna's they analysed. (1) A given part of a fossil, such as a holdfast, if it was separated from the rest of the fossil, was treated as a species even though it had a much simpler morphology than the animal it came from. (2) A given genus was used to construct the morphospace of a younger fauna, even if it had already appeared in an older fauna. This carry-over tends to make the faunas similar and masks morphospace change. (3) Even though the Shen et al. paper is primarily concerned with the Avalon-Charnwood, White-Sea-Flinders and Namibia faunas, data was also collected on the faunas from the Makenzie Mountains, Great Basin, British Columbia, Wernecke Mountains, Olenek uplift, Finmark and Podolia. The only fauna excluded from the study was from the Yangtse Gorge. The reason given for excluding this fauna was its compressed state. Much of the Yangtse Gorge fauna is three-dimensional and is better preserved than the classic Ediacaran assemblages; it includes the oldest and most informative Ediacaran fossils.
When the appearance of new Ediacaran genera is plotted through time in ca. 10 million year bins, between 570–615 MA only a relatively small number of genera appeared compared to later in the Ediacaran Period. This could indicate inadequate sampling; it could also reflect a low intrinsic level of genus origin. Of far greater importance is the appearance of representatives of two radially symmetrical crown group phlya: Porifera and Cnidaria, prior to the appearance of Avalon fauna fossils. There is evidence for new genera in these phyla at later points in the Ediacaran period. Molecular phylogenetic analyses (Dunn et al. Citation2008) indicate that radially symmetrical phyla are the sister group of the bilaterians. There are several lines of evidence, body fossils, trace fossils and embryonic stages, that bilaterians were present during this early period; however, a significant number of bilaterian genera did not appear in the fossil record until 550–560 MA and crown group bilaterians probably did not appear until the Cambrian.
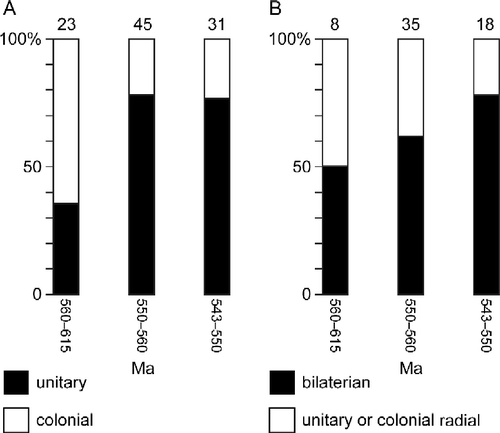
The increase in the proportion of new bilaterian genera and the decrease in colonial genera during the Ediacaran period are shown in Figure . The bar on the left side of each set of graphs covers a time period that is 4.5 times as long as each bar on the right. The graphs on the left (A) shows the relative increase in new unitary genera and the decrease in colonial and tubular genera with time. Since colonial and tubular genera can be either radially or bilaterally symmetrical the graph at the right (B) shows the percentage increase in new bilaterian unitary genera and the relative decrease in radially symmetrical unitary genera and colonial genera that are known to be composed of radially symmetrical zooids. During the periods 615–560 to 550–543 MA, the percentage of new bilaterian genera went from 50 to 72% and the percentage of colonial genera went from 65% to less than 20%. The change from a radial to a bilateral body plan and from a colonial to a unitary body plan are each major changes in morphospace. One does not need 50 characters to define Ediacaran morphospace; a few characters that will distinguish between the promorphological features of body plans will do.
Figure 3 A) The percentage of new genera in each time interval that are either unitary or colonial. Both categories may include some bilaterians. (B) The percentage of new bilaterian genera at each time interval included in radially symmetrical unitary and colonial genera with radially symmetrical zooids. The top of each graph gives the number of genera.
The new bilaterian genera that originated between 560 and 543 MA exhibited a variety of different morphologies that could be characterised in part as vendomorphs, Dipleurozoa with forms covered with a dorsal carapace and worms. Given the genesis of possible stem group Arthropoda and Mollusca during this period, possible groups would have to be around that the precursors of these extant phyla would evolve from; this was certainly the case. At the same time there was a marked increase in the size of bilaterians even though small forms still persisted (Table ).
Why was it that the proportion of new bilaterian genera did not begin to increase significantly until 560–550 MA? There were two sources of food for Ediacaran animals: bacteria, small unicells and multicellular organisms in the pelagic environment and (2) benthic organisms on and in the substrate (Narbonne Citation2005). The first animals to form extant phyla were the Porifera and Cnidaria. These animals were filter feeders (Porifera) and possible passive trappers of small pelagic animals (Cnidaria). The stem groups that gave rise to these extant groups had probably been using these feeding mechanisms for millions of years and would continue to use them. The early bilaterians were small worm-like (trace fossils) and flatworm-like (Vernanimalcula) animals that had recently evolved from ancestors that were radially symmetrical. The part of the ecosystem that they would feed in as grazers would be a benthos dominated by bacteria coated substrates and algae. It probably took these initial bilaterians a period of time to adapt to this new niche within their ecosystem. This may explain the delay in the proportional generic increase in bilaterians. Near the end of the Ediacaran period another modification took place in this ecosystem involving the introduction of predation by bilaterians on other metazoan animals, documented by the appearance of bore holes in Cloudina shells (Bengston and Zhao Citation1992).
Shen et al. (Citation2008) argue that the morphospace of Ediacarans did not change through time even though the species that make up the fauna do. This suggests that there were no major changes in their utilisation of the ecosystem. The picture that emerges in the data presented here demonstrates that there was a shift in the proportionate generation of new bilaterian genera during the late Ediacaran that may have been tied to their ability to utilise the benthic environment.
Postscript
Xiao and Laflamme (Citation2009) have recently produced a review that includes a section on the evolution of the Ediacaran fauna. This review focuses on the Avalon, White Sea and Nama assemblages. A possible phylogenetic placement of radial and bilaterally symmetrical fossils is given in their Figure . Avalon Rangeomorphs are the oldest Ediacaran clade shown and are indicated as the stock that the sponges and cnidarians evolved from. Sponges and cnidarians are not given crown group status until the Cambrian. Early bilaterians are shows evolving from stem group cnidarians during the late Ediacaran. This speculative phylogeny does not fit the facts. The fossils in the early Doushantuo Formation of the Yangtse Gorge predate the Avalon assemblage and include crown group sponges and cnidarians in addition to early bilaterians. It is probable that many of the bilaterian fossils found during the late Ediacaran (560–543 MA) evolved from these early bilaterians. If one is interested in the origin and early evolution of fossil bilaterians a good place to look for the appropriate fossils is the early Doushantuo Formation.
Acknowledgements
I want to thank Dennis Trombatore, the Geoscience librarian for his help, Marianna Grenadier for preparing the figures, and Judith Lundelius for reading the manuscript.
Related Research Data
References
- Babcock , LE , Grunow , AM , Sadowski , GR and Leslie , SA . 2005 . Corumbella an Ediacaran-grade organism from the Late Neoproterozoic of Brazil . Palaeogeo Palaeoclim Palaeoecol , 220 : 7 – 18 .
- Bamforth , EL , Narbonne , GM and Anderson , MM . 2008 . Growth and ecology of a multibranched Ediacaran rangeomorph from the Mistaken Point assemblage, Newfoundland . J Paleontol , 82 : 763 – 777 .
- Barford , GH , Albarede , F , Knoll , AH , Xiao , S , Telouk , P , Frei , R and Baker , J . 2002 . New Lu–Hf and Pb–Pb age constraints on the earliest animal fossils . Earth Planet Sci Lett , 201 : 203 – 212 .
- Beklemishev , WN . 1969 . “ Principles of comparative anatomy of invertebrates ” . In Promorphology , Vol. 1 , Chicago (IL) : University of Chicago Press .
- Bemus , AP . 1988 . Sedimentological context of a deep-water Ediacaran fauna (Mistaken Point, Avalon Zone of eastern Newfoundland) New York State . Mus Bull , 463 : 8 – 9 . abstract
- Bengston S, Budd G. 2004. Comment on ‘small bilaterian fossils from 40–55 million years before the Cambrian’. Science. 306:1291. Available from: http://www.sciencemag.org/cgi/content/full/306/5700/1291a
- Bengston , S and Rasmussen , B . 2009 . New and ancient trace makers . Science , 323 : 346 – 347 .
- Bengston , S and Zhao , Y . 1992 . Predatorial borings in late Precambrian mineralized exoskeletons . Science , 257 : 367 – 369 .
- Bowring , S , Myrow , P , Landing , E and Ramerzani , J . Geochronological constraints on terminal Neoproterozoic events and the rise of metazoans . NASA Astrobiological Institute (NIA General Meetings) . pp. 113 – 114 . abstract)
- Bowring , S , Myrow , P , Landing , E , Ramerzani , J and Grotzinger , J . 2003b . Geochronological constraints on terminal Neoproterozoic events and the rise of metazoans . Geophys Res Abst , 5 : 13219
- Boynton , HE and Ford , DT . 1979 . Pseudovendia charnwoodensis – a new Precambrian arthropod of Charnwood Forest, Leicestershire . Mercian Geol , 7 : 175 – 177 .
- Brasier , MD and Antcliffe , JB . 2009 . Evolutionary relationships within the Avolonian Ediacara biota: new insights from laser analysis . J Geol Soc Lond , 166 : 363 – 384 .
- Chen , DF , Dong , WQ , Zhu , BQ and Chen , XP . 2004 . Pb–Pb ages of Neoproterozoic Doushantuo phosphorites in South China: constraints on early metazoan evolution and glaciation events . Precamb Res , 132 : 123 – 132 .
- Chen , J-Y , Bottjer , DJ , Oliveri , P , Dornbos , SQ , Gao , F , Ruffins , S , Chi , H , Li , C-W and Davidson , EH . 2004a . Small bilaterian fossils from 40–55 million years before the Cambrian . Science , 305 : 218 – 222 .
- Chen , J-Y , Bottjer , DJ , Davidson , EH , Dornbos , SQ , Gao , X , Yang , Y-H , Li , C-W , Li , G , Wang , X-Q Xian , D-C . 2006 . Phosphatized polar lobe-forming embryos from the Precambrian of Southwestern China . Science , 312 : 1644 – 1646 .
- Chen J-Y, Oliveri P, Davidson E, Bottjer D. 2004b. Response to comment on ‘small bilaterian fossils from 40–55 million years before the Cambrian’. Science. 306, 1291. Available from: http://www.sciencemag.org.cgi/content/full/306/5700/1291b
- Chen , J-Y , Oliveri , P , Li , C-W , Zhou , G-Q , Gao , G-Q , Hagadorn , JW , Peterson , KJ and Davidson , EH . 2000 . Precambrian animal diversity: Putative phosphatized embryos from the Doushantuo Formation of China . Proc Nat Acad Sci USA , 97 : 4457 – 4462 .
- Chen , J-Y , Oliveri , P , Gao , F , Dornbos , SQ , Li , C-W , Bottjer , DJ and Davidson , EH . 2002 . Precambrian animal life: probable developmental and adult cindarian forms from Southwest China . Develop Biol , 248 : 182 – 196 .
- Chen , Z , Bengtson , S , Zhou , CM , Hua , H and Yue , Z . 2008 . Tube structure and original composition of Sinotubulites shelly fossils from the late Neoproterozoic in southern Shaanxi, China . Lethaia , 41 : 37 – 45 .
- Clapham , ME , Narbonne , GM , Gehling , JG , Greentree , C and Anderson , MM . 2004 . Thectardis avalonensis: a new Ediacaran fossil from the Mistaken Point Biota, Newfoundland . J Paleontol , 78 : 1031 – 1036 .
- Compston , W , Wright , AE and Toghill , P . 2002 . Dating the Late Precambrian vocanicity of England and Wales . J Geol Soc Lond , 159 : 323 – 339 .
- Condon , D , Zhu , M , Bowring , S , Wang , W , Yang , A and Jin , Y . 2005 . U–Pb ages from the Neoproterozoic Doushantuo Formation, China . Science , 308 : 95 – 98 .
- Dornbos , SQ , Bottjer , DJ , Chen , J-Y , Gao , F , Oliveri , P and Li , C-W . 2006 . Environmental controls on the taphonomy of phosphatized animals and animal embryos from the Neoproterozoic Doushantuo formation, Southwest, China . Palaios , 21 : 3 – 14 .
- Droser , ML and Gehling , JG . 2008 . Synchronous aggregate growth in an abundant new Ediacaran tubular organism . Science , 319 : 1660 – 1662 .
- Dunn , CW , Hejonl , A , Matus , DQ , Pang , K , Browne , WA , Smith , SA , Seaver , E , Rouse , GW , Obst , M Edgecombe , GD . 2008 . Broad phylogenetic sampling improves resolution of the animal tree of life . Nature , 452 : 745 – 749 .
- Dzik , J and Ivantsov , AY . 1999 . An asymmetric segmented organism from the Vendian of Russia and the status of the Dipleurozoa . Hist Biol , 13 : 255 – 268 .
- Dzik , J and Ivantsov , AY . 2002 . Internal anatomy of new Precambrian Dickinsoniid Dipleurozoan from northern Russia . N Jb Paläont Mh , 2002 : 385 – 398 .
- Fedonkin , MA . 1990a . “ Non-skeletal fauna of the Vendian: promorphological analysis ” . In The Vendian system. Vol. 1. Paleontology , 7 – 70 . Berlin : Springer-Verlag .
- Fedonkin , MA . 1990b . “ Systematic description of Vendian metazoa ” . In The Vendian system. Vol. 1. Paleontology , 71 – 120 . Berlin : Springer-Verlag .
- Fedonkin , MA . 2002 . Andiva ivantsovi gen. et. sp. n. and related carapace-bearing Ediacaran fossils from the Vendian of the Winter Coast, White Sea Russia . Ital J Zool , 69 : 175 – 181 .
- Fedonkin , MA , Gehling , JG , Grey , K , Narbonne , GM and Vickers-Rich , P . 2007a . The rise of animals , Baltimore (MD) : Johns Hopkins University Press .
- Fedonkin , MA and Ivantsov , AY . 2007b . Ventrogyrus, a possible siphonophore-like trilobozoan coelenterate from the Vendian sequence (late Neoproterozoic), northern Russia. The Rise and Fall of the Ediacaran Biota . Geol Soc Lond Spec Publ , 286 : 187 – 194 .
- Fedonkin , MA , Simonetta , A and Ivantsov , AY . 2007c . New data on Kimberella, the Vendian mollusk-like organism (White Sea region, Russia): Palaeoecological and evolutionary implications. The Rise and Fall of the Ediacaran Biota . Geol Soc Lond Spec Publ , 286 : 157 – 179 .
- Freeman , G and Lundelius , J . 1992 . Evolutionary implications of the mode of D quadrant specification in coelomates with spiral cleavage . J Evol Biol , 5 : 205 – 247 .
- Gaucher , C and Sprechmann , P . 1999 . Upper Vendian skeletal fauna of the Arroyo del Soldado Group, Uruguay . Beringeria , 23 : 55 – 91 .
- Gehling , JG . 1987 . Earliest known echinoderm – a new Ediacaran fossil from the Pound Subgroup of South Australia . Alcheringa , 11 : 337 – 345 .
- Gehling , JG . 1988 . A cnidarian of actinian-grade from the Ediacaran Pound Subgroup, South Australia . Alcheringa , 12 : 299 – 314 .
- Gehling , JG and Narbonne , GM . 2007 . Spindle-shaped Ediacaran fossils from the mistaken point assemblage, Avalon Zone, Newfoundland . Can J Earth Sci , 44 : 367 – 387 .
- Gehling , JG and Rigby , JK . 1996 . Long-expected sponges from the Neoproterozoic Ediacara Fauna, Pound Subgroup, South Australia . J Paleontol , 70 : 185 – 195 .
- Gehling , JG , Narbonne , GM and Anderson , MM . 2000 . The first named body fossil, Aspidella terranovica Billings 1872 . Palaeont , 43 : 427 – 456 .
- Gehling , JG , Droser , ML , Jensen , S and Runnegar , BN . 2005 . “ Ediacara organisms: relating form to function ” . In Evolving form and function: fossils and development , 43 – 66 . New Haven (CT) : Peabody Museum of Natural History, Yale University .
- Germs , GJB . 1973 . A reinterpretation of Rangea schneiderhoehni and the discovery of a related new fossil from the Nama group, South West Africa . Lethaia , 6 : 1 – 10 .
- Geyer , G and Uchman , A . 1995 . “ Ichnofossil assemblages from the Nama group (Neoproterozoic-Lower Cambrian) in Namibia and the Proterozoic-Cambrian boundary problem revisited. Morocco ’95 ” . In The Lower-Middle Cambrian standard of western Gondwana , Beringeria Special Issue Vol. 2 , 175 – 202 .
- Glaessner , MF . 1969 . Trace fossils from Precambrian and basal Cambrian . Lethaia , 2 : 368 – 393 .
- Glaessner , MF . 1977 . Re-examination of Archaeichinum, a fossil from the Nama Group . Ann S Afr Mus , 74 : 335 – 342 .
- Glaessner , MF . 1979 . An echiurid worm from the late Precambrian . Lethaia , 12 : 121 – 124 .
- Glaessner , MF . 1980 . Parvancornia – an arthropod from the Late Precambrian (Ediacaran) of South Australia . Ann Naturhist Mus Wien , 83 : 83 – 90 .
- Glaessner , MF and Wade , M . 1966 . The Late Precambrian fossils from Ediacara, South Australia . Palaeontology , 9 : 599 – 628 .
- Grazhdankin , DV . 2000 . The Ediacaran genus Inaria: a taphonomic morphodynamic analysis . N Jahrb Geol Palåont Abhand , 216 : 1 – 34 .
- Grazhdankin , D . 2004 . Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution . Paleobiology , 30 : 203 – 221 .
- Grotzinger , JP , Bowering , SA , Saylor , BZ and Kaufman , AJ . 1995 . Biostratigraphic and geochronologic constraints on early animal evolution . Science , 270 : 598 – 604 .
- Grotzinger , JP , Watters , WA and Knoll , AH . 2000 . Calcified metazoans in thrombolite-stromatolite reefs of the terminal Proterozoic Nama group, Namibia . Paleobiology , 26 : 334 – 359 .
- Hagadorn , JW and Waggoner , B . 2000 . Ediacaran fossils from the Southwestern Great Basin, United States . J Paleontol , 74 : 349 – 359 .
- Hofmann , HJ , O'Brien , SJ and King , AF . 2008 . Ediacaran biota on Bonavista Peninsula, Newfoundland, Canada . J Paleontol , 82 : 1 – 36 .
- Hua , H , Zhang , L-Y , Zhang , Z-F and Wang , J-P . 2000 . Fossil evidence of latest Neoproterozoic Gaojiashan biota and their characteristics . Acta Palaeontol Sinica , 39 : 511 – 515 .
- Hua , H , Chen , Z , Yuan , X , Zhang , L and Xiao , S . 2005 . Skeletogenesis and asexual reproduction in the earliest biomineralizing animal Cloudina . Geology , 33 : 277 – 280 .
- Ivantsov , AY . 1999 . A new Dickinsonid from the Upper Vendian of the White Sea Winter Coast (Russia Arkhangelsk Region) . Paleontol J , 33 : 211 – 221 .
- Ivantsov , AY . 2001 . Vendia and other Precambrian ‘Arthropods’ . Paleontol J , 35 : 335 – 343 .
- Ivantsov , AY . 2004 . New Proarticulata from the Vendian of the Arkhangel'sk region . Paleontol J , 38 : 247 – 253 .
- Ivantsov , AY . 2007 . Small Vendian transversely articulated fossils . Paleontol J , 41 : 113 – 122 .
- Ivantsov , AY and Fedonkin , MA . 2002 . Conulariid-like fossils from the Vendian of Russia: a metazoan clade across the Proterozoic/Palaeozoic boundary . Palaentol , 45 : 1219 – 1229 .
- Ivantsov , AY and Malakhovskaya , YE . 2002 . Giant traces of Vendian animals . Doklady Akad Nauk , 385 : 382 – 387 . (in Russian)
- Ivantsov , AY , Malakhovskaya , YE and Serezhnikova , EA . 2004 . Some problematic fossils from the Vendian of the southeastern White Sea region . Paleontol J , 38 : 1 – 9 .
- Jenkins , RLF . 1985 . The enigmatic Ediacaran (late Precambrian) genus Rangea and related forms . Paleobiology , 11 : 336 – 355 .
- Jenkins , RLF . 1992 . “ Functional and ecological aspects of Ediacaran assemblages ” . In Origin and early evolution of Metazoa , 131 – 176 . New York : Plenum Press .
- Jenkins , RJF . 1995 . The problems and potential of using animal fossils and trace fossils in terminal Proterozoic biostratigraphy . Precamb Res , 73 : 51 – 69 .
- Jenkins , RJF and Gehling , JG . 1978 . A review of frond like fossils of the Ediacaran assemblage . Rec S Aust Mus , 17 : 347 – 359 .
- Jenkins , RLF and Nedin , C . 2007 . The provenance and palaeobiology of a new multi-vaned chambered frondose organism from the Ediacaran (Later Neoproterozic) of South Australia. The rise and fall of the Ediacaran biota . Geol Soc Lond Spec Publ , 286 : 195 – 222 .
- Jensen , S . 2003 . The Proterozoic and Earliest Cambrian trace fossil record; patterns, problems and perspectives . Integr Comp Biol , 43 : 219 – 228 .
- Jensen , S , Saylor , BZ , Gehling , JG and Germs , GJB . 2000 . Complex trace fossils from the terminal Proterozoic of Namibia . Geology , 28 : 143 – 146 .
- Jensen , S , Droser , ML and Gehling , JG . 2005 . Trace fossil preservation and the early evolution of animals . Palaeogeo Palaeoclim Palaeoecol , 220 : 19 – 29 .
- Jensen , S , Droser , ML and Gehling , JG . 2006 . “ A critical look at the Ediacaran trace fossil record ” . In Neoproterozoic Geobiology and Paleobiology , 115 – 157 . New York : Springer-Verlag .
- LaFlamme , M , Narbonne , GM and Anderson , MM . 2004 . Morphometric analysis of the Ediacaran frond Charniodiscus from the Mistaken Point Formation, Newfoundland . J Paleontol , 78 : 827 – 837 .
- Leonov , MV . 2007 . Comparative taphonomy of Vendian genera Beltanelloides and Nemiana: taxonomy and lifestyle. The rise and fall of the Ediacaran Biota . Geol Soc Lond Spec Publ , 286 : 259 – 267 .
- Li , C-W , Chen , J-Y and Hua , T-E . 1998 . Precambrian sponges with cellular structures . Science , 279 : 879 – 882 .
- Lin , J-P , Gon , SM III , Gehling , JG , Babcock , LE , Zhao , Y-L , Zhang , X-L , Hu , S-X , Yuan , J-L , Yu , MY and Peng , J . 2006 . A Parvancornia-like arthropod from the Cambrian of South China . Hist Biol , 18 : 33 – 45 .
- Liu , P , Xiao , S , Yin , C , Zhou , C , Gao , L and Tang , F . 2008 . Systematic description and phylogenetic affinity of tubular microfossils from the Ediacaran Doushantuo formation at Weng'an, South China . Palaeontol , 2008 : 339 – 366 .
- Love , GD , Grosjean , E , Stalvies , C , Fike , DA , Grotzinger , JP , Bradley , AS , Kelly , AE , Bhatia , M , Meredith , W Snape , CE . 2009 . Fossil steroids record the appearance of Demospongiae during the Cryogenian period . Nature , 457 : 718 – 721 .
- Martin , MW , Grazhdankin , DV , Bowring , SA , Evans , DAD , Fedonkin , MA and Kirschvink , JL . 2000 . Age of Neoproterozoic bilaterian body and trace fossils, White Sea, Russia: implications for metazoan evolution . Science , 288 : 841 – 845 .
- Narbonne , GM . 1994 . New Ediacaran fossils from the Mackenzie Mountains northwestern Canada . J Paleontol , 68 : 411 – 416 .
- Narbonne , GM . 2004 . Modular construction of Early Ediacaran complex life forms . Science , 305 : 1141 – 1144 .
- Narbonne , GM . 2005 . The Ediacaran biota: Neoproterozoic origin of animals and their ecosystems . Ann Rev Earth Planet Sci , 33 : 421 – 442 .
- Narbonne , GM and Aitken , JD . 1990 . Ediacaran fossils from the Sekwi Brook Area Mackenzie Mountains, Northwestern Canada . Palaeontology , 33 : 945 – 980 .
- Narbonne , GM , Saylor , BZ and Grotzinger , JP . 1997 . The youngest ediacaran fossils from southern Africa . J Paleontol , 71 : 953 – 967 .
- Narbonne , GM , Laflamme , M , Greentree , C and Trusler , P . 2009 . Reconstructing a lost world: Ediacaran rangeomorphs from Spaniard's Bay, Newfoundland . J Paleontol , 83 : 503 – 523 .
- Pflug , HD . 1972 . Zur Fauna der Nama-Schichten in Sudwest-Afrika III. Erinettomorpha, Bau und Systematik. Sonder-Abdruk . Palaeontographica , 139A : 134 – 170 . Beitrage zur Naturgeschichte Der Vorzeit
- Preiss , WV . 2000 . The Adelaide Geosyncline of South Australia and its significance in Neoproterozoic continental reconstruction . Precamb Res , 100 : 21 – 63 .
- Runnegar , B . 1982 . Oxygen requirements, biology and phylogenetic significance of the Late Precambrian worm Dickisonia, and the evolution of the burrowing habit . Alcheringa , 6 : 223 – 239 .
- Runnegar , BN . 1992 . “ Evolution of the earliest animals ” . In Major events in the history of life , 65 – 93 . Boston (MA) : Jones and Bartlet .
- Shen , B , Dong , L , Xiao , S and Kowalewski , M . 2008 . The Avolon explosion: evolution of Ediacara morphospace . Science , 319 : 81 – 84 .
- Slack , JM . 1991 . From egg to embryo , 2nd ed , Cambridge : Cambridge University Press .
- Sun , WG . 1986 . Late Precambrian pennatulids (sea pens) from the eastern Yangtze Gorge, China: Paracharnia gen. nov . Precamb Res , 31 : 361 – 375 .
- Tang , F , Yin , C , Bengtson , S , Liu , Y , Wang , Z , Liu , P and Gao , L . 2006 . A new discovery of macroscopic fossils from the Ediacaran Doushantuo Formation in the Yangtze Gorges area . Chin Sci Bull , 51 : 1487 – 1493 .
- Tangri , SK , Bhargava , ON and Pande , AC . 2003 . Late-Precambrian-Early Cambrian trace fossils from the Tethyan Himalaya, Bhutan and their bearing on the Precambrian–Cambrian boundary . J Geol Soc India , 62 : 708 – 716 .
- Vickers-Rich , P . 2007 . “ The Nama of Southern Africa ” . In The rise of animals , 69 – 87 . Baltimore (MD) : Johns Hopkins University Press .
- Wade , M . 1969 . Medusae from uppermost Precambrian or Cambrian sandstones, central Australia . Palaeontology , 12 : 351 – 365 .
- Wade , M . 1972 . Hydrozoa and Scyphozoa and other medusoids from the Precambrian Ediacaran fauna, South Australia . Palaeontology , 15 : 197 – 255 .
- Wood , RA , Grotzinger , JP and Dickson , JAD . 2002 . Proterozoic modular biomineralized metazoan from the Nama group . Science , 296 : 2383 – 2386 .
- Xiao , S and Laflamme , M . 2009 . On the eve of animal radiation: phylogeny, ecology and evolution of the Ediacaran biota . Trends Ecol Evol , 24 : 31 – 40 .
- Xiao , S , Yuan , X , Steiner , M and Knoll , AH . 2002 . Macroscopic carbonaceous compressions in a terminal Proterozoic shale: a systematic reassessment of the Miaohe biota, South China . J Paleontol , 76 : 347 – 374 .
- Zhang , S , Jiang , G , Zhang , J , Song , B , Kennedy , MJ and Christie-Blick , N . 2005 . U–Pb sensitive high-resolution ion microprobe ages from the Doushantuo Formation of south China: constraints on late Neoproterozoic glaciations . Geology , 33 : 473 – 476 .