
ABSTRACT
Most faba bean research is about protein content while less attention is paid to nutrient content. Thus, the purpose of this study was to assess the effect of sowing rate and variety on the nutrient content of faba beans. Five varieties were tested: Bauska, Gloria, Julia, Jõgeva, and Lielplatones. Two sowing rates were used: 30 seeds m−2 (100%) and 36 seeds m−2 (120%). The higher sowing rate decreased the K content in varieties Gloria, Julia, and Lielplatones in 2016. In 2015 the higher sowing rate increased the Ca content while in 2016 the sowing rate decreased the Ca content. The higher sowing rate increased the Mg content in varieties Gloria and Jõgeva in 2016. The highest N, K, and Mg contents were found in variety Gloria. In 2015 the highest K content was found in varieties Gloria and Jõgeva, while in 2016 the highest K content was in variety Lielplatones. The highest Ca content was in variety Bauska. In conclusion, an increased sowing rate did not have a consistent effect on the nutrient content of faba beans. Thus, variety Gloria can be recommended for faba bean production in Northern Europe because it was the variety with the highest nutrient content.
Introduction
Faba bean (Vicia faba L.) is one of the most important pulse crops grown around the world. It is cultivated as a cool season-crop, for food, feed and green manure purposes (Olle Citation2018). Vicia faba is a traditional legume with high genetic diversity cultivated in a wide range of European climates. Cultivated faba bean is mainly used as human food in developing countries and as animal feed for pigs, horses, poultry and pigeons in industrialized countries (Feverole Citation2004). Seeds are small in Vicia faba var. minor (field beans), large in Vicia faba var. major (broad beans), while Vicia faba var. equina genotypes have an intermediate seed size (Crépon et al. Citation2010; Pietrzak et al. Citation2016). Therefore, V. faba has promise to identify stable and high yielding genotypes for specific target environments (Karkanis et al. Citation2018). The Nordic-Baltic region is challenging for legume production due to a short vegetation period and heat/drought stress in continental climates (Bodner et al. Citation2018).
Faba bean production acreage has declined from 4.8 million ha in 1961–2.4 million ha in 2008 (FAOSTAT Citation2018) despite a clear increase in the productivity per unit because the susceptibility to abiotic (Link et al. Citation2010; Singh et al. Citation2012) and biotic (Sillero et al. Citation2010; Singh et al. Citation2012) stresses has decreased. The global dry bean production area was 34 million ha, the total yield was 30 million tonnes and the average yield was 0.9 t ha−1 in 2018 (FAOSTAT Citation2018). Some countries have seen yield increase during the past 50 years more than others, such as France (64 kg year−1) and Egypt (35 kg year−1) compared with China (22 kg year−1). In the UK, Australia or Canada, average yield has been more stable during the past three decades while still highly variable (Jensen et al. Citation2010).
Yield potential for current faba bean cultivars in Europe is not far from 8.0 to 9.0 t ha−1. It is not a limiting factor anymore like it used to be in the 1970s. Breeders are working to improve yield stability and seed quality (Metayer Citation2004).
Genotype by environment interactions is more common in faba beans than in most other crops because faba beans are sensitive to the environment, and interactions of genotype by seasons generally make a greater contribution than genotype by location interactions (Bond Citation1987). Improved varieties have been appearing on national lists with increasing yield rates as a result of yields that are higher than a group of control varieties (Bond Citation1987).
The sowing rate of pulse crops is affected by environmental factors such as rainfall, irrigation or soil type; the more favourable environment, the higher the optimum seed density. In addition, seeding rates for faba bean vary according to seed size, sowing time and region (Olle Citation2018).
Proteins are major components of legume seeds. Their nutritional and functional properties dramatically affect the overall quality of the seed (Duranti and Gius Citation1997). Faba beans are an important source of protein in the diet of many people (Bond Citation1987). The protein content varied between 31.8% and 38.0% of seed dry matter. The average protein content was 26.1% of seed dry matter (Duc et al. Citation2011). Faba bean plants, due to their high nitrogen-fixing ability, can play a significant role as N sources in cropping systems. Thus the inclusion of faba bean in crop rotations could reduce the need for the use of inorganic N fertilizers (Ntatsi et al. Citation2018).
Most faba bean research is about protein content while less attention is paid to nutrient content (N, P, K, Ca and Mg). Thus the purpose of this study was to assess the effect of sowing rate and variety on the nutrient content of faba beans.
Materials and methods
Field experiments with faba bean (Vicia faba L.) crop were conducted at the experimental station of the Estonian Crop Research Institute, located in Jogeva alevik, in middle of Estonia (58°46′10.6″N 26°24′24.9″E) in 2015 and 2016.
Treatments used in trials with faba bean were followed:
Two sowing rates were used: 30 seeds m−2 (100%) and 36 seeds m−2 (120%). The varieties were: Bauska (Latvia), Gloria (Austria), Julia (Austria), Jõgeva (Estonia) and Lielplatones (Latvia).
A completely randomized experimental design was used with four replications. Plot size was 10 m2. Soil conditions in 2015 were as follows: soil organic matter content was 3.46% and pH was 6.29. The soil type was calcaric cambisol soil in the Estonian system. The preceding crop was barley. Soil conditions in 2016 were as follows: soil organic matter content was 3.39% and pH was 6.22. The soil type was soddy-calcarous podzolic soil in the Estonian system. The preceding crop was oat.
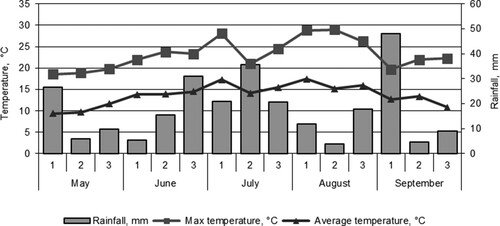
In 2015, the weather was very different from the historical average with a cold spring and cold summer (). There were some periods with too much rain and some with too little rain.
Figure 1. Weather conditions over field pea vegetation period in 2015, data from Jõgeva Meteorological Station.
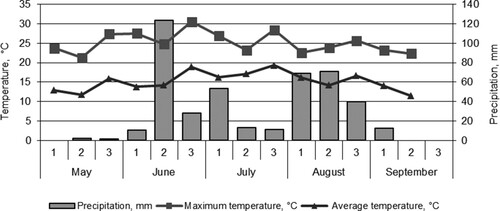
Weather in Estonia, in May 2016 (), was very warm, which in combination with little rain led to very low soil moisture compared to a normal year. June and July were characterized by colder temperatures than average. A lot of rainfall came at the middle of June and beginning of July which provided good conditions for vegetative growth and flowering. Overall, the precipitation was much higher than the average. The rainy season, up to harvesting had a positive impact on the seed yield of faba beans but which caused lodging because the average weight of seeds per plant was high.
Figure 2. Weather conditions over field pea vegetation period in 2016, data from Jõgeva Meteorological Station.
The experimental fields were ploughed 1 year before the trials were established at the end of the 2014 and 2015 growing seasons, respectively. Cultivation was done twice before sowing in the spring of 2015 and 2016. Seed was sown on 4–5 May 2015 and on 5–6 May 2016 at a depth of 4–5 cm. Row distribution was 5 rows between tractor wheel tracks, spaced 20 cm apart. The plot size was 10 m2.
The trials were not irrigated. Fertilization was done with Yara Mila 7-12-25 (300 kg ha−1). In 2015, weeds were controlled by Activus 330 (pendimethalin 330 g L−1) EC at 1.5 L ha−1 + Basagran 480 (bentazon 480 g L−1) 1.5 L ha−1, on 5 June 2015. In 2016, the same herbicides were used, but Activus 330 was applied at the higher rate of 3.0 L ha−1, on 19 May 2016. No control measures against insects and diseases were applied.
The faba beans were harvested between 31 August and 16 September in 2015 and between 27 August and 11 September in 2016. The seeds were dried by forced drying at 30–40°C until the faba bean seeds reached 14% moisture content.
The N, P, K, Ca and Mg contents were determined in faba bean seeds dry matter. Nitrogen content was determined according to the Copper Catalyst Kjeldahl Method (984.13). Phosphorus content was found with a Kjeldahl Digest by Fiastar 5000 (AN 5242; Stannous Chloride method, ISO/FDIS 15681). Potassium content was by the Flame Photometric Method (956.01). Calcium content was by the o-Cresolphthalein Complexone method (ISO 3696, in Kjeldahl Digest by Fiastar 5000). Magnesium content was by Fiastar 5000 (ASTN90/92; Titan Yellow method).
Analyses of variance were used to test the significance of variety and sowing rate on disease incidence. In case of significant effects, differences between treatments were tested post-hoc using Fisher’s LSD test. The statistical analyses were performed using Agrobase (Agrobase Generation II 38.32.3) software. Signs used: ***p < 0.001; **p = 0.001–0.01; *p = 0.01–0.05; NS not significant, p > 0.05.
Results
Nitrogen content
Different genotypes influenced the N content of faba beans both years ( and ). The highest N content was in variety Gloria and lowest N content was in varieties Bauska and Jõgeva. The sowing rate did not influence the N content either year ( and ).
Table 1. Nitrogen, phosphorus, potassium, calcium and magnesium contents in faba bean dry matter in 2015 at ECRI by variety and sowing rate*.
Table 2. Nitrogen, phosphorus, potassium, calcium and magnesium contents in faba bean dry matter in 2016 at ECRI by variety and sowing rate*.
Phosphorus content
Different genotypes influenced the P content of faba beans both years ( and ). In 2015, the highest P content was in varieties Gloria and Jõgeva, while in 2016 the highest P content was in variety Lielplatones. In 2015, the lowest P content was in varieties Bauska and Lielplatones, while in 2016 the lowest P content was in Julia. The sowing rate did not influence the P content either year ( and ).
Potassium content
Different genotypes influenced the K content of faba beans both years ( and ). In 2015, the highest K content was in varieties Gloria and Jõgeva, while in 2016 the highest K content was in variety Gloria. In 2015, the lowest K content was in variety Lielplatones, while in 2016 the lowest K content was in Julia. The sowing rate did not influence the K content in 2015 (). The sowing rate did influence the K content in 2016 (). The higher sowing rate decreased the K content in varieties Gloria, Julia and Lielplatones.
Calcium content
Different genotypes influenced the Ca content of faba beans both years ( and ). The highest Ca content was in variety Bauska. In 2015, the lowest Ca content was in variety Lielplatones, while in 2016 the lowest Ca content was in Gloria. The sowing rate influenced the Ca content both years ( and ). In 2015, the higher sowing rate increased the Ca content of faba beans, while in 2016 it decreased the Ca content.
Magnesium content
Different genotypes influenced the Mg content of faba beans both years ( and ). The highest Mg content was in variety Gloria. In 2015, the lowest Mg content was in variety Bauska, while in 2016 the lowest Mg content was in Jõgeva. The sowing rate did not influence the Mg content in 2015 (). The sowing rate influenced the Mg content in 2016 (). The higher sowing rate increased the Mg content in varieties Gloria and Jõgeva.
The increase in sowing rate by 20% influenced faba bean K, Ca and Mg contents. The higher sowing rate decreased the K content in varieties Gloria, Julia and Lielplatones in 2016. In 2015, the higher sowing rate increased the Ca content of Calcium, while in 2016 it decreased the Ca content. The higher sowing rate increased the Mg content in varieties Gloria and Jõgeva in 2016. The different varieties influenced faba bean nutrient content. The highest N content was in variety Gloria. In 2015, the highest P content was in varieties Gloria and Jõgeva, while in 2016 the highest P content was in variety Lielplatones. In 2015, the highest K content was in varieties Gloria and Jõgeva, while in 2016 the highest K content was in variety Gloria. The highest Ca content was in variety Bauska. The highest Mg content was in variety Gloria. The higher sowing rate did not have a consistent effect on the nutrient content of faba beans. Thus variety Gloria can be recommended for faba bean production in Northern Europe because it was the variety with the highest nutrient content.
Discussion
The mineral content of pulses is generally high, but the bioavailability is poor due to the presence of phytate which is a main inhibitor of Fe and Zn absorption. Some legumes also contain considerable amounts of Fe-binding polyphenols thus inhibiting Fe absorption (Sandberg Citation2002). Legume seeds are an important source of dietary minerals, with the potential to provide all 15 of the essential minerals required by humans. The concentrations of certain minerals (especially Ca), however, are low relative to animal food products (Wang et al. Citation2003).
Increased nutrient uptake could influence plants negatively because a high nitrate accumulation in plants, which was present in this study, might be undesirable. High nitrate accumulation in plants results in nitrite production which is converted to nitric oxide by nitrate reductase and then is converted into the extremely toxic compound peroxynitrite under aerobic conditions; peroxynitrite is harmful to plant growth (Reddy and Menary Citation1990). Even more serious, the accumulation of peroxynitrite in humans may result in conditions such as chronic heart failure, diabetes, chronic inflammatory diseases, cancer and neurodegenerative disorders (Pacher et al. Citation2007).
This study showed that P content varied by faba bean variety. Similarly, Hossain and Mortuza (Citation2006) and Singh et al. (Citation2014) wrote that P content varies between cultivars, while a high P is needed especially for good root growth (Durner Citation2013).
This study showed that K content varied by faba bean variety. This agrees with Hossain and Mortuza (Citation2006) and Singh et al. (Citation2014) who found that K content varies between cultivars, while a K high content is very important in stomatal function and water relations of plants (Durner Citation2013).
Calcium content in this study varied by faba bean variety. In accordance scientists, Hossain and Mortuza (Citation2006) described that calcium content is different between different cultivars. A higher Ca content is beneficial, suppressing insect and disease attack and increasing transportability and storage quality (Olle Citation2013; Olle Citation2015).
Magnesium content in this study varied by faba bean variety. Hossain and Mortuza (Citation2006) also showed that Mg content varies between cultivars. A higher Mg content could be desirable because a higher Mg content reduces the incidence of insect pests and diseases (Cakmak Citation2013).
Vegetative tissues play a central role in the collection, temporary storage and subsequent redistribution of minerals. In fact, the leaf concentration of several minerals declines as seeds develop. What is still not known is (a) whether the remobilized minerals are phloem-loaded apoplastically or symplastically, (b) if phloem mineral loading can occur throughout all minor and major veins, (c) what role the mesophyll tissues play in the storage and possible release of minerals and (d) which transporters might be required at the sieve element-companion cell periphery (Wang et al. Citation2003).
Because of the many unknowns pertaining of spatial mineral distribution and leaf anatomy, it is worth noting that a multitude of leaf morphology mutants is available in field pea which could serve as unique experimental tools to investigate the contribution of various tissue types and vascular patterns on seed mineral accretion (Wang et al. Citation2003).
Acknowledgements
This investigation has been developed with the help of the project EUROLEGUME, funded by the European Union Seventh Framework Programme for Research, Technological Development and Demonstration under the grant agreement No. 613781.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
Margit Olle
Margit Olle (before marriage on 19.07.2003 Margit Kleemann) was born on 27.05.1971. She finished Secondary School No. 5 in Tartu with a Silver medal. She studied for her B.Sc. at the Estonian Agricultural University and finished in 1993 Cum Laude, passing all exams excellently. In 1995, she defended her M.Sc. at the same Institution. She took her doctoral studies in 1995–1999 in Norway at the Agricultural University of Norway and defended her Doctor Scientiarum degree on 27.05.1999. In 2008–2020, she has worked as senior researcher at the Estonian Crop Research Institute (former Jogeva Plant Breeding Institute). Since 2021, she works at NPO Veggies Cultivation as chairman of the board and senior researcher. She has developed three discoveries in greenhouse vegetable growing. She has written five research monographs in English, one popular – scientific book in English and one book of vegetable growing in Estonian. She has organized conferences, has been invited speaker at international conferences and is member of editorial board of scientific journals. She also has some international awards.
Sirje Tamm
Sirje Tamm currently works at Estonian Crop Research institute. She graduated Estonian Agricultural Academy (now Estonian University of Life Sciences) Magister of Science in field of Agronomy. Her research areas are breeding of forage grasses (mainly forage legume grasses) and seed production investigations of forage grasses. Sirje Tamm deals with questions of grassland husbandry and collecting and describing genetic resources of forage grasses. She has published with co-authors research articles in her field and in organic farming and in theme about cover crops.
References
- Bodner G, Kronberga A, Lepse L, Olle M, Vågen IM, Rabante L, Fernández JA, Ntatsi G, Balliu A, Rewald B. 2018. Trait identification of faba bean ideotypes for Northern European environments. Eur J Agron. 96:1–12.
- Bond DA. 1987. Recent developments in breeding field beans (Vicia faba L.). Plant Breed. 99:1–26.
- Cakmak I. 2013. Magnesium in crop production, food quality and human health. Plant Soil. 368:1–4.
- Crépon K, Marget P, Peyronnet C, Marget P, Peyronnet C, Carrouée B, Arese P, Duc G. 2010. Nutritional value of faba bean ( Vicia faba L.) seeds for feed and food. Field Crop Res. 115:329–339.
- Duc G, Marget P, Arese P. 2011. Breeding priorities for improved nutritional value of Vicia faba seeds. Grain Legumes. 56:17–18.
- Duranti M, Gius C. 1997. Legume seeds: protein content and nutritional value. Field Crops Res. 53:31–45.
- Durner EF. 2013. Principles of horticultural physiology. Tarxien, Malta: Gutenberg Press Ltd. 405.
- FAOSTAT. 2018. [Accessed 2020 April 6]. http://www.fao.org/faostat/.
- Feverole G. 2004. Vicia Faba breeding for sustainable agriculture in EUROPE. D7: WP 1- EUFABA, pp. 29.
- Hossain MS, Mortuza MG. 2006. Chemical composition of kalimatar, a locally grown strain of faba bean (Vicia faba L.). Pak J Biol Sci 9:1817–1822.
- Jensen ES, Peoples MB, Hauggaard-Nielsen H. 2010. Faba bean in cropping systems. Field Crops Res. 115:203–216.
- Karkanis A, Ntatsi G, Lepse L, Fernández JA, Vågen IM, Rewald B, Alsiņa I, Kronberga A, Balliu A, Olle M, Bodner G. 2018. Faba bean cultivation–revealing novel managing practices for more sustainable and competitive European cropping systems. Front Plant Sci. 9:1–14.
- Link W, Balko C, Stoddard FL. 2010. Winter hardiness in faba bean: physiology and breeding. Field Crops Res. 115:287–296.
- Metayer N. 2004. Vicia faba breeding for sustainable agriculture in Europe. Giefeverole. 20(2). C:/Users/TOSHIBA/Downloads/EUFABA_Metayer_04_V-faba-for-sust-agric-in-Europe-GIE-f%C3%A9verole.
- Ntatsi G, Karkanis A, Yfantopoulos D, Olle M, Travlos I, Thanopoulos R, Bilalis D, Bebeli P, Savvas D. 2018. Impact of variety and farming practices on growth, yield, weed flora and symbiotic nitrogen fixation in faba bean cultivated for fresh seed production. Acta Agric Scand B Soil Plant Sci. 68:619–630.
- Olle M. 2013. The effect of effective microorganisms (Em) on the yield, storability and calcium content in Swede. XVII. In: Coskun D, Britto DT, Li M, Kronzucker HJ, editors. International plant nutrition colloquium and boron satellite meeting proceedings book, 2013. Istanbul: Sabanci University; p. 714–715.
- Olle M. 2015. Methods to avoid Calcium deficiency on greenhouse grown leafy crops. Frankfurt: LAP LAMBERT Academic Publishing. 118.
- Olle M. 2018. Suitable sowing rate for peas and beans - A review. JOJ Hortic & Arboric. 1(3):1–3.
- Pacher P, Beckman JS, Liaudet L. 2007. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 87:315–424.
- Pietrzak W, Kawa-Rygielska J, Król B, Lennartsson PR, Taherzadeh MJ. 2016. Ethanol, feed components and fungal biomass production from field bean (Vicia faba var. equina) seeds in an integrated process. Bioresour Technol. 216:69–76.
- Reddy KS, Menary RC. 1990. Nitrate reductase and nitrate accumulation in relation to nitrate toxicity in boronia megastigma. Physiol Plant. 78:430–434.
- Sandberg AS. 2002. Bioavailability of minerals in legumes. Br J Nutr. 88:281–285.
- Sillero JC, Villegas-Fernandez AM, Thomas J, Rojas-Molina MM, Emeran AA, Fernandez-Aparicio M, Rubiales D. 2010. Faba bean breeding for disease resistance. Field Crop Res. 115:297–307.
- Singh AK, Bhardwaj R, Singh IS. 2014. Assessment of nutritional quality of developed faba bean (Vicia faba L. lines). J AgriSearch. 1:96–101.
- Singh AK, Bhatt BP, Upadhyaya A, Kumar S, Sundaram PK, Singh BK, Chandra N, Bharati RC. 2012. Improvement of faba bean (Vicia faba L.) yield and quality through biotechnological approach: A review. Afr J Biotechnol. 11:15264–15271.
- Wang TL, Domoney C, Hedley CL, Casey R, Grusak MA. 2003. Can we improve the nutritional quality of legume seeds? Plant Physiol. 131:886–891.