Keywords:
Introduction
Volumetric measurement of water uptake can be made by using potometry (Scholander et al., Citation1955, Citation1957; Kramer, Citation1959). A potometric method has been modified to study water flow through the fruit pedicel into detached sweet cherry fruit (Hovland & Sekse, Citation2004).
The cherry fruit has a double sigmoid growth pattern (Coombe, Citation1976) and during the lag phase (stage II) the embryo develops with little change in total fruit mass (31–37 days after full bloom (DAFB)). Stage III (final swell) of development, characterized primarily by mesocarp development, starts about 43 DAFB and accounts for the rapid increase in fruit mass (Lilleland & Newsom, Citation1934). The cuticular membrane (CM) of the fruit, which is a primary barrier for water transport into and out of the fruit over its surface (Franke, Citation1967), develops simultaneously and its conductance varies with fruit development (Knoche et al., Citation2001). Water uptake through the vascular tissues in developing fruit is of importance in stage III in the growth curve, particularly for understanding the underlying mechanisms of cuticular fracture development and of fruit cracking (Sekse, Citation1995; Yamamoto et al., Citation1990).
This paper presents results from potometric studies of water uptake through the pedicel of two cultivars of sweet cherry in relation to fruit development.
Materials and methods
The potometric apparatus
The potometric apparatus was constructed according to descriptions given by Hovland & Sekse (Citation2004). It consisted of a waterfilled glass tube mounted parallel to a metric scale. Sweet cherry fruit with a pedicel end (2 cm) were mounted in the waterfilled end of a capillary glass tube through a rubber tube. The glass tube was mounted parallel to a metric scale on a rectangular piece of cardboard. The apparatus was placed horizontally on a grate and readings of the water meniscus position were made every 15 min. The relative humidity (RH) of the surrounding air in the laboratory during the experimental period was 50±5% (Hanna Instruments, HI 9065 Thermo Hygrometer™).
Plant material
Fruits grown in the experimental field at Ullensvang Research Centre, Lofthus, Norway on trees receiving standard treatments regarding drip-irrigation, fertilization and plant protection spray applications were used. Standard conditions, such as morning picked fruit similar in appearance regarding size and ripeness, and approximately standardized environmental conditions were applied.
Fruit samples of ‘Van’ were harvested at ten different dates throughout the 1995 season at 3- to 4-day intervals starting 13 July and ending 14 August (). Similarly, fruit samples of ‘Van’ and ‘Sunburst’ were harvested at ten different dates throughout the 1997 season at 3- to 7-day intervals starting 30 June and ending 11 August ( and ).
Table 1. Colour and weight development of ‘Van’ sweet cherries (1995) at different fruit developmental stages (DAFB=days after full bloom, FB: 22 May). Mean values of 10 fruits
Table 2. Colour and weight development and water uptake of ‘Van’ sweet cherries (1997) at different fruit developmental stages (DAFB=days after full bloom, FB: 22 May) Mean values of 10 fruits
Table 3. Colour and weight development and water uptake of ‘Sunburst’ sweet cherries (1997) at different fruit developmental stages (DAFB=days after full bloom, FB: 22 May). Mean values of 10 fruits
Fruits were immediately after harvest positioned in the potometer and water uptake was measured through their pedicels for 4 h in 1995 and 6 h in 1997. Ten representative fruits were weighted gravimetrically and examined for colour using a scale ranging from 1–7 where 1=light red and 7=mahogany/black (Afco fel, Ctifl, France) (Planton, Citation1995).
Results and discussion
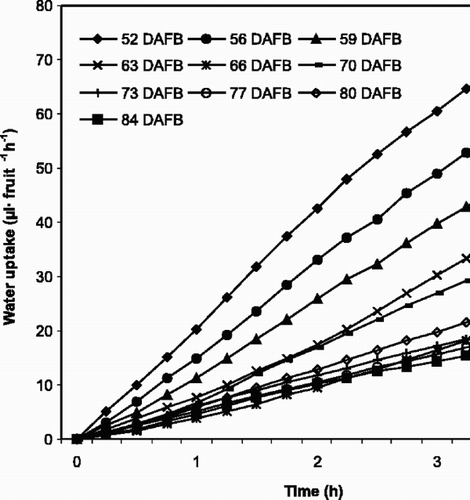
Water uptake was significantly highest (P≤0.0001) during the first dates of harvest (early stage III) 52–56, 40–54 and 39–49 DAFB for Van in 1995 (), Van in 1997 () and Sunburst in 1997 (), respectively. The numbers for water uptake were compared by Tukey's pairwise comparison and the highest water loss is marked by the letter c in and . In the numbers for water loss were statistically higher at 52 and 56 DAFB compared to the other dates. The measured water uptake then decreased significantly in the periods 59–70 (), 43–68 () and 42–67 DAFB (), respectively, and continued to decrease with increasing stage of fruit maturity. If water uptake through the pedicel is calculated on the basis of total fruit surface area, fruit of ‘Van’ (1997) lost 5.17 10−3 g m−2 s−1 and 0.88 10−3 g m−2 s−1 at 40 and 82 DAFB, respectively, which represents a pronounced reduction of water uptake during fruit development.
Fig. 1. Water uptake through sweet cherry fruit pedicels over a 4 h period for ‘Van’ (1995) at different fruit developmental stages (DAFB=days after full bloom, FB: 22 May). Each point represents 6 fruits.
Knoche et al. (Citation2001) concluded that the total conductance for the CM was related to fruit development. During stage II (31–37 DAFB) the total conductance was constant, in early stage III (43–64 DAFB) it increased, while it markedly decreased in later developmental stages. After 64 DAFB the conductance decreased, while the water uptake continued to decrease. This can explain the variation in the potometrically measured water transport through the fruit pedicel; decreased conductance in the CM towards the final stage of fruit development allows more of the water to be utilized for fruit size expansion. This also explains why the fruit continued to increase their size at the end of the season although the total amount of absorbed water was decreasing. However, according to Knoche et al. (Citation2001), the CM of stage III fruit was thinner and more permeable to water than the thicker CM of stage II fruit.
When the fruit continued to ripen the intercellular spaces decreased; they could with difficulty be observed in stage III (Tukey & Young, Citation1939). The cells became thin-walled and closer to each other. This physiological change can explain why overripe fruit act differently from fruit at earlier stages. It is often observed that overripe fruit do not crack in rain (Christensen, Citation1973). The cell wall structures become nearly disintegrated, making the total fruit act more or less as an osmotic entity.
The fruit weight constantly increased until 77, 68 and 67 DAFB for Van in 1995 (), Van in 1997 () and Sunburst in 1997 (), respectively. Similarly, the maximum increase in weight occured around 59, 54 and 53 DAFB, respectively. Assuming that this weight increase caused increase in fruit surface area, the results are well in accordance with Knoche et al. (Citation2001) who found that the maximum rate of surface area expansion occurred at 53 DAFB.
In conclusion, in late stage III, the fruit expansion was high while both the rate of water uptake and the CM conductance was simultaneously decreasing. Decreased conductance allowed more of the absorbed water to be utilized for size expansion. In this period the fruit were more vulnerable to develop cuticular fractures (Hovland et al., Citation2004) and to crack (Christensen, Citation1973). In order to avoid this, it seems to be important to control the amount of available water to the roots. This can be achieved by covering the trees and/or by the use of drip irrigation to provide the tree with regular water supply.
Additional information
Notes on contributors
Kari Louise Hovland
Hovland, K. L. and Sekse, L. (The Norwegian Crop Research Institute, Apelsvoll Research Centre Division Kise, NO-2350 Nes, Norway and The Norwegian Crop Research Institute, Ullensvang Research Centre, NO-5781 Lofthus, Norway). Water uptake through sweet cherry (Prunus avium L.) fruit pedicels in relation to fruit development.Notes
Hovland, K. L. and Sekse, L. (The Norwegian Crop Research Institute, Apelsvoll Research Centre Division Kise, NO-2350 Nes, Norway and The Norwegian Crop Research Institute, Ullensvang Research Centre, NO-5781 Lofthus, Norway). Water uptake through sweet cherry (Prunus avium L.) fruit pedicels in relation to fruit development.
References
References
- Christensen , JV . 1973 . Cracking in cherries. VI. Cracking susceptibility in relation to the growth rhythm of the fruit . Acta Agric. Scand. , 23 : 52 – 54 .
- Coombe , BG . 1976 . The development of fleshy fruits . Ann. Rev. Plant Phys. , 27 : 507 – 528 .
- Franke , W . 1967 . Mechanisms of foliar penetration of solutions . Ann. Rev. Plant Phys. , 18 : 281 – 300 .
- Hovland , KL and Sekse , L . 2004 . Water uptake through sweet cherry (Prunus avium L.) fruit pedicels: influence of fruit surface water status and intact fruit skin . Acta Agric. Scand., Sect. B, Soil and Plant Sci. , 54 : 91 – 96 .
- Hovland KL Vangdal E Sekse L Vestrheim S 2004 Cuticular fracture development and water retention of sweet cherry (Prunus avium L.) fruit Acta Agr. Scand. Sect. B, Soil and Plant Sci. (re-submitted during July 2004).
- Knoche , M , Peschel , S , Hinz , M and Bukovac , MJ . 2001 . Studies of water transport through the sweet cherry fruit surface: II. Conductance of the cuticle in relation to fruit development . Planta , 213 : 927 – 936 .
- Kramer JP Transpiration and the water economy of plants In: Steward F. C. (ed.) Plant Physiology II Academic Press New York pp. 607–730 1959
- Lilleland , O and Newsom , L . 1934 . A growth study of the cherry fruit . Proc. Am. Soc. Hort. Sci. , 32 : 291 – 299 .
- Planton , G . 1995 . Cherries: a colour code to improve crop quality. Centre Technique Interprofessionel des Fruits et Legumes, France . Infos No. , 112 : 38 – 41 .
- Scholander , PF , Love , WE and Kanwisher , JW . 1955 . The rise of sap in tall grapevines . Plant Phys. , 30 : 93 – 104 .
- Scholander , PF , Ruud , B and Leivestad , H . 1957 . The rise of sap in a tropical liana . Plant Phys. , 32 : 1 – 6 .
- Sekse , L . 1995 . Cuticular fracturing in fruits of sweet cherry (Prunus avium L.) resulting from changing soil water contents . J. Hort. Sci. , 70 : 631 – 635 .
- Tukey , HB and Young , JO . 1939 . Histological study of the developing fruit of the sour cherry . Bot. Gaz. , 100 : 723 – 749 .
- Yamamoto , T , Hosoi , K and Watanabe , S . 1990 . Relationship between the degree of fruit cracking of sweet cherries and the distribution of surface stress of the fruit analysed by a newly developed system . J. Jap. Soc. Hort. Sci. , 59 : 509 – 517 .